Джунггарзух
| Джунггарзух | |
|---|---|
| |
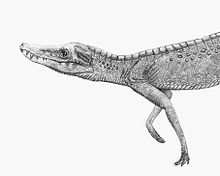
| Реконструкция жизни Junggarsuchus с использованием гипотетических остеодерм. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | Архозаврия |
| Клэйд : | Псевдозухия |
| Клэйд : | Крокодиломорфа |
| Клэйд : | Солидокрания |
| Род: | † Джунггарзух Кларк и др. , 2004 г. |
| Типовой вид | |
| † Junggarsurus sloani Кларк и др. , 2004 г.
| |
Junggarsurusus ( / ˌ d ʒ ə ŋ ɡ ə r ˈ s u k ə s / — вымерший род сфенозуховых из крокодиломорфов Среднего ) [ 2 ] или поздняя юра [ 1 ] [ 3 ] период Китая . Тип и единственный вид — J. sloani . Родовое название Junggarsuchus англизация происходит от котловины ( Джунгарского ) Джунгарской , [ 4 ] где было найдено ископаемое, и греческое слово « souchos », означающее «крокодил » . Видовое название « слоани » дано в честь К. Слоана, которому приписывают открытие голотипа . [ 2 ]
Открытие
[ редактировать ]Junggarsuruchus был обнаружен в верхней части нижней пачки формации Шишугоу в Синьцзяне , Китай , в местонахождении Уцайвань. [ 2 ] [ 5 ] Тип и единственный экземпляр был описан в 2004 году Джеймсом Кларком, Сюй Синем , Кэтрин Форестер и Юань Ван в журнале Nature . [ 2 ] но полное остеологическое описание он получил только в 2022 году, когда Александр Рубеншталь, Майкл Кляйн и И Хунъю опубликовали монографию вместе с двумя первоначальными описателями Джеймсом Кларком и Сюй Синем. [ 3 ]
Это относительно небольшое животное: длина черепа голотипа составляет всего 144 миллиметра (5,7 дюйма). Правая передняя конечность имела длину около 29 сантиметров (11 дюймов) от плеча до пястных костей . [ 3 ] Это сделало бы животное в жизни размером примерно с домашнюю кошку . Однако его общая длина неизвестна, поскольку у голотипа сохранился только один позвонок из хвоста. [ 2 ]
Голотип, получивший обозначение ИВПП 14010 , [ 2 ] состоит из почти целого черепа с неповрежденной черепной коробкой и нижними челюстями, большей части левой передней конечности, проксимальных концов локтевой и лучевой кости правой передней конечности вместе с правой плечевой костью, пятнадцати шейных и спинных позвонков, а также большей части грудная клетка и связанный с ней хвостовой позвонок. [ 3 ] Большая часть скелета была полностью сочленена, при этом некоторые элементы находились в ассоциации и разъединились. [ 2 ] На момент публикации монографии большая часть экземпляра еще не была подготовлена, а некоторые кости оставались скрыты матрицей на момент ее повторного описания. [ 3 ]
Череп голотипа был перевезен из Института палеонтологии и палеоантропологии позвоночных , где он был первоначально репонирован, в Университет Джорджа Вашингтона , где он был изучен и всесторонне переописан Рубенсталем и его коллегами с использованием современных компьютерной томографии технологий . Подготовка скелета была настолько тщательной, что элементы скелета, поврежденные во время окаменения, удалось склеить с минимальным несоответствием. [ 3 ]
Описание
[ редактировать ]В своем первоначальном описании черепа Кларк и его коллеги отметили, что там было очень мало краниального кинезиса и были места прикрепления очень мощных челюстных мышц, которые являются производными чертами, обнаруженными у современных крокодилов. [ 2 ] Что касается посткраниального скелета, они отметили, что позвоночник голотипа, вероятно, имел очень низкую вертикальную подвижность по всей длине и был в основном приспособлен для бокового движения, что соответствует современным крокодилам. [ 3 ] [ 2 ] Однако очень существенным различием между Junggarsurusus и его современными родственниками было полное отсутствие остеодерм у этого экземпляра, несмотря на отсутствие подвижности позвоночника, хотя предполагалось, что эти признаки развились одновременно. Они предполагают, что отсутствие сохранившихся остеодерм может быть результатом тафономических условий экземпляра или что на момент смерти он мог быть молодым. [ 2 ]
Как и у других сфенозухов, конечности Junggarsuchus были приспособлены к наземному передвижению (передвижению по суше), а не к полуводному передвижению, наблюдаемому у современных крокодилов. [ 3 ] К этим адаптациям относятся: вертикально ориентированная кость плеча , шаровидный плечевой сустав и функционально трехпалая (трехпалая) кисть вследствие редукции 5-го пальца и отсутствия 1-го пальца. [ 2 ] Однако тот факт, что эти наземные адаптации, по-видимому, являются общими для Junggarsurusus и более базальных животных, таких как Saltoposuchus , означает, что это, вероятно, представляет собой состояние крокодиломорфов наследственное , а не производную черту «сфенозухов» и полуводные адаптации коронная группа появилась в их эволюции гораздо позже. [ 3 ] [ 6 ]
Череп
[ редактировать ]Специализации, наблюдаемые в черепе Junggarsuchus, в первую очередь связаны с усилением структуры черепа и развитием более сильной мускулатуры челюстей. Диапсидные черепа изначально были относительно легкими, поэтому для обеспечения высокой силы укуса без повреждения черепа требуется значительная специализация. [ 7 ] Это можно увидеть у динозавра тираннозавра , у которого полностью сросшаяся носовая кость (кость на дорсальной средней линии морды), тогда как у большинства других теропод есть швы в топологии кости в этом месте. [ 8 ] Некоторые из окон черепа были уменьшены в размерах по сравнению с другими ранними крокодиломорфами, в первую очередь предглазничное окно и надвисочное окно . [ 2 ] Кроме того, некоторые кости неба стали увеличенными по сравнению с более базальными крокодиломорфами . Квадратная кость также намного ближе к латеро-клиновидной кости нет шва , и между теменными костями . [ 3 ] Оба из них уменьшают гибкость черепа, что, как было предложено, позволяет приложить большую силу укуса, не повреждая череп. [ 2 ] [ 9 ] Однако эта тенденция увеличения прочности черепа неоднородна, и двумя заметными изменениями, противоречащими этой тенденции, являются появление дополнительного отверстия в квадратной кости и отсутствие значительного контакта между носовыми и слезными костями . Хотя большинство характеристик черепа демонстрируют промежуточное положение Junggarsurusus на генеалогическом древе крокодиломорфов, череп демонстрирует несколько совершенно новых адаптаций. Латеральная поверхность угловой кости имеет обширные места прикрепления челюстной мускулатуры, что наблюдается у некоторых крокодилов, но не у других промежуточных форм. Это, как и некоторые другие адаптации, скорее всего, способствовало увеличению силы укуса потенциальной добычи. [ 3 ]
Другой примечательной чертой черепа Junggarsurusus является появление пневматических пространств в квадратной кости, парабазисфеноиде и, возможно, крыловидной кости . Хотя это, казалось бы, противоречит тенденции затвердевания черепа у Junggarsurusus и родственных ему таксонов (базальных солидокраний), Рубеншталь и его коллеги предполагают, что вполне возможно, что эти пневматические пространства позволяли тканям черепа противостоять и поглощать удары, которые в противном случае повредили бы структуру, состоящую из твердая кость. [ 9 ] Однако авторы отмечают, что это только предположение, и эволюция краниальной пневматичности у крокодиломорфов вместо этого может отражать инвазию этих тканей черепными синусами . [ 3 ] Плохая сохранность крыловидного отростка голотипа также затрудняет определение точной степени пневматичности или предположения относительно какой-либо из его возможных функций. [ 3 ]
Примечательно, что внутреннего уха анатомия Junggarsuchus демонстрирует значительную наземную адаптацию. Полукружный канал высокий и узкий, в отличие от водных крокодилов, который, как полагают, помогает животному ориентировать голову и взгляд. Это было необходимо для охоты на наземную добычу и для поддержания равновесия при движении по суше. [ 3 ]
Несмотря на исключительную сохранность черепа, есть несколько ключевых областей, где расположение костей черепа является неопределенным из-за деформации или повреждения, нанесенного черепу в процессе окаменения . Эти области представляют собой контакт между угловой и надугольной костями , а также контакт между чешуйчатой и заглазничной костями . На основании данных КТ Рубеншталь и его коллеги предполагают, что надугловая челюсть, вероятно, короткая и не имеет значительного контакта с угловой. Однако возможно, что надугловая имеет значительный контакт на значительной длине как суставной , так и угловой костей, что делает ее похожей на состояние ее близкого родственника Dibothrosuchus .
Другая неопределенность связана с контактами между квадратной костью и заглазничными костями в задней части черепа. Переломы сохранившейся кости затрудняют определение того, какие переломы отражают тафономическое повреждение, а какие — настоящие швы на кости. Авторы предполагают, что заглазничная кость вносит вклад только в дорсальный гребень надвисочного окна, хотя они рассматривают возможность того, что заглазничная кость простирается вниз по заднему гребню окна, проходя параллельно чешуйчатой кости, а не сходясь с ней прямо. [ 3 ]
Посткраниальный скелет
[ редактировать ]Junggarsurus демонстрирует многочисленные приспособления для беглости . [ 3 ] К ним относится сокращение количества пальцев, соприкасающихся с землей, с пяти, что является наследственным состоянием у псевдозухий , до всего лишь трех в передних конечностях Junggarsuchus . [ 2 ] Ранее было признано, что редукция внешних пальцев так, что они не соприкасаются с землей, является тенденцией, сохранившейся среди бегающих четвероногих. [ 3 ] [ 10 ] первая и пятая цифры сильно редуцированы а у Junggarsurusus . [ 2 ] Другие особенности конечностей включают увеличенную поверхность на переднем крае лопатки , которая интерпретируется как место прикрепления расширенных мышц, способствующих ретракции конечностей. Суставы плеч и костей конечностей также обычно ориентированы таким образом, что конечности удерживаются прямо под телом и имеют возможность сгибаться под телом животного. [ 3 ] Эти дистальные кости руки ( локтевая и лучевая кость ) также намного длиннее плечевой кости , чем у других архозавров (хотя они все же не такие длинные, как плечевая кость). Это было интерпретировано как дополнительный признак беглости других крокодиломорфов. [ 11 ] Задние конечности у голотипа не сохранились, [ 2 ] поэтому неясно, имеют ли бедренная , большеберцовая и малоберцовая кость такие же взаимоотношения. [ 3 ]
Заметное отсутствие остеодерм , которое в первоначальном описании считалось чисто тафономическим, [ 2 ] Поздние авторы полагают, что это отражает подлинное отсутствие этих особенностей как часть общей тенденции к большей наземной мобильности и гибкости, которая не наблюдается у современных родственников Junggarsuruchus . Характеристики позвоночника, включая зигапофизы, позволяют предположить гораздо более гибкий общий диапазон движений, которого полностью лишено большинство современных крокодилов. [ 3 ] Этот уровень земной гибкости характерен для крокодиломорфов, рано расходящихся, а также для архозавров в целом. Большинство близких родственников Junggarsuruchus , таких как Dibothrosuchus , [ 12 ] Террестрисух , [ 13 ] и дромикозух [ 14 ] столь же грациозны и приспособлены к быстрому передвижению по земле. Это говорит о том, что водные адаптации, наблюдаемые у современных крокодилов, а также у многих их вымерших родственников, таких как талаттозухи , не являются наследственным состоянием этой группы. [ 3 ]
ничего не известно, О задних конечностях, бедрах и хвосте Junggarsuchus поскольку голотип, единственный известный экземпляр, не сохраняет эти элементы. [ 2 ] Однако в голотипе могут быть элементы, которые еще предстоит подготовить, и которые могли бы раскрыть информацию об этих участках скелета после того, как камень будет удален и будут описаны любые новые кости. [ 3 ]
Классификация
[ редактировать ]Первоначальное описание Junggarsurus , сделанное Джеймсом А. Кларком и его коллегами, показало, что он является сестринским таксоном клады Crocodyliformes , а также обнаружило, что традиционная группа Sphenosuchia не была монофилетической группой, а скорее образовывала большую политомию от базального до базального уровня. крокодилиформные. Синапоморфии клады Junggarsuchus -crocodyliformes в их анализе включали: экзоципитали , которые встречаются по средней линии над большим затылочным отверстием , большое расширение на нижних сторонах экзоципиталей, которое контактирует с квадратной костью , сильно изогнутые скуловые кости , сужение задней части. часть теменной кости и отверстие в квадратной кости . Их анализ показан ниже. [ 2 ]
| Сухая |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
В 2017 году Хуан Мартин Леарди и его коллеги переописали близкородственный таксон Macelognathus , который первоначально был описан О. К. Маршем в 1884 году как вид динозавров. [ 15 ] Они использовали тот же набор данных, что и Кларк и его коллеги, но их анализ включал гораздо больше таксонов, включая недавно описанные таксоны Almadasurus и Carnufex . Они восстанавливают немного более четкое филогенетическое дерево, за исключением политомии в основании. Их анализ примечателен тем, что Junggarsurus включен в него как несколько более базальный, чем монофилетический Hallopodidae . Синапоморфии клады, включающей Junggarsurusus , Hallopodidae и crocodyliformes, включают следующее: расширенную базисфеноидную кость , экзозатылочные кости, которые контактируют с квадратной костью, закрытый проход для внутренних сонных артерий , большое поствисочное окно, окруженное чешуйчатой и экзоципитальной костями. и радиус короче плечевой кости. Они также обнаруживают Macelognathus как близкого родственника Hallopus . [ 16 ] Другие авторы предположили, что этот результат трудно подтвердить или даже оспорить, поскольку Macelognathus известен в основном по материалу черепа, а у Hallopus не сохранилось ни одного черепа. [ 3 ] [ 15 ] Результаты их анализа приведены ниже. [ 16 ]
| Крокодиломорфа |
| |||||||||||||||
В своем переописании Рубеншталь и его коллеги обнаружили новую кладу Solidocrania , что означает «твердые черепа», в связи с отсутствием краниального кинезиса. [ 2 ] [ 3 ] Эта клада была определена как наименее инклюзивная клада, включающая Junggarsuchus , Macelognathus и Almadasurus . Результаты их филогении показали, что эти три таксона образуют базальную ступень по отношению к Crocodyliformes, и, таким образом, сами Crocodyliformes будут включены в Solidocrania. [ 3 ] Сообщалось, что синапоморфиями этой клады являются: две большие глазные кости , чешуйчатая кость , которая контактирует с задней поверхностью квадратной кости , ограждение черепно-квадратного канала чешуйчатой и затылочной костями, сближение латеросфеноидальной и квадратной костей, соприкосновение отокципитальной и квадратной кости, экспансивный и пневматический парабазисфеноид, развитый гребень на задней поверхности эктоптеригоид, выступающий вдоль внутренней поверхности скуловой кости , передний край лопатки , превышающий задний край, и низкий локтевой отросток локтевой кости . [ 3 ] Были предложены дополнительные синапоморфии, но они получили более слабую поддержку. Результаты их филогенетического анализа, рассчитанные путем обнаружения строгого консенсуса двух наиболее экономных деревьев, показаны ниже. [ 3 ]
Новые результаты этой филогении включали обнаружение Hallopodidae полифилетичности , а также того, что таксон Calsoyasuchus является сестринским таксоном Thalattosuria , хотя он традиционно считался гониофолидом . Было также обнаружено, что Phyllodontosurus является сестринским таксоном Junggarsurusus . [ 3 ] хотя это было слабо подтверждено, поскольку Phyllodontosurus известен только по одному сильно деформированному черепу и связанным с ним зубам. [ 17 ] и его место в филогении авторов значительно различалось в зависимости от его сходства. [ 3 ] Авторы также отметили, что Hsisosurus , обычно считающийся базальным по отношению к расколу зифосухий - неозухий , на самом деле может быть более тесно связан с зифосухиями. [ 3 ]
Три таксона, которые определяют Solidocrania, Junggarsurusus , Macelognathus и Almadasurus , относятся к позднеюрскому возрасту, однако все они считаются более базальными, чем Crocodyliformes , которые, как известно, возникли в позднем триасе . [ 2 ] [ 3 ] [ 18 ] длиной 50 миллионов лет Рубеншталь и его коллеги утверждают, что это открытие означает, что существует призрачная линия солидокрановых таксонов , которая уходит корнями в триасовый период. Если их гипотеза верна, это будет означать, что Junggarsurus и его родственники представляют собой одних из последних выживших некрокодилиформных крокодиломорфов. Однако авторы осторожно отмечают, что, учитывая позднее появление Junggarsurusus и других базальных таксонов солидокраний, вполне возможно, что Solidocrania не является естественной кладой и что объединяющие характеристики черепа могут быть вторично полученным признаком, являющимся результатом конвергентных эволюция, а не черты, унаследованные от общего предка с крокодилиформными. [ 3 ]
Палеобиология
[ редактировать ]В 2023 году Эмили Лесснер, Кэтлин Доллман, Джеймс Кларк, Сюй Син и Кейси Холлидей провели анализ псевдозуховых лицевых нервов, используя материал черепа более чем 20 различных таксонов, включая Junggarsuchus . Одним из их открытий было то, что Junggarsurus был одним из самых ранних дивергентных крокодиломорфов, у которых было линейное расположение отверстий под зубами нижней челюсти . У всех ранее дивергентных таксонов эти отверстия располагались хаотично. [ 19 ]
Их анализ завершился обнаружением заметной тенденции в тактильной чувствительности псевдозуховых морд по мере их эволюции, при этом все более производные группы обладали увеличивающейся плотностью нервов вдоль морды. Эти нервы используются современными крокодилами для обнаружения движения в воде, но исследование пришло к выводу, что увеличение количества этих нервов предшествовало эволюции полуводных крокодилообразных. Одно из возможных объяснений этого очевидного несоответствия, которое предполагают авторы, заключается в том, что ранее дивергентные наземные крокодиломорфы могли демонстрировать экологию питания , которая включала поиск добычи на земле или вблизи нее. Junggarsuchus заметно отстает от этой тенденции, поскольку предполагаемая плотность этих нервов намного ниже, чем у сопоставимых таксонов, таких как Macelognathus и Litargosuchus . Однако авторы не заявляют о каких-либо возможных последствиях, которые это может иметь для экологии питания конкретно Junggarsurusus . [ 19 ]
Палеоэкология
[ редактировать ]Диета
[ редактировать ]Джунггарзух почти наверняка был плотоядным, как и большинство других крокодиломорфов. [ 3 ] У него были мечевидные зубы с зазубринами , приспособленными для прорезания плоти. [ 2 ] Однако это предположение осложняется наличием близкородственного таксона Phyllodontosuchus , [ 3 ] имевшие гетеродонтные зубы. [ 11 ] Гетеродонтия – это адаптация, которая редко встречается у рептилий, и когда она появилась (например, у Pakasuchus и Chimaerasuchus ), ее считали адаптацией к новым стратегиям питания. [ 20 ] [ 21 ] [ 22 ] Точно так же, хотя и не так поразительно, зубы Macelognathus не имеют зазубрин на коронках, а их нижнечелюстной симфиз полностью беззуб, что было интерпретировано как адаптация к травоядности. [ 11 ] Junggarsurus не имеет подобных адаптаций; у него более традиционные зубчатые зубы, которые доходят до конца зубной кости и предчелюстной кости . [ 22 ] Причина такой резкой изменчивости этих близкородственных таксонов неясна и может быть отражением нехватки и относительной неполноты останков этих таксонов, что позволяет проводить лишь ограниченные сравнения между ними. [ 15 ]
Прямых доказательств того, из чего именно мог состоять рацион джунггарсухуса , нет, но, учитывая его размер и зубной ряд, большинство авторов заявили, что наиболее разумным предположением является то, что он был хищником, преследующим мелкую позвоночную добычу. [ 3 ] [ 11 ] Более того, было показано, что общая форма черепа и соотношение его высоты к ширине (т.е. его «плоскостность») больше похожи на современных крокодилов, чем на современных крокодиломорфов. Это связано с местами прикрепления медиальной крыловидной мышцы , которая является важной мышцей, используемой для смыкания челюстей, а это означает, что Junggarsurus , возможно, был приспособлен для ловли мелкой добычи. [ 23 ] В целом, косвенные данные, по-видимому, указывают на то, что джунггарзухи, скорее всего, питались мелкими животными, такими как примитивные млекопитающие , чешуйчатые и, возможно, вылупившиеся динозавры . [ 11 ]
Палеосреда
[ редактировать ]
Единственные останки Junggarsuruchus , описанные на данный момент, были обнаружены недалеко от города Уцайвань в Синьцзяне , Китай . [ 5 ] Это местонахождение является частью нижней пачки свиты Сишугоу . [ 24 ] который колеблется от 164 до 159 миллионов лет назад. Этот интервал охватывает переход от средней юры к поздней юре , хотя большая часть его недавно была датирована поздней юрой. [ 25 ] Сегодня этот регион является внутренним и засушливым, но в поздней юре он образовал прибрежный бассейн на северных берегах океана Тетис . [ 26 ]
Нижняя (или Уцуйвань) пачка Сишугоу состоит в основном из отложений красного аргиллита и песчаника . Предполагается, что это была лесистая аллювиальная веерная среда, которая подвергалась периодическим наводнениям, что объясняет большое разнообразие окаменелостей мелких животных, сохранившихся в этом районе, а также обилие окаменелых деревьев. Член Wucaiwan сохраняет окаменелости двоякодышащих рыб , амфибий , крокодилов , тритилодонтов и динозавров различных размеров. Однако считается, что верхние части этой пачки, где был обнаружен Junggarsuruchus , представляли собой более традиционную речную или водно-болотную среду с менее интенсивными наводнениями, чем нижние части пачки. [ 25 ] Климат этого района в позднеюрском периоде был умеренным, сезонно влажным и сухим. [ 26 ] Такой характер осадков привел к появлению сезонных болот, что, возможно, усугублялось разжижением субстрата из-за шагов массивных зауроподов , которые создавали «ямы смерти», в которых ловились и хоронили мелких животных. [ 25 ] [ 27 ]

На территории Уцайваня также были обнаружены значительные отложения вулканического пепла , что указывает на то, что вулканическая активность в западной части Китая в это время возрастала. [ 25 ]
Современная фауна
[ редактировать ]было обнаружено множество мелких животных В формации Сишугоу . Различные останки мелких животных были отнесены к различным группам, но им еще не дали биномиальных названий . К ним относятся останки двоякодышащих рыб , брахиопоидных амфибий, докодонтных и тритилодонтных млекопитающих , ящериц и черепах . Некоторые из них сохранились почти полностью и в артикуляционном виде. [ 25 ] Существует также небольшой крокодиломорф, который может быть связан с Junggarsurusus , но еще не получил официального описания или названия. [ 28 ] [ 29 ] В этом районе также были обнаружены различные останки динозавров, которым еще не присвоены названия. К ним относятся стегозавры , анкилозавры , орнитоподы , столбняки и предполагаемый орнитомимозавр . [ 24 ] [ 30 ]
Названные окаменелости включают примитивного родственника млекопитающих Yuanotherium , crocodyliformes Sunosuchus и Nominosuchus , а также птерозавров Sericipterus и Kryptodrakon . [ 25 ] Динозавры — самая распространенная и разнообразная часть наземной фауны, обитающая в Сишуго. [ 26 ] Они представлены мелкими птицетазовыми типами Yinlong , Hualianceratops и Eugongbusaurus , а также зауроподами Klamelisaurus , Bellusaurus и Mamenchisaurus sinocanadorum . Все крупные наземные хищники в экосистеме были тероподами . Они варьировались от маленьких целурозавров, таких как Haplocheirus , Aorun и Guanlong , до крупных карнозавров, таких как Sinraptor . Также примечателен в этом районе небольшой цератозавр Limusaurus , который сохранился в одной из грязных «ям смерти». [ 25 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б Шуаньер, Иона Н.; Кларк, Джеймс М.; Норелл, Марк А.; Сюй, Син (2014). «Краниальная остеология Haplocheirus sollers Choiniere et al., 2010 (Theropoda: Alvarezsauroidea) » Новитаты Американского музея (3816): 1–44. дои : 10.1206/3816.1 . S2CID 4667472 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v Кларк, Джеймс М.; Син Сюй; Форстер, Кэтрин А.; Юань Ван (2004). «Сфенозух средней юры из Китая и происхождение черепа крокодила» (PDF) . Природа . 430 (7003): 1021–1024. Бибкод : 2004Natur.430.1021C . дои : 10.1038/nature02802 . ПМИД 15329719 . S2CID 4420246 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть Рубеншталь, Александр А.; Кляйн, Майкл Д.; Йи, Хунъюй; Сюй, Син; Кларк, Джеймс М. (октябрь 2022 г.). «Анатомия и взаимоотношения ранних дивергентных крокодиломорфов Junggarsuchus sloani и Dibothrosuchus elaphros» . Анатомическая запись . 305 (10): 2463–2556. дои : 10.1002/ar.24949 . ISSN 1932-8486 . ПМК 9541040 . ПМИД 35699105 .
- ^ Койпер, Кэтлин (2014). «Джунгли» . Britannica.com . Британская энциклопедия . Проверено 6 августа 2023 г.
- ^ Jump up to: а б Элрой, Джон (2009). «Типовая местность Юнгарсухус (юра Китая)» . База данных палеобиологии . Получено 0 августа 2023 г.
Также известный как Вукайван
- ^ Холлидей, Кейси М.; Шахнер, Эмма Р. (2022). «Послания эпохи крокодилов: Новые открытия древних родословных» . Анатомическая запись . 305 (10): 2343–2352. дои : 10.1002/ar.25043 . ПМИД 35912969 . S2CID 251221205 .
- ^ Абель, Паскаль; Вернебург, Ингмар (2021). «Морфология височной области черепа четвероногих: история исследований, функциональные объяснения и новая комплексная схема классификации» . Биологические обзоры . 96 (5): 2229–2257. дои : 10.1111/brv.12751 . ПМИД 34056833 . S2CID 235256536 .
- ^ Рэйфилд, Эмили Дж. (2004). «Черепная механика и питание тираннозавра рекса » . Труды Лондонского королевского общества. Серия Б: Биологические науки . 271 (1547): 1451–1459. дои : 10.1098/rspb.2004.2755 . ПМК 1691752 . ПМИД 15306316 .
- ^ Jump up to: а б Уитмер, Лоуренс М. (1997). «Эволюция анторбитальной полости архозавров: исследование реконструкции мягких тканей в ископаемых записях с анализом функции пневматичности» . Журнал палеонтологии позвоночных . 17 :1–76. Бибкод : 1997JVPal..17S...1W . дои : 10.1080/02724634.1997.10011027 .
- ^ Кумбс, Уолтер П. (1978). «Теоретические аспекты беговой адаптации динозавров» . Ежеквартальный обзор биологии . 53 (4): 393–418. дои : 10.1086/410790 . S2CID 84505681 .
- ^ Jump up to: а б с д и Ирмис, Рэндалл Б.; Несбитт, Стерлинг Дж.; Сьюс, Ханс-Дитер (2013). «Ранние крокодиломорфы» . Геологическое общество, Лондон, специальные публикации . 379 (1): 275–302. Бибкод : 2013GSLSP.379..275I . дои : 10.1144/SP379.24 . S2CID 219190410 .
- ^ Ву, Сяо-Чун; Чаттерджи, Санкар (1993). « Dibothrosuchus elaphros , крокодиломорф из нижней юры Китая и филогения Sphenosuchia» . Журнал палеонтологии позвоночных . 13 (1): 58–89. Бибкод : 1993JVPal..13...58W . дои : 10.1080/02724634.1993.10011488 .
- ^ Лекуона, А.; Эскурра, доктор медицины; Ирмис, РБ (2016). «Ревизия раннего крокодиломорфа Trialestes romeri (Archosauria,uchia) из нижнего верхнего триаса формации Ишигуаласто в Аргентине: один из старейших известных крокодиломорфов» . Статьи по палеонтологии . 2 (4): 585–622. дои : 10.1002/spp2.1056 . hdl : 11336/66393 . S2CID 88719030 .
- ^ Сьюс, Ханс-Дитер; Олсен, Пол Э.; Картер, Джозеф Г.; Скотт, Дайан М. (2003). «Новый крокодиломорфный архозавр из верхнего триаса Северной Каролины» (PDF) . Журнал палеонтологии позвоночных . 23 (2): 329–343. doi : 10.1671/0272-4634(2003)023[0329:ANCAFT]2.0.CO;2 . S2CID 17319720 .
- ^ Jump up to: а б с Гёлих, Урсула Б.; Кьяппе, Луис М.; Кларк, Джеймс М.; Сьюс, Ханс-Дитер (2005). «Систематическое положение предполагаемого динозавра позднеюрского периода Macelognathus (Crocodylomorpha: Sphenosuchia)» . Канадский журнал наук о Земле . 42 (3): 307–321. Бибкод : 2005CaJES..42..307G . дои : 10.1139/E05-005 .
- ^ Jump up to: а б Леарди, Хуан Мартин; Пол, Диего; Кларк, Джеймс Мэтью (2017). «Подробная анатомия черепной коробки Macelognathus vagans Marsh, 1884 (Archosauria, Crocodylomorpha) с использованием томографии высокого разрешения и новых взглядов на базальную филогению крокодиломорфов» . ПерДж . 5 : е2801. дои : 10.7717/peerj.2801 . ПМК 5251941 . ПМИД 28133565 .
- ^ Харрис, Джеральд Д.; Лукас, Спенсер Г.; Эстеп, JW; Цзяньцзюнь Ли (2000). «Новый и необычный сфенозухий (Archosauria: Crocodylomorpha) из нижнеюрской формации Луфэн, Китайская Народная Республика» . Новогодний альбом по геологии и палеонтологии, Трактаты . 215 (1): 47–68. дои : 10.1127/njgpa/215/2000/47 .
- ^ Мартинес, Рикардо Н.; Алькобер, Оскар А.; Пол, Диего (2018). «Новый протозухид крокодилиформный (Pseudosuria, Crocodylomorpha) из формации Нориан-лос-Колорадос, северо-запад Аргентины» . Журнал палеонтологии позвоночных . 38 (4): (1)-(12). дои : 10.1080/02724634.2018.1491047 . hdl : 11336/98862 . S2CID 109740761 .
- ^ Jump up to: а б Лесснер, Эмили Дж.; Доллман, Кэтлин Н.; Кларк, Джеймс М.; Сюй, Син; Холлидей, Кейси М. (2023). «Экоморфологические закономерности в разветвлении тройничного канала у зауропсидов обнаруживают сенсорный сдвиг у таких животных» . Журнал анатомии . 242 (5): 927–952. дои : 10.1111/joa.13826 . PMC 10093182. PMID 36680380 . S2CID 256055306 .
- ^ Новас, Фернандо Э.; Паис, Диего Ф.; Пол, Диего; Карвальо, Исмар Де Соуза; Сканферла, Агустин; Монес, Альваро; Риглос, Марио Суарес (2009). «Причудливая нотозуховая крокодилиформная форма с соответствующими яйцами из верхнего мела Боливии» . Журнал палеонтологии позвоночных . 29 (4): 1316–1320. Бибкод : 2009JVPal..29.1316N . дои : 10.1671/039.029.0409 . hdl : 11336/54001 . S2CID 140589478 .
- ^ Ву, Сяо-Чун; Сьюс, Ханс-Дитер (1996). «Анатомия и филогенетические взаимоотношения Chimaerasuchus paradoxus , необычной крокодилообразной рептилии из нижнего мела провинции Хубэй, Китай» . Журнал палеонтологии позвоночных . 16 (4): 688–702. Бибкод : 1996JVPal..16..688W . дои : 10.1080/02724634.1996.10011358 .
- ^ Jump up to: а б Оси, Аттила (2014). «Эволюция механизма челюсти и функции зубов у гетеродонтных крокодилиформ» (PDF) . Историческая биология . 26 (3): 279–414. дои : 10.1080/08912963.2013.777533 . S2CID 85707749 .
- ^ Селлерс, Калеб К.; Ньето, Мауро Николас; Дегранж, Федерико Дж.; Пол, Диего; Кларк, Джеймс М.; Миддлтон, Кевин М.; Холлидей, Кейси М. (2022). «Влияние уплощения черепа на эволюцию мышц челюсти» . Анатомическая запись . 305 (10): 2791–2822. дои : 10.1002/ar.24912 . ПМИД 35661427 . S2CID 249387665 .
- ^ Jump up to: а б Вейшампель, Дэвид Б; и др. (2004). «Распространение динозавров (средняя юра, Азия)». В: Вейшампель, Дэвид Б.; Додсон, Питер; и Осмольска, Хальшка (ред.): Динозаврия, 2-е место, Беркли: University of California Press. Стр. 541–542. ISBN 0-520-24209-2 .
- ^ Jump up to: а б с д и ж г Сюй, Син; Кларк, Джеймс М.; Эберт, Дэвид А.; Карри, Филип Дж. (2022). «Фауна Шишуго средне-позднеюрского переходного периода в Джунгарской котловине Западного Китая» . Acta Geologica Sinica — английское издание . 96 (4): 1115–1135. дои : 10.1111/1755-6724.14996 . S2CID 251934171 .
- ^ Jump up to: а б с Ното, Кристофер Р.; Гроссман, Ари (2010). «Широкомасштабные закономерности палеоэкологии позднеюрских динозавров» . ПЛОС ОДИН . 5 (9): е12553. Бибкод : 2010PLoSO...512553N . дои : 10.1371/journal.pone.0012553 . ПМЦ 2933236 . ПМИД 20838442 .
- ^ Эберт, Д.А.; Син, X .; Кларк, Дж. М. (2010). «Ямы смерти динозавров из юрского периода Китая». ПАЛЕОС . 25 (2): 112–125. дои : 10.2110/palo.2009.p09-028r . S2CID 131520314 .
- ^ Крылья, Оливер; Шварц-Уингс, Даниэла; Пфретчнер, Ганс-Ульрих; Мартин, Томас (2010). «Обзор мезозойских крокодиломорфов из Джунгарского бассейна, Синьцзян, Северо-Западный Китай, и описание изолированных крокодилоформных зубов из позднеюрского местонахождения Люхуангоу». Палеобиоразнообразие и палеосреда . 90 (3): 283–294. дои : 10.1007/s12549-010-0033-1 . S2CID 129583852 .
- ^ Сюй, Х.; Кларк, Дж. М.; Мо, Дж; Шуаньер, Дж; Форстер, Калифорния; Эриксон, генеральный директор; Хоун, Д.В.; Салливан, К; и др. (2009). «Юрский цератозавр из Китая помогает выяснить цифровую гомологию птиц» (PDF) . Природа . 459 (7249): 940–944. Бибкод : 2009Natur.459..940X . дои : 10.1038/nature08124 . ПМИД 19536256 . S2CID 4358448 .
- ^ Вейшампель, Дэвид Б; и др. (2004). «Распространение динозавров (поздняя юра, Азия)». В: Вейшампель, Дэвид Б.; Додсон, Питер; и Осмольска, Хальшка (ред.): Динозаврия, 2-е место, Беркли: University of California Press. Стр. 550–552. ISBN 0-520-24209-2 .
Внешние ссылки
[ редактировать ]- В «Эволюции крокодилов» укус был раньше тела (статья в National Geographic)
