Лалиедоринх
| Лалиедоринх Временной диапазон: гваделупский
| |
|---|---|

| |
| Частичный посткраниальный скелет (слепок) Lalieudorhynchus gandi на выставке в Музее Лодева. Крестцовых позвонков здесь нет. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Сторона синапса |
| Клэйд : | † Казеазаврия |
| Семья: | † Казеиды |
| Род: | † Лалиедоринх Вернебург и др. , 2022 г. |
| Разновидность | |
Lalieudorhynchus — вымерший род синапсидов казеидных , живший в Гваделупе (= средней перми ) на территории современного юга Франции . Род известен только по виду типовому Lalieudorhynchus gandi , который был назван в 2022 году Ральфом Вернебургом, Фредериком Шпиндлером, Джоселин Фальконнет, Жаном-Себастьяном Штайером, Моник Виани-Лиод и Йоргом В. Шнайдером. Lalieudorhynchus представлен частичным посткраниальным скелетом, обнаруженным в Лодев бассейне в центральной части Эро департамента в Окситании регионе . Он принадлежит человеку длиной примерно 3,75 м (12,3 фута). Однако степень окостенения его костей указывает на то, что это был поздний юношеский возраст или еще растущая молодая взрослая особь . Основываясь на внутреннем строении его костей, описывающие авторы интерпретировали Lalieudorhynchus как полуводное животное, которое, возможно, вело образ жизни, сходный с образом жизни гиппопотама , проводя часть времени в воде, но возвращаясь на сушу за пищей, хотя идея о том, что казеиды были полуводный вариант ранее оспаривался другими авторами. Геологически это один из самых молодых известных представителей казеид. филогенетический анализ, предложенный Вернебургом и его коллегами, идентифицировал Lalieudorhynchus как производное казеида, тесно связанное с североамериканским видом «Cotylorhynchus» Hancocki . [ 1 ]
Этимология
[ редактировать ]Название рода представляет собой комбинацию слов La Lieude, расположенного недалеко от типового местонахождения, и «rhynchus», латинизированной формы греческого «rhynchos» (нос), иногда используемого в названиях казеид. Конкретный эпитет посвящен Жоржу Ганду, который десятилетиями работал над бассейном Лодев, особенно над плитой со следами Ла-Льед, а также был одним из организаторов и пропагандистов раскопок этого казеида. [ 1 ]
Описание
[ редактировать ]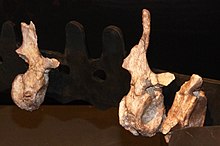
Lalieudorhynchus представлен частичным и разрозненным, но хорошо сохранившимся посткраниальным скелетом. Голотип 02–37, UM-LIE 39–41, UM - , представленный серией костей, каталогизированных UM-LIE LIE 45 и UM-LIE 47, состоит примерно из десяти позвонков ( спинных , крестцовых и хвостовых ), около пятнадцати ребер. , полный правый скапулокоракоид длиной 50 см (20 дюймов), дорсальная ветвь левой подвздошной кости , правая и левая бедренные кости длиной 35,5 см (14,0 дюймов) и несколько костей стопы ( таранная кость , два элемента предплюсны и пять фаланг ) . Общая длина тела этого экземпляра оценивается в 3,75 м (12,3 фута). Голотип Lalieudorhynchus демонстрирует смесь зрелых и неполовозрелых признаков по всему скелету, что указывает на то, что этот экземпляр был позднею ювенильной особью или все еще растущей молодой взрослой особью . на момент смерти [ 1 ] Череп неизвестен, но, как и его ближайшие родственники, он , вероятно, был очень маленьким по сравнению с размером тела, имел треугольную форму при взгляде сверху и оканчивался спереди наклонной вперед мордой с очень большими наружными ноздрями . Форма ребер указывает на то, что у Lalieudorhynchus была бочкообразная грудная клетка, как и у других производных казеидов. [ 1 ] Здесь, должно быть, располагался большой пищеварительный тракт , что позволяет предположить, что животному приходилось питаться большим количеством растений с низкой питательной ценностью. [ 2 ]
Для Lalieudorhynchus характерно несколько апоморфий . Нейральные отростки крестцовых и идущий вверх и передних хвостовых позвонков имеют на поперечном сечении очень тонкий вперед килевидный отросток, начинающийся над презигапофизами по всему вертикальному краю до вершины нервного отростка. Нервные отростки спинных и хвостовых позвонков имеют тонкий дорсальный конец, а не латеральное утолщение. Первое крестцовое ребро имеет узкий дистальный конец. Скапулокоракоид имеет ямку на трехглавом отростке метакоракоида (одна из трех костей, образующих скапулокоракоид с прокоракоидом и лопаткой ). Для стопы характерна очень крупная дистальная предплюсна 1 такой же ширины, как и таранная кость, почти со всех сторон слегка вогнутая. [ 1 ]
Lalieudorhynchus также отличается уникальным сочетанием признаков. Как и другие казеиды, он отличается от рутенозавра прямыми нервными отростками, а не наклоненными вперед. Средние хвостовые позвонки имеют относительно длинные центры , удлиненные ниже постзигапофизов , но с низкими нервными дугами, в отличие от Alierasaurus . Три крестцовых позвонка и самый передний хвостовой позвонок имеют короткий и очень широкий в поперечном направлении центр, как у Ruthenosaurus эти же центры значительно уже , тогда как у Cotylorhynchus romeri . Нервный отросток первого крестцового и первого хвостового позвонка очень удлинен дорсально, как у «Cotylorhynchus» hancocki , чего нет у C. romeri и Ruthenosaurus . Гипсфен C. присутствует под постзигапофизами дорсальных и хвостовых позвонков, особенность, ранее описанная только у Hancocki . супрагленоидное отверстие Имеется , открывающееся латерально в супрагленоидной ямке и медиально в дорсальной части подлопаточной ямки, как у "С." Ханкоки . Стержень у лопаточной лопатки значительно шире, чем Алиеразавра . Переднемедиальный край лопатки выпуклый из-за наличия слегка закругленного лопаточного отростка - признак, общий для буквы «C». Ханкоки , «С.» bransoni и «Angelosaurus» romeri , но не с C. romeri и Alierasaurus . Две части суставной ямки имеют угол примерно 130°, как у C. bransoni . Бедренная кость имеет задний мыщелок, занимающий гораздо более дистальное положение, чем передний мыщелок, в отличие от Rutenosaurus . Подколенная область бедренной кости Lalieudorhynchus относительно широкая, с мощными бороздками, значительно крупнее и глубже, чем у C. romeri . Проксимальная головка кости более массивна в дорсо-вентральном направлении, чем у C. romeri . Бедренная кость также имеет большой выраженный внутренний вертел и небольшой четвертый вертел в дистальной половине, что является общей чертой с буквой «С». hancocki , Angelosaurus romeri и A. greeni , и который отличается от C. romeri , Angelosaurus dolani , Casea broilii и Рутенозавр . Межмыщелковая ямка бедренной кости очень широкая, что предполагает узкий задне-дорсальный мыщелок, в отличие от C. romeri и Ruthenosaurus . В отличие от большинства других казеидов астрагал почти такой же ширины, как и длина, но очень похож на астрагал «C». Ханкоки . кость I плюсневая крепкая и увеличенная, как у Alierasaurus . Фаланги короткие и широкие. Они короче, чем у Алиеразавра . [ 1 ]
Палеобиология
[ редактировать ]ребер Гистология Lalieudorhynchus структурой, чрезвычайно выявила кость с очень губчатой тонкой корой и отсутствием отчетливой костномозговой полости . Эти характеристики, обнаруженные также у других крупных казеид, таких как Cotylorhynchus , позволяют предположить полуводный образ жизни. [ 3 ] [ 1 ] Однако эта гипотеза оспаривается Кеннетом Ангельчиком и Кристианом Каммерером, а также Робертом Рейсом и его коллегами на основании палеонтологических и тафономических данных в сочетании с отсутствием у этих крупных казеид морфологических адаптаций к водному образу жизни. Однако эти авторы пока не дают альтернативных объяснений внутренней структуры костей крупных казеидов. [ 4 ] [ 5 ] Вернебург и его коллеги считают, что Lalieudorhynchus и крупные казеиды в целом могли вести полуводный образ жизни, сравнимый с образом жизни гиппопотамов , проводя большую часть своего времени в воде, практикуя своего рода подводную ходьбу, а не плавание, и, возможно, возвращаясь на сушу, чтобы питаться. наземные растения. Однако палеонтологи не знают, питался ли Lalieudorhynchus наземными и/или водными растениями. Окаменелости растений, связанные со скелетом Lalieudorhynchus, идентифицированы как наземные формы, адаптированные к сухому сезонному климату, в то время как водные растения отсутствуют. Однако последние редко сохраняются в пермских стоянках. Осадочный анализ типового местонахождения Lalieudorhynchus указывает на существование нескольких потенциальных водных местообитаний. Частично алевритистые пласты аргиллита толщиной от 15 см (5,9 дюйма) до 1,8 м (5,9 футов) образовались в результате взвешенных отложений в массе стоячей воды после сильного наводнения. Некоторые остатки водных членистоногих приспособились к временному застою вод, например, conchostracans и триопсиды на этих уровнях были обнаружены . Другая возможная среда обитания - речные русла, достигающие глубины от 3,5 м (11 футов) до 5 м (16 футов) в формации Ла-Льед. Наличие корней на нескольких уровнях указывает на то, что эти каналы не могли быть заполнены текущей водой круглый год из-за сезонного климата того времени. Однако в засушливый сезон вполне вероятно, что массы стоячей воды могли существовать в заброшенных каналах, озерах или прудах, что позволяло выжить животным, ведущим более водный образ жизни, таким как тупилакозавриды- двинозавры , один экземпляр которых был связан с Скелет Lalieudorhynchus . [ 1 ]
Открытие и тафономия
[ редактировать ]Первые останки Lalieudorhynchus были обнаружены в 2001 году Йоргом В. Шнайдером и Франком Кёрнером в ручье Салагу во время геологического картирования полей в бассейне Лодев. Другие элементы были обнаружены во время раскопок, проводившихся в период с 2004 по 2008 год, всего около пятидесяти костей. Диапазон размеров костей указывает на то, что они принадлежат одному человеку. Присутствие костей одной и той же особи, спорадически распределенных в толще мощностью 1,4 м (4,6 фута) (с концентрацией костей в слое толщиной 40 см (16 дюймов)) в горизонтах с разной литологией , можно объяснить тем, что разложение туши на покрытой растительностью территории (о чем свидетельствуют многочисленные остатки растений), попеременно обнаженной и затопленной. Кости, по-видимому, подвергались переработке и переотложению несколько раз, но на очень коротком расстоянии и в течение очень короткого периода времени, поскольку кости очень хорошо сохранились. Они не изнашиваются при транспортировке по воде и не разрушаются при длительном воздействии резко сезонного климата. [ 1 ]
Географическое и стратиграфическое распространение
[ редактировать ]Голотип Lalieudorhynchus происходит из верхней части формации Ла-Льед, в Лодев бассейне , расположенном в Эро департаменте , в Окситания регионе . Разрозненный скелет был обнаружен в пластах, расположенных примерно на высоте 140 м (460 футов) над основанием формации (общая толщина которой достигает 175 м (574 фута)). . Радиометрическое датирование формации Ла-Льед отсутствует Лопинский насекомых, радиометрического возраста и скоростей седиментации , возраст (= верхняя пермь) был присвоен ему на основе биостратиграфии рассчитанных для подстилающей формации Салагу. Магнитостратиграфические и палеонтологические данные, скорее всего, предполагают гвадалупский (= среднепермский) возраст. Магнитостратиграфия показывает, что нижняя часть формации Ла Льед не моложе, чем инверсия Иллаварра, глобальное геомагнитное событие, датируемое средним вордианом 266,66 ± 0,76 миллиона лет назад. Следовательно, эта часть формации, вероятно, будет иметь поздний роудский – ранний вордский возраст, тогда как верхняя часть будет иметь как минимум поздний – ранний вордский возраст. Капитанский век. Присутствие в нижней части формации Ла-Льед ихногена Brontopus также указывает на гваделупский возраст. Первоначально обнаруженный в Ла-Льеде, бронтопус с тех пор был обнаружен в формации Абрахамскраал в Южной Африке , которая радиометрически датирована вордианским и капитанским периодами. Предполагаемые производители следов Brontopus , диноцефаловые терапсиды , также соответствуют гвадалупскому возрасту формации Ла-Льед, поскольку кости этих животных, обнаруженные в Южной и Восточной Африке , а также в России , Китае и Бразилии , известны исключительно в гвадалупских отложениях. [ 1 ] Вордийско-капитанский возраст Lalieudorhynchus делает его одним из последних известных казеид. Что касается родов Ennatosaurus и Alierasaurus , это подтверждает сохранение казеид в период Гваделупы, по крайней мере, в Европе. [ 1 ]
Палеосреды
[ редактировать ]

В гвадалупское время большая часть суши была объединена в один суперконтинент — Пангею . Оно имело примерно С-образную форму: его северная ( Лавразия ) и южная ( Гондвана ) части были соединены с западом, но разделены с востока очень большим морем Тетис . [ 6 ] Длинная цепочка микроконтинентов, сгруппированных под названием Киммерия , делила Тетис на две части: Палео-Тетис на севере и Нео-Тетис на юге. [ 7 ] Бассейн Лодеве располагался в экваториальном поясе того времени, на уровне 10-й параллели северной широты , и по отношению к берегам Тетиса находился примерно в 400 км вглубь суши. [ 8 ] [ 9 ] Герцинские горы неизвестной топографии отделяли бассейн Лодеве от Тефии. [ 9 ] В это время очень влажный климат, обычно характеризующий экваториальный климат, сменился распространением более сухого тропического климата (с двумя сезонами — сухим и влажным) в районы, близкие к экватору. [ 8 ] [ 10 ]
Формация Ла-Льед представлена конгломератами мелко- и крупнозернистых, частично галечников песчаников , красно-коричневых, частично глинистых или мелкопесчаных алевролитов и прослаивающихся алевритистых аргиллитов . Эти породы соответствуют пескам , гравию и гальке переносимым реками , а также речным илам из пойм , отложившимся в разветвленной речной системе. Есть также несколько слоев селей, свидетельствующих об очень сильных дождях во время сезона дождей. Большая часть костей Lalieudorhynchus происходит из этих горизонтов селей, а часть - из селевых потоков. Эти отложения селей также содержат обильные растительные остатки, представленные многочисленными ланцетными листьями длиной от 5 до 11 см (2,0–4,3 дюйма), вероятно, принадлежащими Plagiozamites , некоторые остатки Podozamites -подобных хвойных фитофитов, Supaia -подобные фрагменты, несколько стволов деревьев до 2 м (6,6 футов) в длину и от 15 до 20 см (от 5,9 до 7,9 дюймов) в ширину, а оси растений меньшего размера. [ 1 ]
Помимо Lalieudorhynchus , в верхней части формации Lieude обнаружены только позвоночник тупилакозаврида , пресноводные членистоногие (конхостраканы и триопсиды), крылья темноспондил насекомых ( Odonata ) и неопределенные следы ног. [ 1 ] Другие элементы фауны формации Ла-Льед присутствуют на плите Ла-Льед, расположенной ближе к основанию формации, где обнажаются многочисленные четвероногих следы . Многие ихновиды четвероногих [ номер 1 ] были названы с этого сайта. Таксономический диноцефалических и морфологический обзор этих следов выделил четыре действительных ихновида и определил их вероятных производителей: Brontopus giganteus и B. antecursor , который, весьма вероятно, представляет собой терапсидов ( соответственно Tapinocephalia и Anteosauridae ), Merifontichnus thalerius , который соответствует следам морадизаврина. captorhinid eureptile и Pachypes ollieri , которые принадлежат к парейазавроморфе Nycteroleteridae . [ 11 ] [ 12 ] [ 13 ]
Филогения
[ редактировать ]Филогенетический анализ, проведенный Вернебагом и его коллегами, определил Lalieudorhynchus gandi как один из наиболее производных казеидов и родственный таксон североамериканскому виду «Cotylorhynchus» Hancocki . Эти два таксона образуют кладу, характеризующуюся наличием гипосфена , а также наличием и положением супрагленоидного отверстия. Эта клада вместе с "Cotylorhynchus" bransoni образует апикальную кладу, характеризующуюся близко расположенными постзигапофизами. Этот анализ также предполагает, что роды Angelosaurus и Cotylorhynchus (каждый из которых состоит из трех видов) будут парафилетическими , а таксоны, отличные от их типовых видов, могут принадлежать к разным родам . [ 1 ]
Ниже представлена кладограмма, опубликованная Вернебургом и его коллегами в 2022 году.
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Один и тот же ихновид мог быть оставлен несколькими близкородственными биологическими видами, имеющими очень похожую анатомию кисти и стопы.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н Вернебург, Р.; Шпиндлер, Ф.; Фальконне, Дж.; Стейер, Дж.-С.; Виани-Лиод, М.; Шнайдер, JW (2022). «Новый казеидный синапсид из перми (Гваделупы) бассейна Лодев (Окситания, Франция)» (PDF) . Палеовертебрата . 45 (45(2)-e2): e2. дои : 10.18563/pv.45.2.e2 . S2CID 253542331 .
- ^ Олсон, ЕС (1968). «Семейство Caseidae». Филдиана: Геология . 17 : 225–349.
- ^ Ламбертц, М.; Шелтон, CD; Шпиндлер, Ф.; Перри, Сан-Франциско (2016). «Казеевская точка эволюции гомолога диафрагмы среди самых ранних синапсидов». Анналы Нью-Йоркской академии наук . 1385 (1): 1–18. Бибкод : 2016NYASA1385....3L . дои : 10.1111/nyas.13264 . ПМИД 27859325 . S2CID 24680688 .
- ^ Ангельчик, К.Д.; Каммерер, CF (2018). «Синапсиды немлекопитающих: глубокие корни генеалогического древа млекопитающих». В Захосе, FE; Ашер, Р.Дж. (ред.). Справочник по зоологии: эволюция, разнообразие и систематика млекопитающих . Берлин: де Грюйтер. стр. 138–139. ISBN 978-3-11-027590-2 .
- ^ Рейс, РР; Скотт, Д.; Модесто, СП (2022). «Краниальная анатомия казеидного синапсида Cotylorhynchus romeri , крупного наземного травоядного животного из нижней перми Оклахомы, США» . Границы в науках о Земле . 10 :1–19. дои : 10.3389/feart.2022.847560 .
- ^ Маклафлин, С. (2001). «История распада Гондваны и ее влияние на докайнозойский флористический провинциализм». Австралийский журнал ботаники . 49 (3): 271–300. дои : 10.1071/BT00023 .
- ^ Шенгёр, AMC (1987). «Тектоника Тетисидов: развитие орогенных коллажей в условиях коллизий». Ежегодный обзор наук о Земле и планетах . 15 : 214–244. Бибкод : 1987AREPS..15..213C . doi : 10.1146/annurev.ea.15.050187.001241 .
- ^ Перейти обратно: а б Шнайдер, Дж.В.; Кернер, Ф.; Рошер, М.; Кронер, У. (2006). «Развитие пермского климата в северной зоне пери-Тетиса - бассейн Лодев, Центральный французский массив, в сравнении в европейском и глобальном контексте». Палеогеография, Палеоклиматология, Палеоэкология . 240 (1–2): 161–183. Бибкод : 2006PPP...240..161S . дои : 10.1016/j.palaeo.2006.03.057 .
- ^ Перейти обратно: а б Мишель, Луизиана; Табор, Нью-Джерси; Монтаньес, IP; Шмитц, М.; Давыдов, В.И. (2015). «Хроностратиграфия и палеоклиматология бассейна Лодев, Франция: свидетельства пантропической аридификации на границе каменноугольного и пермского периодов» . Палеогеография, Палеоклиматология, Палеоэкология . 430 : 118–131. Бибкод : 2015PPP...430..118M . дои : 10.1016/j.palaeo.2015.03.020 .
- ^ Кемп, Т.С. (2006). «Происхождение и раннее распространение терапсидных млекопитающих рептилий: палеобиологическая гипотеза» . Журнал эволюционной биологии . 19 (4): 1231–1247. дои : 10.1111/j.1420-9101.2005.01076.x . ПМИД 16780524 . S2CID 3184629 .
- ^ Маркетти, Л.; Кляйн, Х.; Бухвиц, М.; Рончи, А.; Смит, RMH; Де Клерк, WJ; Шишио, Л.; Гроеневальд, GH (2019). «Следы пермско-триасовых позвоночных из Южной Африки: ихнотаксономия, производители и биостратиграфия в условиях двух крупных фаунистических кризисов» . Исследования Гондваны . 72 (72): 139–168. Бибкод : 2019GondR..72..139M . дои : 10.1016/j.gr.2019.03.009 . S2CID 133781923 .
- ^ Маркетти, Л. (2016). «Новые появления четвероногих ихнотаксонов из пермского бассейна Оробик (Северная Италия) и критическое обсуждение возраста ихноассоциации». Статьи по палеонтологии . 2 (3): 363–386. дои : 10.1002/spp2.1045 . S2CID 133136159 .
- ^ Маркетти, Л.; Фойгт, С.; Мухал, Э.; Лукас, СГ; Францискини, Х.; Фортуни, Дж.; Сантуччи, В.Л. (2021). «Распространение следов Pareiasauromorpha на Приуралье: более раннее появление и более широкая палеобиогеография группы» (PDF) . Статьи по палеонтологии . 7 (3): 1297–1319. дои : 10.1002/spp2.1342 . S2CID 229416421 .