Мосхоринус
| Мосхоринус Временной диапазон: поздняя пермь – ранний триас
| |
|---|---|
| |
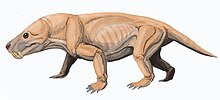
| Реставрация Moschorhinus kitchingi | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Сторона синапса |
| Клэйд : | Терапсида |
| Клэйд : | † Тероцефалия |
| Семья: | † Акидногнатиды |
| Род: | † Мосхоринус Метла , 1920 год. |
| Разновидность: | † М. Китчинги
|
| Биномиальное имя | |
| † Moschorhinus kitchingi Метла, 1920 год.
| |
| Синонимы | |
| |
Moschorhinus — вымерший род тероцефаловых , синапсидов семейства Akidnognathidae M. содержащий только один вид: kitchingi , который был обнаружен в поздней перми — раннем триасе южноафриканской супергруппы Кару . Это был крупный плотоядный терапсид , достигавший 1,1–1,5 метра (3,6–4,9 футов) в общей длине тела с самым большим черепом, сравнимым по размеру с черепом льва , и имел широкую тупую морду с длинными прямыми клыками .
Moschorhinus , по-видимому, экологически заменил горгонопсидов в качестве высшего хищника и охотился так же, как большая кошка . Хотя он был наиболее многочисленным в поздней перми, он в небольших количествах дожил до раннего триаса после пермского вымирания , хотя у этих выживших в триасе видов наблюдалась задержка роста .
Таксономия
[ редактировать ]Название рода Moschorhinus происходит от древнегреческих слов μόσχος (мос-хос) moschos, обозначающих теленка или молодого животного, и rhin/rhino-, обозначающего нос или морду, в связи с его короткой и широкой мордой. Название вида, kitchingi , относится к г-ну Джеймсу Китчингу, который первоначально нашел (но не описал) этот экземпляр. [ 2 ]
Китчинг обнаружил экземпляр голотипа — череп (лучше всего сохранившийся — нёбо ) — в супергруппе Кару в Южной Африке, недалеко от деревни Нью-Бетезда . Впервые он был описан палеонтологом Робертом Брумом в 1920 году. [ 2 ] Сейчас это один из самых известных и узнаваемых терапсидов супергруппы. [ 3 ]
Брум назвал другой вид тероцефалов Ранее в 1907 году из Квазулу-Натала , Scymnosaurus warreni , который он позже перенес в Moschorhinus в 1932 году как M. warreni , сохранив его как отдельный вид. M. warreni Позднее Китчинг в своей неопубликованной докторской диссертации признал вероятным синонимом M. kitchingi , а повторное описание голотипа в 2023 году Дэвидом Грюнвальдом и Кристианом Каммерером подтвердило это предложение. Как старое название, M. warreni будет иметь таксономический приоритет для перед M. kitchingi этого вида . Однако Гроеневальд и Каммерер (2023) полагали, что было бы преждевременно определять M. warreni как правильное название в ожидании пересмотра таксономии тероцефаловых акидногнатид и возможности того, что даже более старые названия могут иметь старшинство. [ 1 ]

Останки Moschorhinus были обнаружены наиболее заметно в от верхней перми до нижнего триаса группе Бофорта . [ 3 ] [ 4 ] [ 5 ]
Классификация
[ редактировать ]Moschorhinus — тероцефал , член клады Eutheriodontia и родственный таксон цинодонтам и современным млекопитающим. Moschorhinus относится к семейству Akidnognathidae , наряду с другими крупными плотоядными терапсидами с крепким черепом и большими верхними клыками. [ 6 ]
Moschorhinus занял нишу , когда-то контролируемую горгонопсидами . Обе группы были построены как большие кошки . После исчезновения Moschorhinus в триасовом периоде цинодонты . аналогичную нишу заняли [ 6 ]
Описание
[ редактировать ]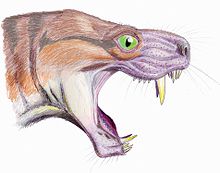
Moschorhinus был крупным хищником, достигавшим общей длины тела 1,1–1,5 метра (3,6–4,9 футов) и весом 84,3 кг (186 фунтов). [ 7 ] [ 8 ] Череп похож на череп горгонопсидов, с большими височными окнами (всего три в виде синапсид) и выпукло изогнутым небом . Размер черепа варьировался от размера варана до льва. У них характерно короткая и широкая морда. У них есть пара заметно длинных резцов , похожих на клыки саблезубых кошек . [ 6 ] [ 9 ]
Вид сбоку челюсти Moschorhinus , показывающий диапазон движений, необходимый для таких больших резцов, и верхние небные отверстия морды. (Из Ван Валкенбурга и Дженкинса, 2002). [ 6 ]
Морда
[ редактировать ]Морда Moschorhinus характерно короткая и широкая. На тупом кончике морды имеется гребень, идущий по средней линии к лобной кости . [ 6 ] [ 4 ] Нижняя челюсть значительно шире, чем у любого другого тероцефала. [ 6 ] [ 4 ] У молодых особей верхняя часть морды немного выступает за резцы. [ 4 ]
Ноздри были большими и располагались ближе к кончику морды. [ 4 ]
Зубы
[ редактировать ]Считается, что у Moschorhinus была зубная формула I6.C1.M3 с 6 резцами , 1 клыком и 3 задниклыками по обе стороны верхней челюсти. [ 2 ]
Резцы расположены в предчелюстных костях . Они большие, слегка изогнутые, имеют колоколообразную форму в поперечном сечении. У них были гладкие режущие поверхности и, в отличие от других тероцефалов, не было граней или бороздок, возникающих в результате истирания и износа. [ 4 ]
Большие саблевидные клыки находятся внутри верхних челюстей и являются быстро распознаваемыми признаками Moschorhinus . Они особенно толстые и прочные и имеют уникальное круглое поперечное сечение. По длине эти сабли сравнимы с горгонопсидами. Хотя реального современного аналога не существует, наиболее похожим живым примером может быть дымчатый леопард ( Neofelis nebulosa ). [ 6 ]
Как и у других тероцефалов, у Moschorhinus было меньше постклыков, расположенных на верхних челюстях. У большинства тероцефалов «зубы», или, вернее, зубчатые выступы (зубцы) крыловидных костей сильно редуцированы или отсутствуют, а у Moschorhinus они отсутствуют. [ 4 ] [ 10 ]
Черепная крыша
[ редактировать ]Прослеживая крышу черепа, Moschorhinus имеет небольшие префронтальные кости над глазами, за которыми следуют большие расширенные лобные кости. образуют Теменные кости узкий сагиттальный гребень вдоль средней линии черепа, в котором находится очень простое шишковидное отверстие . [ 2 ] [ 4 ] можно увидеть углубления В височных ямках , углубления по обе стороны от гребня, что указывает на наличие множества кровеносных сосудов и нервов, снабжающих мозг. [ 11 ]
Глазницы
[ редактировать ]Слезная кость крупнее редуцированной префронтальной кости и образует большую часть глазницы . Слёзная кость имеет костный выступ (округленный выступ) на глазнице и большое отверстие на внутренней стороне. Нижний край глазницы образован скуловой и верхнечелюстной костями. [ 2 ] Скуловая кость заканчивается у глазницы и не выпуклая, как у некоторых более поздних тероцефалов. [ 4 ]
Нёбо
[ редактировать ]В целом небо выпуклое, с широким треугольным сошником, с парными бугорками, округлыми выступами, направленными вентрально, [ 6 ] [ 4 ] сходен с другими акидногнатидами. [ 2 ] Небные кости (образующие заднюю часть неба) увеличены и толсты, особенно на их внешних краях, где они соединяются с верхней челюстью. По своим внутренним краям небные кости соединены с крыловидной костью и сошником на носу, образуя часть окружности полости носа . Между небом и верхней челюстью, сразу за клыками, имеются большие отверстия, предположительно предназначенные для прохождения нервов. Наклонный гребень посередине неба предположительно поддерживал мягкое небо, которое позволяло воздуху проходить между носом и легкими. [ 6 ]
Для использования сабель требуется широко открыть рот, что затрудняет кормление. У близкородственного Promoschorhynchus имеются жесткие складки ( хоанальный гребень) на границе носового прохода и горла, которые используются для того, чтобы держать его открытым и обеспечивать дыхание во время еды. Развитие вторичного неба в черепе постепенно развилось у тероцефалов, а хоанальный гребень присущ всем более поздним тероцефалам. [ 10 ]
Палеобиология
[ редактировать ]
Предполагается, что Moschorhinus был кошачьим хищником, способным протыкать кожу и удерживать борющуюся добычу своими длинными клыками. Это первая запись о подобной охотничьей технике. Учитывая его прочную конструкцию, толстую морду, огромные клыки и мощные челюстные мышцы, Moschorhinus , по-видимому, был устрашающим хищником. [ 6 ]
Палеоэкология
[ редактировать ]В бассейне Кару было обнаружено множество окаменелостей позвоночных. Другими тероцефалами с того же уровня породы являются Tetracynodon и Promoschorhynchus . [ 3 ] Экземпляры Moschorhinus были единственными крупными тероцефалами. [ 12 ] [ 4 ]
Moschorhinus , по-видимому, вымер в раннем триасе на 251 млн лет назад после пермского вымирания на 252 млн лет назад. [ 13 ] [ 14 ] [ 15 ] наряду с 80–95% видов животных из-за массовой гипоксии . Судя по всему, это привело к задержке роста . [ 3 ] интенсивные сезоны, сокращение разнообразия экосистем и потеря лесов. [ 4 ] Ископаемые данные показывают, что триасовый Moschorhinus рос быстрее, чем пермский, что привело к уменьшению размера тела у первых, что, как полагают, является следствием более резкой изменчивости окружающей среды после пермского вымирания ( эффект лилипутии ). [ 3 ] [ 16 ] [ 4 ] Черепа пермского периода имеют среднюю длину 207 мм (8,1 дюйма), а длина черепа триасового периода - всего 179 мм (7,0 дюйма). [ 3 ] Тем не менее, триасовые Moschorhinus были крупнейшими тероцефалами своего времени. [ 3 ] [ 12 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б Гроеневальд, ДП; Каммерер, CF (2023). «Повторная идентификация и обновленный стратиграфический контекст голотипов позднепермских четвероногих Dicynodon ingens и Scymnosaurus warreni из Квазулу-Натал». Палеонтология Африканская . 56 : 171–179. hdl : 10539/37143 .
- ^ Jump up to: а б с д и ж Веник Р. (1920). «О некоторых новых тероцефаловых рептилиях из слоев Карру в Южной Африке» . Труды Лондонского зоологического общества : 351–354.
- ^ Jump up to: а б с д и ж г Хуттенлокер А.К., Бота-Бринк Дж. (2013). «Размер тела и модели роста тероцефалов Moschorhinus kitchingi (Eutheriodontia) до и после вымирания в конце пермского периода в Южной Африке». Палеобиология . 39 (2): 253–77. дои : 10.1666/12020 . S2CID 86490421 .
- ^ Jump up to: а б с д и ж г час я дж к л м Хаттенлокер, Адам (2013). Палеобиология южноафриканских тероцефаловых терапсидов (Amniota, Synapsida) и влияние вымирания в конце пермского периода на размер, рост и микроструктуру костей (доктор философии). Университет Вашингтона.
- ^ Рубидж, бакалавр наук; Сидор, Калифорния (2001). «Эволюционные закономерности пермо-триасовых терапсидов». Ежегодный обзор экологии и систематики . 32 : 449–480. doi : 10.1146/annurev.ecolsys.32.081501.114113 .
- ^ Jump up to: а б с д и ж г час я дж ван Валкенбург Б., Дженкинс I (2002). «Эволюционные закономерности в истории пермо-триасовых и кайнозойских синапсидных хищников» (PDF) . Документы Палеонтологического общества . 8 : 267–88. дои : 10.1017/S1089332600001121 . Архивировано из оригинала (PDF) 17 октября 2013 г.
- ^ Бота, Дж. и Смит, RMH (2005). « Видовой состав Lystrosaurus на границе пермо-триаса в бассейне Кару в Южной Африке». Летайя . 40 (2): 125–137. дои : 10.1111/j.1502-3931.2007.00011.x . Полная версия онлайн по адресу « Видовой состав листрозавров на границе пермо-триаса в бассейне Кару в Южной Африке» (PDF) . Архивировано из оригинала (PDF) 10 сентября 2008 г. Проверено 2 июля 2008 г.
- ^ Стюарт, БП; Хаттенлокер, АК; Бота, Дж. (2024). «Посткраниальная анатомия Moschorhinus kitchingi (Therapsida: Therocephalia) из бассейна Кару в Южной Африке» . ПерДж . 12 . е17765. дои : 10.7717/peerj.17765 . ПМЦ 11326434 . ПМИД 39148680 .
- ^ Хаттенлокер Адам (2009). «Исследование кладистических отношений и монофилии тероцефаловых терапсидов» (PDF) . Зоологический журнал Линнеевского общества . 157 (4): 865–891. дои : 10.1111/j.1096-3642.2009.00538.x . S2CID 84603632 . Архивировано (PDF) из оригинала 19 марта 2023 г.
- ^ Jump up to: а б Майер В., ван ден Хевер Дж., Дюран Ф. (1996). «Новые экземпляры терапсидов и происхождение вторичного твердого и мягкого неба млекопитающих» . Журнал зоологической систематики и эволюционных исследований . 34 : 9–19. дои : 10.1111/j.1439-0469.1996.tb00805.x .
- ^ Дюран Ж. Ф. (1991). «Пересмотренное описание черепа moschorhinus (therapsida, therocephalia)» . Анналы Южноафриканского музея . 99 : 381–413.
- ^ Jump up to: а б Кристиан А. Сидор; Роджер М.Х. Смит; Адам К. Хаттенлокер; Брэндон Р. Пикук (2014). «Новые четвероногие среднего триаса из формации Верхнего Фремоу в Антарктиде и условия их отложения» . Журнал палеонтологии позвоночных . 34 (4): 793–801. дои : 10.1080/02724634.2014.837472 . S2CID 128981733 .
- ^ Питер Д. Уорд; Дженнифер Бота; Роджер Бьюик; Мишель О. Де Кок; Дуглас Х. Эрвин; Джеффри Х. Гаррисон; Джозеф Л. Киршвинк; Роджер Смит (2005). «Резкое и постепенное вымирание наземных позвоночных поздней перми в бассейне Кару, Южная Африка». Наука . 307 (5710): 709–714. Бибкод : 2005Sci...307..709W . CiteSeerX 10.1.1.503.2065 . дои : 10.1126/science.1107068 . ПМИД 15661973 . S2CID 46198018 .
- ^ Бота Дж., Смит Р.М. (2006). «Быстрое восстановление позвоночных в бассейне Кару в Южной Африке после вымирания в конце пермского периода» (PDF) . Журнал африканских наук о Земле . 45 (4–5): 502–14. doi : 10.1016/j.jafrearsci.2006.04.006 . [ мертвая ссылка ]
- ^ Дамиани, Р.; Модесто, С.; Йейтс, А.; Невелинг, Дж. (2003). «Самые ранние свидетельства рытья нор цинодонтами» . Труды Лондонского королевского общества Б. 270 (1525): 1747–1751. дои : 10.1098/rspb.2003.2427 . JSTOR 3592240 . ПМК 1691433 . ПМИД 12965004 .
- ^ Ричард Дж. Твитчетт (2007). «Эффект лилипутии после вымирания в конце Пермского периода» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 252 (1–2): 132–144. дои : 10.1016/j.palaeo.2006.11.038 .


