Внутренне светочувствительные ганглиозные клетки сетчатки

Внутренне светочувствительные ганглиозные клетки сетчатки ( ipRGC ), также называемые светочувствительными ганглиозными клетками сетчатки ( pRGC ), или меланопсин-содержащие ганглиозные клетки сетчатки ( mRGC ), представляют собой тип нейронов в сетчатке глаза млекопитающих . О наличии дополнительного фоторецептора впервые заподозрили в 1927 году, когда мыши, у которых отсутствовали палочки и колбочки, все еще реагировали на изменение уровня освещенности сужением зрачков ; [1] это позволило предположить, что палочки и колбочки — не единственные светочувствительные ткани. [2] Однако было неясно, возникает ли эта светочувствительность из-за дополнительного фоторецептора сетчатки или из-за чего-то еще в организме. Недавние исследования показали, что эти ганглиозные клетки сетчатки , в отличие от других ганглиозных клеток сетчатки, по своей природе светочувствительны из-за присутствия меланопсина , светочувствительного белка. Следовательно, они составляют третий класс фоторецепторов, помимо палочек и колбочек . [3]
Обзор
[ редактировать ]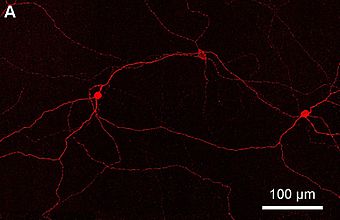
По сравнению с палочками и колбочками ipRGC реагируют более вяло и сигнализируют о присутствии света в течение длительного времени. [5] Они представляют собой очень небольшую подгруппу (~ 1%) ганглиозных клеток сетчатки. [6] Их функциональные роли не являются образообразующими и принципиально отличаются от ролей образного зрения; они обеспечивают стабильное представление интенсивности окружающего света. У них есть как минимум три основные функции:
- Они играют важную роль в синхронизации циркадных ритмов с 24-часовым циклом света и темноты, предоставляя в первую очередь информацию о продолжительности дня и ночи. Они посылают световую информацию через ретиногипоталамический тракт к циркадному — водителю ритма мозга супрахиазматическому ядру гипоталамуса (RHT) непосредственно . Физиологические свойства этих ганглиозных клеток соответствуют известным свойствам механизма ежедневного уноса света ( синхронизации ), регулирующего циркадные ритмы. Кроме того, ipRGC могут также влиять на периферические ткани, такие как регенерация волосяных фолликулов, через симпатическую нервную цепь SCN. [7]
- Фоточувствительные ганглиозные клетки иннервируют другие цели мозга, такие как центр зрачкового контроля , оливковое претектальное ядро среднего мозга . Они способствуют регулированию размера зрачков и другим поведенческим реакциям на условия окружающего освещения. [8]
- Они способствуют фоторегуляции и острому фотоподавлению высвобождения гормона мелатонина . [8]
- У крыс они играют определенную роль в сознательном зрительном восприятии, включая восприятие обычных решеток, уровней освещенности и пространственной информации. [8]
Фоторецепторные ганглиозные клетки были выделены у людей, где, как было показано, помимо регуляции циркадного ритма, они опосредуют определенную степень распознавания света у субъектов без палочек и колбочек, страдающих нарушениями фоторецепторов палочек и колбочек. [9] Работа Фархана Х. Заиди и его коллег показала, что фоторецепторные ганглиозные клетки могут выполнять у людей некоторую зрительную функцию.
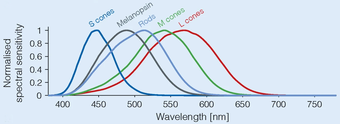
Фотопигмент фоторецепторных ганглиозных клеток, меланопсин, возбуждается светом преимущественно в синей части видимого спектра (пики поглощения при ~480 нанометрах). [10] ). Механизм фототрансдукции в этих клетках до конца не изучен, но, по-видимому, он напоминает таковой в рабдомерных фоторецепторах беспозвоночных. Помимо прямой реакции на свет, эти клетки могут получать возбуждающее и тормозящее влияние от палочек и колбочек посредством синаптических связей в сетчатке.
Аксоны этих ганглиев иннервируют области мозга, связанные с распознаванием объектов, включая верхний холмик и дорсально -латеральное коленчатое ядро . [8]
Структура
[ редактировать ]рецептор ipRGC
[ редактировать ]
Эти фоторецепторные клетки проецируются как по всей сетчатке, так и в мозг. Они содержат фотопигмент меланопсин в различных количествах вдоль клеточной мембраны, в том числе на аксонах до диска зрительного нерва, соме и дендритах клетки. [3] ipRGC содержат мембранные рецепторы для нейромедиаторов глутамата, глицина и ГАМК . [11] Фоточувствительные ганглиозные клетки реагируют на свет деполяризацией, тем самым увеличивая скорость подачи нервных импульсов, что противоположно скорости других фоторецепторных клеток, которые гиперполяризуются в ответ на свет. [12]
Результаты исследований на мышах позволяют предположить, что аксоны ipRGCs немиелинизированы . [3]
Меланопсин
[ редактировать ]В отличие от других пигментов фоторецепторов, меланопсин обладает способностью действовать как возбудимый фотопигмент и как фотоизомераза. В отличие от зрительных опсинов в палочках и колбочках , которые используют стандартные зрительные циклы для перезарядки полностью транс- ретиналя обратно в светочувствительный 11-цис -ретиналь , меланопсин способен изомеризовать полностью транс- ретиналь в 11-цис-ретиналь. сама сетчатка при стимуляции другим фотоном. [11] Таким образом, ipRGC не полагается на клетки Мюллера и/или клетки пигментного эпителия сетчатки для этого преобразования.
Две изоформы меланопсина различаются по своей спектральной чувствительности: изоформа 11-цис -ретиналя более чувствительна к более коротким длинам волн света, тогда как полностью транс- изоформа более чувствительна к более длинным длинам волн света. [13]
Синаптические входы и выходы
[ редактировать ]
Входы
[ редактировать ]ipRGC являются как пре-, так и постсинаптическими по отношению к дофаминергическим амакриновым клеткам (DA-клеткам) через реципрокные синапсы, при этом ipRGC посылают возбуждающие сигналы клеткам DA, а клетки DA посылают тормозящие сигналы ipRGC. Эти ингибирующие сигналы передаются через ГАМК , которая высвобождается из клеток DA вместе с дофамином . Дофамин участвует в процессе адаптации к свету, усиливая транскрипцию меланопсина в ipRGC и тем самым повышая чувствительность фоторецепторов. [3] Параллельно с ингибированием амакриновых клеток DA, амакриновые клетки, высвобождающие соматостатин, которые сами ингибируются амакриновыми клетками DA, ингибируют ipRGC. [14] Другие синаптические входы в дендриты ipRGC включают биполярные клетки колбочек и биполярные клетки палочек. [11]
Выходы
[ редактировать ]Одной из постсинаптических мишеней ipRGC является супрахиазматическое ядро (SCN) гипоталамуса, которое служит циркадными часами в организме. ipRGC высвобождают как белок, активирующий аденилатциклазу гипофиза (PACAP), так и глутамат в SCN через моносинаптическую связь, называемую ретиногипоталамическим трактом (RHT). [15] Глутамат оказывает возбуждающее действие на нейроны SCN, а PACAP, по-видимому, усиливает эффекты глутамата в гипоталамусе. [16]
Другие постсинаптические мишени ipRGC включают: межлопаточный листок (IGL), скопление нейронов, расположенных в таламусе, которые играют роль в циркадном смещении; оливковое претектальное ядро (OPN), скопление нейронов в среднем мозге, которое контролирует зрачковый световой рефлекс; вентролатеральное преоптическое ядро (ВЛПО), расположенное в гипоталамусе и являющееся центром управления сном; а также чтобы [ объяснить ] миндалевидное тело. [3]
Функция
[ редактировать ]Зрачковый рефлекс на свет
[ редактировать ]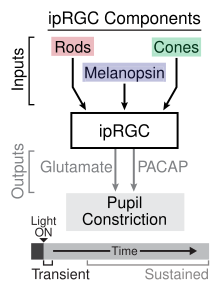
Используя мышей с нокаутом различных фоторецепторов, исследователи определили роль ipRGC как в временной, так и в постоянной передаче сигналов зрачкового светового рефлекса (PLR). [17] Временный PLR происходит при слабой и умеренной интенсивности света и является результатом фототрансдукции, происходящей в палочковидных клетках , которые обеспечивают синаптический вход на ipRGC, которые, в свою очередь, передают информацию в оливарное претектальное ядро в среднем мозге . [18] Нейромедиатором, участвующим в передаче информации в средний мозг от ipRGC во время временного PLR, является глутамат . При более яркой интенсивности света происходит устойчивая PLR, которая включает как фототрансдукцию палочки, обеспечивающей вход в ipRGC, так и фототрансдукцию самих ipRGC через меланопсин. Исследователи предположили, что роль меланопсина в устойчивом PLR обусловлена отсутствием у него адаптации к световым раздражителям в отличие от палочек, которые демонстрируют адаптацию. Устойчивый PLR поддерживается PACAP из ipRGC. пульсирующим высвобождением [17]
Возможная роль в сознательном зрении
[ редактировать ]Эксперименты с людьми без стержней и шишек позволили изучить еще одну возможную роль рецептора. В 2007 году была обнаружена новая роль фоторецепторных ганглиозных клеток. Заиди и его коллеги показали, что у людей фоторецепторы ганглиозных клеток сетчатки способствуют сознательному зрению, а также функциям, не связанным с формированием изображения, таким как циркадные ритмы, поведение и зрачковые реакции. [9] Поскольку эти клетки реагируют в основном на синий свет, было высказано предположение, что они играют роль в мезопическом зрении. [ нужна ссылка ] и что старая теория о чисто дуплексной сетчатке со палочковидным (темным) и колбочковым (светлым) зрением была упрощенной. Таким образом, работа Заиди и его коллег с людьми без стержней и колбочек также открыла дверь в формирующую изображение (визуальную) роль фоторецептора ганглиозных клеток.
Было сделано открытие, что существуют параллельные пути зрения: один классический, основанный на палочках и колбочках, исходящий из внешней сетчатки, другой - рудиментарный детектор визуальной яркости, исходящий из внутренней сетчатки. Последнее, кажется, активируется светом раньше первого. [9] Классические фоторецепторы также участвуют в новой системе фоторецепторов, и постоянство цвета может играть важную роль, как предположил Фостер. [ нужна ссылка ] .
Авторы модели человека без стержней и конусов предположили, что этот рецептор может сыграть важную роль в понимании многих заболеваний, включая основные причины слепоты во всем мире, такие как глаукома , заболевание, поражающее ганглиозные клетки.
Доказано, что у других млекопитающих светочувствительные ганглии играют реальную роль в сознательном зрении. Тесты, проведенные Дженнифер Экер и др. обнаружили, что крысы, у которых не было палочек и колбочек, могли научиться плыть к последовательностям вертикальных полос, а не к одинаково люминесцентному серому экрану. [8]
Свет от фиолетового до синего
[ редактировать ]Большинство работ предполагает, что пиковая спектральная чувствительность рецептора находится между 460 и 484 нм. Локли и др. в 2003 году [19] показали, что свет с длиной волны 460 нм (синий) подавляет мелатонин в два раза сильнее, чем свет с длиной волны 555 нм (зеленый), пиковая чувствительность фотопической зрительной системы. В работе Заиди, Локли и соавторов с использованием человека без стержней и конусов было обнаружено, что очень интенсивный стимул длиной 481 нм привел к некоторому сознательному восприятию света, а это означает, что было реализовано некоторое элементарное видение. [9]
Открытие
[ редактировать ]В 1923 году Клайд Э. Килер заметил, что зрачки глаз слепых мышей, которых он случайно вывел, все еще реагировали на свет. [2] Способность мышей без палочек и колбочек сохранять зрачковый световой рефлекс наводила на мысль о наличии дополнительных фоторецепторных клеток. [11]
В 1980-х годах исследования на крысах с дефицитом палочек и колбочек показали регуляцию дофамина в сетчатке, известного нейромодулятора, отвечающего за световую адаптацию и фотоувлечение. [3]
Исследования продолжились в 1991 году, когда Рассел Г. Фостер и его коллеги, в том числе Игнасио Провенсио , показали, что палочки и колбочки не необходимы ни для фотосинхронизации, визуального привода циркадного ритма , ни для регуляции мелатонина секреции шишковидной железой через палочки. - и мыши с выбитыми конусами. [20] [11] Более поздняя работа Провенсио и его коллег показала, что этот фотоответ опосредован фотопигментом меланопсином , присутствующим в слое ганглиозных клеток сетчатки. [21]
Фоторецепторы были идентифицированы в 2002 году Самером Хаттаром , Дэвидом Берсоном и коллегами, где было показано, что они представляют собой ганглиозные клетки, экспрессирующие меланопсин, которые обладают внутренней реакцией на свет и проецируются на ряд областей мозга, участвующих в зрении, не формирующем изображения. [22] [23]
В 2005 году Панда, Мелян, Цю и коллеги продемонстрировали, что фотопигмент меланопсин является пигментом фототрансдукции в ганглиозных клетках. [24] [25] Деннис Дейси и его коллеги показали на обезьянах Старого Света, что гигантские ганглиозные клетки, экспрессирующие меланопсин, проецируются в латеральное коленчатое ядро (LGN). [26] [6] только проекции на средний мозг (претектальное ядро) и гипоталамус ( супрахиазматические ядра Ранее были показаны , SCN). Однако визуальная роль рецептора до сих пор не подозревалась и не была доказана.
Исследовать
[ редактировать ]Исследования на людях
[ редактировать ]Были предприняты попытки выследить рецептор у людей, но люди поставили перед собой особые задачи и потребовали новую модель. В отличие от других животных, исследователи не могли с этической точки зрения вызвать потерю палочек и колбочек ни генетически, ни с помощью химических веществ, чтобы напрямую изучать ганглиозные клетки. В течение многих лет о рецепторе у человека можно было сделать только выводы, хотя иногда они были уместны.
В 2007 году Заиди и его коллеги опубликовали свою работу о людях без стержней и шишек, показав, что эти люди сохраняют нормальную реакцию на невизуальные эффекты света. [9] [27] Было обнаружено, что фоторецептор без палочек и колбочек у людей представляет собой ганглиозную клетку во внутренней сетчатке, как было показано ранее на моделях без палочек и колбочек у некоторых других млекопитающих. Работа проводилась на пациентах с редкими заболеваниями, которые уничтожили функцию классических фоторецепторов палочек и колбочек, но сохранили функцию ганглиозных клеток. [9] [27] Несмотря на отсутствие палочек и колбочек, у пациентов продолжали проявляться циркадные фотоэнцефалические изменения, циркадные поведенческие модели, подавление мелатонина и реакции зрачков с пиковой спектральной чувствительностью к окружающему и экспериментальному свету, которая соответствует фотопигменту меланопсину. Их мозг также мог ассоциировать зрение со светом этой частоты. Клиницисты и ученые сейчас пытаются понять роль нового рецептора в заболеваниях человека и слепоте. [ нужна ссылка ] Внутренне светочувствительные RGC также участвуют в усилении головной боли под воздействием света во время приступов мигрени. [28]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Киллер, Клайд Э. (1927). «Движения радужной оболочки у слепых мышей». Американский журнал физиологии . 81 (1): 107–112. дои : 10.1152/ajplegacy.1927.81.1.107 .
- ^ Перейти обратно: а б Килер CE (октябрь 1928 г.). «Слепые мыши». Журнал экспериментальной зоологии . 51 (4): 495–508. Бибкод : 1928JEZ....51..495K . дои : 10.1002/jez.1400510404 .
- ^ Перейти обратно: а б с д и ж До МТ, Яу К.В. (октябрь 2010 г.). «Самостоятельно светочувствительные ганглиозные клетки сетчатки» . Физиологические обзоры . 90 (4): 1547–81. doi : 10.1152/physrev.00013.2010 . ПМЦ 4374737 . ПМИД 20959623 .
- ^ Блюм С., Гарбацца С., Спицан М. (2019). «Влияние света на циркадные ритмы человека, сон и настроение» . Сомнология (Берл) . 23 (3): 147–156. дои : 10.1007/s11818-019-00215-x . ПМК 6751071 . ПМИД 31534436 .
- ^ Вонг К.Ю., Данн Ф.А., Берсон Д.М. (декабрь 2005 г.). «Адаптация фоторецепторов в внутренне светочувствительных ганглиозных клетках сетчатки» . Нейрон . 48 (6): 1001–10. дои : 10.1016/j.neuron.2005.11.016 . ПМИД 16364903 .
- ^ Перейти обратно: а б Берсон Д.М. (июнь 2003 г.). «Странное видение: ганглиозные клетки как циркадные фоторецепторы». Тенденции в нейронауках . 26 (6): 314–20. дои : 10.1016/S0166-2236(03)00130-9 . ПМИД 12798601 . S2CID 15149809 .
- ^ Фань С.М., Чанг Ю.Т., Чен С.Л., Ван В.Х., Пан М.К., Чен В.П. и др. (июль 2018 г.). «Внешний свет активирует стволовые клетки волосяного фолликула через глаза через симпатический нервный путь ipRGC-SCN» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (29): E6880–E6889. Бибкод : 2018PNAS..115E6880F . дои : 10.1073/pnas.1719548115 . ПМК 6055137 . ПМИД 29959210 .
- ^ Перейти обратно: а б с д и Экер Дж.Л., Думитреску О.Н., Вонг К.Ю., Алам Н.М., Чен С.К., Легейтс Т. и др. (июль 2010 г.). «Меланопсин-экспрессирующие фоторецепторы ганглиозных клеток сетчатки: клеточное разнообразие и роль в образном зрении» . Нейрон . 67 (1): 49–60. дои : 10.1016/j.neuron.2010.05.023 . ПМК 2904318 . ПМИД 20624591 .
- ^ Перейти обратно: а б с д и ж Заиди Ф.Х., Халл Дж.Т., Пирсон С.Н., Вульф К., Эшбах Д., Гули Дж.Дж. и др. (декабрь 2007 г.). «Коротковолновая световая чувствительность циркадного, зрачкового и зрительного восприятия у людей, лишенных внешней сетчатки» . Современная биология . 17 (24): 2122–8. Бибкод : 2007CBio...17.2122Z . дои : 10.1016/j.cub.2007.11.034 . ПМК 2151130 . ПМИД 18082405 .
- «Слепые люди, у которых нет палочек и колбочек, сохраняют нормальную реакцию на невизуальные эффекты света» . ЭврекАлерт! (Пресс-релиз). 13 декабря 2007 г.
- ^ Берсон Д.М. (август 2007 г.). «Фототрансдукция в фоторецепторах ганглиозных клеток» . Архив Пфлюгерса . 454 (5): 849–55. дои : 10.1007/s00424-007-0242-2 . ПМИД 17351786 .
- ^ Перейти обратно: а б с д и Колб Х., Фернандес Э., Нельсон Р. (1 января 1995 г.). «Меланопсин-экспрессирующие внутренне фоточувствительные ганглиозные клетки сетчатки (IpRGC)». В Колб Х., Фернандес Э., Нельсон Р. (ред.). Webvision: организация сетчатки и зрительной системы . Центр медицинских наук Университета Юты. ПМИД 21413389 .
- ^ До М.Т., Кан С.Х., Сюэ Т., Чжун Х., Ляо Х.В., Берглс Д.Э., Яу К.В. (январь 2009 г.). «Захват фотонов и передача сигналов меланопсиновыми ганглиозными клетками сетчатки» . Природа . 457 (7227): 281–7. Бибкод : 2009Natur.457..281D . дои : 10.1038/nature07682 . ПМК 2794210 . ПМИД 19118382 .
- ^ Челлаппа С.Л., Ли Дж.К., Мейер С., Балто Е., Дегельдре С., Люксен А. и др. (апрель 2014 г.). «Фотическая память для исполнительных реакций мозга» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (16): 6087–91. Бибкод : 2014PNAS..111.6087C . дои : 10.1073/pnas.1320005111 . ПМК 4000819 . ПМИД 24616488 .
- ^ Вуонг Х.Э., Харди С.Н., Барнс С., Бреча, Северная Каролина (декабрь 2015 г.). «Параллельное ингибирование дофамин-амакриновых клеток и внутренне светочувствительных ганглиозных клеток сетчатки в зрительной цепи сетчатки мыши, не формирующей изображение» . Журнал неврологии . 35 (48): 15955–70. doi : 10.1523/jneurosci.3382-15.2015 . ПМК 4666919 . ПМИД 26631476 .
- ^ Колвелл К.С., Мишель С., Итри Дж., Родригес В., Тэм Дж., Лельевр В. и др. (ноябрь 2004 г.). «Селективный дефицит циркадной реакции на свет у мышей, лишенных PACAP». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 287 (5): Р1194-201. дои : 10.1152/ajpregu.00268.2004 . ПМИД 15217792 .
- ^ Мясник GQ, Ли Б., Ченг Х.И., Обриетан К. (июнь 2005 г.). «Свет стимулирует активацию MSK1 в супрахиазматическом ядре посредством киназно-зависимого механизма PACAP-ERK/MAP» . Журнал неврологии . 25 (22): 5305–13. doi : 10.1523/jneurosci.4361-04.2005 . ПМК 6724997 . ПМИД 15930378 .
- ^ Перейти обратно: а б Кинан В.Т., Рупп А.К., Росс Р.А., Сомасундарам П., Хирияна С., Ву З. и др. (сентябрь 2016 г.). «Зрительная цепь использует дополнительные механизмы для поддержки временного и устойчивого сужения зрачков» . электронная жизнь . 5 . дои : 10.7554/eLife.15392 . ПМК 5079752 . ПМИД 27669145 .
- ^ Гамлин П.Д., Макдугал Д.Х., Покорни Дж., Смит В.К., Яу К.В., Дейси Д.М. (март 2007 г.). «Реакция зрачков человека и макака, обусловленная меланопсинсодержащими ганглиозными клетками сетчатки» . Исследование зрения . 47 (7): 946–54. дои : 10.1016/j.visres.2006.12.015 . ЧВК 1945238 . ПМИД 17320141 .
- ^ Локли С.В., Брейнард Г.К., Чейслер, Калифорния (сентябрь 2003 г.). «Высокая чувствительность циркадного ритма мелатонина человека к сбросу коротковолнового света» . Журнал клинической эндокринологии и метаболизма . 88 (9): 4502–5. дои : 10.1210/jc.2003-030570 . ПМИД 12970330 .
- ^ Фостер Р.Г., Провенсио I, Хадсон Д., Фиске С., Де Грип В., Менакер М. (июль 1991 г.). «Циркадный фоторецепция у мышей с дегенерацией сетчатки (rd/rd)». Журнал сравнительной физиологии A: Сенсорная, нервная и поведенческая физиология . 169 (1): 39–50. дои : 10.1007/BF00198171 . ПМИД 1941717 . S2CID 1124159 .
- ^ Провенсио I, Родригес И.Р., Цзян Г., Хейс В.П., Морейра Э.Ф., Роллаг, доктор медицинских наук (январь 2000 г.). «Новый человеческий опсин во внутренней сетчатке» . Журнал неврологии . 20 (2): 600–5. doi : 10.1523/jneurosci.20-02-00600.2000 . ПМК 6772411 . ПМИД 10632589 .
- ^ Берсон Д.М., Данн Ф.А., Такао М. (февраль 2002 г.). «Фототрансдукция ганглиозными клетками сетчатки, которые устанавливают циркадные часы». Наука . 295 (5557): 1070–3. Бибкод : 2002Sci...295.1070B . дои : 10.1126/science.1067262 . ПМИД 11834835 . S2CID 30745140 .
- ^ Хаттар С., Ляо Х.В., Такао М., Берсон Д.М., Яу К.В. (февраль 2002 г.). «Меланопсин-содержащие ганглиозные клетки сетчатки: архитектура, проекции и внутренняя фоточувствительность» . Наука . 295 (5557): 1065–70. Бибкод : 2002Sci...295.1065H . дои : 10.1126/science.1069609 . ПМЦ 2885915 . ПМИД 11834834 .
- ^ Панда С., Наяк С.К., Кампо Б., Уокер Дж.Р., Хогенеш Дж.Б., Джегла Т. (январь 2005 г.). «Освещение сигнального пути меланопсина». Наука . 307 (5709): 600–4. Бибкод : 2005Sci...307..600P . дои : 10.1126/science.1105121 . ПМИД 15681390 . S2CID 22713904 .
- ^ Цю X, Кумбаласири Т., Карлсон С.М., Вонг К.Ю., Кришна В., Провенсио I, Берсон Д.М. (февраль 2005 г.). «Индукция фоточувствительности за счет гетерологичной экспрессии меланопсина». Природа . 433 (7027): 745–9. Бибкод : 2005Natur.433..745Q . дои : 10.1038/nature03345 . ПМИД 15674243 . S2CID 24999816 .
- ^ Дейси Д.М., Ляо Х.В., Петерсон Б.Б., Робинсон Ф.Р., Смит В.К., Покорный Дж. и др. (февраль 2005 г.). «Ганглионарные клетки, экспрессирующие меланопсин, в сетчатке приматов сигнализируют о цвете и освещенности и проецируются на LGN». Природа . 433 (7027): 749–54. Бибкод : 2005Natur.433..749D . дои : 10.1038/nature03387 . ПМИД 15716953 . S2CID 4401722 .
- ^ Перейти обратно: а б Коглан А (2007). «Как слепые видят восход и закат» . Новый учёный . 196 (2635–2636): 9. doi : 10.1016/S0262-4079(07)63172-8 .
- ^ Нозеда Р. (2010). «Нейральный механизм усиления головной боли от света» . Природная неврология . 13 (2): 239–245. дои : 10.1038/nn.2475 . ПМЦ 2818758 . ПМИД 20062053 .
