Визуальный цикл
Эта статья нуждается в дополнительных цитатах для проверки . ( май 2017 г. ) |
Зрительный цикл — это процесс в сетчатке, который пополняет молекулы сетчатки для ее использования в зрении . Ретиналь является хромофором большинства зрительных опсинов , то есть он захватывает фотоны, чтобы начать каскад фототрансдукции . Когда фотон поглощается, 11-цис-ретиналь фотоизомеризуется в полностью транс-ретиналь, выбрасываясь из белка опсина. Каждая молекула ретиналя должна пройти путь от фоторецепторной клетки к РПЭ и обратно, чтобы обновиться и соединиться с другим опсином. Этот закрытый ферментативный путь иногда 11-цис сетчатки называют зрительным циклом Уолда в честь Джорджа Уолда (1906–1997), получившего Нобелевскую премию в 1967 году за работу по его открытию.
сетчатка
[ редактировать ]Ретиналь представляет собой хромофор , который образует светочувствительные ретинилиденовые белки при ковалентной связи с белками, называемыми опсинами . Ретиналь может быть фотоизомеризован сам по себе, но его необходимо связать с белком опсином, чтобы запустить каскад фототрансдукции и настроить спектральную чувствительность на более длинные волны, что обеспечивает цветовое зрение .
это разновидность ретиноида и альдегидной формы витамина А. Ретиналь – Ретиналь взаимопревращается с ретинолом , формой транспорта и хранения витамина А. Во время зрительного цикла ретиналь перемещается между несколькими различными изомерами, а также превращается в ретинол и ретиниловый эфир . Ретиноиды могут быть получены в результате окисления каротиноидов, таких как бета-каротин , или могут потребляться напрямую. Чтобы достичь сетчатки, он связывается с ретинол-связывающим белком (RBP) и транстиретином , что предотвращает его фильтрацию в клубочках .
Как и при транспорте по пути RBP-транстиретин, ретиноиды всегда должны быть связаны с молекулами шаперона по нескольким причинам. Ретиноиды токсичны, нерастворимы в водных растворах и склонны к окислению, поэтому при попадании в организм их необходимо связывать и защищать. Для транспортировки ретиноидов организм использует различные шапероны, особенно в сетчатке.
Обзор
[ редактировать ]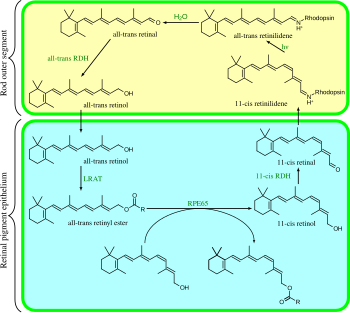
одинаков Зрительный цикл у млекопитающих и резюмируется следующим образом:
- полностью транс- ретиниловый эфир + H 2 O → 11 -цис -ретинол + жирная кислота ; RPE65 ; изомерогидролазы [1]
- 11- цис -ретинол + НАД + → 11- цис -ретиналь + НАДН + Н + ; 11- цис -ретинолдегидрогеназы;
- 11- цис -ретиналь + апорходопсин → родопсин + H 2 O; образует связь основания Шиффа с лизином , -CH=N + ЧАС-;
- родопсин + hν → метародопсин II (т.е. 11- цис- фотоизомеризуется до полностью- транс ):
- (родопсин + hν → фотородопсин → батородопсин → люмирродопсин → метародопсин I → метародопсин II);
- метародопсин II + H 2 O → апорходопсин + all- транс -ретиналь;
- полностью транс -ретиналь + НАДФН + H + → полностью транс -ретинол + НАДФ + ; полностью транс ретинолдегидрогеназы ; -
- полностью транс -ретинол + жирная кислота → полностью транс- ретиниловый эфир + H 2 O; лецитин-ретинолацилтрансферазы (LRAT). [2]
Шаги 3, 4, 5 и 6 происходят во внешних сегментах палочек ; Этапы 1, 2 и 7 происходят в клетках пигментного эпителия сетчатки (RPE).
Описание
[ редактировать ]При фотона поглощении 11- цис -ретиналь превращается в полностью транс -ретиналь и перемещается к месту выхода родопсина . Он не покинет белок опсин до тех пор, пока на его место не придет другой свежий хромофор, за исключением пути ABCR. Пока он все еще связан с опсином, полностью транс -ретиналь превращается в полностью транс -ретинол под действием полностью транс -ретинолдегидрогеназы. Затем он переходит к клеточной мембране палочки, где связывается с пигментным эпителием сетчатки (RPE) с помощью межфоторецепторного ретиноид-связывающего белка (IRBP). Затем он проникает в клетки РПЭ и переносится в шаперон клеточного ретинол-связывающего белка (CRBP).
Находясь внутри клетки RPE, связанный с CRBP, полностью транс -ретинол этерифицируется лецитин-ретинол-ацилтрансферазой (LRAT) с образованием сложного эфира ретинила. Ретиниловые эфиры RPE сопровождаются белком, известным как RPE65 . Именно в этой форме РПЭ хранит большую часть своих ретиноидов, поскольку РПЭ хранит в 2-3 раза больше ретиноидов, чем сама нейронная сетчатка. Когда требуется дополнительный хромофор, сложные эфиры ретинила подвергаются воздействию изомерогидролазы с образованием 11- цис -ретинола, который переносится на клеточный ретинальдегид-связывающий белок (CRALBP). 11- цис -ретинол трансформируется в 11-цис-ретиналь под действием 11-цис-ретинолдегидрогеназы , затем доставляется обратно в фоторецепторные клетки посредством IRBP . Там он заменяет отработанный хромофор в молекулах опсина, делая опсин светочувствительным.
Путь ABCR
[ редактировать ]В нормальных условиях отработанный хромофор высвобождается из белка поступающим «перезаряженным» хромофором. Однако иногда отработанный хромофор может покинуть белок опсин до его замены, когда он связан с белком ABCA4 (также известным как ABCR). На этом этапе он также трансформируется в полностью транс -ретинол, а затем покидает внешний сегмент фоторецептора через шаперон IRBP. Затем следует обычный визуальный цикл. Именно этим путем можно объяснить наличие опсина без хромофора.
регулирование РГР
[ редактировать ]Зрительный цикл может регулироваться системой рецепторов, связанных с G-белком (RGR-опсин) . Когда свет активирует RGR-опсин, рециркуляция хромофора в RPE ускоряется. Этот механизм обеспечивает дополнительный хромофор после интенсивного отбеливания и может рассматриваться как важный механизм на ранних стадиях темновой адаптации и пополнения хромофора.
Альтернативные циклы
[ редактировать ]Визуальный цикл, специфичный для конуса
[ редактировать ]Считается, что существует альтернативный зрительный цикл, в котором используются глиальные клетки Мюллера вместо пигментного эпителия сетчатки . В этом пути колбочки восстанавливают полностью транс-ретинол до полностью-транс-ретинола с помощью полностью-транс-ретинолдегидрогеназы, а затем транспортируют полностью-транс-ретинол в клетки Мюллера. Там он трансформируется в 11-цис-ретинол под действием полностью транс-ретинол-изомеразы и может либо храниться в виде эфиров ретинила в клетках Мюллера, либо транспортироваться обратно к фоторецепторам колбочек, где он трансформируется из 11-цис-ретинола в 11-цис. сетчатки с помощью 11-цис-дегидрогеназы сетчатки. Этот путь помогает объяснить быструю адаптацию к темноте в системе колбочек и присутствие 11-цис-дегидрогеназы сетчатки в фоторецепторах колбочек, поскольку она не обнаружена в палочках, а только в RPE. [3]
Зрительный цикл меланопсина
[ редактировать ]Меланопсин представляет собой зрительный опсин, присутствующий в внутренне светочувствительных ганглиозных клетках сетчатки (ipRGC), а также с хромафором сетчатки. Однако, в отличие от пигментов палочек и колбочек, меланопсин обладает способностью действовать как возбудимый фотопигмент, так и как фотоизомераза . Таким образом, меланопсин способен изомеризовать весь транс- ретиналь в 11-цис- ретиналь при стимуляции другим фотоном. Таким образом, ipRGC не полагается на клетки Мюллера и/или клетки пигментного эпителия сетчатки для этого преобразования. [4]
Врожденный амороз Лебера
[ редактировать ]Возможный механизм врожденного амавроза Лебера был предложен как дефицит RPE65 . Без белка RPE65 RPE не может хранить сложные эфиры ретинила, и поэтому зрительный цикл прерывается. На начальных стадиях заболевания колбочки не поражаются, поскольку они могут полагаться на альтернативный зрительный цикл клеток Мюллера. Однако стержни не имеют доступа к этой альтернативе и становятся инертными. Таким образом, LCA проявляется как никталопия (куриная слепота). На более поздних стадиях заболевания наблюдается общая ретинопатия, поскольку палочки теряют способность передавать сигналы. В результате палочки постоянно выделяют глутамат , нейромедиатор, со скоростью, которую клетки Мюллера не могут усвоить. Уровни глутамата будут накапливаться в сетчатке, где они достигнут нейротоксичного уровня. Дефицит RPE65 может иметь генетическое происхождение и является лишь одной из многих предполагаемых возможных патофизиологий заболевания. Тем не менее, существует генная терапия сетчатки для восстановления нормальных генов RPE65, одобренная FDA с 2017 года. [5]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Моисеев Геннадий; Чен, Ин; Такахаси, Юсуке; Ву, Билл X.; Ма, Цзянь-син (2005). «RPE65 — это изомерогидролаза ретиноидного зрительного цикла» . Труды Национальной академии наук . 102 (35): 12413–12418. Бибкод : 2005PNAS..10212413M . дои : 10.1073/pnas.0503460102 . ПМЦ 1194921 . ПМИД 16116091 .
- ^ Цзинь, Минхао; Юань, Цюань; Ли, Сунгари; Трэвис, Габриэль Х. (2007). «Роль LRAT в активности ретиноид-изомеразы и мембранной ассоциации Rpe65» . Журнал биологической химии . 282 (29): 20915–20924. дои : 10.1074/jbc.M701432200 . ПМЦ 2747659 . ПМИД 17504753 .
- ^ Ван, Цзинь-Шань; Кефалов, Владимир Дж. (март 2011 г.). «Визуальный цикл, характерный для Конуса» . Прогресс в исследованиях сетчатки и глаз . 30 (2): 115–128. doi : 10.1016/j.preteyeres.2010.11.001 . ПМЦ 3073571 . ПМИД 21111842 .
- ^ Колб Х., Фернандес Э., Нельсон Р. (1 января 1995 г.). «Меланопсин-экспрессирующие внутренне фоточувствительные ганглиозные клетки сетчатки (IpRGC)». В Колб Х., Фернандес Э., Нельсон Р. (ред.). Webvision: организация сетчатки и зрительной системы . Центр медицинских наук Университета Юты. ПМИД 21413389 .
- ^ «Одобренная продукция – Luxturna» . Центр FDA по оценке и исследованиям биологических препаратов. 19 декабря 2017 г.