Казеиды
| Казеиды Временной диапазон: поздний карбон – средняя пермь ,
| |
|---|---|

| |
| Ископаемый скелет Cotylorhynchus romeri | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Сторона синапса |
| Клэйд : | † Казеазаврия |
| Семья: | † Казеиды Уиллистон , 1912 год. |
| Роды | |
Caseidae — вымершее семейство базальных , жившее синапсид от позднего карбона до средней перми примерно 300–265 миллионов лет назад. Окаменелости этих животных происходят из южно-центральной части США ( Техас , Оклахома и Канзас ) , из различных частей Европы ( европейская часть России , Франция , Германия , Сардиния и Польша ) и, возможно, из Южной Африки , если Род Eunotosaurus действительно является казеидом, как предположили некоторые авторы в 2021 году. Казеиды демонстрируют большое таксономическое и морфологическое разнообразие. Наиболее базальными таксонами были мелкие насекомоядные и всеядные формы, обитавшие преимущественно в верхнем карбоне и нижней перми, такие как Eocasea , Callibrachion и Martensius . Этот тип казеида сохраняется до средней перми с Phreatophasma и может быть Eunotosaurus . В раннюю пермь клада представлена в основном многими видами , перешедшими на растительноядный рацион. Некоторые из них превратились в гигантские формы, которые могут достигать 6–7 метров (20–23 футов) в длину, например Cotylorhynchus hancocki. и Alierasaurus ronchii , что делает их крупнейшими пермскими синапсидами. Казеиды считаются важными компонентами ранних наземных экосистем в истории позвоночных, поскольку многочисленные травоядные виды этого семейства являются одними из первых наземных четвероногих , которые взяли на себя роль основного потребителя . Казеиды испытали значительную эволюционную радиацию в конце ранней перми, став вместе с капторинидными эврептилиями доминирующими травоядными наземных экосистем вместо эдафозавридов и диадектид .
В 2016 и 2022 годах палеонтологи предположили полуводный образ жизни для наиболее производных родов, таких как Cotylorhynchus и Lalieudorhynchus , но эта гипотеза оспаривается другими исследователями.
Летопись окаменелостей простирается от позднего карбона до средней перми, и казеиды являются одной из групп базальных синапсид (ранее известных как « пеликозавр »), имеющей самый большой стратиграфический диапазон. Они также представляют собой одну из двух групп базальных синапсид (наряду с Varanopidae ), выживших в наземных сообществах с доминированием терапсидов . Таким образом, последние известные казеиды происходят из толщ средней перми (верхнего рога — ворда ) Франции и Европейской России, где они сожительствовали преимущественно с диноцефалами . [ номер 1 ] Эти последние казеиды все еще демонстрируют определенное морфологическое разнообразие со средними и крупными травоядными ( Ennatosaurus и Lalieudorhynchus ), а также мелкими хищными или всеядными формами, такими как Phreatophasma и, возможно, Eunotosaurus . Казеиды пока неизвестны в толщах верхней перми и, вероятно, исчезли в конце средней перми. На смену им пришли парейазавры (со схожим зубным рядом) и дицинодонты .
Описание
[ редактировать ]


Казеиды имеют длину от менее 1 метра (3 футов 3 дюйма) до 7 метров (23 фута). [ 1 ] [ 2 ] [ 3 ] У них была небольшая голова шириной больше высоты и с наклоненной вперед мордой, очень короткая шея, длинный хвост, крепкие передние конечности и тело различных пропорций в зависимости от их рациона. Мелкие насекомоядные виды, такие как Eocasea, имели нерасширенный туловище. [ 4 ] Другие, придерживавшиеся всеядной диеты, такие как Мартенсиус, имели едва увеличенную грудную клетку, более удлиненный череп, меньшие ноздри и морду, менее наклоненную вперед, чем у травоядных казеидов. [ 5 ] Последние отличались непропорционально маленьким черепом по сравнению с размером тела. Посткраниальный скелет действительно демонстрирует впечатляющее увеличение объема грудной клетки , которая становится очень широкой и бочкообразной, вероятно, для размещения особенно развитого кишечника , необходимого для переваривания растений с высоким содержанием клетчатки. У этих форм череп имеет очень большие наружные ноздри и очень короткую лицевую область с сильным наклоном вперед конца рыла, который явно нависает над зубным рядом. Височные отверстия также относительно крупные (особенно у эннатозавра ), надвисочные отверстия имеют большие размеры, а на затылочной околозатылочные отростки поверхности массивно развиты , устанавливающие прочные опорные контакты с чешуйчатыми костями . [ 6 ] Дорсальная поверхность черепа покрыта многочисленными мелкими ямками. Это позволяет предположить наличие крупной чешуи на голове этих животных. [ 7 ] Многочисленные губные отверстия, идущие параллельно вентральному краю предчелюстной и верхней челюсти , а также вдоль дорсального края зубной кости , предполагают наличие чешуйчатых «губ», которые, должно быть, скрывали зубной ряд, когда челюсти были закрыты. [ 8 ] [ 7 ] Зубы , просто конические и заостренные у насекомоядных видов, у травоядных принимают листовидную или лопатчатую форму и снабжены более или менее многочисленными бугорками. Многочисленные мелкие зубы украшали также несколько костей неба. Травоядные виды не демонстрируют простой эволюционной тенденции к увеличению сложности зубов. [ 9 ] Так, зубы базальных таксонов Casea и Arisierpeton имеют три бугорки. [ 10 ] [ 11 ] точно так же, как и в более производных формах Cotylorhynchus и Caseopsis . [ 10 ] Эннатозавр и евромиктер , занимающие промежуточное филогенетическое положение, имеют зубы с 5–7 и 5–8 бугорками соответственно. [ 9 ] [ 12 ] Ангелозавр , один из самых производных казеидов, имеет зубы с 5 бугорками. [ 10 ] У ангелозавра зубы имеют луковицеобразную форму с очень короткими и широкими коронками. Их прочность и значительный износ указывают на то, что ангелозавр, должно быть, питался более жесткими растениями, чем те, которыми питалось большинство других травоядных казеидов. [ 10 ] Травоядные казеиды также демонстрируют совсем другие диетические адаптации по сравнению с теми, которые наблюдаются у другой группы базальных синапсид, Edaphosauridae . Последние имели, помимо краевого зубного ряда, зубную батарею, состоящую из многочисленных зубов, расположенных как на небе , так и на внутренней поверхности нижних челюстей. У травоядных казеидов небные зубы мельче, а внутренняя поверхность нижних челюстей зубов не имеет. Вместо зубной батареи у них был массивный язык (на что указывает наличие высокоразвитого подъязычного аппарата, обнаруженного у эннатозавра и евромиктера ), возможно, шероховатый, которым им приходилось прижимать пищу к небным зубам. [ 10 ] [ 8 ]


Передние конечности казеидов часто более крепкие, чем задние. Действительно, было замечено, что кости передних конечностей приобрели прочность с самого начала эволюции группы, до появления крупных видов, тогда как задние конечности оставались более тонкими. Эти характеристики позволяют предположить, что первоначальное усиление передних конечностей, вероятно, было связано с определенной функцией, такой как рытье, и что эта черта позже была использована более развитыми и более крупными казеидами, чтобы выдерживать их вес до нескольких сотен килограммов. [ 1 ] В ходе своей эволюционной истории казеиды также демонстрируют редукцию формулы фаланг. [ номер 2 ] Наиболее базальные казеиды, такие как Eoasea , Callibrachion и Martensius , обладали плезиоморфным состоянием ранних амниот с фаланговой формулой 2-3-4-5-3 для кисти и 2-3-4-5-4 для стопы . [ 4 ] [ 13 ] [ 5 ] У Euromycter рука имеет формулу 2-3-4-4-3 (пес неизвестен). [ 12 ] У более производных форм, таких как Cotylorhynchus, кисть и стопа имеют фаланговую формулу 2-2-3-3-2. [ номер 3 ] [ 14 ] [ 15 ] [ 10 ] [ 16 ] Наряду с этим уменьшением количества фаланг у производных казеидов также изменяются пропорции автоподий: пястные кости , плюсневые кости и фаланги становятся короче и шире. На крайнем уровне этой специализации род Angelosaurus имеет короткие, широкие и гладкие когтевые фаланги, которые напоминают копыта, а не когти . [ 10 ] [ 17 ]
Следы
[ редактировать ]Было предложено отнести следы многих позвоночных к Caseidae. В начале 2000-х годов считалось, что большие следы, известные как Brontopus giganteus из пермского бассейна Лодев на юге Франции, принадлежат либо казеидам, либо диноцефаловым терапсидам . [ 18 ] Однако в 2019 году Лоренцо Маркетти и его коллеги определили, что диноцефалы, скорее всего, были следопытами ихногена Brontopus . [ 19 ] В 2012 году Рафаэль Коста да Силва и его коллеги предположили, что ихноген Chelichnus , широко распространенный в пермских пустынных фациях (ископаемых дюнах ) Европы , Северной Америки и Южной Америки , может представлять собой следы казеид. [ 20 ] Однако в 2019 году Маркетти и его коллеги переосмыслили Chelichnus как nomen dubium и тафотаксон , этот тип следов демонстрирует ложные анатомические особенности, возникающие в результате передвижения по песчаной наклонной палеоповерхности. [ 21 ] В 2014 году Ева Сакки и ее коллеги описали ихнотаксон Dimetropus osageorum примерно по сотне изолированных следов и нескольких следов из ранней пермской формации Веллингтон в Оклахоме . Морфологическое изучение этих следов и сравнение их со скелетами четвероногих показало, что они, вероятно, принадлежат крупному казеиду, сравнимому по размерам с Cotylorhynchus romeri (последний из более поздних слоев). [ 22 ] По мнению Сакки и его коллег, некоторые следы из бассейна Лодев (неуточненная геологическая формация) аналогичны следам из формации Веллингтон, и эти авторы обозначают их как Dimetropus cf. осагеорум . [ 22 ] Исследование Sacchi et al. также демонстрирует, что ихнород Dimetropus демонстрирует большую морфологическую изменчивость и что его продуценты могут быть отнесены к различным зоологическим группам среди нетерапсидных синапсидов, а не только к Sphenacodontidae , как когда-то считалось. [ 22 ] В 2021 году Рафель Маталес-Андреу и его коллеги приписали следы, обнаруженные в формации Порт-де-Канож нижней перми ( артинско - кунгурский период ) на Майорке , казеиду. Эти следы напоминают следы D. osageorum, но имеют и различия. Эти следы оставлены в открытой номенклатуре, см. Dimetropus sp. Однако они меньше, чем у D. osageorum , и могли быть произведены казеидами скромных размеров с пропорциями, сравнимыми с эннатозаврами . [ 23 ]
Палеобиология
[ редактировать ]Диеты
[ редактировать ]

В ходе своей эволюционной истории казеиды перешли от фауноядной к травоядной диете, и эта закономерность также наблюдалась независимо у других каменноугольных и пермских групп четвероногих, таких как Captorhinidae и Edaphosauridae . Самые ранние и большинство базальных казеидов, такие как позднекаменноугольная Eocasea и раннепермский Callibrachion, имели нерасширенную грудную клетку и зубной ряд, состоящий из очень маленьких конических зубов, что позволяет предположить насекомоядную диету. Другой базальный казеид, Martensius , имеет слегка увеличенный бочкообразный туловище и зубной ряд, в котором зубы, указывающие на насекомоядную диету у молодых особей, были онтогенетически заменены у взрослых зубами, предполагающими всеядную диету. У Martensius взрослая особь все еще могла питаться насекомыми, но она также обладает некоторыми чертами растительноядной диеты, присутствующими у более поздних казеид, такими как относительно короткая, слегка наклоненная вперед морда и почти гомодонтный зубной ряд . верхних челюстях и полностью гомодонтных на нижних челюстях. Последовательность приобретения зубных признаков у Мартенсиус предполагает, что кишечной обработка растительности предшествовала пероральной в эволюции казеидных травоядных обработке. Диета насекомоядных молоди обеспечила бы возможность успешного введения в кишечник микроорганизмов, способных к эндосимбиотическому целлюлолизу , особенно если потребляемая добыча была травоядными насекомыми, которые содержат такие микроорганизмы в своих внутренних органах . Впоследствии казеиды перешли на строго травоядную диету и превратились в гигантские формы. Эти травоядные казеиды имели лопатчатые зубы, снабженные более или менее многочисленными бугорками, а также очень увеличенную бочкообразную грудную клетку, в которой, должно быть, располагался высокоразвитый кишечник, необходимый для переваривания растений с низкой пищевой ценностью. Эта адаптация частично объясняет диверсификацию и расширение группы в конце нижней и средней перми, поскольку она позволила им использовать богатые волокнами растительные ресурсы, которые к тому времени стали обильными и широко распространенными. Тем не менее, небольшие, вероятно фауноядные казеиды, такие как Фреатофазма , по-видимому, сохранялась до средней перми. [ 4 ] [ 13 ] [ 24 ] [ 5 ]
Полуводный и наземный образ жизни
[ редактировать ]
Казеиды обычно считаются преимущественно наземными животными. Эверетт К. Олсон, в частности, считал, что степень окостенения скелета, относительно короткие ступни и руки, массивные когти, конечности с очень мощными мышцами-разгибателями и прочный крестец убедительно свидетельствуют о наземном образе жизни. Олсон не исключал, что казеиды некоторое время проводят в воде, но считал передвижение по суше важным аспектом их образа жизни. [ 10 ] Было высказано предположение, что очень мощные передние конечности с сильными и очень сухожильными мышцами-разгибателями, а также очень массивными когтями можно было использовать для выкапывания корней или клубней. [ 10 ] Однако очень короткая шея подразумевала низкую амплитуду вертикальных движений головы, что не позволяло крупным видам питаться на уровне земли. [ 25 ] Другая гипотеза предполагает, что казеиды могли использовать свои мощные передние конечности, чтобы сгибать к себе крупные растения, которые они бы отрывали своими мощными когтями. [ 10 ] Другие гипотезы предполагают, что некоторые казеиды, такие как Cotylorhynchus, использовали свои конечности с мощными когтями для защиты от хищников или во время внутривидовой деятельности, связанной, в частности, с размножением . По словам Олсона, интересно то, что почти у всех известных экземпляров вида Cotylorhynchus hancocki в течение жизни сломано и зажило от одного до десяти ребер. [ 15 ] [ 10 ] Наконец, по мнению некоторых авторов, крупные производные казеиды были полуводными животными, которые использовали свои руки с большими когтями, похожими на лопасти , которые также можно было использовать для манипулирования растениями, которыми они питались. [ 25 ]
В 2016 году Маркус Ламбертц и его коллеги поставили под сомнение наземный образ жизни крупных казеид, таких как Cotylorhynchus . Эти авторы показали, что микроструктура костей плечевой, бедренной кости и ребер взрослых и неполовозрелых экземпляров Cotylorhynchus romeri больше напоминала таковую водных животных, чем наземных, кости имели очень губчатое строение, чрезвычайно тонкую кору и не имели отчетливый медуллярный канал . Такая низкая плотность костей была бы помехой для животных весом в несколько сотен килограммов, ведущих строго наземный образ жизни. Ламбертц и др. также утверждали, что суставы между позвонками и спинными ребрами допускают лишь небольшой диапазон движений грудной клетки, тем самым ограничивая реберную вентиляцию. Чтобы преодолеть эту проблему, они предположили наличие протодиафрагмы для облегчения дыхания, особенно в водной среде. Эти авторы также считают, что засушливый палеоклимат , которому соответствуют казеидные отложения, не является несовместимым с водным образом жизни этих животных. Эти палеосреды фактически включали значительное количество водоемов (рек, озер и лагун). Засушливые условия могли быть причиной того, что животные иногда собирались вместе и в конечном итоге умирали. Кроме того, в засушливой среде низкая плотность растений, что потребует еще больше двигательных усилий для поиска пищи. Таким образом, по мнению Ламбертца и др., крупные казеиды, такие как Cotylorhynchus , должно быть, были главным образом водными животными, пришедшими на сушу только с целью размножения или терморегуляции . [ 25 ]
Однако эта гипотеза оспаривается Кеннетом Ангельчиком и Кристианом Каммерером, а также Робертом Рейсом и его коллегами на основании палеонтологических и тафономических данных в сочетании с отсутствием у этих крупных казеид морфологических адаптаций к водному образу жизни. По данным Angielczyk и Kammerer, низкая плотность костной ткани казеидов, выявленная Lambertz et al. не похож на скелет полуводных животных, которые, как правило, имеют более сильно окостеневший скелет, обеспечивающий пассивный контроль плавучести и повышенную устойчивость к воздействию течения и волн. Микроструктура костей Cotylorhynchus больше похожа на ту, что наблюдается у животных, живущих в открытом океане , таких как китообразные и ластоногие , у которых подчеркивается высокая маневренность, быстрое ускорение и гидродинамический контроль плавучести. Однако морфология казеидов была совершенно несовместима с пелагическим образом жизни. Таким образом, из-за этих необычных данных Ангельчик и Каммерер считают, что имеющихся доказательств все еще недостаточно, чтобы поставить под сомнение более широко предполагаемый наземный образ жизни казеидов. [ 2 ] Роберт Рейс и его коллеги также оспаривают предполагаемый полуводный образ жизни казеид на том факте, что последние не обладают морфологическими адаптациями к водному образу жизни, а в случае вида Cotylorhynchus romeri - на интерпретации, согласно которой это животное жило в сухой среде в течение длительного времени. часть года, о чем свидетельствует присутствие многочисленных скелетов амфибий Brachydectes, сохранившихся в зимнем состоянии, и двоякодышащей рыбы Gnathorhiza , еще одного известного эстиватора. [ 8 ]
В 2022 году Вернебург и его коллеги предложили несколько иной полуводный образ жизни, при котором крупные казеиды, такие как Lalieudorhynchus (чья текстура костей еще более остеопорозна, чем у Cotylorhynchus ), будут экологическими эквивалентами современных бегемотов , проводящих часть своего времени в воде (находясь под водой). ходоки, а не плавающие животные), но приходящие на сушу за едой. [ 26 ]
Эволюция
[ редактировать ]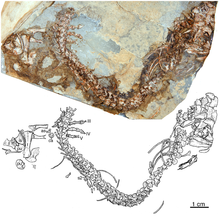

Казеиды впервые появляются в летописи окаменелостей в конце каменноугольного периода и уже присутствуют как в Северной Америке , так и в Европе вместе с небольшими хищными формами, такими как Eocasea и Datheosaurus . [ 4 ] [ 13 ] Эти фауноядные казеиды существуют и в начале нижней перми, например Callibrachion , [ 13 ] и, возможно, существовали до основания средней перми с Phreatophasma . [ 24 ] В перми представители клады в основном эволюционировали в сторону травоядности. Эти травоядные казеиды испытали важную эволюционную радиацию во второй половине нижней перми, став вместе с морадизауриновыми капторинидными эврептилиями основными травоядными наземных экосистем вместо эдафозавридов и диадектид . Уменьшение численности эдафозавридов и распространение казеид совпадает с аридизацией окружающей среды, произошедшей с артинского века и благоприятствовавшей распространению казеид. [ 27 ] [ 26 ] Седиментологические и тафономические данные свидетельствуют о том, что большинство эдафозавридов предпочитало влажные местообитания (их останки встречаются преимущественно в озерных и болотистых отложениях), тогда как травоядные казеиды обычно обитали в более сухих биотопах , где вода в прудах и ручьях была эфемерной. [ 28 ] Последние казеиды датируются средней перми и известны в Европе главным образом вместе с Phreatophasma и Ennatosaurus из роудско - вордского периода России, [ 29 ] [ 30 ] [ 31 ] Алиеразавр из Родиана-(Вордиана?) Сардинии, [ 32 ] [ 33 ] [ 26 ] и Lalieudorhynchus из позднего слова - раннего капитана южной Франции. [ 26 ] В Северной Америке Angelosaurus romeri и Cotylorhynchus bransoni из формации Чикаша в Оклахоме, вероятно, относятся к раннему роудиану, тогда как казеиды из формации Сан-Анджело в Техасе немного старше и могут располагаться между кунгурскими и роадскими границами, но пока неясно, являются ли они они датируются поздним кунгурским или самым ранним роудским периодом. [ 34 ]
Палеобиогеография
[ редактировать ]

казеидов Окаменелости были найдены на юге и в центральной части США ( Техас , Оклахома , Канзас ), Франции , Германии , Польше , Сардинии и европейской части России . Такое географическое распространение соответствует палеоэкваториальному поясу Пангеи в каменноугольном периоде и перми, за исключением российских местонахождений, располагавшихся на уровне 30-й параллели с.ш. , где преобладал более умеренный климат. Отсутствие в России местонахождений наземных позвоночных до кунгурского периода не позволяет узнать точный момент прибытия казеидов в этот регион мира. [ 24 ] По мнению Вернебурга и его коллег, присутствие казеид на этой палеошироте предполагает их возможное существование в умеренных регионах Гондваны . [ 26 ] Возможное появление в Гондване было предложено еще в 1990-х годах Майклом С. Ю. Ли, а в 2021 году Ашером Дж. Лихтигом и Спенсером Г. Лукасом, которые переосмыслили среднепермский таксон Eunotosaurus из Южной Африки как небольшой роющий казеид. [ 35 ] [ 36 ] Между этими двумя датами другие исследователи классифицировали евнотозавра как парарептилию. [ 37 ] или на пантестудины . [ 38 ] [ 39 ] Если первые три автора правы, евнотозавр будет первым казеидом, обнаруженным в Южном полушарии , и последним известным представителем семейства самого позднего капитанского возраста для самых последних экземпляров. [ 40 ] В других частях Гондваны присутствие ихнотаксона Dimetropus в перми Марокко предполагает будущее открытие базальных синапсид (включая казеиды) в пермских бассейнах Северной Африки , которые также были частью палеоэкваториального пояса и составляли один из основных миграционных путей. между Северным и Южным полушариями. [ 41 ] [ 42 ]
Филогения
[ редактировать ]Семейство Caseidae было создано Сэмюэлем Венделлом Уиллистоном в 1911 году. [ 43 ] В 1940 году Альфред Шервуд Ромер и Л.В. Прайс поместили Caseidae и Edaphosauridae в один подотряд, который они назвали Edaphosauria. [ 44 ] В настоящее время эта группа заброшена, поскольку эдафозавриды находятся ближе к Sphenacodontia (с которой они образуют кладу Sphenacomorpha ), чем к казеидам. [ 28 ] Последние группируются с Eothyrididae в кладе Caseasauria , которые представляют собой наиболее базальные синапсиды . [ 45 ]
Первый филогенетический анализ казеидов был опубликован в 2008 году Хиллари К. Мэддин и его коллегами. В этом анализе род Oromycter занимает самое базальное положение внутри клады. Эннатозавр — сестринская группа клады, в которую входят Cotylorhynchus romeri и Angelosaurus dolani . Этот анализ впервые выявляет парафилию рода Casea представляющего , вида Casea rutena, отдельный род, который в 2011 году будет назван Euromycter . [ 9 ] [ 46 ]
Ниже представлена первая кладограмма казеидов, опубликованная Мэддином и др. в 2008 году. [ 9 ]
Другой филогенетический анализ, проведенный в 2012 году Бенсоном, включает род Trichasaurus , который занимает промежуточное положение между Casea broilii и Casea rutena . Эннатозавр идентифицирован как сестринская группа клады, включающей Angelosaurus и три вида Cotylorhynchus . В рамках последнего рода Cotylorhynchus romeri является сестринской группой двух видов C. hancocki и C. bransoni . [ 47 ]
Ниже представлена кладограмма Caseasauria, опубликованная Бенсоном в 2012 году. [ 47 ]
В 2015 году Марко Романо и Умберто Никосия опубликовали первое кладистическое исследование, включающее почти все Caseidae (за исключением Alierasaurus ronchii из Сардинии , который тогда считался слишком фрагментарным). Это исследование снова определяет род Casea как парафилетический, а вид Casea nicholsi представляет собой отдельный безымянный род. В наиболее скудном анализе Рутенозавр тесно связан с Cotylorhynchus hancocki . Эннатозавр занимает более высокое положение, чем в предыдущих анализах, и идентифицируется как более тесно связанный с Angelosaurus dolani . Однако тесная связь между A. dolani и эннатозавром может быть искажена крайней неполнотой материала по североамериканским видам. [ 16 ]
Ниже представлена кладограмма казеидов, опубликованная Романо и Никосией в 2015 году. [ 16 ]
Филогенетический анализ, опубликованный в 2016 году Нилом Броклхерстом и его коллегами, впервые включает роды Callibrachion и Datheosaurus , которые идентифицированы как базальные казеиды, занимающие промежуточное положение между Eocasea и Oromycter . [ 45 ]
Ниже представлен филогенетический анализ, опубликованный Нилом Броклхерстом и его коллегами в 2016 году. [ 45 ]
В 2017 году Романо и его коллеги опубликовали первый филогенетический анализ, включающий род Alierasaurus . Последний обнаружен как сестринский таксон рода Cotylorhynchus . [ 48 ]
Ниже представлен филогенетический анализ, опубликованный Романо и его коллегами в 2017 году. [ 48 ]
Описывая вид Martensius bromacckerensis в 2020 году, Берман и его коллеги опубликовали две кладограммы , на которых Martensius идентифицирован как один из наиболее базальных казеид и расположен чуть выше позднекаменноугольного Eocasea martini . На первой кладограмме положение остальных казеидов выяснено плохо. На второй кладограмме Casea broilii расположена над Martensius bromacckerensis , за ней следует политомия между Oromycter dolesorum , Trichasaurus texensis и кладой оставшегося казеида. Внутри этих оставшихся таксонов последовательность ветвящихся таксонов начинается с Casea nicholsi , за которым следует Euromycter rutenus , затем Ennatosaurus tecton , затем Angelosaurus romeri , затем апикальная клада трех видов Cotylorhynchus плюс Alierasaurus ronchii . В этой последней кладе Cotylorhynchus hancocki и C. bransoni являются сестринскими таксонами, и клада образует политомию с C. romeri и Alierasaurus ronchii . [ 5 ]
Ниже представлены две кладограммы казеидов, опубликованные Берманом и его коллегами в 2020 году. [ 5 ]
| Казеиды | |
| Казеиды | |
В 2022 году Вернебург и его коллеги описали род Lalieudorhynchus и опубликовали филогенетический анализ, который пришел к выводу, что роды Angelosaurus и Cotylorhynchus являются парафилетическими и представлены только типовыми видами. В этом анализе Cotylorhynchus romeri расположен чуть выше рода Angelosaurus и образует политомию с кладой, содержащей Ruthenosaurus и Caseopsis , и другой кладой, содержащей Alierasaurus , два других вида Cotylorhynchus и Lalieudorhynchus . В рамках этой последней клады Alierasaurus является сестринской группой «Cotylorhynchus» bransoni и более производной кладой, включающей Lalieudorhynchus и «Cotylorhynchus» Hancocki . [ 26 ]
Ниже представлена кладограмма, опубликованная Вернебургом и его коллегами в 2022 году. [ 26 ]
| Казеиды | |
Примечания
[ редактировать ]- ^ Диноцефалы известны во Франции только по их следам, приписываемым ихногену Brontopus . Последний также был обнаружен в формации Абрахамскраал в Южной Африке, где в изобилии находятся скелетные останки диноцефалов. В России казеид эннатозавр присутствует в мезенском комплексе, который включает также еще не описанный вид антеозаврид- диноцефалов.
- ^ Формула фаланг соответствует количеству фаланг, составляющих каждый палец кисти и стопы четвероногих . Он указан начиная с цифры I (соответствующей у мужчин большому и большому пальцу ноги) до цифры V (соответствующей мизинцу и мизинцу ноги).
- ^ В 1962 году Олсон и Баргузен сообщили о наличии у вида Cotylorhynchus bransoni стопы с формулой фаланг 2-2-2-3-2, то есть меньшей формулой, чем у типового вида C. romeri . Однако в 2015 году Романо и Никосия продемонстрировали, что третий палец стопы этого вида действительно имел три фаланги, а не две. Следовательно, фаланговая формула стопы C. bransoni была 2-2-3-3-2, как и у C. romeri .
Ссылки
[ редактировать ]- ^ Jump up to: а б Романо, М. (2017). «Масштабирование казеидных синапсид длинных костей: комбинированный морфометрический и кладистический подход». Летайя . 50 (4): 511–526. дои : 10.1111/лет.12207 .
- ^ Jump up to: а б Ангельчик, К.Д.; Каммерер, CF (2018). «Синапсиды немлекопитающих: глубокие корни генеалогического древа млекопитающих». В Захосе, FE; Ашер, Р.Дж. (ред.). Справочник по зоологии: эволюция, разнообразие и систематика млекопитающих . Берлин: де Грюйтер. стр. 117–198. ISBN 978-3-11-027590-2 .
- ^ Романо, М.; Циттон, П.; Магануко, С.; Сакки, Э.; Карателли, М.; Рончи, А.; Никосия, США (2018). «Новое открытие базальных синапсид на пермском обнажении Торре-дель-Портиччоло (Альгеро, Италия)». Геологический журнал . 54 (3): 1554–1566. дои : 10.1002/gj.3250 . S2CID 133755506 .
- ^ Jump up to: а б с д Рейс, РР; Фрёбиш, Дж. (2014). «Старейший казеидный синапсид из позднего пенсильванского периода в Канзасе и эволюция травоядных у наземных позвоночных» . ПЛОС ОДИН . 9 (4): е94518. Бибкод : 2014PLoSO...994518R . дои : 10.1371/journal.pone.0094518 . ПМЦ 3989228 . ПМИД 24739998 .
- ^ Jump up to: а б с д и Берман, Д.С.; Мэддин, ХК; Хенричи, AC; Сумида, СС; Скотт, Д.; Рейс, Р.Р. (2020). «Новый примитивный казеид (Synapsida, Caseasauria) из ранней перми Германии». Анналы музея Карнеги . 86 (1): 43–75. дои : 10.2992/007.086.0103 . S2CID 216027787 .
- ^ Кемп, Т.С. (1982). «Пеликозавры». В Кемпе, Т.С. (ред.). Млекообразные рептилии и происхождение млекопитающих . Лондон: Академическая пресса. п. 39. ИСБН 978-0124041202 .
- ^ Jump up to: а б Романо, М.; Броклхерст, Н.; Фрёбиш, Дж. (2017). «Посткраниальный скелет тектона эннатозавра (Synapsida, Caseidae)». Журнал систематической палеонтологии . 16 (13): 1097–1122. дои : 10.1080/14772019.2017.1367729 . S2CID 89922565 .
- ^ Jump up to: а б с Рейс, РР; Скотт, Д.; Модесто, СП (2022). «Краниальная анатомия казеидного синапсида Cotylorhynchus romeri , крупного наземного травоядного животного из нижней перми Оклахомы, США» . Границы в науках о Земле . 10 :1–19. дои : 10.3389/feart.2022.847560 .
- ^ Jump up to: а б с д Мэддин, ХК; Сидор, Калифорния; Рейс, Р.Р. (2008). «Краниальная анатомия тектона эннатозавров (Synapsida: Caseidae) из средней перми России и эволюционные связи Caseidae». Журнал палеонтологии позвоночных . 28 (1): 160–180. doi : 10.1671/0272-4634(2008)28[160:CAOETS]2.0.CO;2 . S2CID 44064927 .
- ^ Jump up to: а б с д и ж г час я дж к Олсон, ЕС (1968). «Семейство Caseidae». Филдиана: Геология . 17 : 225–349.
- ^ Рейс, Р. (2019). «Небольшой казеидный синапсид, Arisierpeton simplex gen. et sp. nov., из ранней перми Оклахомы, с обсуждением разнообразия синапсид в классическом местонахождении Ричардс Спур» . ПерДж . 7e6615: 1–20. дои : 10.7717/peerj.6615 . ПМК 6462398 . ПМИД 30997285 .
- ^ Jump up to: а б Сигоно-Рассел, Д.; Рассел, Д.Э. (1974). «Изучение первых казеид (Reptilia, Pelycosauria) из Западной Европы» . Вестник Национального музея естественной истории . Серия 3. 38 (230): 145–215.
- ^ Jump up to: а б с д Шпиндлер, Ф.; Фальконне, Дж.; Фрёбиш, Дж. (2015). « Каллибрахион и Датеозавр , два исторических и ранее ошибочных базальных казеазавровых синапсида из Европы» . Acta Palaeontologica Polonica . 361 (3): 597–616. дои : 10.4202/app.00221.2015 .
- ^ Стовалл, Дж.В.; Цена, ЛИ; Ромер, А.С. (1966). «Посткраниальный скелет гигантского пермского пеликозавра Cotylorhynchus romeri ». Вестник Музея сравнительной зоологии . 135 (1): 1–30.
- ^ Jump up to: а б Олсон, ЕС (1962). «Позднепермские наземные позвоночные, США и СССР». Труды Американского философского общества . Новая серия. 52 (2): 1–224. дои : 10.2307/1005904 . JSTOR 1005904 .
- ^ Jump up to: а б с Романо, М.; Никосия, У. (2015). «Кладистический анализ Caseidae (Caseasauria, Synapsida): использование метода взвешивания пробелов для включения таксонов на основе неполных экземпляров» . Палеонтология . 58 (6): 1109–1130. Бибкод : 2015Palgy..58.1109R . дои : 10.1111/пала.12197 . S2CID 86489484 .
- ^ Рончи, А.; Сакки, Э.; Романо, М.; Никосия, У. (2011). «Огромный казеидный пеликозавр из северо-западной Сардинии и его влияние на стратиграфию и палеобиогеографию европейской перми» . Acta Palaeontologica Polonica . 56 (4): 723–738. дои : 10.4202/app.2010.0087 . S2CID 55085495 .
- ^ Ганд, Г.; Гаррик, Дж.; Дематье, Г.; Элленбергер, П. (2000). «Палихнофауна четвероногих позвоночных верхней перми бассейна Лодев (Лангедок-Франция)» . Палевертебрата . 29 (1): 1–82.
- ^ Маркетти, Л.; Кляйн, Х.; Бухвиц, М.; Рончи, А.; Смит, RMH; Де Клерк, WJ; Шишио, Л.; Гроеневальд, GH (2019). «Следы пермско-триасовых позвоночных из Южной Африки: ихнотаксономия, производители и биостратиграфия в условиях двух крупных фаунистических кризисов» . Исследования Гондваны . 72 : 139–168. Бибкод : 2019GondR..72..139M . дои : 10.1016/j.gr.2019.03.009 . S2CID 133781923 .
- ^ Сильва, RC; Седор, ФА; Фернандес, ACS (2012). «Ископаемые следы поздней перми Бразилии: пример скрытого биоразнообразия». Журнал южноамериканских наук о Земле . 38 : 31–43. Бибкод : 2012JSAES..38...31D . дои : 10.1016/j.jsames.2012.05.001 .
- ^ Маркетти, Л.; Фойгт, С.; Лукас, СГ (2019). «Анатомически последовательное исследование лопингских эоловых следов Германии и Шотландии обнаруживает первые свидетельства массового вымирания в конце Гуадалупа на низких палеоширотах Пангеи». Исследования Гондваны . 73 : 32–53. Бибкод : 2019ГондР..73...32М . дои : 10.1016/j.gr.2019.03.013 . S2CID 146780505 .
- ^ Jump up to: а б с Сакки, Э.; Чифелли, Р.; Циттон, П.; Никосия, США; Романо, М. (2014). « Dimetropus osageorum n. isp. из ранней перми Оклахомы (США): след и его след». Ихнос: Международный журнал по следам растений и животных . 21 (3): 175–192. дои : 10.1080/10420940.2014.933070 . S2CID 129567990 .
- ^ Маталес-Андреу, Р.; Мухал, Э.; Галобарт, А.; Фортуни, Дж. (2021). «Понимание эволюции синапсидной локомоции на основе следов четвероногих из нижней перми Майорки (Балеарские острова, западное Средиземноморье)» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 579 : 110589. Бибкод : 2021PPP...579k0589M . дои : 10.1016/j.palaeo.2021.110589 .
- ^ Jump up to: а б с Броклхерст, Н.; Фрёбиш, Дж. (2017). «Повторное исследование загадочного русского четвероногого Phreatophasma aenigmaticum и его эволюционное значение» . Ископаемый рекорд . 20 (1): 87–93. doi : 10.5194/fr-20-87-2017 .
- ^ Jump up to: а б с Ламбертц, М.; Шелтон, CD; Шпиндлер, Ф.; Перри, Сан-Франциско (2016). «Казеевская точка эволюции гомолога диафрагмы среди самых ранних синапсидов». Анналы Нью-Йоркской академии наук . 1385 (1): 1–18. Бибкод : 2016NYASA1385....3L . дои : 10.1111/nyas.13264 . ПМИД 27859325 . S2CID 24680688 .
- ^ Jump up to: а б с д и ж г Вернебург, Р.; Шпиндлер, Ф.; Фальконне, Дж.; Стейер, Дж.-С.; Виани-Лиод, М.; Шнайдер, JW (2022). «Новый казеидный синапсид из перми (Гваделупы) бассейна Лодев (Окситания, Франция)» (PDF) . Палеовертебрата . 45(2)-e2 (2): e2. дои : 10.18563/pv.45.2.e2 . S2CID 253542331 .
- ^ Маркетти, Л.; Форте, Г.; Кустачер, Э.; ДиМишель, Вашингтон; Лукас, СГ; Роги, Г.; Жункал, Массачусетс; Харткопф-Фредер, К.; Крайнер, К.; Морелли, К.; Рончи, А. (2022). «Артинское потепление: евроамериканское изменение климата и земной биоты в ранней перми». Обзоры наук о Земле . 226 : 103922. Бибкод : 2022ESRv..22603922M . doi : 10.1016/j.earscirev.2022.103922 . S2CID 245892961 .
- ^ Jump up to: а б Шпиндлер, Ф.; Фойгт, С.; Фишер, Дж. (2019). «Edaphosauridae (Synapsida, Eupelycosauria) из Европы и их отношение к представителям Северной Америки». ПалЗ . 94 (1): 125–153. дои : 10.1007/s12542-019-00453-2 . S2CID 198140317 .
- ^ Голубев, В.К. (2000). «Фаунистические комплексы пермских наземных позвоночных Восточной Европы» (PDF) . 34 (Приложение 2): S211–S224.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Голубев В.К. (2001). «Даже стратиграфия и корреляция казанских морских отложений в стратотипической области» . Стратиграфия и геологическая корреляция . 9 (5): 454–472.
- ^ Голубев, В.К. (2015). «Диноцефалийский этап в истории пермской фауны четвероногих Восточной Европы». Палеонтологический журнал . 49 (2): 1346–1352. дои : 10.1134/S0031030115120059 . S2CID 130694755 .
- ^ Циттон, П.; Рончи, А.; Магануко, С.; Карателли, М.; Никосия, США; Сакки, Э.; Романо, М. (2019). «Первые следы четвероногих из перми Сардинии и их палеонтологическое и стратиграфическое значение». Геологический журнал . 54 (4): 2084–2098. дои : 10.1002/gj.3285 . S2CID 135216684 .
- ^ Шнайдер, Дж.В.; Лукас, СГ; Шольце, Ф.; Фойгт, С.; Маркетти, Л.; Кляйн, Х.; Оплюштил, С.; Вернебург, Р.; Голубев В.К.; Баррик, Дж. Э.; Немировска, Т.; Рончи, А.; Дэй, Миссури; Силентьев В.В.; Рёсслер, Р.; Сэйбер, Х.; Линнеманн, У.; Жаринова В.; Шен, СЗ. (2020). «Континентальная биостратиграфия позднего палеозоя – раннего мезозоя — ссылки на стандартную глобальную хроностратиграфическую шкалу». Палеомир . 29 (2): 186–238. дои : 10.1016/j.palwor.2019.09.001 . S2CID 210316208 .
- ^ Лорин, М.; Крюк, RW (2022). «Возраст самых молодых палеозойских континентальных позвоночных Северной Америки: обзор данных среднепермских групп реки Пиз (Техас) и Эль-Рино (Оклахома)» . BSGF — Бюллетень наук о Земле . 193 (10): 10. doi : 10.1051/bsgf/2022007 . S2CID 248955905 .
- ^ Ли, MSY (1995). Эволюционная морфология парейазавров (кандидатская диссертация). Кембридж: Кафедра зоологии Кембриджского университета. стр. 151–155.
- ^ Лихтиг, AJ; Лукас, СГ (2021). « Чинлечелис из верхнего триаса Нью-Мексико, США, и происхождение черепах» . Электронная палеонтология . 24 ((1): а13): 1–49. дои : 10.26879/886 . S2CID 233454789 .
- ^ Модесто, СП (2000). « Eunotosaurus africanus и гондванское происхождение анапсидных рептилий» . Палеонтология Африканская . 36 : 15–20.
- ^ Бевер, Г.С.; Лисон, ТР; Филд, диджей; Бхуллар, Б.С. (2015). «Эволюционное происхождение черепа черепахи». Природа . 525 (7568): 239–242. Бибкод : 2015Natur.525..239B . дои : 10.1038/nature14900 . ПМИД 26331544 . S2CID 4401555 .
- ^ Лисон, ТР; Рубидж, бакалавр наук; Шайер, ТМ; де Кейрос, К.; Шахнер, ER; Смит, RMH; Бота-Бринк, Дж.; Бевер, Г.С. (2016). «Ископаемое происхождение панциря черепахи» . Современная биология . 26 (14): 1887–1894. дои : 10.1016/j.cub.2016.05.020 . ПМИД 27426515 . S2CID 3935231 .
- ^ Дэй, Миссури; Смит, RMH (2020). «Биостратиграфия зоны комплекса эндотиодонов (группа Бофорта, супергруппа Кару), Южная Африка». Южноафриканский геологический журнал . 123 (2): 165–180. Бибкод : 2020SAJG..123..165D . дои : 10.25131/sajg.123.0011 . S2CID 225834576 .
- ^ Фойгт, С.; Лагнауи, А.; Хминна, А.; Сэйбер, Х.; Шнайдер, JW (2011). «Редакционные заметки по ихнофауне пермских четвероногих из бассейна Тиддас, центральное Марокко». Палеогеография, Палеоклиматология, Палеоэкология . 302 (3–4): 474–483. Бибкод : 2011PPP...302..474В . дои : 10.1016/j.palaeo.2011.02.010 .
- ^ Фойгт, С.; Сэйбер, Х.; Шнайдер, Дж.В.; Хмич, Д.; Хминна, А. (2011). «Ихнофауна четвероногих позднего карбона-ранней перми из бассейна Хенифры, Центральное Марокко». Геобиос 44 (4): 399–407. Бибкод : 2011Geobi..44..399V . дои : 10.1016/j.geobios.2010.11.008 .
- ^ Уиллистон, Юго-Запад (1911). «Пермские рептилии» . наук . 33 (851): 631–632. Бибкод : 1911Sci....33..631W . дои : 10.1126/science.33.851.631 . ПМИД 17798180 .
- ^ Ромер, А.С.; Прайс, Л.И. (1940). «Обзор пеликозаврии». Специальный доклад Геологического общества Америки . Специальные статьи Геологического общества Америки. 28 : 1–538. дои : 10.1130/SPE28-p1 .
- ^ Jump up to: а б с Броклхерст, Н.; Рейс, Р.; Фернандес, В.; Фрёбиш, Дж. (2016). «Переописание кузнецов миктерозавра, раннепермского эотиридида, и его влияние на филогению синапсидов пеликозаврового класса» . ПЛОС ОДИН . 11 (6): e0156810. Бибкод : 2016PLoSO..1156810B . дои : 10.1371/journal.pone.0156810 . ПМЦ 4917111 . ПМИД 27333277 .
- ^ Рейс, РР; Мэддин, ХК; Фрёбиш, Дж.; Фальконне, Дж. (2011). «Новый крупный казеид (Synapsida, Caseasauria) из перми Родеза (Франция), включая переоценку «Casea» rutena Sigogneau-Russell & Russell, 1974» . Георазнообразие . 33 (2): 227–246. дои : 10.5252/g2011n2a2 . S2CID 129458820 .
- ^ Jump up to: а б Бенсон, RBJ (2012). «Взаимоотношения базальных синапсид: краниальные и посткраниальные морфологические перегородки предполагают разные топологии». Журнал систематической палеонтологии . 10 (4): 601–624. дои : 10.1080/14772019.2011.631042 . S2CID 84706899 .
- ^ Jump up to: а б Романо, М.; Рончи, А.; Магануко, С.; Никосия, У. (2017). «Новый материал Alierasaurus ronchii (Synapsida, Caseidae) из перми Сардинии (Италия) и его филогенетическое сходство» . Электронная палеонтология . 20.2.26А: 1–27. дои : 10.26879/684 . hdl : 11573/1045550 . S2CID 73577681 .





