Орнитозухиды
| Орнитозухиды Временной диапазон: поздний триас ,
| |
|---|---|

| |
| Восстановление жизни орнитозуха , питающегося ринхозавром Hyperodapedon | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | Архозаврия |
| Клэйд : | Псевдозухия |
| Семья: | † Орнитозухиды Хюэне , 1908 г. |
| Типовой вид | |
| † Орнитозух лесварди Ньютон, 1894 г.
| |
| Роды | |
|
† Энигмаспина ? | |
Орнитозухиды — вымершее семейство псевдозуховых ( дальних архозавров родственников современных крокодилов ) триасового периода . Орнитозухиды были четвероногими и факультативно двуногими (например, как шимпанзе), что означает, что они обладали способностью ходить на двух ногах в течение коротких периодов времени. У них были характерные опущенные морды, уникальные лодыжки, перевернутые как крокодилы, и несколько других особенностей, которые отличали их от других архозавров. Орнитозухиды были географически широко распространены во время карнийского и норийского этапов позднего триаса ; их представители были известны из Аргентины , Бразилии и Великобритании . Четыре рода, включающие Ornithosurus , Venaticosurus , Dynamosuchus , [1] и Riojasuchus известны в настоящее время. [2] Семья была впервые установлена немецким палеонтологом Фридрихом фон Хюне в 1908 году. [3]
Описание
[ редактировать ]Череп
[ редактировать ]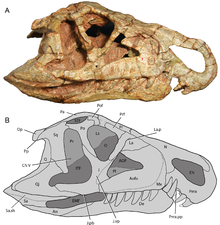

Орнитозухид можно идентифицировать по наличию дугообразной диастемы — промежутка между зубами в передней части морды. Когда челюсть сомкнута, два больших изогнутых зубных зуба (нижней челюсти) входят в диастему, которая расположена между предчелюстной и верхней челюстью . На стенке диастемы имеются две неглубокие впадины для размещения этих зубов. Большие зубные зубы Ornithosurus и Riojasurus расположены позади меньшего, лежащего зубного зуба, который торчит из челюсти. Такого типа положения зубов не наблюдается ни у каких других базальных архозавров. Другой характерной особенностью орнитозухид является их необычная опущенная, нависающая морда, наблюдаемая у Riojasuchus и Venaticosurus , но не у Ornithosurus . [4]
Несколько других особенностей отличают орнитозухид от всех других ранних архозавров. И Ornithosurus , и Riojasuchus имеют небольшое окно или отверстие между небными и крыловидными костями неба , то есть нёба. Контакт носовых и предлобных костей черепа небольшой или отсутствует, исключается большим контактом лобных и слезных костей . У других архозавров, в том числе райзухий , этозавров , птерозавров и динозавроморфов , носово-префронтальный контакт отделяет лобную часть от слезной. [4]
Посткраниальный скелет
[ редактировать ]Посткраниальный скелет почти полностью известен у Riojasuchus , неполный у Ornithosurus и совершенно неизвестен у Venaticosurus . В результате неясно, все ли посткраниальные черты, кажущиеся уникальными для орнитозухид, на самом деле наблюдались у всех членов семейства. Орнитозухиды, известные по приличным посткраниальным останкам, обычно имели около 9 шейных (шея), 14-15 спинных (спина), три крестцовых (бедра) и более 20 хвостовых (хвостовых) позвонков . Над каждым позвонком располагалась пара костных щитков, известных как остеодермы . [2]
Бедренная кость (бедренная кость) имеет выраженный передний вертел. Передний вертел, иногда известный как «малый вертел» (но не связанный с малым вертелом бедренной кости у человека), представляет собой гребень на внешней поверхности бедренной кости, рядом с головкой бедренной кости . Вероятно, это была точка вставки m. iliofemoralis cranialis, помогающая поднять ногу. У большинства архозавров и родственников архозавров отсутствует отчетливый передний вертел, но орнитозухиды являются исключением, наряду с большинством динозавроморфов (динозавров и их близких родственников). [5]
Как и бедренная кость, малоберцовая кость (внешняя большеберцовая кость) также имеет особое место прикрепления мышц. Речь идет о подвздошно-фибулярной мышце, которая помогает выпрямлять конечности. У большинства архозавров подвздошно-фибулярная мышца прикрепляется к малоберцовой кости с помощью крошечного гребня в проксимальной части малоберцовой кости, возле колена. Однако у орнитозухид есть гораздо более крупная точка прикрепления подвздошно-фибулярной мышцы в форме ручки, расположенная примерно на середине стержня малоберцовой кости. У фитозавров и этозавров также есть общая точка прикрепления в форме бугорка посередине малоберцовой кости, поэтому неясно, является ли случай у орнитозухид уникальным случаем конвергентной эволюции или, альтернативно, сохранением признака, независимо утраченного несколькими линиями архозавров. [6]
В отличие от большинства других ранних архозавров, когтевые кости стоп (самые дистальные кости стоп, образующие когти) сжаты с боков. Они острые и загнутые. Ногтевые кости очень глубокие, их высота превышает длину, особенно на внутренних пальцах. Этот тип когтей не встречается ни у одного другого триасового архозавра, за исключением птерозавров. [4]

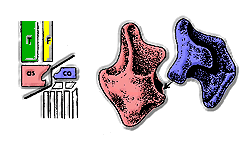
Основные группы архозавров часто отличали друг от друга по строению их лодыжек. У большинства круротарсанов таранная кость имеет выпуклый выступ, который вписывается в вогнутое пространство пяточной кости . Это состояние часто называют «крокодиловой» лодыжкой, поскольку это наиболее распространенный тип лодыжки у круротарзанов. Орнитозухиды уникальны среди круротарсанов и всех других архозавров тем, что у них есть лодыжка, перевернутая крокодилом, в которой расположение вогнутости обратное; вместо того, чтобы находиться на пяточной кости, он находится на таранной кости. У орнитозухид пяточная кость имеет выпуклый выступ, аналогичный выпуклому выступу на «крокодиловой» таранной кости. [4]
Филогения
[ редактировать ]Орнитозухиды обычно считаются частью более крупной клады Сухая , которая включает этозавров, рауизухий и крокодиломорфов. [7] [8] [9] Ниже представлена кладограмма, основанная на Несбитте (2011), показывающая расположение Ornithosuchidae среди Archosauriformes. [6]
| Архозавриформы |
| ||||||||||||||||||||||||||||||||||||||||||||||||

Ссылки
[ редактировать ]- ^ Jump up to: а б Мюллер, Родриго Т.; Фон Бачко, М. Белен; Дезохо, Джулия Б.; Несбитт, Стерлинг Дж. (31 января 2020 г.). «Первый орнитозухид из Бразилии и его макроэволюционное и филогенетическое значение для фаун позднего триаса в Гондване» (PDF) . Acta Palaeontologica Polonica . 65 . дои : 10.4202/app.00652.2019 .
- ^ Jump up to: а б Бачко, М. Белен фон; Эскурра, Мартин Д. (1 января 2013 г.). «Орнитозухиды: группа триасовых архозавров с уникальным голеностопным суставом» . Геологическое общество, Лондон, специальные публикации . 379 (1): 187–202. Бибкод : 2013GSLSP.379..187B . дои : 10.1144/SP379.4 . hdl : 11336/41617 . ISSN 0305-8719 . S2CID 130687362 .
- ^ Хюне, Ф. фон. 1908. «Динозавры европейской триасовой формации с учетом неевропейских находок». Геологические и палеонтологические трактаты 1 (Приложение): 1–419.
- ^ Jump up to: а б с д Серено, ПК (1991). «Базальные архозавры: филогенетические связи и функциональные последствия» . Журнал палеонтологии позвоночных . 11 (Приложение 4): 1–53. Бибкод : 1991JVPal..11S...1S . дои : 10.1080/02724634.1991.10011426 .
- ^ Лангер, Макс К.; Бентон, Майкл Дж. (6 ноября 2006 г.). «Ранние динозавры: филогенетическое исследование» (PDF) . Журнал систематической палеонтологии . 4 (4): 309–358. Бибкод : 2006JSPal...4..309L . дои : 10.1017/s1477201906001970 . ISSN 1477-2019 . S2CID 55723635 .
- ^ Jump up to: а б Несбитт, SJ (2011). «Ранняя эволюция архозавров: взаимоотношения и происхождение основных клад» . Бюллетень Американского музея естественной истории . 352 : 1–292. дои : 10.1206/352.1 . hdl : 2246/6112 .
- ^ Несбитт, С.Дж.; Норрелл, Массачусетс (2006). «Чрезвычайное сближение планов тела ранних зузиев (Archosauria) и орнитомимидных динозавров (Theropoda)» . Труды Королевского общества Б. 273 (1590): 1045–1048. дои : 10.1098/rspb.2005.3426 . ПМК 1560254 . ПМИД 16600879 .
- ^ Несбитт, SJ (2007). «Анатомия Effigia okeeffeae (Archosauria,uchia), тероподоподобная конвергенция и распределение родственных таксонов» (PDF) . Бюллетень Американского музея естественной истории . 302 : 1–84. doi : 10.1206/0003-0090(2007)302[1:TAOEOA]2.0.CO;2 . hdl : 2246/5840 . S2CID 55677195 .
- ^ Брусатте, СЛ; Бентон, MJ; Дезохо, Дж.Б.; Лангер, MC (2010). «Филогения высшего уровня Archosauria (Tetrapoda: Diapsida)» (PDF) . Журнал систематической палеонтологии . 8 (1): 3–47. Бибкод : 2010JSPal...8....3B . дои : 10.1080/14772010903537732 . hdl : 20.500.11820/24322ff3-e80e-45f2-8d53-d35fd104195c . S2CID 59148006 .
