2024 год в палеонтологии архозавров
| |||
|---|---|---|---|
| +... |
В этой статье зафиксированы новые таксоны всех видов ископаемых архозавров которых запланировано , описание на 2024 год, а также другие важные открытия и события, связанные с палеонтологией архозавров, которые будут опубликованы в 2024 году.
Псевдозухи
[ редактировать ]Новые таксоны псевдозухий
[ редактировать ]| Имя | Новинка | Статус | Авторы | Возраст | Тип населенного пункта | Страна | Примечания | Изображения |
|---|---|---|---|---|---|---|---|---|
СП. ноябрь | В печати | Мартинс и др. | Поздний мел | Баурусухид . Анонсировано в 2023 году; окончательная версия статьи была опубликована в 2024 году. | ||||
СП. ноябрь | Действительный | Фернандес Дюмон и др. | Поздний мел (сеноман) | |||||
СП. ноябрь | Нарваес и др. | Эоцен (Лютет) | Базальный представитель Crocodyloidea . | |||||
| Бенггвишингасух [4] | ген. это сп. ноябрь | Действительный | Смит и др. | Средний триас (анизийский) | Формация Фавре | ( | Представитель Paracrocodylomorpha , вероятно, принадлежащий к группе Poposauroidea . Типовой вид — B. eremicarminis . |  |
СП. ноябрь | Иори и др. | Поздний мел | ||||||
ген. это сп. ноябрь | Действительный | Сакс и др. | Ранний мел (валанжин) | Метриоринхид . Типовой вид — E. schroederi . | ||||
ген. это сп. ноябрь | Действительный | Рейес, Марц и Смолл | Поздний триас (норийский) | Этозавр . Типовой вид — G. muelleri . | ||||
ген. это сп. ноябрь | Действительный | Лопес-Рохас и др. | Поздняя юра (кимеридж-титон) | Гониофолидид крокодиломорф . Типовой вид — O. paimogonectes . | ||||
| Парвозух [9] | ген. это сп. ноябрь | Мюллер | Триас (ладинский-карнийский период) | Последовательность Пиньейрос-Чиникуа суперпоследовательности Санта-Мария | псевдозухий Грацилизухидный . Типовой вид — P. aurelioi. |  | ||
ген. и расчесать. ноябрь | Десохо и Раухут | Триас (ладинский-карнийский период) | Последовательность Пиньейрос-Чиникуа суперпоследовательности Санта-Мария | Представитель Paracrocodylomorpha, вероятно, принадлежащий к группе Poposauroidea. Типовой вид — Prestosurus loricatus von Huene (1938). | ||||
Джин и др. ноябрь | В печати | Почат-Котийу и др. | Ранний мел | Атопозаврид . Типовой вид — V. sakonnakhonensis . |
Общее исследование псевдозухий
[ редактировать ]- Сенников (2024) трактует орнитозухид как хищников-макрофагов со специализированным челюстным аппаратом и отмечает аналогии между ними и саблезубыми терапсидами (в том числе млекопитающими). [12]
- Исследование передвижения Riojasuchus tenuisceps опубликовано фон Бачко и др. (2024), которые реконструируют R. tenuisceps как человека с прямой осанкой и парасагиттальной походкой, но не пришли к окончательному выводу, был ли он двуногим или четвероногим. [13]
- Исследование анатомии черепа и неврологии Tarjadia ruthae опубликовано Desojo et al. (2024). [14]
- Исследование гистологии плечевой кости Benggwigwishingasurus eremacarminis опубликовано Кляйном (2024), который не обнаружил никаких доказательств вторичной водной адаптации, но сообщает о доказательствах, указывающих на более медленную скорость роста, чем у Effigia и Sillosuchus . [15]
- Переописание анатомии скелета Shuvosaurus inexpectatus опубликовано Nesbitt & Chatterjee (2024). [16]
- Мастрантонио и др. (2024) описывают анатомию посткраниального скелета наиболее полного экземпляра Prestosuchus chiniquensis, зарегистрированного на сегодняшний день, и пересматривают диагноз P. chiniquensis . [17]
Исследование этозавра
[ редактировать ]Исследование крокодиломорфа
[ редактировать ]- Описание анатомии и гистологии костей посткраниального скелета Terrestrisuchus gracilis опубликовано Spiekman, Butler & Maidment . [18]
- Вудворд и др. (2024) отмечают корреляцию между объемом бедра аллигатора и массой тела и используют объем бедра для определения массы тела гониофолидид , дирозавров , нотозухов и талаттозухий . [19]
- Исследование морфологического разнообразия тазового пояса талаттозухий и дирозавридов на протяжении всей их эволюционной истории опубликовано Scavezzoni et al. (2024). [20]
- Янг и др. (2024) обеспечивают систематизацию Thalattosuria более высокого уровня как в соответствии с Филокодом , так и в Международном кодексе зоологической номенклатуры , называя новые таксоны Neothalattosuria , Euthalattosuria и Dakosaurina . [21]
- Исследование морфологии остеодерм Indosinosuchus из и неназванного представителя Mesoeucrocodylia позднеюрского места раскопок Фу Ной ( Таиланд ) опубликовано Bhuttarach et al. (2024). [22]
- Веринский и др. (2024) идентифицировали рострум телеозавра из известняковой формации Ченстохова Губка ( Польша ) как принадлежащий немахимозавру махимозавриду , питающемуся крупной добычей, с морфологическим сходством с Neosteneosaurus edwardsi и Proexochokefalos heberti , предоставляя доказательства того, что такие телеозавроиды присутствовали за пределами Западной Европы. во время Оксфорда . [23]
- Шейер и др. (2024) описывают зубные коронки телеозавроидов, связанные с останками ихтиозавров (со следами мусора, также оставленными телеозавроидами) из байосской формации Хаутрогенштайн ( Швейцария ), представляющие собой самый старый ископаемый материал члена трибы Machimosaurini, о котором сообщалось на сегодняшний день. [24]
- Хуа, Листон и Табуэль (2024) описывают экземпляр Metriorhynchus cf. superciliosus из келловейских слоев скал «Ваш-Нуар» в Виллер-сюр-Мер ( Франция ), сохранившихся с содержимым желудка, включающим остатки жаберного аппарата Leedsichthys , и интерпретируют изученный образец как свидетельство того, что Metriorhynchus питался останками. из Лидсихтиса . [25]
- Исследование гистологии костей Araripesuchus buitreraensis , доказывающее в целом медленную, ежегодно прерываемую скорость роста, опубликовано Navarro et al. (2024). [26]
- Доказательства непрерывной и скоординированной замены зубов у Armadillosuchus arrudai , гарантирующей, что животное не потеряет слишком много зубов одновременно и что потеря зубов не повлияет на его способность к питанию, представлены Borsoni, Carvalho & Marinho (2024). [27]
- Дос Сантос и др. (2024) описывают анатомию скелета наиболее полного юного экземпляра баурузухид , зарегистрированного на сегодняшний день, и сообщают о доказательствах различий в украшении черепа и развитии мышц между молодыми и взрослыми экземплярами баурузухид, которые могут указывать на онтогенетическое разделение ниш . [28]
- Ископаемый материал гониофолидида , интерпретируемый как базальная из нижнемеловой формации Китадани ( Япония ). форма, которая имела несколько общих анатомических черт с производными членами группы, описан Обусе и Сибатой (2024) [29]
- Форе и др. (2024) изучают факторы, стимулирующие эволюцию тетисухий , сообщая о доказательствах смены видов после пограничного события сеномана и турона , когда фауна с доминированием дирозаврид заменила фауну с доминированием фолидозавридов , увеличения биоразнообразия тетисухий после события вымирания в мел-палеогеновом периоде и положительного результата. корреляция между длиной тела и температурой. [30]
- Кузьмин и др. (2024) представляют реконструкцию Kansajsuchus extensus и отмечают наличие значительных различий в строении черепной коробки фолидозавридов и дирозаврид, ставя под сомнение близкое родство этих двух групп. [31]
- Рокки и Вила (2024) описывают ископаемый материал Allodaposuchus cf. subjuniperus из нижнемаастрихтских отложений местонахождения Сутерранья-Мина-де-лигнит ( формация Ла-Поса ; Лерида, Испания ), что свидетельствует о присутствии третьего раннемаастрихтского вида Allodaposuchus (помимо A. palustris и A. hulki ) в Группа Тремп. [32]
- Йейтс и Штейн (2024) интерпретируют Ultrastenos willisi и «Baru» Huberi как синонимы , но сохраняют Ultrastenos как отдельный род мекозухин , в результате чего образуется новая комбинация Ultrastenos Huberi . [33]
- Обзор летописи окаменелостей и осморегуляции представителей Alligatoroidea опубликован Стаутом (2024), который утверждает, что ископаемые представители этой группы могли быть солеустойчивыми и более приспособленными к плаванию в океане, чем современные аллигатороиды. [34]
- Переописание Arambourgia gaudryi опубликовано Conedera et al. (2024), которые обнаружили A. gaudryi как аллигатора и интерпретировали его как полуземное животное. [35]
- Переописание Crocodylus palaeindicus и исследование филогенетических взаимоотношений представителей Crocodyloidea опубликовано Chabrol et al. (2024), которые считают Crocodylus sivalensis младшим синонимом C. palaeindicus , находят доказательства тесного родства Crocodylus checchiai и Crocodylus falconensis с современными американскими крокодилами, обнаруживают киньян как крокодилин, а не остеолемин , обнаруживают Albertosuchus knudsenii , Prodiplocynodon langi. и "Crocodylus" affinis за пределами Crocodyloidea, и считают, что размещение аллигатороидов для клады Orientalosuchina очень лабильно. [36]
Нептичьи динозавры
[ редактировать ]Новые таксоны динозавров
[ редактировать ]| Имя | Новинка | Статус | Авторы | Возраст | Тип населенного пункта | Страна | Примечания | Изображения |
|---|---|---|---|---|---|---|---|---|
ген. это сп. ноябрь | Действительный | Чжэн и др. | Поздний мел ( Маастрихт ) | Теропод -тираннозавр . Типовой вид — A. xui . |  | |||
ген. это сп. ноябрь | Действительный | Нин и др. | Средняя юра (бат) |  | ||||
ген. это сп. ноябрь | Альварес Ногейра и др. | Поздний мел (сеноман-турон) | Эласмариевый орнитопод . Типовой вид — C. nekul . |  | ||||
| Комптонированный [40] | ген. это сп. ноябрь | Локвуд и др. | Ранний мел ( баррем ) | Формирование Уэссекса | Игуанодонтид - орнитопод. Типовой вид — C. Chasei . |  | ||
ген. это сп. ноябрь | Действительный | Син и др. | Поздний мел (турон-ранний коньяк) | Анкилозаврид . Типовой вид — D. yingliangis . |  | |||
ген. это сп. ноябрь | Действительный | Порфири и др . | Поздний мел (сантон) | теропод Неэнлагийный . Типовой вид — D. lechiguanae . | 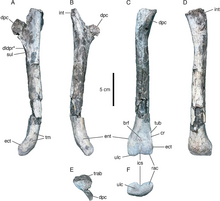 | |||
ген. это сп. ноябрь | Действительный | барон | Ранняя юра (геттанг-синемюр) | Аверострановый . теропод Типовой вид — D.normani . |  | |||
ген. это сп. ноябрь | Аткинс-Вельтман и др. | Поздний мел (Маастрихт) | Ценагнатидный . теропод Типовой вид — E. infernalis . |  | ||||
| Фон [45] | ген. это сп. ноябрь | Авраами и др. | Поздний мел (сеноман) | Кедровая горная формация | ( | Тецелозаврид птицетазовый . Типовой вид — F. herzogae. |  | |
ген. это сп. ноябрь | Действительный | Хан и др. | Поздний мел (сеноман-турон) | Чжутянская формация | Титанозавр-зауропод. Типовой вид — G. cavocaudatus . |  | ||
ген. это сп. ноябрь | Ли и др. | Поздний мел | Троодонтидный . теропод Типовой вид — H. prima . |  | ||||
ген. это сп. ноябрь | Действительный | Ротатори и др. | Поздняя юра | Ранний дивергентный игуанодонтический орнитопод, возможно, дриоморфан . Типовой вид — H. martinhotomasorum . |  | |||
ген. это сп. ноябрь | Действительный | Кубота, Кобаяши и Икеда | Ранний мел (альб) | Троодонтидный . теропод Типовой вид — H. matsubaraetoheorum . |  | |||
ген. это сп. ноябрь | Действительный | Филиппи и др. | Поздний мел (сантон) | Титанозавр-зауропод. Типовой вид — I. oslatus . Анонсировано в 2023 году; окончательная версия статьи была опубликована в 2024 году. |  | |||
ген. это сп. ноябрь | Рен и др. | Поздняя юра | маменчизаврид Зауропод . Типовой вид — J. dongxingensis . Первоначально предложенное название принадлежит Джингиа Чен, 1983 год. [52] Заменяющее название было опубликовано в приложении. [53] |  | ||||
ген. это сп. ноябрь | Аверьянов и др. | Ранний мел (апт) | Ноазавридный . теропод Типовой вид — K. longipes . |
| ||||
ген. это сп. ноябрь | Pol et al. | Поздний мел (кампан-маастрихт) | Абелизавридный . теропод Типовой вид — К. неточность. |  | ||||
ген. это сп. ноябрь | Действительный | Лоуэн и др. | Поздний мел (кампан) | Центрозавровый . цератопс Типовой вид – L. rangiformis. |
| |||
ген. это сп. ноябрь | Лонгрич и др. | Поздний мел (Маастрихт) | Ламбеозавровый принадлежащий гадрозаврид, к трибе Arenysaurini . Типовой вид - M. bata. |  | ||||
| Не стесняйся [58] | ген. это сп. ноябрь | Барретт и др. | Поздний триас ( норийский ) | Галечная аркосовая свита | Массоподан - зауроподоморф. Типовой вид — M. sanyatiensis . |  | ||
ген. это сп. ноябрь | Исасменди и др. | Ранний мел (баррем-апт) | Спинозавридный . теропод Типовой вид — R. lacustris . |  | ||||
ген. это сп. ноябрь | Действительный | Лерзо и др. | Поздний мел (сеноман-турон) | зауропод Реббахизаврид- . Типовой вид — S. marae . |  | |||
ген. это сп. ноябрь | Зафати и др. | Средняя юра | Стегозавр . Типовой вид — T. atlasicus. |  | ||||
ген. это сп. ноябрь | Перейра и др. | Мел (альб–сеноман) | Базальный зауропод - титанозавр. Типовой вид — T. valdecii . |  | ||||
| Тиетазаура [63] | ген. это сп. ноябрь | Бандейра и др. | Ранний мел ( валанжин - готерив ) | Формация Слоновой Кости | Эласмариевый орнитопод. Типовой вид — T. derbyiana . |  | ||
ген. это сп. ноябрь | Перес-Морено и др. | Поздний мел (кампан-маастрихт) | Титанозавр-зауропод. Типовой вид — T. gimenezi . |  | ||||
СП. ноябрь | Действительный | Далман и др. | Поздний мел (кампан-маастрихт) | Тираннозавр ; разновидность тираннозавра . |
| |||
ген. это сп. ноябрь | В печати | Сото и др. | Поздний мел | Титанозавр-зауропод, принадлежащий к отряду Saltasauroidea . Типовой вид — U. celeste . |  | |||
СП. ноябрь | Ван и др. | Поздний мел | Троодонтидный теропод ; разновидность урбакодона . | |||||
ген. это сп. ноябрь | Действительный | Лонгрич и др. | Ранний мел (баррем) | Гипсилофодонтид . Типовой вид — V. insularis. Анонсировано в 2023 году; окончательная версия статьи была опубликована в 2024 году. |
| |||
ген. это сп. ноябрь | Действительный | Цзя и др. | Ранний мел (альб) | Стегозавр. Типовой вид — Y. ultimus . |  |




Общее исследование нептичьих динозавров
[ редактировать ]- Обзор исследований филогенетических взаимоотношений основных групп динозавров за предыдущие годы опубликован Lovegrove, Upchurch & Barrett (2024). [70]
- Доказательства, указывающие на то, что эволюция рострального кератинового покрова была связана с частичной редукцией зубов на протяжении всей эволюционной истории динозавров, но не объясняют полную потерю зубов в линиях динозавров, представлены Aguilar-Pedrayes, Gardner & Organ (2024). [71]
- Исследование скорости эволюции механики укуса травоядных динозавров опубликовано Кунцем и Сакамото (2024), которые интерпретируют свои результаты как указание на то, что скорость биомеханической эволюции может выявить экологические признаки на разных линиях и стадиях онтогенеза. [72]
- Каспар и др. (2024) представляют пересмотренные оценки энцефализации и числа телэнцефальных нейронов у динозавров, оспаривая оценки количества нейронов и относительного размера мозга, представленные в исследовании Herculano-Housel (2023), [73] и, в частности, противоречивые оценки исключительного количества нейронов и относительного размера мозга у крупных теропод по сравнению с другими динозаврами, представленные цитируемым автором. [74]
- Исследование эволюции ландшафта климатических ниш динозавров на протяжении мезозоя опубликовано Chiarenza et al. (2024), которые сообщают, что распространение зауроподоморфов указывает на их предпочтение теплой среды, в то время как птицетазовые и тероподы исследовали более широкий диапазон сред с разнообразным климатом, и интерпретируют колонизацию тероподами областей с более холодным климатом начиная с ранней юры как вероятно связанную к развитию эндотермии. [75]
- Предполагаемые фрагменты костей крупнотелых динозавров из ретских отложений во Франции , Германии и Великобритании как ископаемый материал крупнотелых ихтиозавров . были переосмыслены Перилло и Сандером (2024) [76]
- Ромилио и др . (2024) описывают следы динозавров из раннеюрских ( синемюрских ) слоев Razorback (Австралия), представляющие собой самые старые на сегодняшний день следы динозавров в стране. [77]
- Трояно и др. (2024) сообщают об открытии ассоциации следов динозавров раннего мела и петроглифов на стоянке Серроте-ду-Летрейру ( Бразилия ). [78]
- Обзор окаменелостей динозавров позднего триаса и юры из Индии опубликован Хосла и Лукас (2024). [79]
- Мейдмент (2024) описывает разнообразие динозавров из верхней части формации Моррисон (США) во времени и пространстве и находит доказательства, подтверждающие кладогенез как средство увеличения разнообразия диплодоцинов с течением времени, а также пространственной сегрегации видов аллозавров и камаразавров . [80]
- Бандейра и др. (2024) пересматривают останки динозавров из групп нижнего мела Массакара и Ильяс (бассейн Реконкаво, Бразилия ), собранные между 1859 и 1906 годами, и интерпретируют изученные окаменелости как указывающие на присутствие группы динозавров раннего мела, включая теропод, зауроподов и орнитопод. [63]
- Киркланд и др . (2024) описывают биоразнообразие меловых динозавров из штата Юта (США). [81]
- Исследование диверсификации нептичьих динозавров, сделанное на основе имеющихся филогений динозавров, опубликовано Allen et al. считают невозможным сделать окончательный вывод о том, произошло ли сокращение разнообразия динозавров до мел-палеогенового вымирания (2024), которые на основании имеющихся данных , отмечая влияние использованных в исследовании филодинамических моделей (в частности, их предположений о выборке и изменения числа видов во времени) на оценках темпов эволюции динозавров. [82]
Сауришские исследования
[ редактировать ]- Исследование бедренной гистологии амниот из триасовой формации Ишигуаласто ( Аргентина ), включая ранних динозавров Chromogisaurus novasi , Eodromaeus murphi , Eoraptor lunensis , Herrerasaurus ischigualastensis и Sanjuansaurus gordilloi , опубликовано Curry Rogers et al. (2024), которые обнаружили, что ранние динозавры, известные из этой формации, росли по крайней мере так же быстро, как динозавры-зауроподоморфы и тероподы из позднего мезозоя, и что их повышенные темпы роста не отличали их от других амниот, живших в то же время. [83]
- Юань и др. (2024) описывают новые следы зауроподов и теропод из верхнеюрско-нижнемеловой формации Хоучэн (Хэбэй, Китай) и интерпретируют изученные следы как свидетельствующие о последовательной эволюции биоты Янляо и биоты Джехол , без каких-либо доказательств полной смена или вымирание биоты, а также предположение о том, что разнообразие динозавров в Северном Китае в раннем меловом периоде находилось под влиянием вулканической активности. [84]
- Пайо и др. (2024) описывают новый фрагмент ребра кампан-маастрихтского возраста из формации Марилия ( Бразилия ) и интерпретируют его как представляющий неопределенный ящеротаз . [85]
Исследования тероподов
[ редактировать ]- Исследование вариаций формы бедренных костей у теропод, доказывающее эволюцию подобных адаптаций к гигантизму у крупных теропод независимо от их филогенетического сходства, опубликовано Pintore et al. (2024). [86]
- Дриди и др. (2024) описывают следы теропод среднего и крупного размера из слоев нижнего мела ( готерив - баррем ) из местонахождения Джебель-Кебар (формация Бухедма, Тунис ), расширяя известный географический ареал нептичьих теропод до более высоких широт в пределах Гондваны . [87]
- Исследование сходства сброшенных зубных коронок теропод из туронско - коньякской формации Портесуэло (Аргентина), доказывающее ранее недокументированное разнообразие теропод из этой формации, опубликовано Meso et al. (2024). [88]
- Исасменди и др. (2024) описывают новые и пересматривают известные зубы теропод из маастрихтских слоев Южно-Пиренейского бассейна ( Испания ), расширяя известное разнообразие теропод из этого бассейна и сообщая о доказательствах оборота теропод в течение маастрихта. [89]
- McLarty & Esperante (2024) описывают следы теропод из маастрихтских отложений на территории следа Каррерас Пампа ( Боливия ), интерпретируемые как вероятные свидетельства того, что следостроители останавливались во время движения, обходили препятствия и приседали. [90]
- Мохабей и др. (2024) рассмотрели и переописали Laevisuchus indicus , Jubbulpuria tenuis и Compsosurus Solus , а также описали новую зубную кость ноазаврид из центральной Индии с расположенным рядом зубным рядом, похожим на тот, что присутствует у Masiakasaurus . [91]
- Исследование сходства изолированных зубов теропод из группы Кем-Кем ( Марокко ) опубликовано Hendrickx et al. (2024), которые идентифицировали зубы абелизавридов , спинозавринов , кархародонтозаврид и неабелизавроидного цератозавра или мегарапторана . [92]
- Вероятная зубная кость цератозаврид описана (2024) из тоарской формации Каньядон-Асфальто ( Аргентина Праделли, Полом и Эскуррой ) , что расширяет известное разнообразие теропод из этой формации. [93]
- Исследование сходства изолированных зубов теропод из бассейна Бауру ( Бразилия ) опубликовано Delcourt et al. (2024), которые утверждают, что на географическое распространение абелизаврид в Южной Америке повлияли климатические условия. [94]
- Рибейро и др. ) верхней юры и нижнего мела (2024) идентифицируют зуб теропода из формации Миссан-Велья ( Бразилия как самую старую на сегодняшний день находку абелизаврид в Южной Америке. [95]
- Исследование гистологии костей абелизаврида среднего размера из верхнемеловой формации Серра-да-Гальга ( Бразилия ) опубликовано Aureliano et al. (2024), которые сообщают, что, несмотря на проживание в полузасушливой тропической среде, изученный экземпляр имел скорость роста, аналогичную темпам роста патагонских абелизаврид. [96]
- Исследование скелетных патологий, поражающих известные экземпляры брахиространовых абелизаврид, опубликовано Baiano et al. (2024), которые диагностируют сращение двух хвостовых позвонков голотипа Aucasaurus garridoi как врожденный порок развития , а частичное сращение пяти хвостовых позвонков голотипа Elemgasem nubilus - как спондилораптропатию, что в обоих случаях представляет собой первые проявления диагностированной патологии. среди нетентануарных теропод . [97]
- Черрони, Отеро и Новас (2024) представляют реконструкцию мускулатуры таза и задних конечностей Skorpiovenator bustingorryi . [98]
- Исследование микроархитектуры костей осевого скелета Majungasaurus Rahonavis и et , доказывающее увеличение пневматической сложности у ранних паравианов по сравнению с представителями Ceratosauria, опубликовано Aureliano al. (2024). [99]
- Кау (2024) интерпретирует экземпляры « комсогнатидных » теропод как молодых представителей неманирапториформных групп столбняков . [100]
- Монтеалегре, Кастильо-Виза и Селлес (2024) описывают ранее неопубликованный ископаемый материал теропод ( ср. Protathlitis и кархародонтозаврид, который может отличаться от Concavenator ) из барремской формации Arcillas de Morella ( Испания ). [101]
- Юн (2024) выявляет конвергентные сходства в краниодентальной анатомии спинозавров и фитозавров . [102]
- Мирвольд и др . (2024) используют статистический анализ, чтобы пересмотреть предыдущие описания Fabbri et al . (2022) спинозавров, таких как Spinosaurus, как подводных собирателей, [103] и предоставить доказательства того, что спинозавр, вероятно, не был водным хищником. [104]
- Данные изучения особенностей формы черепа, которые интерпретируются как указывающие на то, что спинозавр питался водной добычей и, вероятно, использовал стратегию хищничества «стой и жди», представлены Smart & Sakamoto (2024). [105]
- Баффетаут и Тонг (2024) дали новую интерпретацию предполагаемого зуба ихтиозавра из формации Сан-Кхуа, собранного в 1962 году и описанного в 1963 году как зуб спинозаврид, и стали первой находкой окаменелости нептичьего динозавра, зарегистрированной в Таиланде . [106]
- Роу и Рэйфилд (2024) изучают биомеханические возможности черепов тираннозавроидных теропод с разными размерами тела и морфологией черепа и обнаруживают, что более крупные тираннозавроиды испытывали более высокие абсолютные напряжения в черепах во время кормления по сравнению с их мелкотелыми родственниками, и что широкие черепа тираннозаврид позволили им лучше переносить высокие стрессы во время кормления. [107]
- Исследование схемы замены зубов Guanlong wucaii опубликовано Ke, Pei & Xu (2024). [108]
- Зубы вероятного базального тираннозавроида описаны из верхнеюрской формации Фу Крадунг ( Таиланд ) Чоучувечом и др. (2024). [109]
- Син и др. (2024) описывают большие зубы тираннозавроидов из маастрихтской формации Даланшань , что представляет собой самую южную на сегодняшний день находку тираннозавроидов в Китае. [110]
- ЛеБлан и др. (2024) сообщают, что дошедшие до нас вараны Комодо сохраняют режущие края своих зубов благодаря обогащенному железом покрытию на зазубринах и кончиках зубов, утверждают, что секвестрация железа, вероятно, широко распространена в эмали рептилий, но также не находят доказательств наличия железных покрытий вдоль зубцов зубов тероподовых динозавров. , сообщают, что тираннозавриды имели специализированную волнистую эмаль вдоль зубчатых зазубрин, которая, вероятно, поддерживала режущие края зубов, и интерпретируют эти результаты либо как указание на разные стратегии питания тираннозаврид и варанов Комодо, либо как указание на то, что только у крупных теропод зубная эмаль имела такую структуру. был достаточно толстым, чтобы существенно влиять на механический износ зубьев зубьев. [111]
- Словяк, Брусатте и Щигельски (2024) переоценивают ископаемый материал, приписываемый Bagaraatan ostromi , интерпретируя голотип как неопределенный молодой тираннозаврид и сообщая, что некоторые из окаменелостей, первоначально приписываемых B. ostromi, на самом деле представляют собой кости ценугнатидов. [112]
- Юн (2024) оценивает профили силы нижней челюсти у Alioramus altai и Qianzhousaurus sinensis , интерпретируя челюсти изученных теропод как вероятно неподходящие для нанесения мощных укусов и выдерживания высоких нагрузок, вызванных захватом, удержанием и расчленением крупной добычи. [113]
- Исследование родства тираннозавров опубликовано Уоршоу, Баррера Гевара и Фаулер (2024), которые оспаривают выводы исследования Шерера и Войкулеску-Холвада (2023). [114] восстановление признанных видов дасплетозавров как представляющих единую анагенетическую линию, предковую для тираннозавров - линии тираннозавров. [115]
- Лонгрич и Саитта (2024) рассматривают таксономический статус Nanotyrannus и утверждают, что многочисленные доказательства подтверждают, что он является отдельным таксоном с небольшим телом, возможно, не принадлежащим к тираннозаврам , а не незрелой формой тираннозавра . [116]
- Мэллон и Хоун (2024) подсчитали, что прошлые усилия по отбору проб, вероятно, привели к выборке даже 99-го процентиля массы тела, достигнутой тираннозавром рексом , и что самые крупные представители этого вида могли быть на 70% крупнее, чем самые крупные известные в настоящее время экземпляры. , достигая примерно 15 000 (± 3750) кг массы тела. [117]
- Исследование филогенетических взаимоотношений Kinnareemimus khonkaenensis опубликовано Самати (2024). [118]
- Описание анатомии скелета Nothronychus graffami и N. mckinleyi , свидетельствующее о наличии черт, сходных с современными птицами, птицетазовыми динозаврами и зауроподами-титанозаврами, опубликовано Smith & Gillette (2024). [119]
- Парк и др. (2024) предполагают, что ранние пеннарапторы могли использовать свои перистые перья , чтобы спугивать прячущихся насекомых, а также создавать подъемную силу или сопротивление во время преследования покрасневших насекомых, и предполагают, что такое использование перистых перьев могло способствовать эволюции более крупных и жестких перьев. перья. [120]
- Характеристика того, как количество и форма маховых перьев коррелирует со стилем передвижения у современных птиц, опубликована Киатом и О'Коннором (2024). Экстраполируя эти закономерности на мезозойских пеннарапторов , авторы предполагают, что каудиптерикс и анхиорнитины, возможно, были вторично нелетающими . [121]
- Исследование эволюции грудного пояса пеннарапторанов опубликовано Wu et al. (2024), которые сообщают о доказательствах модификаций, изменяющих диапазон движений передних конечностей, которые предшествовали началу полета у паравианов , а также о доказательствах последующих адаптивных к полету модификаций у авиаланов . [122]
- Мид и др. (2024) сообщают о доказательствах, указывающих на то, что способность черепа противостоять большим механическим нагрузкам появилась на ранних этапах эволюции овирапторозавров , до появления сильно модифицированной черепной архитектуры овирапторидов . [123]
- Первый ископаемый материал ценагнатид из верхнего кампанского слоя Дена-зин формации Киртланд ( Нью-Мексико , США ) описан Funston, Williamson & Brusatte (2024). [124]
- Цю и др. (2024) описывают скелетную анатомию запястья Heyuannia huangi , предоставляя доказательства специализированной морфологии запястья, которая функционально сходилась с морфологией запястья современных птиц. [125]
- Описание анатомии скелета оксоко аварсана опубликовано Funston (2024). [126]
- Чжу, Ван и Ван (2024) изучают микроструктурные вариации яиц элонгатоолитид из Китая и интерпретируют изученные вариации как указывающие на то, что не все яичные скорлупы элонгатоолитид могут быть связаны с овирапторозаврами. [127]
- Исследование формы черепа и механики укуса дромеозаврид опубликовано Це, Миллером и Питтманом (2024), которые интерпретируют Deinonychus antirhopus как приспособленного к поимке крупной позвоночной добычи, а Halszkaraptor escuilliei считают , что он вряд ли питается рыбой и с большей вероятностью будет питаться рыбой. имеют экологию питания, аналогичную таковой у современных водоплавающих птиц. [128]
- Возможные яйца дромеозаврид описаны из верхнемеловой формации Лианхэ (Китай) Wu et al. (2024), которые назвали новый оотаксон Gannanoolithus yingliangi и интерпретировали открытие парных яиц Gannanoolithus как возможное свидетельство того, что у дромеозаврид были парные функциональные яйцеводы . [129]
- Джанечини, Колли и Маковицки (2024) представляют реконструкцию мускулатуры таза и задних конечностей Buitreraptor gonzalezorum . [130]
- позы суставов стопы дейнониха во время его цикла ходьбы . Манафзаде, Гейтси и Бхуллар (2024) на основе сравнения с современными птицами реконструировали [131]
- Описание черепной коробки и черепной эндокасты Sinovenator changii , интерпретируемого как морфологически промежуточное между базальными тероподами и современными птицами, опубликовано Yu et al. (2024). [132]
- Син и др. (2024) описывают следы из верхнемеловой формации Шаксиан (Фуцзянь, Китай), которые могли быть оставлены крупнотелым (оценочная высота бедра более 1,8 м) троодонтидом , и называют новый ихнотаксон Fujianipus yingliangi . [133]
Исследование зауроподоморфов
[ редактировать ]- Сильва и др. (2024) описывают ископаемый материал члена или родственника группы Bagualosauria из стоянки Вила-Ботукарай (последовательность Канделария суперпоследовательности Санта-Мария, Бразилия ), представляющей собой первого зауроподоморфа, зарегистрированного с этого участка. [134]
- Доказательства изменчивости структуры пневмонии шейных и спинных позвонков у платеозавра представлены Регаладо Фернандесом (2024). [135]
- Переописание голотипа и исследование родства Plateosaurus trossingensis опубликовано Шеффером (2024). [136]
- Шеффер и др. (2024) описывают патологии шевронов хвоста у двух экземпляров Plateosaurus trossingensis из местонахождения Обере Мюле в Троссингене ( Германия ), сообщают о патологиях хвостовых шевронов у других образцов, указывающих на то, что шевроны были уязвимой частью хвоста, и интерпретируют пострадавшие люди могут выздороветь без особых осложнений, если не будет нанесен серьезный функциональный ущерб. [137]
- Чжао и др . (2024) описывают новый экземпляр ювенильно-половозрелых массоспондилид из нижнеюрской формации Луфэн (Юньнань, Китай), увеличивая известное разнообразие массоспондилид из Азии. [138]
- «Gyposaurus» sinensis интерпретируется как вероятный младший синоним Lufengosaurus huenei . Ван, Чжао и Ю (2024) [139]
- Barrett & Choiniere (2024) переописывают анатомию скелета Melanorosaurus readi и определяют лектотип этого вида. [140]
- На примере спинофорозавра Видал (2024) объясняет, как виртуальные 3D-модели зауроподов позволили понять их биомеханику. [141]
- Агусти, Алькала и Сантос-Кубедо (2024) предполагают, что гигантизм зауроподов был адаптацией, которая увеличила способность зауроподов преодолевать большие расстояния, что было вызвано выраженными сезонными изменениями. [142]
- Батлер и др. (2024) описывают совокупность следов, оставленных крупными зауроподами, проходящими через прибрежную лагуну из самых ранних меловых слоев формации Дарлстон (Дорсет, Великобритания ), представляющих собой крупнейшее место следов динозавров, доступное в пределах группы Пурбек, о котором сообщалось на сегодняшний день. [143]
- Буасверт и др. (2024) описывают новый экземпляр Haplocanthosaurus sp. из карьера динозавров Сухой Мезы ( Колорадо , США ), что расширяет известный ареал этого рода до настоящей части бассейна Браши формации Моррисон и, вероятно, представляет собой геологически самое молодое местонахождение гаплокантозавра на плато Колорадо. [144]
- Кинг и др. (2024) сообщают о доказательствах ранее неизвестной формы пневматическости в ребре представителя рода Apatosaurus и предполагают, что пневматическость ребер среди апатозавринов индивидуально варьируется. [145]
- Виндхольц и др. (2024) описывают новый хвостовой позвонок реббахизаврид из сеноманской формации Канделерос ( Аргентина ), предоставляя новую информацию о хвостовой анатомии и пневматическости реббахизаврид. [146]
- Гомес и др. (2024) описывают новые титанозавроформ окаменелости из формации Портесуэло (Аргентина), расширяя известное разнообразие зауроподов из этой формации. [147]
- Бистон и др. (2024) описывают новый материал зауроподов из формации Винтон ( Австралия ) и интерпретируют Australotitan cooperensis как неопределенного диамантиназавра , который, вероятно, является младшим синонимом Diamantinasaurus matildae . [148]
- Филиппи и др. (2024) изучают ископаемый материал зауроподов из района Серро Оверо — Ла Инвернада ( формация Бахо-де-ла-Карпа ; провинция Неукен , Аргентина ), интерпретируемый как свидетельствующий о наличии разнообразной фауны титанозаврообразных, сосуществовавших в окружающей среде в течение сантона . [149]
- Исследование тафономии ископаемого материала Кайютитан-мауи и гистологии его костей опубликовано Филиппи, Превитера и Гарридо (2024). [150]
- Видал и др. (2024) изучают диапазон движений осевого ряда Trigonosaurus Pricei и интерпретируют его как способность к высокому поднятию шеи. [151]
- Исследование степени посткраниальной пневматическости у сальтазавринов и других производных титанозавров опубликовано Зурриагузом (2024). [152]
- Описание и исследование морфологической изменчивости аппендикулярных останков зауропод из маастрихтских стоянок бассейнов Хацег, Трансильвании и Руска Монтана ( Румыния ) опубликовано Мочо, Пересом-Гарсиа и Кодреа (2024), которые интерпретируют изученные останки как свидетельства присутствие четырех или пяти таксонов зауроподов на острове Хацег в течение маастрихта, включая линию титанозавров с чрезвычайно удлиненной кистью . [153]
- Обзор крупнейших известных зауроподов Аргентины опубликован Кальво (2024). [154]
Орнитишские исследования
[ редактировать ]- Исследование филогенетических взаимоотношений птицетазовых опубликовано Fonseca et al. (2024), которые назвали новые клады Pyrodontia и Tenontosauridae . [155]
- Исследование таксономического сходства изолированных птицетазовых зубов из батских стоянок микропозвоночных в Соединенном Королевстве , доказывающее наличие ранее неизвестной разнообразной птицетазовой фауны, опубликовано Wills, Underwood & Barrett (2024). [156]
- Исследование структуры смены зубов у Jeholosaurus shangyuanensis , доказывающее, что скорость смены зубов замедлялась в процессе онтогенеза, опубликовано Hu et al. (2024). [157]
- Переописание анатомии скелета и исследование сходства Oryctodromeus Cubularis опубликованы Krumenacker et al. (2024). [158]
- Остеологический и филогенетический анализ Ajkaceratops kozmai , предполагающий, что первоначальная классификация этого вида как цератопса как неопределенная и, таким образом, рассматриваемая как загадочное птицетазовое животное, был опубликован Чепинским и Мадзией (2024). [159]
- Lee et al., (2024) описали однопедальную фалангу базального неорнитисха (NHCG 10972) из нижнемеловых отложений Тандо Южной Кореи, наиболее сходного с Jeholosauridae . [160]
исследование тиреофора
[ редактировать ]- Сатчелл (2024) повторно идентифицировал фрагмент проксимального отдела бедренной кости (BELUM K3998) из группы Лиас в Северной Ирландии как неопределенный остаток динозавра, а не как потенциальный образец сцелидозавра или птицетазового животного. [161]
- Кастанера, Мампель и Кобос (2024) описывают новые следы стегозавров из верхнеюрской формации Виллар-дель-Арсобиспо ( Испания ), предоставляя доказательства стадного поведения стегозавров. [162]
- Санчес-Феноллоса, Эскасо и Кобос (2024) описывают новый экземпляр Dacentrurus Armatus из верхнеюрской формации Виллар-дель-Арсобиспо ( Испания ), предлагают новый диагноз этого вида и интерпретируют Miragaia longicollum как младший синоним D. Armatus . [163]
- Латегано, Конти и Лозар (2024) изучают стрессоустойчивость хвоста Miragaia longicollum , интерпретируют его хвост как способный развивать высокую скорость и давление, но также интерпретируют его хвостовые шипы как менее прочные, чем у Stegosaurus stenops , и рассматривают свои результаты. чтобы свидетельствовать об оборонительной стратегии, в которой запугивание отдавалось приоритету, а не прямому физическому бою. [164]
- Первый ископаемый материал стегозавров из Ганьсу ( Китай ), отнесенный к Stegosaurus sp., описан из группы Hekou нижнего мела Li et al. (2024). [165]
- Кросс и Арбор (2024) описывают бедренную кость анкилозавра из сеноманской формации Данвеган ( Британская Колумбия , Канада). [166]
- Сото Акунья, Варгас и Калуза (2024) переописывают голотип Antarctopelta из формации острова Сноу-Хилл (Антарктида) и подтверждают его филогенетическое положение внутри Parankylosauria . [167]
Исследование цераподов
[ редактировать ]- Обзор раннемеловых испанских стиракостернов из бассейна Маэстрата, опубликованный Сантос-Кубедо (2024). [168]
- Эсканеро-Агилар и др. (2024) описывают материал черепа гадрозавриформного орнитопода из нижнемеловой формации Кастрильо-де-ла-Рейна ( Испания ), который интерпретируется как более производный, чем игуанодон , но более базальный , чем Проа , и расширяет известное разнообразие орнитопод из бассейна Камерос. [169]
- Хаяши и др. (2024) сообщают об открытии вероятного гадрозавроидного позвонка из верхнемеловой формации Хикета (группа Идзуми) в Сануки, префектура Кагава, что представляет собой дополнительные доказательства расселения гадрозавроформ в районе современной Японии в кампанском периоде . [170]
- Николов, Дочев и Брусатте (2024) проверяют онтогенетический возраст небольших костей гадрозавроидов из позднемеловой ( маастрихтской ) формации Кайлака (Болгария) и определяют, что этот экземпляр, вероятно, принадлежал позднему молодому или молодому полувзрослому человеку, а не карлику взрослому . и предполагают, что крупные наземные животные смогли заселить некоторые европейские острова циклическим или кратковременным путем расселения. [171]
- Van der Linden et al. (2024) describe spheroolithid eggshells from the Maastrichtian Argiles et Grès à Reptiles Formation, probably representing the first hadrosauroid eggshells reported from France, and name a new ootaxon Paraspheroolithus porcarboris.[172]
- The first described hadrosaurid footprints from the Horseshoe Canyon Formation are described by Powers et al. (2024), who assign them to the ichnospecies Hadrosauropodus langstoni.[173]
- Evidence from the study of a skull of a juvenile hadrosaurine from the Campanian Dinosaur Park Formation (Alberta, Canada), interpreted as indicative of differences in the dental battery development between hadrosaurid species which might have been related to dietary differences during early ontogeny, is presented by Warnock-Juteau et al. (2024).[174]
- Sharpe et al. (2024) describe fossil material of a probable immature specimen of Edmontosaurus regalis from the Horseshoe Canyon Formation, and interpret its similarities to Ugrunaaluk kuukpikensis as supporting the referral of the Alaskan saurolophine material to Edmontosaurus cf. regalis.[175]
- Description of the morphology of the skull and endocranium of Psittacosaurus sibiricus, based on the study of both juvenile and adult specimens, is published by Podlesnov et al. (2024).[176]
- A description endocranial anatomy of the Psittacosaurus lujiatunensis published by Sakagami et al. (2024).[177]
- Yang et al. (2024) describe a well-preserved scaled skin of a specimen of Psittacosaurus from the Early Cretaceous Jehol Biota of China, providing evidence of preservation of epidermal layers, corneocytes and melanosomes, and interpret the studied specimen as indicative of co-occurrence of feathers and reptile-type skin in non-feathered regions of the skin in Psittacosaurus.[178]
- Witton & Hing (2024) argue that there is no compelling evidence indicating that the development of the idea of the griffin was inspired by the discovery of fossils of Protoceratops.[179]
- Barrera Guevara et al. (2024) reinterpret fossil material of Coahuilaceratops magnacuerna as derived from Cerro Huerta Formation (and representing the first dinosaur taxon described from this formation) rather than from Cerro del Pueblo Formation.[180]
Birds
[edit]New bird taxa
[edit]| Name | Novelty | Status | Authors | Age | Type locality | Country | Notes | Images |
|---|---|---|---|---|---|---|---|---|
Sp. nov | Valid | Tennyson et al. | A species of Ardenna. | |||||
Sp. nov | Valid | Agnolín, Álvarez Herrera & Tomassini | Pleistocene | A species of Chloephaga. | ||||
Gen. et sp. et comb. nov | Valid | Zelenkov | Pliocene and Pleistocene | Crimea | A relative of the grey partridge. The type species is E. voinstvenskyi; genus also includes "Phasianus" etuliensis Bocheński & Kurochkin (1987) from Moldova. | |||
| Eocypselus geminus[184] | Sp. nov | In press | Mayr & Kitchener | Eocene | London Clay | A species of Eocypselus. | ||
| Eocypselus grandissimus[184] | Sp. nov | In press | Mayr & Kitchener | Eocene | London Clay | A species of Eocypselus. | ||
| Eocypselus paulomajor[184] | Sp. nov | In press | Mayr & Kitchener | Eocene | London Clay | A species of Eocypselus. | ||
| Fluvioviridavis michaeldanielsi[185] | Sp. nov | Mayr & Kitchener | Eocene | London Clay | A species of Fluvioviridavis. | |||
| Fluvioviridavis nazensis[185] | Sp. nov | Mayr & Kitchener | Eocene | London Clay | A species of Fluvioviridavis. | |||
Gen. et sp. nov | In press | Wang et al. | An enantiornithine. The type species is I. attenboroughi. | |||||
Gen. et comb. nov | Zelenkov | Late Eocene | An anseriform of uncertain placement; a new genus for "Cygnavus" formosus. | |||||
Sp. nov | Zelenkov | Early Oligocene | A species of Mionetta. | |||||
| Nasiornis[188] | Gen. et sp. nov | In press | Mayr & Kitchener | Eocene | London Clay | A messelornithid. The type species is N. messelornithoides. | ||
Gen. et sp. nov | Valid | Musser & Clarke | Eocene | An early member of Anseriformes. The type species is P. grandei. | ||||
Gen. et sp. nov | Ando et al. | Late Oligocene | An early penguin. The type species is P. hakataramea. | |||||
Gen. et comb. nov | Valid | Zelenkov | Pliocene and Pleistocene | A grouse; a new genus for "Lagopus lagopus" atavus Jánossy (1974), originally described from the Rębielice Królewskie 1 locality in Poland, subsequently also described from the Taurida Cave in Crimea.[191] | ||||
Sp. nov | Mayr & Kitchener | Eocene (Ypresian) | London Clay | A member of Galliformes, possibly belonging to the family Gallinuloididae. | ||||
| ?Parvirallus incertus[188] | Sp. nov | In press | Mayr & Kitchener | Eocene | London Clay | A messelornithid; a possible species of Parvirallus. | ||
| Phalacrocorax bakonyiensis[193] | Sp. nov | Valid | Horváth, Futó, & Kessler | Miocene | A cormorant; a species of Phalacrocorax. | |||
Gen. et 2 sp. et comb. nov | Valid | Mayr & Kitchener | Eocene | London Clay | A possible member of Piciformes. The type species is P. minor; genus also includes new species P. major, as well as "Neanis" kistneri Feduccia (1973). | |||
Sp. nov | Valid | Mayr & Kitchener | Eocene (Ypresian) | London Clay | A stem group roller belonging or related to the family Primobucconidae. | |||
Sp. nov | Valid | Gorbatcheva & Zelenkov | Pleistocene | A vulture, a species of Torgos. | ||||
Gen. et sp. nov | Zelenkov | Early Oligocene | An anatid of uncertain placement. The type species is U. chalkarica. | |||||
| Walbeckornis waltonensis[188] | Sp. nov | In press | Mayr & Kitchener | Eocene | London Clay | A species of Walbeckornis. | ||
Gen. et sp. nov | Valid | Mayr & Kitchener | Eocene (Ypresian) | London Clay | A member of Upupiformes. The type species is W. tendringensis. | |||
Gen. et sp. nov | Mayr & Kitchener | Eocene (Ypresian) | London Clay | A member of Galliformes, the type genus of the new family Waltonortygidae. The type species is W. bumbanipodiides. | ||||
Gen. et comb. nov | Valid | De Mendoza, Degrange & Tambussi | A member of Anseriformes of uncertain affinites; a new genus for "Telmabates" howardae. |
Avian research
[edit]- A study performing quantitative functional imaging of the brain during rest and flight in rock doves with implications for the evolution of avian flight is published by Balanoff et al. (2024). They found increased neural activity in the cerebellum during flight, and through comparisons with cranial endocasts of extinct theropods, suggest that cerebellar expansion underlying such activity occurred at the base of Maniraptora, prior to the origin of avian flight.[197]
- The Cretaceous fossil record of avialans from China is reviewed by Zhou & Wang (2024).[198]
- Evidence of gradual and sequential moult of wing flight feathers in two probable members of Confuciusornithiformes from the Lower Cretaceous Yixian Formation (China) is presented by Wang et al. (2024).[199]
- A morphometric study of a large sample of specimens of Confuciusornis sanctus is published by Zhou et al. (2024), who interpret their findings as indicative of the presence of sexual dimorphism in this species.[200]
- The fossil record of avialans from the Upper Cretaceous Maastricht Formation (Belgium and the Netherlands) is reviewed by Field et al. (2024), who additionally present new data on the bone histology and hindlimb length of Asteriornis maastrichtensis.[201]
- Stoicescu et al. (2024) describe partial femur of an avialan belonging or related to the species Elopteryx nopcsai from the Maastrichtian strata at the Nălaț-Vad locality (Romania), interpret E. nopcsai as a probable secondarily flightless avialan, and argue that Balaur bondoc might be a junior synonym of E. nopcsai.[202]
- A study the relationship between the morphology of cervical vertebrae and dietary modes in extant and extinct birds is published by Liu et al. (2024), who report that Bohaiornis, Brevirostruavis and Longipteryx had cervical morphologies resembling those of extant insectivorous or raptorial birds, while Yanornis and Iteravis had cervical morphologies closer to those of extant generalist or herbivorous birds, falling into the ecological niches of aquatic or semiaquatic birds.[203]
- A study aiming to determine the diets of members of the family Bohaiornithidae is published by Miller et al. (2024), who interpret their findings as indicating that the family included taxa adapted to diverse diets, and predict the ancestral member of Enantiornithes to have been a generalist which ate a wide variety of foods.[204]
- A study on the limb bone histology and growth dynamics of Musivavis amabilis is published by Kundrát et al. (2024).[205]
- The Cretaceous fossil record of avialans from Antarctica is reviewed by Acosta Hospitaleche et al. (2024).[206]
- A study on the antiquity of the crown group of birds is published by Brocklehurst & Field (2024), who argue that the crown group originated between 110.5 and 90.3 million years ago, and that the majority of higher-order diversification within the crown group either spanned or postdated the Cretaceous-Paleogene transition.[207]
- A study on patterns of avian molecular evolution is published by Berv et al. (2024), who interpret their findings as indicating that the Cretaceous–Paleogene extinction event influenced the evolution of bird genomes, physiology and life history traits that in turn influenced the diversification of modern birds.[208]
- Widrig, Navalón & Field (2024) describe the external and internal morphology of the braincase of Lithornis vulturinus, interpret its neuroanatomy as likely similar to the neuroanatomy of the ancestral crown bird, and interpret L. vulturinus as a diurnal bird that likely was reliant on visual cues and had a well-developed sense of smell.[209]
- The histochemistry of an ostrich eggshell from the Miocene Liushu Formation (China) is examined by Wu et al. (2024).[210]
- Schroeter (2024) presents a characterization of diagenetiforms in a moa proteome.[211]
- Pickford (2024) revises fossil eggshells from the Miocene strata from the Karingarab aeolianite succession (Namibia), originally described as Struthio karingarabensis, and transfers this oospecies to the genus Diamantornis.[212]
- A draft genome of the little bush moa is presented by Edwards et al. (2024).[213]
- Tomlinson et al. (2024) reconstruct the range and extinction dynamics of six species of moa, and interpret their findings as indicating that the studied species likely had similar spatial patterns of geographic range collapse, and that their final populations persisted in cold, mountainous areas that continue to function as sanctuaries for New Zealand's remaining flightless birds.[214]
- Fossil material of a possible member of Galloanserae is described from the Upper Cretaceous (Maastrichtian) Lance Formation (Wyoming, United States) by Brownstein (2024), who interprets this finding as supporting a cosmopolitan distribution of early crown birds.[215]
- McInerney, Blokland & Worthy (2024) redescribe the skull morphology of Genyornis newtoni and study its phylogenetic affinities, recovering the family Dromornithidae as more likely to be members of Anseriformes related to screamers than close relatives of the family Gastornithidae.[216]
- A study on the vertebral column of Annakacygna hajimei is published by Matsuoka, Seoka & Hasegawa (2024), who reconstruct the neck of this bird with a curve at its base that increased the buoyancy and stability of the bird's body when it was in the water by helping it to put the base of the neck with its air sacs below the water surface.[217]
- A case for the validity of Miotadorna catrionae is presented by Tennyson et al. (2024),[218] in response to Worthy et al. (2022)[219] considering it a junior synonym of Miotadorna sanctibathansi.
- Evidence from the study of mitogenomes of the extant Brazilian merganser and extinct Auckland Island merganser, interpreted as indicating that the studied mergansers are not sister taxa and that their ancestors moved into the Southern Hemisphere in two separate colonization events at least 7 million years ago, is presented by Rawlence et al. (2024).[220]
- A study on the evolutionary history of neoavians, as indicated by genomic data, is published by Wu et al. (2024), who argue that the initial diversification of the crown group of birds was correlated with the rise of flowering plants in the Cretaceous, that modern birds survived the Cretaceous–Paleogene extinction event relatively well, and that the Paleocene–Eocene Thermal Maximum had a significant impact on the diversification of the seabirds.[221]
- Zelenkov (2024) describes a fragmentary humerus of a buttonquail from the Lower Pleistocene strata from the Taurida Cave (Crimea), representing the first record of a member of the family Turnicidae from Eurasia from the Pliocene to Middle Pleistocene interval.[222]
- Goodman & Rasolonjatovo (2024) study the carpal spur of the Malagasy lapwing, find it to be larger than wing spurs of living lapwings, and interpret it as likely used for defence against predators.[223]
- A study on the internal structure and resistance to bending forces of tarsometatarsi of extant and Eocene penguins is published by Jadwiszczak, Krüger & Mörs (2024).[224]
- A new specimen of Palaeeudyptes is described by Xia, Pei & Li (2024).[225]
- A study on the long limb bone microstructure of extant king penguins throughout their ontogeny is published by Canoville, Robin & de Buffrénil (2024), who find evidence of substantial intraspecific variability regardless of the ontogenetic stage, and evidence indicating that limb bones of king penguins reach adult size early in the development while their microstructure continues to change until adulthood; on the basis of their findings the authors do not consider the conclusions of Cerda, Tambussi & Degrange (2014)[226] and Ksepka et al. (2015)[227] about the paleobiology of fossil penguins to be properly supported by their data.[228]
- The evolutionary dynamics of microsatellites in Adélie penguins based on both modern and ancient genetic samples (up to 46.5 thousand years old) are studied by McComish et al. (2024).[229]
- Leoni et al. (2024) describe the first fossil material of a turkey vulture from cave deposits in northeastern Brazil, which preserves trace marks likely produced by a felid and indicating that the vulture died in the cave it was discovered in.[230]
- A study on the age of remains of California condors from the Mule Ears Peak Cave (Texas, United States) is published by Emslie (2024), who find evidence of the presence of condors at the studied site beginning at ~15,000 calendar years before present and evidence of definite nesting ~13,000 calendar years before present, reports evidence from stable isotope analysis of bone collagen interpreted as indicating that the studied condors fed on megafauna living in a desert grassland ecosystem, and interprets these findings as indicating that the disappearance of the California condor from the inland west of North America as related to the extinctions of megafauna the end of the Pleistocene.[231]
- The colonization of the Mediterranean Basin by Bonelli's eagle is studied by Moleón et al. (2024), drawing on data from environmental favorability, genetic structure, the fossil record, and ecological relationships with golden eagles.[232]
- Acosta Hospitaleche & Jones (2024) describe fossil material of a large-bodied (with an estimated body mass of around 100 kg) phorusrhacid or phorusrhacid-like bird from the Eocene La Meseta Formation (Seymour Island, Antarctica), interpreted by the authors as likely apex predator of Antarctica during the Eocene.[233]
- A study on the phylogenetic relationships and on the evolution of body size and cursoriality in phorusrhacids, providing evidence of niche partitioning and competitive exclusion that controlled phorusrhacid diversity, is published by LaBarge, Gardner & Organ (2024).[234]
- Acosta Hospitaleche & Jones (2024) describe partial tibiotarsus of a psilopterine phorusrhacid from the Eocene (Lutetian) Sarmiento Formation (Argentina), interpreted as belonging to a bird with an estimated body mass of approximately 5 kg.[235]
- A carpometacarpus of a Cuban macaw is described from the Pleistocene of El Abrón Cave (Cuba) by Zelenkov (2024).[236]
- A study on the phylogenetic relationships of Wieslochia weissi, Crosnoornis nargizia, Jamna szybiaki, Resoviaornis jamrozi and an unnamed passerine from the Oligocene of France described by Riamon, Tourment & Louchart (2020)[237] is published by Lowi-Merri et al. (2024).[238]
- Pöllath & Peters (2024) study the composition of early Holocene bird assemblages from southeast Turkey, northern Syria and northern Iraq, providing evidence of changes of bird species ranges related to climatic changes during the Pleistocene-Holocene transition, aridification during the Holocene and human activities.[239]
Pterosaurs
[edit]New pterosaur taxa
[edit]| Name | Novelty | Status | Authors | Age | Type locality | Country | Notes | Images |
|---|---|---|---|---|---|---|---|---|
| Ceoptera[240] | Gen. et sp. nov | Valid | Martin-Silverstone et al. | Middle Jurassic | Kilmaluag Formation | A darwinopteran. The type species is C. evansae. | ||
Gen. et sp. nov | Valid | Pentland et al. | Early Cretaceous (Albian) | A member of Anhangueria. The type species is H. peterseni. |  | |||
Gen. et sp. nov | Valid | Spindler | Painten Formation | A transitional monofenestratan. The type species is P. frankerlae. |
| |||
| Torukjara[243] | Gen. et sp. nov | Valid | Pêgas | Early Cretaceous | Caiuá Group | A tapejarid. The type species is T. bandeirae. |  |

Pterosaur research
[edit]- A study on the cervical osteology of Anhanguera piscator, Azhdarcho lancicollis and Rhamphorhynchus muensteri, aiming to reconstruct the cervical arthrology of pterosaurs and the position of the pterosaur neck at rest, is published by Buchmann & Rodrigues (2024).[244]
- A study on the palate structure in Kunpengopterus, Hongshanopterus, Hamipterus and Dsungaripterus, providing new information on the relations between the palatine, ectopterygoid, maxilla and pterygoid in the studied pterosaurs resulting in reinterpretation of the main palatal openings, and identifying an opening bordered anteriorly by the maxilla and posteriorly by the palatine that is unique within Diapsida and might be a synapomorphy of Pterosauria, is published by Chen et al. (2024).[245]
- A study aiming to determine the aerodynamic impact of large heads and head crests of pterosaurs is published by Henderson (2024).[246]
- Yun (2024) uses geometric morphometric analyses to investigate the relationships of pterosaur specimens from the Early Cretaceous Jinju and Hasandong formations (South Korea), and suggests that the material likely cannot be assigned to the Boreopteridae, as had previously been assumed.[247]
- Habib & Hone (2024) study the variation seen in elements and body parts of specimens of Rhamphorhynchus muensteri, providing evidence of high levels of constraint throughout the appendicular and axial elements that were likely important for flight, and evidence of increased variability of tails of larger individuals, possibly related to the signalling function of the tail.[248]
- So, Kim & Won (2024) describe a nearly complete skeleton of a probable member of the genus Jeholopterus from the Lower Cretaceous Sinuiju Formation, representing the first pterosaur recond from North Korea reported to date.[249]
- Heredia et al. (2024) describe new tracks of pterodactyloid pterosaurs from the Cenomanian Candeleros Formation (Argentina) with a different morphology from previously recorded tracks from this formation, interpreted as more likely produced by individuals of different ages rather than different species.[250]
- Partial finger phalanx of a member of Ctenochasmatoidea with an estimated wingspan of at least 3 m, representing one of the first records of Jurassic pterodactyloids from the United Kingdom, is described from the Kimmeridge Clay of Abingdon, Oxfordshire by Etienne et al. (2024).[251]
- Description of the anatomy of the ankle of Pterodaustro guinazui is published by Burlot et al. (2024).[252]
- Redescription and a study on the affinities of Haopterus gracilis is published by Xu, Jiang & Wang (2024), who recover H. gracilis as a member of Istiodactyliformes.[253]
- Ciaffi & Bellardini (2024) describe isolated teeth of indeterminate members of Ornithocheiriformes from the Lohan Cura Formation (Neuquén Province, Argentina), providing evidence of a more abundant and diversified ornithocheiriform fauna in the south of the Neuquén Basin (at least in the Albian) than previously known.[254]
- A study evaluating the ability of different proposed take-off motions of pterosaurs to produce leverage during the launch phase, as indicated by tests using a musculoskeletal model based on an indeterminate ornithocheiraean pterosaur with a 5 m wingspan, is published by Griffin et al. (2024).[255]
- Jung & Huh (2024) describe pterosaur tracks from the Turonian Jangdong Formation (South Korea), interpreted as likely produced by small-bodied or immature azhdarchids and as probable evidence of gregariousness of the trackmakers.[256]
Other archosaurs
[edit]Other archosaur research
[edit]- Garcia et al. (2024) describe two new lagerpetid specimens from the Carnian strata of the upper Santa Maria Formation (Brazil), interpreted as indicative of a sympatric occurrence of lagerpetids representing different morphotypes.[257]
- Agnolín et al. (2024) revise the anatomy of the pelvic girdle of Lagerpeton chanarensis, reinterpreting it as likely to have a sprawling gait.[258]
General research
[edit]- A study on the evolution of locomotion in archosauromorph reptiles is published by Shipley et al. (2024), who interpret their findings as indicative of greater range in limb form and locomotor modes of dinosaurs compared to other archosauromorph groups, and argue that the ability to adopt a wider variety of limb forms and modes might have given dinosaurs a competitive advantage over pseudosuchians.[259]
- A study on the body size evolution of non-avian dinosaurs and Mesozoic birds is published by Wilson et al. (2024), who find no evidence that Bergmann's rule applied to the studied taxa.[260]
- Knoll, Ishikawa & Kawabe (2024) present a new method which can be used to determine the brain volume of extinct archosaurs on the basis their endocranial cavity volume.[261]
- Malafaia et al. (2024) revise fossils from Portugal that were historically assigned to Megalosaurus, and find that the majority of this fossil material represents bones of members of different theropod groups, but also that the studied material includes stegosaurian, iguanodontian, sauropod and thalattosuchian bones.[262]
- Dinosaur and probable crocodylomorph tracks, including some of the largest sauropod tracks worldwide, are described from the Bathonian strata in the El Mers area (Morocco) by Amzil et al. (2024).[263]
References
[edit]- ^ Martins, K. C.; Queiroz, M. V.; Ruiz, J. V.; Langer, M. C.; Montefeltro, F. C. (2024). "A new Baurusuchidae (Notosuchia, Crocodyliformes) from the Adamantina Formation (Bauru Group, Upper Cretaceous), with a revised phylogenetic analysis of Baurusuchia". Cretaceous Research. 153. 105680. Bibcode:2024CrRes.15305680M. doi:10.1016/j.cretres.2023.105680. S2CID 261182849.
- ^ Fernández Dumont, M. L.; Pol, D.; Bona, P.; Apesteguía, S. (2024). "A new species of Araripesuchus with durophagous dentition increases the ecological disparity among uruguaysuchid crocodyliforms". Journal of Systematic Palaeontology. 22 (1). 2373987. doi:10.1080/14772019.2024.2373987.
- ^ Narváez, I.; de Celis, A.; Escaso, F.; Martín de Jesús, S.; Pérez-García, A.; Ortega, F. (2024). "A new Crocodyloidea from the middle Eocene of Zamora (Duero Basin, Spain)". The Anatomical Record. doi:10.1002/ar.25422. PMID 38444286.
- ^ Smith, Nathan D.; Klein, Nicole; Sander, P. Martin; Schmitz, Lars (July 2024). "A new pseudosuchian from the Favret Formation of Nevada reveals that archosauriforms occupied coastal regions globally during the Middle Triassic". Biology Letters. 20 (7). 20240136. doi:10.1098/rsbl.2024.0136. ISSN 1744-957X. PMC 11286145. PMID 38982977.
- ^ Iori, F. V.; Ghilardi, A. M.; Fernandes, M. A.; Dias, W. A. F. (2024). "A new species of vocalizing crocodyliform (Notosuchia, Sphagesauridae) from the Late Cretaceous of Brazil". Historical Biology: An International Journal of Paleobiology: 1–12. doi:10.1080/08912963.2024.2364332.
- ^ Sachs, Sven; Young, Mark T.; Hornung, Jahn J.; Cowgill, Thomas; Schwab, Julia A.; Brusatte, Stephen L. (2024-12-31). "A new genus of metriorhynchid crocodylomorph from the Lower Cretaceous of Germany". Journal of Systematic Palaeontology. 22 (1). Bibcode:2024JSPal..2259946S. doi:10.1080/14772019.2024.2359946. ISSN 1477-2019.
- ^ Reyes, W. A.; Martz, J. W.; Small, B. J. (2024). "Garzapelta muelleri gen. et sp. nov., a new aetosaur (Archosauria: Pseudosuchia) from the Late Triassic (middle Norian) middle Cooper Canyon Formation, Dockum Group, Texas, USA, and its implications on our understanding of the morphological disparity of the aetosaurian dorsal carapace". The Anatomical Record. 307 (4): 1271–1299. doi:10.1002/ar.25379. PMID 38206046. S2CID 266931123.
- ^ López-Rojas, V.; Mateus, S.; Marinheiro, J.; Mateus, O.; Puértolas-Pascual, E. (2024). "A new goniopholidid crocodylomorph from the Late Jurassic of Portugal". Palaeontologia Electronica. 27 (1). 27.1.5a. doi:10.26879/1316.
- ^ Müller, Rodrigo T. (2024-06-20). "A new small-sized predatory pseudosuchian archosaur from the Middle-Late Triassic of Southern Brazil". Scientific Reports. 14 (1): 12706. Bibcode:2024NatSR..1412706M. doi:10.1038/s41598-024-63313-3. ISSN 2045-2322. PMC 11189902. PMID 38902259.
- ^ Desojo, J. B.; Rauhut, O. W. M. (2024). "Reassessment of the enigmatic "Prestosuchus" loricatus (Archosauria: Pseudosuchia) from the Middle-Late Triassic of southern Brazil". The Anatomical Record. 307 (4): 974–1000. doi:10.1002/ar.25401. PMID 38344898.
- ^ Pochat-Cottilloux, Yohan; Lauprasert, Komsorn; Chanthasit, Phornphen; Manitkoon, Sita; Adrien, Jérôme; Lachambre, Joël; Amiot, Romain; Martin, Jeremy E. (2024-01-09). "New Cretaceous neosuchians (Crocodylomorpha) from Thailand bridge the evolutionary history of atoposaurids and paralligatorids". Zoological Journal of the Linnean Society. In press: 1–27. doi:10.1093/zoolinnean/zlad195.
- ^ Sennikov, A. G. (2024). "Ornithosuchidae—Early Archosaurs with a Hyperspecialized Jaw Apparatus". Paleontological Journal. 58 (1): 1–19. Bibcode:2024PalJ...58....1S. doi:10.1134/S0031030124010064.
- ^ von Baczko, M. B.; Zariwala, J.; Ballentine, S. E.; Desojo, J. B.; Hutchinson, J. R. (2024). "Biomechanical modeling of musculoskeletal function related to the terrestrial locomotion of Riojasuchus tenuisceps (Archosauria: Ornithosuchidae)". The Anatomical Record. doi:10.1002/ar.25528. PMID 38943347.
- ^ Desojo, J. B.; von Baczko, M. B.; Ezcurra, M. D.; Fiorelli, L. E.; Martinelli, A. G.; Bona, P.; Trotteyn, M. J.; Lacerda, M. (2024). "Cranial osteology and paleoneurology of Tarjadia ruthae: An erpetosuchid pseudosuchian from the Triassic Chañares Formation (late Ladinian-?early Carnian) of Argentina". The Anatomical Record. 307 (4): 890–924. doi:10.1002/ar.25382. PMID 38263705. S2CID 267198765.
- ^ Klein, N. (2024). "Diverse growth rates in Triassic archosaurs—insights from a small terrestrial Middle Triassic pseudosuchian". The Science of Nature. 111 (4). 38. Bibcode:2024SciNa.111...38K. doi:10.1007/s00114-024-01918-4. PMC 11239758. PMID 38990382.
- ^ Nesbitt, S. J.; Chatterjee, S. (2024). "The osteology of Shuvosaurus inexpectatus, a shuvosaurid pseudosuchian from the Upper Triassic Post Quarry, Dockum Group of Texas, USA". The Anatomical Record. 307 (4): 1175–1238. doi:10.1002/ar.25376. hdl:10919/117738. PMID 38258540.
- ^ Mastrantonio, B. M.; Lacerda, M. B.; de Farias, B. D. M.; Pretto, F. A.; Rezende, L. O.; Desojo, J. B.; Schultz, C. L. (2024). "Postcranial anatomy of Prestosuchus chiniquensis (Archosauria: Loricata) from the Triassic of Brazil". The Anatomical Record. 307 (4): 925–956. doi:10.1002/ar.25383. PMID 38299218. S2CID 267362910.
- ^ Spiekman, S. N. F.; Butler, R. J.; Maidment, S. C. R. (2024). "The postcranial anatomy and osteohistology of Terrestrisuchus gracilis (Archosauria, Crocodylomorpha) from the Late Triassic of Wales". Papers in Palaeontology. 10 (4). e1577. doi:10.1002/spp2.1577.
- ^ Woodward, H. N.; Aubier, P.; Sena, M. V. A.; Cubo, J. (2024). "Evaluating extinct pseudosuchian body mass estimates using a femur volume-based model". The Anatomical Record. doi:10.1002/ar.25452. PMID 38634509.
- ^ Scavezzoni, I.; Fischer, V.; Johnson, M. M.; Jouve, S. (2024). "Form and function of the pelvic girdle of Thalattosuchia and Dyrosauridae (Crocodyliformes)". Geodiversitas. 46 (6): 135–326. doi:10.5252/geodiversitas2024v46a6. hdl:2268/308796.
- ^ Young, M. T.; Wilberg, E. W.; Johnson, M. M.; Herrera, Y.; Brandalise de Andrade, M.; Brignon, A.; Sachs, S.; Abel, P.; Foffa, D.; Fernández, M. S.; Vignaud, P.; Cowgill, T.; Brusatte, S. L. (2024). "The history, systematics, and nomenclature of Thalattosuchia (Archosauria: Crocodylomorpha)". Zoological Journal of the Linnean Society. 200 (2): 547–617. doi:10.1093/zoolinnean/zlad165.
- ^ Bhuttarach, S.; Deesri, U.; Warapeang, P.; Taesuk, N.; Lauprasert, K. (2024). "Morphology of teleosaurid osteoderms from the Phu Kradung Formation of Thailand". Annales de Paléontologie. 109 (4). 102653. doi:10.1016/j.annpal.2023.102653. S2CID 267691380.
- ^ Weryński, Ł.; Błażejowski, B.; Szczygielski, T.; Young, M. T. (2024). "The first occurrence of machimosaurid crocodylomorphs from the Oxfordian of south-central Poland provides new insights into the distribution of macrophagous teleosauroids". PeerJ. 12. e17153. doi:10.7717/peerj.17153. PMC 10981889. PMID 38560470.
- ^ Scheyer, T. M.; Johnson, M. M.; Bastiaans, D.; Miedema, F.; Maxwell, E. E.; Klug, C. (2024). "Oldest record of Machimosaurini (Thalattosuchia, Teleosauroidea): teeth and scavenging traces from the Middle Jurassic (Bajocian) of Switzerland". Royal Society Open Science. 11 (4). 240071. Bibcode:2024RSOS...1140071S. doi:10.1098/rsos.240071. PMC 11004672. PMID 38601027.
- ^ Hua, S.; Liston, J.; Tabouelle, J. (2024). "The Diet of Metriorhynchus (Thalattosuchia, Metriorhynchidae): Additional Discoveries and Paleoecological Implications". Fossil Studies. 2 (1): 66–76. doi:10.3390/fossils2010002.
- ^ Navarro, T. G.; Cerda, I. A.; Fernández Dumont, M. L.; Apesteguía, S.; Pol, D. (2024). "New data on the bone histology of Araripesuchus buitreraensis (Crocodylomorpha: Notosuchia) from the Late Cretaceous of Argentinean Patagonia". Historical Biology: An International Journal of Paleobiology: 1–11. doi:10.1080/08912963.2023.2301140. S2CID 266948988.
- ^ Borsoni, B. T.; Carvalho, I. S.; Marinho, T. S. (2024). "Armadillosuchus arrudai (Sphagesauridae, Crocodyliformes), Adamantina Formation (Turonian - Santonian), Bauru Basin, southeastern Brazil: Dental development aspects". Cretaceous Research. 158. 105838. Bibcode:2024CrRes.15805838D. doi:10.1016/j.cretres.2024.105838. S2CID 267077357.
- ^ dos Santos, D. M.; de Carvalho, J. C.; de Oliveira, C. E. M.; de Andrade, M. B.; Santucci, R. M. (2024). "Cranial and postcranial anatomy of a juvenile baurusuchid (Notosuchia, Crocodylomorpha) and the taxonomical implications of ontogeny". The Anatomical Record. doi:10.1002/ar.25419. PMID 38429867. S2CID 268121558.
- ^ Obuse, S.; Shibata, M. (2024). "New goniopholidid specimens from the Lower Cretaceous Kitadani Formation, Tetori Group, Japan". Annales de Paléontologie. 110 (1). 102661. Bibcode:2024AnPal.11002661O. doi:10.1016/j.annpal.2023.102661. S2CID 268190726.
- ^ Forêt, T.; Aubier, P.; Jouve, S.; Cubo, J. (2024). "Biotic and abiotic factors and the phylogenetic structure of extinction in the evolution of Tethysuchia". Paleobiology. 50 (2): 285–307. Bibcode:2024Pbio...50..285F. doi:10.1017/pab.2024.5.
- ^ Kuzmin, I. T.; Sichinava, E. A.; Mazur, E. V.; Gombolevskiy, V. A. (2024). "Virtual reconstruction of the neurocranial anatomy of Kansajsuchus extensus (Neosuchia: Paralligatoridae) from the Upper Cretaceous of Tadzhikistan with a review of braincase osteology in Neosuchia". Cretaceous Research. 164. 105959. doi:10.1016/j.cretres.2024.105959.
- ^ Rocchi, R.; Vila, B. (2024). "New eusuchian cranial remains from the Upper Cretaceous of the southern Pyrenees". Historical Biology: An International Journal of Paleobiology: 1–9. doi:10.1080/08912963.2024.2350551.
- ^ Yates, A. M.; Stein, M. (2024). "A reinterpretation and taxonomic revision of Ultrastenos willisi Stein, Hand and Archer, 2016, a short-snouted mekosuchine crocodylian from the Oligocene of northern Australia". Palaeontologia Electronica. 27 (1). 27.1.a22. doi:10.26879/1355.
- ^ Stout, J. B. (2024). "Osmoregulation in Alligatoroidea: shifting the paradigm untethers biogeographic questions". Historical Biology: An International Journal of Paleobiology: 1–6. doi:10.1080/08912963.2024.2379029.
- ^ Conedera, D.; Pochat-Cottilloux, Y.; Rinder, N.; Adrien, J.; Martin, J. E. (2024). "An anatomical reappraisal of the dwarf crocodylian Arambourgia gaudryi from the Eocene of Quercy (France) using CT data and its implications for the phylogeny and paleoecology of basally branching alligatoroids". Journal of Vertebrate Paleontology. 43 (4). e2313612. doi:10.1080/02724634.2024.2313612.
- ^ Chabrol, N.; Jukar, A. M.; Patnaik, R.; Mannion, P. D. (2024). "Osteology of Crocodylus palaeindicus from the late Miocene–Pleistocene of South Asia and the phylogenetic relationships of crocodyloids". Journal of Systematic Palaeontology. 22 (1). 2313133. Bibcode:2024JSPal..2213133C. doi:10.1080/14772019.2024.2313133.
- ^ Zheng, W.; Jin, X.; Xie, J.; Du, T. (2024). "The first deep-snouted tyrannosaur from Upper Cretaceous Ganzhou City of southeastern China". Scientific Reports. 14 (1). 16276. doi:10.1038/s41598-024-66278-5. ISSN 2045-2322. PMC 11272791. PMID 39054316.
- ^ Ning, Li; Maidment, Susannah C. R.; Daqing, Li; Hailu, You; Guangzhao, Peng (2024-07-02). "A new stegosaur (Dinosauria: Ornithischia) from the Middle Jurassic of Gansu Province, China". Scientific Reports. 14 (1): 15241. Bibcode:2024NatSR..1415241N. doi:10.1038/s41598-024-66280-x. ISSN 2045-2322. PMC 11219857. PMID 38956140.
- ^ Альварес Ногейра, Р.; Розадилья, С.; Аньолин, Флорида; Гарсиа Марса, JA; Мотта, MJ; Новас, FE (2024). «Новый орнитопод из верхнего мела (формация Уинкул) северо-западной Патагонии, Аргентина. Влияние на посткраниальную анатомию эласмариев». Меловые исследования . 159 . 105874. Бибкод : 2024CrRes.15905874N . дои : 10.1016/j.cretres.2024.105874 .
- ^ Локвуд, Джереми А.Ф.; Мартилл, Дэвид М.; Мейдмент, Сюзанна ЧР (31 декабря 2024 г.). «Comptonatus Chasei, новый игуанодонтический динозавр из нижнемеловой формации Уэссекс на острове Уайт, южная Англия» . Журнал систематической палеонтологии . 22 (1). Бибкод : 2024JSPal..2246573L . дои : 10.1080/14772019.2024.2346573 . ISSN 1477-2019 .
- ^ Син, Л.; Ню, К.; Мэллон, Дж.; Мияшита, Т. (2024). «Новый панцирный динозавр с двойными рогами из раннего позднего мела на юго-востоке Китая» . Анатомия позвоночных Морфология Палеонтология . 11 : 113–132. дои : 10.18435/vamp29396 .
- ^ Порфирий, Джон Д.; Баяно, Маттиа А.; дос Сантос, Доменика Д.; Джанекини, Федерико А.; Питтман, Майкл; Ламанна, Мэтью К. (14 июня 2024 г.). « Diuqin lechiguanae gen. et sp. nov., новый уненлагиин (Theropoda: Paraves) из формации Бахо-де-ла-Карпа (группа Неукен, верхний мел) провинции Неукен, Патагония, Аргентина» . BMC Экология и эволюция . 24 (1): 77. Бибкод : 2024BMCEE..24...77P . дои : 10.1186/s12862-024-02247-w . ISSN 2730-7182 . ПМЦ 11177497 . ПМИД 38872101 .
- ^ Барон, Мэтью Г. (29 апреля 2024 г.). «Новое название для старых костей: переоценка останков раннеюрских теропод из Дорсета, Англия» . Электронная палеонтология . 27 (1): 1–12. дои : 10.26879/1346 . ISSN 1094-8074 .
- ^ Аткинс-Вельтман, КЛ; Саймон, диджей; Вудворд, Х.Н.; Фанстон, Г.Ф.; Снайвли, Э. (2024). «Новый овирапторозавр (Dinosauria: Theropoda) из формации Хелл-Крик конца Маастрихта в Северной Америке» . ПЛОС ОДИН . 19 (1). е0294901. Бибкод : 2024PLoSO..1994901A . дои : 10.1371/journal.pone.0294901 . ПМЦ 10807829 . ПМИД 38266012 .
- ^ Авраами, Хавив М.; Маковицкий, Питер Дж.; Такер, Райан Т.; Занно, Линдси Э. (9 июля 2024 г.). «Новый полуфоссорный динозавр-тецелозавр из сеноманского возраста, члена Муссентучит, формации Кедровая гора, штат Юта» . Анатомическая запись . дои : 10.1002/ar.25505 . ISSN 1932-8486 . ПМИД 38979930 .
- ^ Хан, Ф.; Ян, Л.; Лу, Ф.; Салливан, Дж.; Сюй, Х.; Цю, В.; Лю, Х.; Ю, Дж.; Ву, Р.; Ключ.; Сюй, М.; Ху, Дж.; Лу, П. (2024). «Новый зауропод-титанозавр, Gandititan cavocaudatus gen. et sp. nov., из позднего мела южного Китая» . Журнал систематической палеонтологии . 22 (1). 2293038. Бибкод : 2024JSPal..2293038H . дои : 10.1080/14772019.2023.2293038 . S2CID 267107071 .
- ^ Ли, С.; Ли, Ю.-Н.; Парк, Ж.-Ю.; Ким, С.-Х.; Бадамхатан, З.; Идерсайхан, Д.; Цогтбаатар, К. (2024). «Первый троодонтид (Dinosauria: Theropoda) из верхнемеловой баруунгойотской свиты Монголии » Журнал палеонтологии позвоночных . е2364746. дои : 10.1080/02724634.2024.2364746 .
- ^ Ротатори, FM; Феррари, Л.; Секеро, К.; Камило, Б.; Матеус, О.; Морено-Азанза, М. (2024). «Неожиданный раннерасходящийся игуанодонтический динозавр (Ornithischia, Ornithopoda) из верхней юры Португалии». Журнал палеонтологии позвоночных . 43 (4). е2310066. дои : 10.1080/02724634.2024.2310066 .
- ^ Кубота, К.; Кобаяши, Ю.; Икеда, Т. (2024). «Раннемеловой троодонтиновый троодонтид (Dinosauria: Theropoda) из формации Охьямашимо в Японии раскрывает раннюю эволюцию Troodontinae» . Научные отчеты . 14 (1). 16392. дои : 10.1038/s41598-024-66815-2 . ПМЦ 11272788 . ПМИД 39054320 .
- ^ Филиппи, Леонардо С.; Хуарес Вальери, Рубен Д.; Галлина, Пабло А.; Мендес, Ариэль Х.; Джанекини, Федерико А.; Гарридо, Альберто К. (2024). «Титанозавр, имитирующий реббахизавридов, и свидетельства нарушения фауны в позднем меловом периоде на юго-западе Гондваны». Меловые исследования . 154 . Бибкод : 2024CrRes.15405754F . дои : 10.1016/j.cretres.2023.105754 . ISSN 0195-6671 . S2CID 264792693 .
- ^ Рен, Х.-Х.; Ван, X.-R.; Джи, Ю.-Н.; Го, З.; Джи, К. (2024). «Первый маменчизаврид из верхнеюрской формации Дунсин в Гуанси, самый южный Китай». Историческая биология: Международный журнал палеобиологии : 1–14. дои : 10.1080/08912963.2024.2309287 . S2CID 267947729 .
- ^ «Джингия» .
- ^ «Дополнение» . Историческая биология : 1. 15 апреля 2024 г. дои : 10.1080/08912963.2024.2325806 . ISSN 0891-2963 .
- ^ Аверьянов, АО; Скутщас, ПП; Атучин А.А.; Слободин Д.А.; Феофанова О.А.; Владимирова, ОН (2024). «Последний цератозавр Азии: новый ноазаврид из раннемелового Великого сибирского рефугиума». Труды Королевского общества B: Биологические науки . 291 (2023). 20240537. дои : 10.1098/rspb.2024.0537 . ПМИД 38747705 .
- ^ Пол, Диего; Баяно, Маттиа Антонио; Черны, Давид; Новас, Фернандо Э.; Серда, Игнасио А.; Питтман, Майкл (21 мая 2024 г.). «Новый динозавр-абелизаврид из конца мела Патагонии и темпы эволюции среди цератозавров» . Кладистика . 40 (3): 307–356. дои : 10.1111/cla.12583 . ISSN 0748-3007 . ПМИД 38771085 .
- ^ Лоуэн, Марк А.; Сертич, Джозеф JW; Сэмпсон, Скотт ; О'Коннор, Цзинмай К .; Карпентер, Саванна; Сиссон, Брок; Оленшлегер, Анна; Фарке, Эндрю А.; Маковицкий, Питер Дж.; Лонгрич, Ник; Эванс, Дэвид К. (20 июня 2024 г.). « Lokiceratops rangiformis gen. et sp. nov. (Ceratopsidae: Centrosaurinae) из кампанской формации Джудит-Ривер в Монтане обнаруживает быструю региональную радиацию и крайний эндемизм среди центрозавровых динозавров» . ПерДж . 12 : е17224. дои : 10.7717/peerj.17224 . ISSN 2167-8359 . ПМЦ 11193970 . ПМИД 38912046 .
- ^ Лонгрич, Северная Каролина; Переда-Субербиола, X.; Бардет, Н.; Джалиль, Н.-Э. (2024). «Новый небольшой утконосый динозавр (Hadrosauridae: Lambeosaurinae) из Марокко и разнообразие динозавров в позднем маастрихте Северной Африки » Научные отчеты 14 (1). 3665. Бибкод : 2024НатСР..14.3665Л . дои : 10.1038/ s41598-024-53447-9 ПМЦ 10864364 . ПМИД 38351204 .
- ^ Барретт, Пол М .; Шапель, Кимберли Э.Дж.; Шишио, Лара; Бродерик, Тимоти Дж.; Зондо, Мишель; Муньиква, Дарлингтон; Шуаньер, Иона Н. «Новый динозавр-зауроподоморф позднего триаса из бассейна Средней Замбези, Зимбабве» . Acta Palaeontologica Polonica . 69 (2): 227–241. дои : 10.4202/app.01100.2023 .
- ^ Исасменди, Э.; Слоуп, Э.; Диас-Мартинес, И.; Компания, Дж.; Саес-Бенито, П.; Виера, Л.И.; Торичес, А.; Переда-Субербиола, П. (2024). «Увеличение количества теропод в Европе: новый базальный спинозаврид из группы Энсизо бассейна Камерос (Ла-Риоха, Испания). Эволюционные последствия и палеобиоразнообразие» . Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlad193 .
- ^ Лерзо, Лукас Николас; Галлина, Пабло Ариэль; Канале, Хуан Игнасио; Отеро, Алехандро; Карбаллидо, Хосе Луис; Апестегия, Себастьян; Маковицкий, Петер Юрай (3 января 2024 г.). «Последний из старых: базальный реббахизаврид (Sauropoda, Diplodocoidea) из раннего позднего мела (сеноман-турон) Патагонии, Аргентина» . Историческая биология : 1–26. дои : 10.1080/08912963.2023.2297914 . ISSN 0891-2963 . S2CID 266865502 .
- ^ Зафати, Омар; Укассу, Мостафа; Ригуетти, Факундо; Компания Хулио; Бендриуа, Саад; Табусе, Родольф; Шарьер, Андре; Переда-Субербиола, Хавьер (29 марта 2024 г.). «Новый стегозавровый динозавр (Ornithischia: Thyreophora) с замечательным кожным панцирем из средней юры Северной Африки» . Исследования Гондваны . 131 : 344–362. Бибкод : 2024GondR.131..344Z . дои : 10.1016/j.gr.2024.03.009 . ISSN 1342-937X .
- ^ Перейра, PVLGC; Флаг, КЛН; Видаль, Л.С.; Рибейро, ТБ; Кандейро, CRA; Бергквист, LP (2024). «Новый вид зауроподов из северо-западной Бразилии: биомеханика и радиация титанозаврий (Sauropoda: Somphospondyli)». Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlae054 .
- ^ Перейти обратно: а б Бандейра, КЛН; Наварро, бакалавр; Пегас, Р.В.; Брильанте, Н.С.; Брум, А.С.; де Соуза, LG; да Силва, RC; Галло, В. (2024). «Повторная оценка исторических находок окаменелостей из штата Баия (северо-восток Бразилии) выявила разнообразную фауну динозавров в нижнем меловом периоде Южной Америки» . Историческая биология: Международный журнал палеобиологии : 1–42. дои : 10.1080/08912963.2024.2318406 .
- ^ Перес-Морено, А.; Сальгадо, Л.; Карбаллидо, JL; Отеро, А.; Пол, Д. (2024). «Новый титанозавр из формации Ла Колония (кампан-маастрихт), провинция Чубут, Аргентина» . Историческая биология: Международный журнал палеобиологии : 1–20. дои : 10.1080/08912963.2024.2332997 .
- ^ Далман, С.Г.; Лоуэн, Массачусетс; Пайрон, РА; Ясински, SE; Малинзак, Делавэр; Люк, СГ; Фиорилло, Арканзас; Карри, Пи Джей; Лонгрич, Северная Каролина (2024 г.). «Гигантский тираннозавр из кампана-маастрихта на юге Северной Америки и эволюция гигантизма тираннозаврид » Научные отчеты 13 (1). Артикул 22124. doi : 10.1038/s41598-023-47011-0 . ПМЦ 10784284 . ПМИД 38212342 .
- ^ Сото, М.; Карбаллидо, JL; Лангер, MC; Сильва Джуниор, JCG; Черногория, Ф.; Переа, Д. (2024). «Филогенетические связи нового титанозавра (Dinosauria, Sauropoda) из верхнего мела Уругвая». Меловые исследования . 160 . 105894. Бибкод : 2024CrRes.16005894S . дои : 10.1016/j.cretres.2024.105894 .
- ^ Ван, Шуо; Дин, Нуо; Тан, Цинвэй; Ян, Руи; Чжан, Циюэ; Тан, Лин (17 июля 2024 г.). «Новый урбакодон (Theropoda, Troodontidae) из верхнемеловой формации Ирен-Дабасу, Китай: значение для филогении троодонтид и биологии зубов». Кладистика . дои : 10.1111/cla.12592 . ISSN 0748-3007 . ПМИД 39016633 .
- ^ Лонгрич, Николас Р.; Мартилл, Дэвид М.; Мунт, Мартин; Грин, Мик; Пенн, Марк; Смит, Шон (2024). «Vectidromeus insularis, новый гипсилофодонтидный динозавр из нижнемеловой формации Уэссекс на острове Уайт, Англия» . Меловые исследования . 154 : 105707. Бибкод : 2024CrRes.15405707L . дои : 10.1016/j.cretres.2023.105707 . ISSN 0195-6671 . S2CID 261933503 .
- ^ Цзя, Лэй; Ли, Лиян; Кан, Чжишуай; Сюй, Шичао; «Новый стегозавр из позднего мела Цзоюня». , провинция Шаньси, Китай». Историческая биология : 1–10. doi : 10.1080/08912963.2024.2308214 . ISSN 0891-2963 . S2CID 267465456 .
- ^ Лавгроув, Дж.; Апчерч, П.; Барретт, премьер-министр (2024). «Распутывание дерева или распутывание консенсуса? Последние события в поисках разрешения широкомасштабных отношений внутри Dinosauria» . Журнал систематической палеонтологии . 22 (1). 2345333. Бибкод : 2024JSPal..2245333L . дои : 10.1080/14772019.2024.2345333 .
- ^ Агилар-Педраес, И.; Гарднер, доктор медицинских наук; Орган, CL (2024). «Коэволюция рострального кератина и распределения зубов у динозавров» . Труды Королевского общества B: Биологические науки . 291 (2015). 20231713. дои : 10.1098/rspb.2023.1713 . ПМЦ 10792295 . ПМИД 38229513 .
- ^ Кунц, С; Сакамото, М (2024). «Повышенные темпы эволюции биомеханики кусания открывают закономерности необычайных краниодентальных адаптаций у некоторых травоядных динозавров» . Палеонтология . 67 (1): 12689. Бибкод : 2024Palgy..6712689K . дои : 10.1111/пала.12689 .
- ^ Эркулано-Хаузель, С. (2023). «Динозавры-тероподы имели количество телэнцефальных нейронов, подобное приматам». Журнал сравнительной неврологии . 531 (9): 962–974. дои : 10.1002/cne.25453 . ПМИД 36603059 . S2CID 249994109 .
- ^ Каспар, КР; Гутьеррес-Ибаньес, К.; Бертран, ОК; Карр, Т.; Колбурн, JAD; Эрб, А.; Джордж, Х.; Хольц, ТР; Нэйш, Д.; Уайли, доктор медицинских наук; Херлберт, Греция (2024). «Насколько умен был тираннозавр ? Проверка заявлений об исключительном познании динозавров и применение оценок количества нейронов в палеонтологических исследованиях» . Анатомическая запись . дои : 10.1002/ar.25459 . ПМИД 38668805 .
- ^ Кьяренца, А.А.; Канталапьедра, JL; Джонс, Луизиана; Гамбоа, С.; Гальван, С.; Фарнсворт, AJ; Вальдес, П.Дж.; Сотело, Г.; Варела, С. (2024). «Раннеюрское происхождение птичьей эндотермии и термофизиологического разнообразия динозавров» . Современная биология . 34 (11): 2517–2527.e4. дои : 10.1016/j.cub.2024.04.051 . ПМИД 38754424 .
- ^ Перилло, М.; Сандер, премьер-министр (2024 г.). «Динозавры, которых не было: остеогистология подтверждает родство загадочных крупных сегментов костей европейского рэта с гигантскими ихтиозаврами» . ПерДж . 12 . е17060. дои : 10.7717/peerj.17060 . ПМЦ 11011611 . ПМИД 38618574 .
- ^ Ромилио, А.; Дик, Р.; Скиннер, Х.; Миллар, Дж. (2024). «Обнаружение скрытых следов: ревизия нижнеюрских (синемюрских) отложений Razorback – места, где обнаружены самые ранние следы динозавров в Австралии» . Историческая биология : 1–8. дои : 10.1080/08912963.2024.2320184 .
- ^ Трояно, LP; душ Сантос, HB; Аурелиано, Т.; Гиларди, AM (2024). «Замечательное собрание петроглифов и следов динозавров на северо-востоке Бразилии» . Научные отчеты . 14 (1). 6528. Бибкод : 2024НатСР..14.6528Т . дои : 10.1038/s41598-024-56479-3 . ПМЦ 10948842 . ПМИД 38499621 .
- ^ Хосла, А.; Лукас, СГ (2024). «Триасово-юрские динозавры Индии, их возраст и палеобиогеографическое значение». Историческая биология: Международный журнал палеобиологии : 1–26. дои : 10.1080/08912963.2024.2336992 .
- ^ Мейдмент, ЮКР (2024 г.). «Разнообразие во времени и пространстве в верхнеюрской формации Моррисон, запад США». Журнал палеонтологии позвоночных . е2326027. дои : 10.1080/02724634.2024.2326027 .
- ^ Киркланд, Дж.И.; Сертич, JJW; Титус, Алабама (2024). «Биостратиграфия динозавров неморского мела штата Юта». Геологическое общество, Лондон, специальные публикации . 545 (1): 211. Бибкод : 2024ГСЛСП.545..211К . дои : 10.1144/SP545-2023-211 .
- ^ Аллен, Би Джей; Волкова Оливейра, МВ; Стадлер, Т.; Воган, Т.Г.; Уорнок, RCM (2024). «Механистические филодинамические модели не дают убедительных доказательств того, что нептичьи динозавры находились в упадке перед своим окончательным вымиранием» . Кембриджские призмы: вымирание . 2 . е6. дои : 10.1017/доб.2024.5 . hdl : 20.500.11850/669070 .
- ^ Карри Роджерс, К.; Мартинес, Р.Н.; Коломби, К.; Роджерс, Р.Р.; Алькобер, О. (2024). «Остеогистологическое понимание динамики роста ранних динозавров и их современников» . ПЛОС ОДИН . 19 (4). е0298242. Бибкод : 2024PLoSO..1998242C . дои : 10.1371/journal.pone.0298242 . ПМЦ 10990230 . ПМИД 38568908 .
- ^ Юань, Т.; Сюй, Х.; Цзян, X.; Лю, Ю.; Куанг, Х.; Пэн, Н.; Чен, Дж.; Цен, К. (2024). «Комплексы следов динозавров поздней юры – раннего мела из северо-западной провинции Хэбэй, Китай: значение для палеосреды и палеоэкологии». Меловые исследования . 163 . 105960. дои : 10.1016/j.cretres.2024.105960 .
- ^ Пайо, Винисиус Хосе Маростика; Джуриган, Изабела; Делькур, Рафаэль; де Фариа, Рафаэль Соуза; Батезелли, Алессандро; Рикарди-Бранко, Фрезия (1 августа 2024 г.). «Тафономия и палеогистология ребра динозавра из формации Марилия, группа Бауру, в штате Минас-Жерайс, Бразилия» . Меловые исследования . 160 : 105899. Бибкод : 2024CrRes.16005899P . дои : 10.1016/j.cretres.2024.105899 . ISSN 0195-6671 .
- ^ Пинторе, Р.; Хатчинсон-младший; Бишоп, ПиДжей; Цай, Х.П.; Уссай, А. (2024). «Эволюция морфологии бедренной кости у гигантских нептичьих динозавров-теропод» . Палеобиология . 50 (2): 308–329. Бибкод : 2024Pbio...50..308P . дои : 10.1017/паб.2024.6 . PMC 7616063. PMID 38846629 .
- ^ Дриди, Дж.; Хоула, Ю.; Салхи, И.; Заграрни, МФ (2024). «Свидетельства существования тероподовых динозавров в верхнем готериве – нижнем барреме Джебель-Кебара (центральный Тунис): палеобиогеографические последствия». Журнал африканских наук о Земле . 216 . 105306. Бибкод : 2024JAfES.21605306D . doi : 10.1016/j.jafrearsci.2024.105306 .
- ^ Мезо, Дж.Г.; Джанекини, Ф.; Гомес, КЛ; Муси, Л.; Баяно, Массачусетс; Пол, Д.; Калуза, Дж.; Гарридо, А.; Питтман, М. (2024). «Выпавшие зубы из формации Портесуэло в Сьерра-дель-Портесуэло свидетельствуют о более высоком разнообразии теропод-хищников в туронско-коньякский период в северной Патагонии» . BMC Экология и эволюция . 24 (1). 59. Бибкод : 2024BMCEE..24...59M . дои : 10.1186/s12862-024-02249-8 . ПМЦ 11083846 . ПМИД 38730384 .
- ^ Исасменди, Э.; Перес-Пуэйо, М.; Морено-Азанза, М.; Алонсо, А.; Пуэртолас-Истер, Э.; Баденас, Б.; Канудо, Джи (2024). «Палеоразнообразие зубов теропод из верхнего мела Южно-Пиренейского бассейна (северо-восток Иберии) и внутримаастрихтский оборот фауны» . Меловые исследования . 162 . 105952. Бибкод : 2024CrRes.16205952I . дои : 10.1016/j.cretress.2024.105952 .
- ^ Макларти, Дж.А.; Эсперанте, Р. (2024). «Остановки и повороты: необычно сохранившиеся модели поведения локомотивов теропод на путях верхнего мела из национального парка Тороторо, Боливия» . Журнал южноамериканских наук о Земле . 143 . 105011. Бибкод : 2024JSAES.14305011M . дои : 10.1016/j.jsames.2024.105011 .
- ^ Мохабей, DM; Самант, Б.; Велес-Росадо, К.И.; Уилсон Мантилья, JA (2024). «Обзор мелких динозавров-теропод из верхнего мела Индии с описанием новых черепных останков ноазаврид (Theropoda: Abelisauria)». Журнал палеонтологии позвоночных . 43 (3). е2288088. дои : 10.1080/02724634.2023.2288088 .
- ^ Хендрикс, К.; Трапман, TH; Уиллс, С.; Холверда, FM; Штейн, KHW; Раухут, OWM; Мельцер, Р.Р.; Ван Воэнсель, Дж.; Реймер, JWF (2024). «Комбинированный подход к идентификации изолированных зубов теропод из сеноманской группы Кем-Кем в Марокко: кладистический, дискриминантный анализ и анализ машинного обучения». Журнал палеонтологии позвоночных . 43 (4). е2311791. дои : 10.1080/02724634.2024.2311791 .
- ^ Праделли, Луизиана; Пол, Д.; Эскурра, доктор медицины (2024 г.). «Новые останки динозавров увеличивают разнообразие теропод в формации Каньядон Асфальто (нижняя юра), провинция Чубут, Аргентина». Журнал систематической палеонтологии . 22 (1). 2318262. Бибкод : 2024JSPal..2218262P . дои : 10.1080/14772019.2024.2318262 .
- ^ Делькур, Р.; Бриллиант, Н.С.; Пирес-Домингос, РА; Хендрикс, К.; Грилло, Онтарио; Огаста, Б.Г.; Масиэль, бакалавр наук; Гиларди, AM; Рикарди-Бранко, Ф. (2024). «Биогеография динозавров-теропод в позднем меловом периоде: данные из центральной части Южной Америки» . Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlad184 .
- ^ Рибейро, ТБ; Купелло, К.; де Майринк, Д.; Перейра, PVLGC; Брито, премьер-министр (2024 г.). «Зуб теропода из формации Миссан-Велья (поздняя юра – ранний мел) бассейна Арарипе: старейшая находка бразильских абелизаврид». Историческая биология: Международный журнал палеобиологии : 1–11. дои : 10.1080/08912963.2024.2336982 .
- ^ Аурелиано, Т.; Гиларди, AM; Фонсека, PHM; Мартинелли, АГ; Мариньо, ТС (2024). «Эволюция и диверсификация стратегий роста абелизавроидных теропод». Журнал палеонтологии позвоночных . 43 (3). е2298395. дои : 10.1080/02724634.2023.2298395 .
- ^ Баяно, Массачусетс; Серда, Айова; Бертоццо, Ф.; Пол, Д. (2024). «Новая информация о палеопатологиях нептичьих динозавров-теропод: на примере южноамериканских абелизаврид» . BMC Экология и эволюция . 24 (1). 6. Бибкод : 2024BMCEE..24....6B . дои : 10.1186/s12862-023-02187-x . ПМЦ 10829224 . ПМИД 38291378 .
- ^ Черрони, Массачусетс; Отеро, А.; Новас, FE (2024). «Аппендикулярная миология Skorpiovenator bustingorryi : первая попытка реконструировать мускулатуру таза и задних конечностей теропода-абелизаврид». Анатомическая запись . дои : 10.1002/ar.25532 . ПМИД 38989612 .
- ^ Аурелиано, Т.; Алмейда, В.; Расаона, М.; Гиларди, AM (2024). «Эволюция системы воздушных мешков у динозавров-теропод: данные верхнего мела Мадагаскара». Журнал анатомии . дои : 10.1111/joa.14113 . ПМИД 39022807 .
- ^ Кау, А. (2024). «Единая основа макроэволюции хищных динозавров» (PDF) . Бюллетень Итальянского палеонтологического общества . 63 (1): 1–19. дои : 10.4435/BSPI.2024.08 .
- ^ Монтеалегре, А.; Кастильо-Виза, О.; Селлес, А. (2024). «Новые останки теропода из позднего баррема (раннего мела) восточной части Пиренейского полуострова». Историческая биология: Международный журнал палеобиологии : 1–11. дои : 10.1080/08912963.2024.2308220 . S2CID 267374161 .
- ^ Юн, Чан Гю (2024). «Спинозавры как имитаторы фитозавров: случай конвергентной эволюции двух вымерших клад архозавроформ» . Acta Palaeontologica Romane . 20 (1): 17–29. doi : 10.35463/апреля 2024.01.02 .
- ^ Фаббри, Маттео; Навалон, Гильермо; Бенсон, Роджер Б.Дж.; Пол, Диего; О'Коннор, Цзинмай; Бхуллар, Бхарт-Анжан С.; Эриксон, Грегори М.; Норелл, Марк А.; Оркнейские острова, Эндрю; Ламанна, Мэтью С.; Зухри, Самир; Беккер, Жюстин; Эмке, Аманда; Даль Сассо, Криштиану; Бинделлини, Габриэле; Магануко, Симоне; Аудиторе, Марко; Ибрагим, Низар (31 марта 2022 г.). «Подводное кормление хищных динозавров» . Природы . 603 (7903): 852–857. Бибкод : 2022Natur.603..852F . дои : 10.1038/s41586-022-04528-0 . ISSN 0028-0836 . ПМИД 35322229 . S2CID 247630374 .
- ^ Мирвольд, Натан П.; Баумгарт, Стефани Л.; Видаль, Дэниел; Фиш, Фрэнк Э.; Хендерсон, Дональд М.; Сайтта, Эван Т.; Серено, Пол К. (06 марта 2024 г.). «Ныряющие динозавры? Предостережения относительно использования компактности костей и pFDA для определения образа жизни» . ПЛОС ОДИН . 19 (3): e0298957. Бибкод : 2024PLoSO..1998957M . дои : 10.1371/journal.pone.0298957 . ISSN 1932-6203 . ПМЦ 10917332 . ПМИД 38446841 .
- ^ Смарт, С.; Сакамото, М. (2024). «Использование линейных измерений для диагностики экологической среды обитания спинозавра » . ПерДж . 12 . е17544. дои : 10.7717/peerj.17544 . ПМЦ 11180429 . ПМИД 38881866 .
- ^ Бюффето, Э.; Тонг, Х. (2024). «Первое открытие спинозаврид остается в Азии: Таиланд, 1962 год». Анналы палеонтологии . 110 (1). 102664. Бибкод : 2024AnPal.11002664B . дои : 10.1016/j.annpal.2024.102664 .
- ^ Роу, Эй Джей; Рэйфилд, EJ (2024). «Морфологическая эволюция и функциональные последствия гигантизма у динозавров-тираннозавроидов» . iScience . 110679. дои : 10.1016/j.isci.2024.110679 .
- ^ Ке, Ю.-Х.; Пей, Р.; Сюй, X. (2024). «Данные компьютерной томографии высокого разрешения показывают структуру замены зубов тираннозавроида позднеюрского периода Guanlong wucaii (Dinosauria, Theropoda)» . Vertebrata PalAsiatica : 1. doi : 10.19615/j.cnki.2096-9899.240715 .
- ^ Чочувеч, В.; Маниткун, С.; Чантасит, П.; Кетвецурия, К. (2024). «Первое появление базального тираннозавра в Юго-Восточной Азии: стоматологические данные из верхней юры северо-восточного Таиланда» . Тропическая естественная история . 24 : 84–95.
- ^ Син, Л.; Лян, З.; Чжан, К.; Ван, Д.; Чжан, X.; Лица, WS; Рен, З.; Лян, З.; Сиань, М.; Цзэн, К. (2024). «Большие зубы теропода из верхнего мела провинции Гуандун, Южный Китай». Меловые исследования . 161 . 105914. Бибкод : 2024CrRes.16105914X . дои : 10.1016/j.cretres.2024.105914 .
- ^ ЛеБлан, ARH; Моррелл, AP; Сировица, С.; Аль-Джавад, М.; Лабонте, Д.; Д'Амор, округ Колумбия; Клемент, К.; Ван, С.; Джулиани, Ф.; МакГилвери, CM; Питтман, М.; Кэй, Т.Г.; Стивенсон, К.; Капон, Дж.; Тэпли, Б.; Спиро, С.; Аддисон, О. (2024). «Покрытые железом зубы дракона Комодо и сложная зубная эмаль хищных рептилий» . Экология природы и эволюция : 1–12. дои : 10.1038/ s41559-024-02477-7 ПМИД 39048730 .
- ^ Славяк, Ю.; Брусатте, СЛ; Щигельский, Т. (2024). «Переоценка загадочного динозавра-теропода позднего мелового периода, Bagaraatan ostromi » . Зоологический журнал Линнеевского общества . дои : 10.1093/zoolinnean/zlad169 .
- ^ Юн, К.-Г. (2024). «Профили сил нижней челюсти Алиорамини (Theropoda: Tyrannosauridae) с последствиями для палеоэкологии этой уникальной линии динозавров тираннозаврид» . Летайя . 57 (2): 1–12. Бибкод : 2024Лета..57....1Г . дои : 10.18261/let.57.2.6 .
- ^ Шерер, ЧР; Войкулеску-Холвад, К. (2023). «Повторный анализ набора данных опровергает утверждения об анагенезе у тираннозавров линии Tyrannosaurus (Theropoda, Tyrannosauridae)» . Меловые исследования . 155 . 105780. дои : 10.1016/j.cretres.2023.105780 .
- ^ Уоршоу, EA; Баррера Гевара, Д.; Фаулер, Д.В. (2024). «Анагенез и родословная тирана: ответ на «Повторный анализ набора данных опровергает утверждения об анагенезе внутри тираннозавров линии Tyrannosaurus (Theropoda, Tyrannosauridae) » . Меловые исследования . 163 . 105957. Бибкод : 2024CrRes.16305957W . дои : 10.1016/j.cretres.2024.105957 .
- ^ Лонгрич, Николас Р.; Сайтта, Эван Т. (01 марта 2024 г.). «Таксономический статус Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea) — особого таксона мелких тираннозавров» . Исследования окаменелостей . 2 (1): 1–65. doi : 10.3390/fossils2010001 . eISSN 2813-6284 .
- ^ Мэллон, Джей Си; Хон, DWE (2024). «Оценка максимального размера тела ископаемых видов: тематическое исследование с использованием тираннозавра рекса » . Экология и эволюция . 14 (7). е11658. дои : 10.1002/ece3.11658 . ПМЦ 11267449 . ПМИД 39050661 .
- ^ Самати, А. (2024). «Филогенетическое положение Kinnareimimus khonkaenensis (Dinosauria: Theropoda: Ornithomimosauria) из нижнего мела Таиланда» Зоотаксы 5448 (1): 67–84. дои : 10.11646/zootaxa.5448.1.4 .
- ^ Смит, ДК; Джилетт, Д.Д. (2024). «Остеология производного теризинозавра Nothronychus с доказательствами конвергенции в эволюции динозавров». Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlad148 .
- ^ Парк, Дж.; Сын, М.; Парк, Дж.; Банг, СИ; Ха, Дж.; Мун, Х.; Ли, Ю.-Н.; Ли, С.; Яблонски, П.Г. (2024). «Поведение добычи и эволюция перистого оперения у динозавров» . Научные отчеты . 14 (1). 549. Бибкод : 2024НатСР..14..549П . дои : 10.1038/s41598-023-50225-x . ПМЦ 10811223 . ПМИД 38272887 .
- ^ Киат, Йосеф; О'Коннор, Цзинмай К. (2024). «Функциональные ограничения на количество и форму маховых перьев». Труды Национальной академии наук . 121 (8): e2306639121. Бибкод : 2024PNAS..12106639K . дои : 10.1073/pnas.2306639121 . ПМЦ 10895369. ПМИД 38346196 . S2CID 267634048 .
- ^ Ву, К.; О'Коннор, Дж. К.; Ван, С.; Чжоу, З. (2024). «Трансформация грудного пояса у пеннарапторанов: важные этапы формирования современного плечевого сустава птиц» . ПерДж . 12 . е16960. дои : 10.7717/peerj.16960 . ПМЦ 10909347 . ПМИД 38436017 .
- ^ Мид, Луизиана; Питтман, М.; Баланов А.; Лаутеншлагер, С. (2024). «Краниальная функциональная специализация по силе предшествует морфологической эволюции овирапторозаврии» . Коммуникационная биология . 7 (1). 436. дои : 10.1038/s42003-024-06137-1 . ПМЦ 11006937 . ПМИД 38600295 .
- ^ Фанстон, Г.Ф.; Уильямсон, TE; Брусатте, СЛ (2024 г.). «Большая кость ценагнатида (Theropoda: Oviraptorosauria) из верхней кампанской формации Киртланд в Нью-Мексико». Меловые исследования . 158 . 105856. Бибкод : 2024CrRes.15805856F . дои : 10.1016/j.cretres.2024.105856 . S2CID 267601272 .
- ^ Цю, Р.; Ду, Ю.; Хуанг, З.; Чжу, X.; Ян, X.; Ван, К.; Ван, X. (2024). «Остеология запястья Heyuannia huangi (Oviraptorosauria) и ее значение для механизма складывания запястья» . ПерДж . 12 . е17669. дои : 10.7717/peerj.17669 .
- ^ Фанстон, Г.Ф. (2024). «Остеология двупалого овирапторида Оксоко аварсан (Theropoda: Oviraptorosauria)» . Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlae011 .
- ^ Чжу, X.; Ван, К.; Ван, X. (2024). «Исследование дифракции обратного рассеяния электронов (EBSD) яиц элонгатоолитид из Китая с микроструктурными и паратаксономическими последствиями». Палеобиология . 50 (2): 330–345. Бибкод : 2024Pbio...50..330Z . дои : 10.1017/паб.2024.9 .
- ^ Це, Ю.Т.; Миллер, резюме; Питтман, М. (2024). «Морфологическое несоответствие и структурные характеристики черепа дромеозаврид играют важную роль в экологии и истории эволюции» . BMC Экология и эволюция . 24 (1). 39. Бибкод : 2024BMCEE..24...39T . дои : 10.1186/s12862-024-02222-5 . ПМК 11020771 . ПМИД 38622512 .
- ^ Ву, Р.; Ню, К.; Чжан, С.; Сюэ, Ю.; Хан, Ф. (2024). «Новый оотип предполагаемых яиц дромеозаврид из верхнего мела южного Китая». Меловые исследования . 161 . 105909. Бибкод : 2024CrRes.16105909W . дои : 10.1016/j.cretres.2024.105909 .
- ^ Джанекини, Ф.А.; Колли, Л.; Маковицкий, П.Дж. (2024). «Реконструкция мышц таза и задних конечностей паравианского теропода Buitreraptor gonzalezorum и ее палеобиологические последствия». Историческая биология: Международный журнал палеобиологии : 1–27. дои : 10.1080/08912963.2023.2301674 . S2CID 266994137 .
- ^ Манафзаде, Армита Р.; Гейтси, Стивен М.; Бхуллар, Бхарт-Анжан С. (2024). «Взаимодействие суставных поверхностей отличает позы локомоторных суставов динозавров» . Природные коммуникации . 15 (1). 854. Бибкод : 2024NatCo..15..854M . дои : 10.1038/s41467-024-44832-z . ПМЦ 10873393 . ПМИД 38365765 .
- ^ Ю, Ц.; Ватанабэ, А.; Цинь, З.; Кинг, Дж.Л.; Уитмер, Л.М.; Ма, Кью; Сюй, X. (2024). «Авиаланоподобная морфология мозга Sinovenator (Troodontidae, Theropoda)» . Коммуникационная биология . 7 (1). 168. дои : 10.1038/s42003-024-05832-3 . ПМЦ 10858883 . ПМИД 38341492 .
- ^ Син, Л.; Ню, К.; Локли, МГ; Ромилио, А.; Дэн, К.; Лица, WS (2024). «Следы дейнонихозавров на юго-востоке Китая фиксируют, возможно, гигантского троодонтида» . iScience . 27 (5). 109598. Бибкод : 2024iSci...27j9598X . дои : 10.1016/j.isci.2024.109598 . ПМЦ 11123545 . ПМИД 38799075 .
- ^ СИЛЬВА, ФО; МАРТИНЕЛЛИ, АГ; ПРЕТТО, Ф.; Фериголо, Дж.; Рибейро, AM (2024). «Первые зауроподоморфы (Dinosauria) на стоянке Вила-Ботукарай ( зона комплекса Hyperodapedon , последовательность Канделария), Риу-Гранди-ду-Сул, Бразилия». Журнал южноамериканских наук о Земле . 140 . 104927. Бибкод : 2024JSAES.14004927O . дои : 10.1016/j.jsames.2024.104927 .
- ^ Регаладо Фернандес, Орегон (2024 г.). «Изменчивость позвоночных пластинок у восьми экземпляров платеозавра (Saurischia, Sauropodomorpha)» . Revue de Paléobiologie, Женева . 43 (1): 85–100.
- ^ Шеффер, Дж. (2024). «Остеологическое переописание голотипа Plateosaurus trossingensis (Dinosauria: Sauropodomorpha) из верхнего триаса юго-западной Германии и его филогенетическое значение» . Журнал систематической палеонтологии . 22 (1). 2335387. Бибкод : 2024JSPal..2235387S . дои : 10.1080/14772019.2024.2335387 .
- ^ Шеффер, Дж.; Вольф, Э.; Витцманн, Ф.; Феррейра, Г.С.; Шох, Р.Р.; Мухал, Э. (2024). «Палеобиологические последствия патологии шеврона у зауроподоморфа Plateosaurus trossingensis из верхнего триаса на юго-западе Германии» . ПЛОС ОДИН . 19 (7). е0306819. дои : 10.1371/journal.pone.0306819 . ПМЦ 11290664 . ПМИД 39083447 .
- ^ Чжао, Р.; Чжан, С.; Ты, Х.; Ван, Ю.; Чжан, К.; Ван, Т. (2024). «Новый экземпляр ранневетвящегося динозавра-зауроподоморфа из бассейна Чуаньцзе, Луфэн, провинция Юньнань» . Acta Palaeontologica Sinica . 63 (1): 102–111. doi : 10.19800/j.cnki.aps.2023030 .
- ^ Ван, Ю.-М.; Чжао, К.; Вы, Х.-Л. (2024). «Повторная оценка Gyposaurus Young, 1941 (Dinosauria: Sauropodomorpha) из sinensis . раннеюрского бассейна Луфэн, провинция Юньнань, Китай» Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlae032 .
- ^ Барретт, премьер-министр; Шуаньер, Ж.Н. (2024). « Melanorosaurus readi Haughton, 1924 (Dinosauria, Sauropodomorpha) из позднего триаса Южной Африки: остеология и обозначение лектотипа». Журнал палеонтологии позвоночных . е2337802. дои : 10.1080/02724634.2024.2337802 .
- ^ Видаль, Д. (2024). «Виртуальные зауроподы: революция в изучении длинношеих динозавров» . Журнал научных исследований Metode . 14 : 77–83. дои : 10.7203/metode.14.24689 .
- ^ Августин Дж.; Алькала, Л.; Сантос-Кубедо, А. (2024). «Способствовали ли крупные миграции в поисках пищи огромным размерам тела гигантских зауроподов? Случай с туриазавром » . Испанский журнал палеонтологии . 39 (1): 103–110. дои : 10.7203/sjp.28176 .
- ^ Батлер, Р.Дж.; Эдгар, К.М.; Халлер, Л.; Мид, Луизиана; Джонс, ХТ; Хилл, О.; Скривен, С.; Ридман, К. (2024). «Следы динозавров зауроподов из группы Пурбек (ранний мел) карьера Спайуэй, Дорсет, Великобритания» . Королевское общество открытой науки . 11 (7). 240583. Бибкод : 2024RSOS...1140583B . дои : 10.1098/rsos.240583 . ПМЦ 11285821 . ПМИД 39076363 .
- ^ Буасверт, К.; Кертис, Б.; Ведель, М.; Уилхайт, Р. (2024). «Описание нового экземпляра гаплокантозавра из карьера динозавров Сухой Мезы» . Анатомическая запись . дои : 10.1002/ar.25520 . ПМИД 38887924 .
- ^ Кинг, Дж.Л.; Макхью, Дж. Б.; Ведель, MJ; Кертис, Б. (2024). «Ранее не сообщавшаяся форма пневматическости спинных ребер у апатозавра (Dinosauria: Sauropoda) и последствия для пневматических вариаций среди диплодоцидных спинных ребер». Журнал палеонтологии позвоночных . е2316665. дои : 10.1080/02724634.2024.2316665 .
- ^ Виндхольц, Дж.Дж.; Порфирий, JD; Дос Сантос, Д.; Беллардини, Ф.; Ведель, MJ (2024). «Хорошо сохранившийся позвонок дает новое представление о каудальных анатомических и пневматических особенностях реббахизаврид-зауроподов» . Acta Palaeontologica Polonica . 69 (1): 39–47. дои : 10.4202/app.01104.2023 .
- ^ Гомес, КЛ; Перес-Морено, А.; Мезо, Дж.Г.; Беллардини, Ф.; Баяно, Массачусетс; Пол, Д.; Гарридо, А.; Калуза, Дж.; Муси, Л.; Питтман, М. (2024). «Раскрытие разнообразия зауропод в формации Портесуэло в Патагонии посредством всестороннего анализа нового и существующего материала» . BMC Экология и эволюция . 24 (1). 96. Бибкод : 2024BMCEE..24...96G . дои : 10.1186/s12862-024-02280-9 . ПМЦ 11234639 . ПМИД 38982364 .
- ^ Бистон, СЛ; Поропат, Сан-Франциско; Мэннион, Полиция; Пентленд, штат Айдахо; Энчелмайер, MJ; Слоан, Т.; Эллиотт, Д.А. (2024). «Переоценка разнообразия динозавров-зауроподов в верхнемеловой формации Винтон в Квинсленде, Австралия, посредством 3D-оцифровки и описания новых экземпляров» . ПерДж . 12 . е17180. дои : 10.7717/peerj.17180 . ПМК 11011616 . ПМИД 38618562 .
- ^ Филиппи, Л.С.; Беллардини, Ф.; Карбаллидо, Дж.Л.; Перес-Морено, А.; Гарридо, AC (2024). «Разнообразие зауропод (Dinosauria: Sauropoda) Серро Оверо – Ла Инвернада (формация Бахо-де-ла-Карпа, сантон), северо-восточной части бассейна Неукен, и палеоэкологические последствия для фауны зауропод верхнего мела» . Электронное издание Аргентинской палеонтологической ассоциации . 24 (1): 71–96. дои : 10.5710/PEAPA.24.02.2024.484 .
- ^ Филип, Л.С.; Превитера, Э.; Гарридо, AC (2024). « Кайютитан мауи , титанозавр-зауропод из верхнего мела (формация Сьерра-Барроса, бассейн Неукен) северной Патагонии, Аргентина: гистологические и тафономические соображения» . Электронное издание Аргентинской палеонтологической ассоциации . 24 (1): 129–148. дои : 10.5710/PEAPA/26.02.2024.481 .
- ^ Видаль, Л.С.; Бергквист, LP; Кандейро, CRA; Флаг, КЛН; Таварес, С.; Брусатте, СЛ; Перейра, ПВЛГВ (2024 г.). «Осевая биомеханика Trigonosaurus Pricei (Neosauropoda: Titanosauria) и важность шейно-спинной области для стратегии кормления зауроподов с высоким уровнем браузера». Зоологический журнал Линнеевского общества . 201 (3). zlae087. doi : 10.1093/zoolinnean/zlae087 .
- ^ Зурриагуз, В.Л. (2024). «Изменение посткраниальной пневматичности у производных титанозавров (Dinosauria: Sauropoda)». Историческая биология: Международный журнал палеобиологии : 1–15. дои : 10.1080/08912963.2024.2377708 .
- ^ Мочо, П.; Перес-Гарсия, А.; Кодря, Вирджиния (2024 г.). «Новые останки аппендикуляров зауроподов из верхнего мела Румынии: доступ к морфологической изменчивости» . Меловые исследования . 163 . 105936. Бибкод : 2024CrRes.16305936M . дои : 10.1016/j.cretres.2024.105936 .
- ^ Кальво, Дж. О. (2024). «Какой самый гигантский зауропод из Аргентины? Разнообразие крупных титанозавров из Патагонии» . Журнал научных исследований Metode . 14 (14): 65–75. дои : 10.7203/metode.14.24556 .
- ^ Фонсека, АО; Рид, Ай-Джей; Веннер, А.; Дункан, Р.Дж.; Гарсия, MS; Мюллер, RT (2024). «Всесторонний филогенетический анализ ранней птицетазовой эволюции». Журнал систематической палеонтологии . 22 (1). 2346577. Бибкод : 2024JSPal..2246577F . дои : 10.1080/14772019.2024.2346577 .
- ^ Уиллс, С.; Андервуд, CJ; Барретт, премьер-министр (2024). «Скрытое разнообразие птицетазовых динозавров: фауна микропозвоночных средней юры Великобритании проливает свет на плохо представленный период». Журнал палеонтологии позвоночных . е2323646. дои : 10.1080/02724634.2024.2323646 .
- ^ Ху, Дж.; Сюй, Х.; Ли, Ф.; Хан, Ф (2024). «Замена зубов у рано расходящихся неорнитисхий Jeholosaurus shangyuanensis и последствия для эволюции зубов и травоядной адаптации у Ornithischia» . BMC Экология и эволюция . 24 (1). 46. Бибкод : 2024BMCEE..24...46H . дои : 10.1186/s12862-024-02233-2 . ПМК 11020315 . ПМИД 38627692 .
- ^ Круменакер, LJ; Варриккио, диджей; Орган, К.; Гарднер, доктор медицинских наук; Бритт, BB; Бойд, К. (2024). «Остеология и филогенетические взаимоотношения неорнитишского динозавра среднего мела Oryctodromeus Cubularis Varricchio, 2007». Журнал палеонтологии позвоночных . е2330581. дои : 10.1080/02724634.2024.2330581 .
- ^ Чепинский, Л.; Мадзия, Д. (2024). «Остеология, филогенетическое сходство и палеобиогеографическое значение причудливого птицетазового динозавра Ajkaceratops kozmai с Европейского архипелага позднего мела». Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlae048 .
- ^ Ли, Сан Юн; Ли, Юнг-Нам; Юнг, Сын Хо (1 июня 2024 г.). «Неорнитишская фаланга из нижнемеловых отложений тандо Ансан-си, Кёнгидо, Южная Корея». Журнал Геологического общества Кореи . 60 (2): 135–142. дои : 10.14770/jgsk.2024.010 .
- ^ Сатчелл, КГ (2024 г.). «Переоценка остатков сцелидозавра из Ирландии и важность идентификации на основе апоморфии». Труды Ассоциации геологов . 135 (3): 349–351. Бибкод : 2024ПрГА..135..349С . дои : 10.1016/j.pgeola.2024.03.002 .
- ^ Кастанера, Д.; Мампель, Л.; Кобос, А. (2024). «Сложность отслеживания стегозавров и их стадное поведение» . Научные отчеты . 14 (1). 14833. Бибкод : 2024NatSR..1414833C . дои : 10.1038/s41598-024-64298-9 . ПМЦ 11222438 . ПМИД 38961126 .
- ^ Санчес-Феноллоса, С.; Эскасо, Ф.; Кобос, А. (2024). «Новый экземпляр Dacentrurus Armatus Owen, 1875 (Ornithischia: Thyreophora) из верхней юры Испании и его таксономическое значение в разнообразии европейских стегозавров». Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlae074 .
- ^ Латегано, Ф.; Конти, С.; Лозар, Ф. (2024). « Биомеханика и защита хвоста мирагайя . Оценка подвижности хвоста и устойчивости к нагрузкам и столкновениям». Rivista Italiana di Paleontologia e Stratigrafia . 130 (2): 475–486. doi : 10.54103/2039-4942/21688 (неактивен 30 июля 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на июль 2024 г. ( ссылка ) - ^ Ли, Н.; Ли, Д.; Пэн, Г.; Ты, Х. (2024). «Первый стегозавровый динозавр из провинции Ганьсу, Китай». Меловые исследования . 158 . 105852. Бибкод : 2024CrRes.15805852L . дои : 10.1016/j.cretres.2024.105852 . S2CID 267286799 .
- ^ Кросс, ЕС; Арбур, ВМ (2024). «Бедренная кость анкилозавра из середины мела региона Мира на северо-востоке Британской Колумбии». Канадский журнал наук о Земле . 61 (6): 678–685. Бибкод : 2024CaJES..61..678C . doi : 10.1139/cjes-2023-0118 . S2CID 267961368 .
- ^ Сото Коин, Серджио; Варгас, Александр О.; Калуза, Джонатан (2024). « Antarctopelta Oliveroi (Ankylosauria: Parankylosauria) » Достижения полярной науки . 35 (1): 78–107. дои : 10.12429/j.advps.2023.0036 .
- ^ Сантос-Кубедо, А. (2024). «Динозавры бассейна Маэстрат: эволюция гадрозавриформ на востоке Пиренейского полуострова» . Журнал научных исследований Metode . 14 : 57–63. дои : 10.7203/metode.14.24534 . hdl : 10234/207308 .
- ^ Эсканеро-Агилар, Д.; Торсида Фернандес-Бальдор, Ф.; Переда-Субервиола, X.; Уэрта, П. (2024). «Материал черепа раннемелового гадрозавриформного динозавра (Ornithopoda) из Салас-де-лос-Инфантес (Бургос, Испания)». Журнал иберийской геологии . 50 (1): 67–82. Бибкод : 2024JIbG...50...67E . дои : 10.1007/s41513-023-00227-5 .
- ^ Хаяши, С.; Накадзима, Ю.; Танака, Ю.; Бриден, Британская Колумбия; Канадзава, Ю.; Цогтбаатар, К. (2024). «Гадрозавроидный позвонок из верхнемеловой группы Идзуми, префектура Кагава, Япония». Палеонтологические исследования . 28 (4): 442–451. дои : 10.2517/PR230027 .
- ^ Николов Владимир; Дочев, Дочо; Брусатте, Стивен Л. (1 января 2024 г.). «Онтогенетический статус небольшого гадрозавроидного динозавра из верхнего мелового периода Болгарии и его значение для палеобиогеографии и сборки европейской островной фауны» . Меловые исследования . 157 . 105819. Бибкод : 2024CrRes.15705819N . дои : 10.1016/j.cretres.2023.105819 . S2CID 266738171 .
- ^ ван дер Линден, ТТП; Зеленицкий, ДК; Фраайе, RHB; Гарсия, Г.; Валентин, X.; Холверда, FM; Шульп, А.С. (2024). «Первая яичная скорлупа гадрозавроидов из бассейна Экс-ан-Прованс (поздний маастрихт) во Франции». Историческая биология: Международный журнал палеобиологии : 1–8. дои : 10.1080/08912963.2024.2380808 .
- ^ Марк Дж. Пауэрс; Мэтью М. Роудс; Аарон Д. Дайер; Стивен Э. Мендонка; Райан Уилкинсон; Майкл Нейлор Хаджинс; Мэтью Дж. Пруден; Филип Дж. Карри; Грегори Ф. Фанстон (2024). «Первая тропа гадрозаврид из формации Каньон Подкова (кампан / Маастрихт) в Альберте, Канада». Ихнос . 30 (3): 179–206. дои : 10.1080/10420940.2024.2307069 . S2CID 267489605 .
- ^ Варнок-Жуто, ТМ; Райан, MJ; Паттерсон, RT; Мэллон, Джей Си (2024). «Компьютерная томография черепа вылупившегося детеныша выявляет онтогенетические изменения в морфологии зубных рядов и окклюзионной поверхности Hadrosauridae (Dinosauria: Ornithischia)» . Анатомия позвоночных Морфология Палеонтология . 11 : 133–150. дои : 10.18435/vamp29395 .
- ^ Шарп, HS; Пауэрс, MJ; Дайер, AD; Родос, ММ; Макинтош, AP; Гаррос, CW; Карри, Пи Джей; Фанстон, Г.Ф. (2024). «Краниомандибулярная анатомия молодого экземпляра Edmontosaurus regalis Lambe, 1917 проясняет вопросы онтогенеза и биогеографии». Журнал палеонтологии позвоночных . е2326644. дои : 10.1080/02724634.2024.2326644 .
- ^ Подлеснов А.В.; Аверьянов, АО; Бурухин А.А.; Феофанова О.А.; Владимирова, ОН (2024). «Новые данные о морфологии черепа Psittacosaurus sibiricus (Dinosauria: Ceratopsia) с использованием микрокомпьютерной томографии». Палеонтологический журнал . 57 (10): 1128–1187. дои : 10.1134/S0031030123100040 . S2CID 267537898 .
- ^ Сакагами, Р.; Кавабе, С.; Хаттори, С.; Чжэн, В.; Джин, X. (2024). «Эндокраниальная анатомия цератопсового динозавра Psittacosaurus lujiatunensis и ее влияние на сенсорные и двигательные способности» (PDF) . Мемуары Музея динозавров префектуры Фукуи . 22 : 1–12.
- ^ Ян, З.; Цзян, Б.; Сюй, Дж.; Макнамара, Мэн (2024). «Клеточная структура чешуи динозавров свидетельствует о сохранении кожи типа рептилий во время эволюционного перехода к перьям» . Природные коммуникации . 15 (1). 4063. Бибкод : 2024NatCo..15.4063Y . дои : 10.1038/s41467-024-48400-3 . ПМЦ 11109146 . ПМИД 38773066 .
- ^ Уиттон, член парламента; Хинг, РА (2024). «Рогатый динозавр Протоцератопс вдохновил грифона?». Междисциплинарные научные обзоры . doi : 10.1177/03080188241255 (неактивен 23 июня 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на июнь 2024 г. ( ссылка ) - ^ Баррера Гевара, член парламента; Эспиноза Чавес, Б.; Серрано Браньяс, CI; де Леон Давила, К.; Посада Мартинес, Д.; Фридман Фаулер, Э.; Фаулер, Д. (2024). «Стратиграфическая переоценка мексиканского хасмозавра Coahuilaceratops magnacuerna как первых диагностических остатков динозавра из формации Серро-Уэрта (нижний маастрихт), подтверждающая южное происхождение трицератопсини» . Разнообразие . 16 (7). 390. дои : 10.3390/d16070390 .
- ^ Теннисон, AJD; Сальвадор, РБ; Томотани, Б.М.; Маркс, ФГ (2024). «Новый ныряющий плиоценовый буревестник Ardenna (Aves: Procellariidae) из Новой Зеландии» . Таксономия . 4 (2): 237–249. дои : 10.3390/taxonomy4020012 . hdl : 10037/33366 .
- ^ Аньолин, Флорида; Альварес Эррера, терапевт; Томассини, Р. (2024). «Плейстоценовая находка Chloephaga Eyton, 1838 (Anseriformes: Anatidae) в аргентинских пампасах с описанием нового вида» . Comptes Рендус Палевол . 23 (18): 241–255. doi : 10.5852/cr-palevol2024v23a18 .
- ^ Зеленков, Н.В. (2024). «Серые куропатки (Phasianidae: Genera Perdix и Enkuria gen. nov.) из раннего плейстоцена Крыма и замечания об эволюции рода Perdix ». Палеонтологический журнал . 58 (3): 335–352. Бибкод : 2024PalJ...58..335Z . дои : 10.1134/S0031030124700084 .
- ^ Перейти обратно: а б с Майр, Г.; Китченер, AC (2024). «Новые окаменелости Eocypselus и Primapus из британской лондонской глины демонстрируют высокое таксономическое и экологическое разнообразие раннеэоценовых быстроподобных аподиформных птиц» . Ibis (предварительная онлайн-публикация). дои : 10.1111/ibi.13323 .
- ^ Перейти обратно: а б Майр, Джеральд; Китченер, Эндрю К. (07.06.2024). «Неаподиформные стризоры (поту, козодои и родственные птицы) из раннеэоценовой лондонской глины Уолтон-на-Нейзе» . Палеобиоразнообразие и палеосреда . Бибкод : 2024PdPe..tmp...21M . дои : 10.1007/s12549-024-00610-9 . ISSN 1867-1594 .
{{cite journal}}: CS1 maint: bibcode ( ссылка ) - ^ Ван, Сяоли; Кларк, Александр Д.; О'Коннор, Цзинмай К.; Чжан, Сянъюй; Ван, Син; Чжэн, Сяотин; Чжоу, Чжунхэ (27 февраля 2024 г.). «Первый беззубый энантиорнитин (Aves: Ornithothoraces) из нижнемеловой авифауны Джехол» . Меловые исследования . 159 (в печати): 105867. Бибкод : 2024CrRes.15905867W . дои : 10.1016/j.cretres.2024.105867 . ISSN 0195-6671 . S2CID 268103010 .
- ^ Перейти обратно: а б с Зеленков, Никита (2024). «Замечательное разнообразие водоплавающих птиц (Aves: Anseriformes) верхнего эоцена и нижнего олигоцена Казахстана». Журнал палеонтологии позвоночных . е2374306. дои : 10.1080/02724634.2024.2374306 .
- ^ Перейти обратно: а б с Майр, Джеральд; Китченер, Эндрю К. (2024). «Месселорнитиды и мессселорнитидоподобные птицы из ранней эоценовой лондонской глины Уолтон-он-Наз (Эссекс, Великобритания)» . Геобиос (в печати). дои : 10.1016/j.geobios.2023.12.011 . ISSN 0016-6995 .
- ^ Мюссер, Г.; Кларк, Дж. А. (2024). «Новая окаменелость палеогена и новый набор данных по водоплавающим птицам (Aves: Anseriformes) проясняют филогению, экологическую эволюцию и эволюцию птиц на границе K-Pg» . ПЛОС ОДИН . 19 (7). е0278737. дои : 10.1371/journal.pone.0278737 . ПМЦ 11288464 . ПМИД 39078833 .
- ^ Андо, Тацуро; Робинсон, Джеффри; Лох, Каролина; Накахара, Тамон; Хаяси, Сёдзи; Ричардс, Маркус Д.; Фордайс, Роберт Юэн (31 июля 2024 г.). «Новый крошечный ископаемый пингвин из позднего олигоцена Новой Зеландии и морфофункциональный переход крыла пингвина» . Журнал Королевского общества Новой Зеландии : 1–22. дои : 10.1080/03036758.2024.2362283 . ISSN 0303-6758 .
- ^ Перейти обратно: а б Зеленков, Н.В. (2024). «Тетерев (Aves: Phasianidae: Tetraonini) из раннего плейстоцена Крыма и таксономический статус Lagopus atavus » . Палеонтологический журнал . 58 (1): 112–123. Бибкод : 2024PalJ...58..112Z . дои : 10.1134/S0031030124010106 .
- ^ Перейти обратно: а б Майр, Г.; Китченер, AC (2024). «Курчатые птицы из нижнего эоцена Лондонской глины Уолтон-он-Наз (Эссекс, Великобритания): новые виды предполагают фаунистические связи с Азией» . Журнал палеонтологии позвоночных . е2374305. дои : 10.1080/02724634.2024.2374305 .
- ^ Хорват, И.; ФУТО, Дж.; Кесслер, Дж. (2024). « Phalacrocorax bakonyiensis n. sp., новый вид баклана из позднего миоцена Венгрии» . Орнис Хунгарика . 32 : 222–230. дои : 10.2478/orhu-2024-0016 .
- ^ Перейти обратно: а б с Майр, Г.; Китченер, AC (2024). «Пикокорациады (удоды, ролики, дятлы и их союзники) из раннего эоцена лондонской глины Уолтон-на-Назе» . ПалЗ . 98 (2): 291. Бибкод : 2024PalZ...98..291M . дои : 10.1007/s12542-024-00687-9 .
- ^ Горбачев, ВО; Зеленков, Н.В. (2024). «Стервятник рода Торгос (Птицы: Accipitridae) в позднем плейстоцене Азербайджана» Палеонтологический журнал . 58 (4): 475–482. дои : 10.1134/S0031030124600367 .
- ^ Де Мендоса, Рикардо Сантьяго; Дегранж, Федерико Хавьер; Тамбусси, Клаудия Патрисия (2024). «Оценка гусеобразного сходства «Telmabates» Howardae » . Журнал южноамериканских наук о Земле . 135 . 104786. Бибкод : 2024JSAES.13504786D . дои : 10.1016/j.jsames.2024.104786 . S2CID 267159455 .
- ^ Баланофф, Эми; Феррер, Элизабет; Салех, Лемизе; Жиньяк, Поль М.; Голд, М. Евгения Л.; Маруган-Лобон, Хесус; Норелл, Марк; Уэллетт, Дэвид; Салерно, Майкл; Ватанабэ, Акинобу; Вэй, Шоуи; Бевер, Габриэль; Васька, Павел (2024). «Количественная функциональная визуализация мозга голубя: значение для эволюции полета птиц». Труды Королевского общества B: Биологические науки . 291 (2015). дои : 10.1098/rspb.2023.2172 . ПМЦ 10827418. ПМИД 38290541 .
- ^ Чжоу, Чжунхэ; Ван, Мин (2024). «Меловые ископаемые птицы из Китая». Геологическое общество, Лондон, специальные публикации . 544 (1). дои : 10.1144/SP544-2023-129 . S2CID 267184802 .
- ^ Ван, X.; О'Коннор, Дж.; Чжэн, X.; Ван, Ю.; Киат, Ю. (2024). «Самые ранние свидетельства первичной линьки перьев птиц». Письма по биологии . 20 (7). 20240106. дои : 10.1098/rsbl.2024.0106 . ПМК 11285806. ПМИД 38955226 .
- ^ Чжоу, Ю.; Пан, Ю.; Ван, М.; Ван, X.; Чжэн, X.; Чжоу, З. (2024). «Ископаемые свидетельства проливают свет на половой отбор на ранней стадии эволюции птиц» . Труды Национальной академии наук Соединенных Штатов Америки . 121 (3). e2309825120. Бибкод : 2024PNAS..12109825Z . дои : 10.1073/pnas.2309825120 . ПМЦ 10801838 . ПМИД 38190528 . S2CID 266871669 .
- ^ Филд, Дэниел Дж.; Бенито, Хуан; Вернинг, Сара; Чен, Альберт; Куо, Пей-Чен; Крейн, Аби; Видриг, Клара Э.; Ксепка, Дэниел Т.; Джагт, Джон ВМ (2024). «Замечательное понимание происхождения современных птиц из области маастрихтского типа (северо-восток Бельгии, юго-восток Нидерландов)». Нидерландский журнал геонаук . 103 . Бибкод : 2024NJGeo.103E..15F . дои : 10.1017/njg.2024.11 .
- ^ Стойческу, В.; Кодря, Вирджиния; Бордеян, М.; Соломон, А.А. (2024). « Элоптерикс в Налац-Ваде: новый материал теропод, описанный из бассейна Хацег (Румыния)» . Северо-Западный зоологический журнал . 20 (1): 73–80.
- ^ Лю, Б.-Ю.; Стидхэм, штат Техас; Ван, Х.-П.; Ли, З.-Х.; Чжоу, З.-Х. (2024). «Морфометрический анализ ряда шейных позвонков современных птиц с учетом экологии питания мезозойских птиц» . Позвоночные Палазиатские . 62 (2): 99–119. doi : 10.19615/j.cnki.2096-9899.240305 .
- ^ Миллер, Кейс Винсент; Брайт, Джен А; Ван, Сяоли; Чжэн, Сяотин; Питтман, Майкл (2024). «Синтетический анализ трофического разнообразия и эволюции Enantiornithes с новыми данными о Bohaiornithidae» . электронная жизнь . 12 . РП89871. дои : 10.7554/eLife.89871.3 . ПМК 11060716 . ПМИД 38687200 .
- ^ Кундрат, М.; Хорват, Д.; Ван, З.; Ван, X. (2024). «Распределение лакун остеоцитов в области развития в коре костей конечностей Musivavis amabilis с обзором адаптации микроструктуры кости у Enantiornithes». Меловые исследования . 158 . 105839. Бибкод : 2024CrRes.15805839K . дои : 10.1016/j.cretres.2024.105839 . S2CID 267250890 .
- ^ Акоста Госпиталече, Каролина; Ирасоки, Факундо; Бона, Паула; Паулина-Карабахал, Ариана (2024). «Обзор мелового разнообразия птиц Антарктиды: меняющийся сценарий эволюции ранних неорнитинских птиц». Достижения полярной науки . 35 (1): 1–13. дои : 10.12429/j.advps.2023.0025 .
- ^ Броклхерст, Н.; Филд, диджей (2024). «Первичное датирование и факторы Байеса дают представление о расхождениях кладов коронованных птиц во время массового вымирания в конце мелового периода» . Труды Королевского общества B: Биологические науки . 291 (2016). 20232618. дои : 10.1098/rspb.2023.2618 . ПМК 10865003 . ПМИД 38351798 .
- ^ Берв, Дж.С.; Сингхал, С.; Филд, диджей; Уокер-Хейл, Н.; Макхью, Юго-Запад; Шипли, младший; Миллер, ET; Кимбалл, RT; Браун, Эль; Дорнбург, А.; Паринс-Фукучи, Коннектикут; Прум, РОД; Вингер, Б.М.; Фридман, М.; Смит, Ю.А. (2024 г.). «Геном и эволюция жизненного цикла связывают диверсификацию птиц с массовым вымиранием в конце мелового периода» . Достижения науки . 10 (31): eadp0114. дои : 10.1126/sciadv.adp0114 . ПМЦ 11290531 . ПМИД 39083615 .
- ^ Видриг, Кентукки; Навалон, Г.; Филд, диджей (2024). «Палеоневрология стволовых палеогнатов проясняет плезиоморфное состояние центральной нервной системы кроновых птиц» . Журнал морфологии . 285 (6). е21710. дои : 10.1002/jmor.21710 . ПМИД 38760949 .
- ^ Ву, Цянь; Пан, Ян-Хонг; Ли, Чжи-Хэн; Чжоу, Чжун-Хе; Байёль, Алида М. (2024). «Первое гистохимическое исследование скорлупы страусиного яйца миоцена с древнейшими пептидами, связанными с минералами». Позвоночные Палазиатские . 62 (2): 120–134. doi : 10.19615/j.cnki.2096-9899.240329 .
- ^ Шретер, Э. (2024). «Характеристика диагенетиформ в расширенном протеоме вымерших моа (Dinornithidae): идентификация биологических, диагенетических, экспериментальных артефактов и неправильно маркированных модификаций в деградировавших тканях» . Минералы . 14 (2): 137. Бибкод : 2024Мой...14..137С . дои : 10.3390/мин14020137 .
- ^ Пикфорд, М. (2024). «Таксономический пересмотр вымерших видов птиц Diamantornis karingarabensis (Senut et al. , 1998) из позднего миоцена Намибии» (PDF) . Сообщения Геологической службы Намибии . 27 : 66–71.
- ^ Эдвардс, Скотт В.; Клотье, Элисон; Кокберн, Гленн; Драйвер, Роберт; Грейсон, Фил; Като, Казутака; Болдуин, Мод В.; Сактон, Тимоти Б.; Бейкер, Аллан Дж. (2024). «Сборка ядерного генома вымершей нелетающей птицы, маленького моа» . Достижения науки . 10 (21): eadj6823. Бибкод : 2024SciA...10J6823E . дои : 10.1126/sciadv.adj6823 . ПМИД 38781323 .
- ^ Томлинсон, С.; Ломолино, МВ; Вуд, младший; Андерсон, А.; Браун, Южная Каролина; Хейторн, С.; Перри, GLW; Уилмшерст, Дж. М.; Остин, Джей-Джей; Фордхэм, Д.А. (2024 г.). «Экологическая динамика вымирания моа обнаруживает конвергентные рефугиумы, в которых сегодня обитают нелетающие птицы». Экология природы и эволюция : 1–10. дои : 10.1038/s41559-024-02449-x . ПМИД 39048729 .
- ^ Браунштейн, CD (2024). «Молодая птица, возможно, имеющая сходство с кроновой группой из богатой динозаврами меловой экосистемы Северной Америки» . BMC Экология и эволюция . 24 (1). 20. Бибкод : 2024BMCEE..24...20B . дои : 10.1186/s12862-024-02210-9 . ПМЦ 10858573 . ПМИД 38336630 .
- ^ Макинерни, Польша; Блокланд, Дж. К.; Достойно, TH (2024). «Морфология черепа загадочного Genyornis newtoni Stirling and Zeitz, 1896 (Aves, Dromornithidae) с последствиями для функциональной морфологии, экологии и эволюции в контексте Galloanserae» . Историческая биология: Международный журнал палеобиологии . 36 (6): 1093–1165. Бибкод : 2024HBio...36.1093M . дои : 10.1080/08912963.2024.2308212 .
- ^ Мацуока, Х.; Сока, Р.; Хасэгава, Ю. (2024). «Повторное исследование предтазовых позвонков, обнаруженных у голотипа Annakacygna hajimei (Aves, Anseriformes, Cygnini), выявило адаптивную морфологию позвоночного столба, связанную с образом жизни «высшей птицы» » (PDF) . Вестник Музея естественной истории Гумма . 28 : 15–44.
- ^ Теннисон, Алан Джей Ди; Грир, Лиам; Люббе, Паскаль; Маркс, Феликс Г.; Джованарди, Симоне; Роуленс, Николас Дж. (2024). «Ответ Уорти и др. 2022. Ископаемое утиное животное размером с лебедя (Aves: Anatidae) из ранней миоценовой фауны Сент-Батанс в Новой Зеландии. Zootaxa , 5168 (1), 39–50». Зоотакса . 5453 (1): 127–128. дои : 10.11646/zootaxa.5453.1.8 .
- ^ Достойно, Тревор Х.; Скофилд, Р. Пол; Хэнд, Сюзанна Дж.; Де Пьетри, Ванеса Л.; Арчер, Майкл (2022). «Ископаемое анатид размером с лебедя (Aves: Anatidae) из ранней миоценовой фауны Сент-Батанс Новой Зеландии». Зоотакса . 5168 (1): 39–50. дои : 10.11646/zootaxa.5168.1.3 . ПМИД 36101302 .
- ^ Роуленс, Нью-Джерси; Верри, AJF; Коул, ТЛ; Шепард, LD; Теннисон, AJD; Уильямс, М.; Вуд, младший; Митчелл, KJ (2024). «Древние митогеномы свидетельствуют о расселении крохалей в Южном полушарии в позднем миоцене» . Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlae040 .
- ^ Ву, С.; Райндт, FE; Чжан, Дж.; Ван, Дж.; Чжан, Л.; Цюань, К.; Ли, З.; Ван, М.; Ву, Ф.; Цюй, Ю.; Эдвардс, СВ; Чжоу, З.; Лю, Л. (2024). «Геномы, окаменелости и одновременное возникновение современных птиц и цветковых растений в позднем меловом периоде» . Труды Национальной академии наук Соединенных Штатов Америки . 121 (8). е2319696121. Бибкод : 2024PNAS..12119696W . дои : 10.1073/pnas.2319696121 . ПМЦ 10895254 . ПМИД 38346181 .
- ^ Зеленков, Н.В. (2024). «Неожиданная находка перепела (Aves: Charadriiformes: Turnicidae) в нижнем плейстоцене Крыма». Доклады биологических наук . 513 (1 дополнение): S1–S4. дои : 10.1134/S0012496623600148 . ПМИД 38190042 . S2CID 266843308 .
- ^ Гудман, С.М.; Расолонятово, ВЭМ (2024). «Описание шпоры крыла у субископаемого малагасийского чибиса Vanellus madagascariensis (Aves: Charadriiformes, Charadriidae): понимание некоторых возможных особенностей его жизненного цикла и причин его исчезновения» . Геобиос . дои : 10.1016/j.geobios.2024.02.002 .
- ^ Ядвищак, П.; Крюгер, А.; Мёрс, Т. (2024). «Ископаемые и современные тарсометатарси пингвинов: полости, васкуляризация и устойчивость» . Интегративная зоология . дои : 10.1111/1749-4877.12852 . ПМИД 38858828 .
- ^ Ся, Бо-Ян; Пей, Цзюнь-Линг; Ли, Цюань-Го (2024). «Новая окаменелость пингвина с острова Сеймур и переоценка таксономии и разнообразия эоценовых антарктических пингвинов». Палеомир . дои : 10.1016/j.palwor.2024.04.007 .
- ^ Серда, Айова; Тамбусси, КП; Дегранж, Ф.Дж. (2014). «Неожиданные микроанатомические вариации среди эоценовых антарктических стволовых пингвинов (Aves: Sphenisciformes)». Историческая биология: Международный журнал палеобиологии . 27 (5): 549–557. дои : 10.1080/08912963.2014.896907 . hdl : 11336/41915 .
- ^ Ксепка, Д.Т.; Вернинг, С.; Склафани, М.; Болес, ЗМ (2015). «Гистология костей современных и ископаемых пингвинов (Aves: Sphenisciformes)» . Журнал анатомии . 227 (5): 611–630. дои : 10.1111/joa.12367 . ПМК 4609197 . ПМИД 26360700 .
- ^ Кановиль, А.; Робин, Ж.-П.; де Буффрениль, В. (2024). «Онтогенетическое развитие микроструктуры костей конечностей у королевского пингвина Aptenodytes patagonicus (Miller, 1778) с учетом палеоэкологических выводов у Sphenisciformes». Зоологический журнал Линнеевского общества . doi : 10.1093/zoolinnean/zlae002 .
- ^ Маккомиш, Беннет Дж; Чарльстон, Майкл А; Паркс, Мэтью; Барони, Карло; Сальваторе, Мария Кристина; Ли, Жуйцян; Чжан, Гоцзе; Миллар, Крейг Д.; Холланд, Барбара Р.; Ламберт, Дэвид М. (2024). «Древние и современные геномы показывают, что микросателлиты поддерживают динамическое равновесие в глубоком времени» . Геномная биология и эволюция . 16 (3): evae017. дои : 10.1093/gbe/evae017 . ПМЦ 10972684 . ПМИД 38412309 .
- ^ Леони, Р; Алвес-Сильва, Л; да Кошта, Дж; де Араужо-Жуниор, Х; Дантас, М (2024). «Первая находка окаменелостей стервятника-индейки (Cathartes aura) на северо-востоке Бразилии: таксономия, ихнология и тафономическая история». Южноамериканские науки о Земле . 136 . Бибкод : 2024JSAES.13604831L . дои : 10.1016/j.jsames.2024.104831 . S2CID 267602385 .
- ^ Эмсли, SD (2024). «Пещера-гнездо Gymnogyps Californianus (Калифорнийский кондор) позднего плейстоцена в Техасе: новый радиоуглеродный анализ и анализ стабильных изотопов». Орнитология . doi : 10.1093/ornithology/ukae032 .
- ^ Молеон, Маркос; Грасия, Ева; Гарсия, Нурия; Хиль-Санчес, Хосе М.; Годиньо, Ракель; Бежа, Педро; Пальма, Луис; Настоящая, Джоан; Эрнандес-Матиас, Антонио; Муньос, А. Роман; Аррондо, Энеко; Санчес-Сапата, Хосе А. (2024). «Дикая жизнь следует за людьми: междисциплинарная оценка древней колонизации Средиземноморского бассейна хищником-долгожителем» . Люди и природа . 6 (3): 1303–1319. Бибкод : 2024PeoNa...6.1303M . дои : 10.1002/pan3.10642 .
- ^ Акоста Госпиталече, К.; Джонс, В. (2024). «Были ли ужасные птицы главными континентальными хищниками Антарктиды? Новые находки в раннем эоцене острова Сеймур» . Электронная палеонтология . 27 (1). 27.1.а13. дои : 10.26879/1340 .
- ^ Лабарж, ТВ; Гарднер, доктор медицинских наук; Орган, CL (2024). «Эволюция и экология гигантизма у ужасных птиц (Aves, Phorusrhacidae)». Труды Королевского общества B: Биологические науки . 291 (2021). 20240235. дои : 10.1098/rspb.2024.0235 . PMC 11040249. PMID 38654650 .
- ^ Акоста Госпиталече, К.; Джонс, В. (2024). «Сведения о старейших птицах-террористах (Aves, Phorusrhacidae) из эоцена Аргентины». Историческая биология: Международный журнал палеобиологии : 1–9. дои : 10.1080/08912963.2024.2304592 . S2CID 267475903 .
- ^ Зеленков, Н.В. (2024). «Кубинский ара трехцветный Ара в верхнем плейстоцене Западной Кубы». Доклады биологических наук . 516 (1): 32–35. дои : 10.1134/S0012496624700947 . ПМИД 38538825 .
- ^ Риамон, С.; Туртур, Н.; Лушар, А. (2020). «Самая ранняя Тираннида (Aves, Passeriformes) из олигоцена Франции» . Научные отчеты . 10 (1). 9776. Бибкод : 2020НатСР..10.9776Р . дои : 10.1038/s41598-020-66149-9 . ПМК 7299954 . ПМИД 32555197 .
- ^ Лоуи-Мерри, ТМ; Гевори, М.; Боченский, З.М.; Вертц, К.; Кларамунт, С. (2024). «Общее датирование и филогенетическое сходство ранних ископаемых воробьинообразных». Журнал систематической палеонтологии . 22 (1). 2356086. Бибкод : 2024JSPal..2256086L . дои : 10.1080/14772019.2024.2356086 .
- ^ Пёллат, Н.; Питерс, Дж. (2024). «Останки орнитофауны раннего неолита из юго-восточной Анатолии дают представление о распространении видов в раннем голоцене и долгосрочных изменениях в их ареале» . Ибис . дои : 10.1111/ibi.13341 .
- ^ Мартин-Сильверстоун, Элизабет; Анвин, Дэвид М.; Кафф, Эндрю Р.; Браун, Эмили Э.; Аллингтон-Джонс, Лу; Барретт, Пол М. (5 февраля 2024 г.). «Новый птерозавр из средней юры Скай, Шотландия, и раннее разнообразие летающих рептилий» . Журнал палеонтологии позвоночных . 43 (4). дои : 10.1080/02724634.2023.2298741 . ISSN 0272-4634 .
- ^ Пентленд, штат Айдахо; Поропат, Сан-Франциско; Дункан, Р.Дж.; Келлнер, AWA; Бантим, РАМ; Бевитт, Джей-Джей; Тейт, AM; Грайс, К. (2024). « Haliskia peterseni , новый ангангерианский птерозавр из позднего раннего мела Австралии» . Научные отчеты . 14 (1). 11789. Бибкод : 2024NatSR..1411789P . дои : 10.1038/s41598-024-60889-8 . ПМЦ 11169243 . PMID 38866826 .
- ^ Шпиндлер, Фредерик (23 июля 2024 г.). «Связующее звено птерозавров из поздней юры Германии» . Электронная палеонтология . 27 (2): 1–27. дои : 10.26879/1366 . ISSN 1094-8074 .
- ^ Пегас, Родриго В. (10 июня 2024 г.). «Таксономическая заметка о птерозаврах Tapejarid с кладбища птерозавров (группа Кайуа, «Ранний мел Южной Бразилии): свидетельства присутствия двух видов» . Историческая биология : 1–22. дои : 10.1080/08912963.2024.2355664 . ISSN 0891-2963 .
- ^ Бухманн, Р.; Родригес, Т. (2024). «Артрологические реконструкции шеи птерозавра и их значение для положения шеи в состоянии покоя» . ПерДж . 12 . е16884. дои : 10.7717/peerj.16884 . ПМЦ 10893864 . ПМИД 38406270 .
- ^ Чен, Х.; Цзян, С.; Келлнер, AWA; Ван, X. (2024). «Новый взгляд на анатомию черепа птерозавра: рентгеновские снимки выявляют структуру неба и тенденции эволюции» . Коммуникационная биология . 7 (1). 456. дои : 10.1038/s42003-024-06132-6 . ПМЦ 11014945 . ПМИД 38609453 .
- ^ Хендерсон, DM (2024). «Использование головы — краниальное управление у птерозавров». Наука о природе . 111 (3). 29. Бибкод : 2024SciNa.111...29H . дои : 10.1007/s00114-024-01915-7 . ПМИД 38713269 .
- ^ Юн, Чан-Гю (2024). «Геометрический морфометрический подход для установления филогенетического сходства загадочных образцов птерозавров из нижнего мела Южной Кореи» . Acta Palaeontologica Romane . 20 (1): 77–86. doi : 10.35463/апреля 2024.01.06 .
- ^ Хабиб, МБ; Хон, DWE (2024). «Внутривидовая изменчивость птерозавра Rhamphorhynchus muensteri — влияние на полет и социально-сексуальную передачу сигналов» . ПерДж . 12 . е17524. дои : 10.7717/peerj.17524 . ПМЦ 11260407 . ПМИД 39035160 .
- ^ Итак, КС; Ким, PH; Выигран, CG (2024). «Первый сочлененный рамфоринхоидный птерозавр из раннего мела Корейской Народно-Демократической Республики». Палеонтологический журнал . 57 (1 дополнение): S90–S94. дои : 10.1134/S003103012360018X .
- ^ Эредиа, AM; Диас-Мартинес, И.; Пасос, П.Дж.; де Вале, С. (2024). «Следы птерозавров из верхнемеловой (сеноманской) формации Канделерос на северо-западе Патагонии, Аргентина: ихнотаксономические и палеоэкологические перспективы из Гондваны». Палеогеография, Палеоклиматология, Палеоэкология . 650 . 112338. Бибкод : 2024PPP...65012338H . дои : 10.1016/j.palaeo.2024.112338 .
- ^ Этьен, JL; Смит, Р.Э.; Анвин, DM; Смит, RSH; Мартилль, DM (2024). «Гигантский птеродактилоидный птерозавр из британской юры» . Труды Ассоциации геологов . 135 (3): 335–348. Бибкод : 2024PrGA..135..335E . дои : 10.1016/j.pgeola.2024.05.002 .
- ^ Бурло, Р.; Кодорню, Л.; Дефенд, Л.; Лаурин, М. (2024). «Голосеностопный сустав Птеродаустро гиназуи » . Acta Palaeontologica Polonica . 69 (2): 329–350. дои : 10.4202/app.01097.2023 .
- ^ Сюй, Ю.; Цзян, С.; Ван, X. (2024). «Повторное исследование Haopterus gracilis из формации Исянь, Ляонин, Китай». Меловые исследования . 162 . 105933. Бибкод : 2024CrRes.16205933X . дои : 10.1016/j.cretres.2024.105933 .
- ^ Чаффи, А.; Беллардини, Ф. (2024). «Зубы птерозавров из бассейна Южного Неукена (Патагония, Аргентина): новые идеи о реконструкции орнитохейриформной анатомии зубов» . Acta Palaeontologica Polonica . 69 (1): 73–86. дои : 10.4202/app.01122.2023 .
- ^ Гриффин, BW; Мартин-Сильверстоун, Э.; Пегас, Р.В.; Свет, Э.А.; Коста, Франция; Палмер, К.; Рэйфилд, EJ (2024). «Моделирование рук взлетного момента у орнитохейрейного птерозавра» . ПерДж . 12 . е17678. дои : 10.7717/peerj.17678 .
- ^ Юнг, Дж.; Ха, М. (2024). «Новые следы птерозавров со тропы Хвасун Союри (Турониан) в Южной Корее: последствия для их экологической ниши и среды обитания». Палеогеография, Палеоклиматология, Палеоэкология . 645 . 112218. Бибкод : 2024PPP...64512218J . дои : 10.1016/j.palaeo.2024.112218 .
- ^ Гарсия, MS; Фонсека, АО; Деринг, М.; да Роза, А. КАК; Мюллер, RT (2024). «Новое симпатрическое появление лагерпетид (Pan-Aves, Pterosauromorpha) в верхнем триасе южной Бразилии». Журнал южноамериканских наук о Земле . 140 . 104897. Бибкод : 2024JSAES.14004897G . дои : 10.1016/j.jsames.2024.104897 .
- ^ Аньолин, Флорида; Новас, FE; Эскурра, доктор медицины; Майнер, С.; Мюллер, RT (2024). «Комментарии к анатомии тазового пояса Lagerpeton chanarensis Romer, 1971 (Archosauria) и ее влияние на осанку и походку ранних птерозавроморфов». Анатомическая запись . 307 (4): 1001–1010. дои : 10.1002/ar.25389 . ПМИД 38263641 . S2CID 267197820 .
- ^ Шипли, AE; Эльслер, А.; Сингх, ЮАР; Стаббс, ТЛ; Бентон, MJ (2024). «Передвижение и успех Archosauromorpha в раннем мезозое» . Королевское общество открытой науки . 11 (2). 231495. Бибкод : 2024RSOS...1131495S . дои : 10.1098/rsos.231495 . ПМЦ 10846959 . ПМИД 38328568 .
- ^ Уилсон, Л.Н.; Гарднер, доктор медицинских наук; Уилсон, JP; Фарнсворт, А.; Перри, ЗР; Дракенмиллер, PS; Эриксон, генеральный директор; Орган, CL (2024). «Глобальные широтные градиенты и эволюция размеров тела динозавров и млекопитающих» . Природные коммуникации . 15 (1). 2864. Бибкод : 2024NatCo..15.2864W . дои : 10.1038/s41467-024-46843-2 . ПМЦ 10997647 . ПМИД 38580657 .
- ^ Нолл, Ф.; Исикава, А.; Кавабе, С. (2024). «Показатель индекса соотношения мозга и эндокраниальной полости у неорнитских динозавров и других вымерших архозавров» . Журнал сравнительной неврологии . 532 (3). е25597. дои : 10.1002/cne.25597 . ПМИД 38588163 .
- ^ Малафайя, Э.; Мочо, П.; Эскасо, Ф.; Нарваес, И.; Ортега, Ф. (2024). «Таксономическое и стратиграфическое обновление материала, исторически приписываемого мегалозавру из Португалии» . Acta Palaeontologica Polonica . 69 (2): 127–171. дои : 10.4202/app.01113.2023 .
- ^ Амзил, М.; Укассу, М.; Лалленсак, JN; Кляйн, Х.; Зафати, О.; Сэйбер, Х.; Шарьер, А.; Мейер, К.; Герлинский, ГД (2024). «Новые следы динозавров из среднеюрских красных отложений Среднего Атласа (Марокко): применение фотограмметрии к ихнологии и сохранению геологического наследия». Труды Ассоциации геологов . дои : 10.1016/j.pgeola.2024.06.004 .