Нундазух
| Нундазух Временной диапазон: Анисийский
~ | |
|---|---|
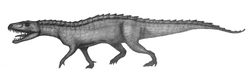
| |
| Карандашный набросок Nundasuchus songeaensis | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | Архозаврия |
| Клэйд : | Псевдозухия |
| Род: | † Нундазух Несбитт и др. , 2014 |
| Типовой вид | |
| † Nundasurus songeaensis Несбитт и др. , 2014
| |
Nundasurus — вымерший род круротарсанов , , возможно, зух архозавр- родственный Paracrocodylomorpha . Остатки этого рода известны из среднетриасовых отложений Манда на юго-западе Танзании . Он содержит единственный вид , Nundasuchus songeaensis , известный по единственному частично полному скелету , включая позвонки, элементы конечностей, остеодермы и черепа . фрагменты [ 1 ]
Нундазух жил на территории современной Танзании около в Африке 240 миллионов лет назад. Представители этого рода, вероятно, были хищниками , длиной от 2,7 до 3 метров (9 футов), с зифодонтами ( похожими на ножи для стейка ) зубами и рядами костных пластинок ( остеодермы ) вдоль спины. [ 2 ] Филогенетический анализ последовательно относит этот род к группе Crurotarsi на основании особенностей лодыжки. Большинство исследований также считают его псевдозухом , а это означает, что он был более тесно связан с современными крокодилами , чем с динозаврами . Тем не менее, нундазух имел вертикальную позицию, с ногами, расположенными прямо под телом, как и у различных других ранних псевдозухий (таких как « рауизухии » и этоозавры ), но в отличие от современных крокодилов.
Классификация Nundasurus относительно других псевдозухов несколько противоречива. Некоторые филогенетические анализы помещают его рядом или в основании группы, иногда вместе с фитозаврами , на основании определенных плезиоморфных (примитивных) особенностей, таких как зубы на небе, короткая лобковая кость и характеристики пяточной кости (пяточной кости). [ 1 ] [ 3 ] [ 4 ] Другая гипотеза, подтвержденная первоначальным описанием 2014 года, считает, что они несколько более «продвинуты», чем эти группы, вместо этого они ближе к Ticinosuchus и паракрокодиломорфам (группе, содержащей « рауизухаев » и предков современных крокодилов). Такая схема классификации обоснована наличием «шахматных» остеодерм, сердцевидных «позвоночных столиков», бороздки на головке бедренной кости . Независимо от этих гипотез, ясно, что Nundasurus представляет собой ранее неизвестную группу рептилий со смесью черт, как плезиоморфных, так и производных от таких архозавров. [ 1 ]
Открытие
[ редактировать ]
Nundasurus известен исключительно по голотипу NMT RB48, частично полностью разобщенной и большей частью разобщенной особи, хранящейся в Национальном музее Танзании , Дар-эс-Салам, Танзания . Голотип состоит из частичной правой крыловидной кости , почти полной правой зубной кости , правого слениального , правого надугольного и изолированных зубов, а также следующих посткраниальных костей: частичного атланта , двух сочлененных среднешейных позвонков , двух сочлененных средних позвонков спины. позвонки , последние задние позвонки, крестец с крестцовыми ребрами , передние хвостовые позвонки , спинной отдел ребра, гастралия , сочлененные и изолированные парамедиальные остеодермы , частичный плечевой пояс, включая межключицу , части обеих ключиц , полные левая и правая лопатки , а также правый клювовидный отросток , левая плечевая кость , обе лобковые кости , обе бедренные кости , левая малоберцовая кость , внутренний и наружный концы левой большеберцовой кости , а также левой таранной кости , левой пяточной кости , левой четвертая предплюсна и все плюсневые кости , кроме четвертой , а также внешние концы правой третьей-пятой плюсневых костей, многочисленные изолированные фаланги , частичный когтевой сустав и многие другие фрагменты. [ 1 ]
NMT RB48 был открыт доктором Стерлингом Несбиттом в 2007 году. [ 5 ] на изолированном обнажении площадью около 100 квадратных метров, известном как местоположение Z41, вместе с останками других архозавров и ринхозавров . Другое местонахождение (Z42) встречается в непосредственной близости и дало четыре цинодонта , включая две неназванные формы, Scalenodon attridgei и Mandagomphodon hirschsoni , четыре дицинодонта, включая Tetragonias njalilus , Sangusaurus parringtonii , Angonisaurus cruickshanki и Rechnisaurus cristarhynchus , а также архозавроморфов . Stenaulorhynchus stockleyi и Asilisaurus kongwe . Эти местонахождения, расположенные между реками Ндатира и Нжалила, относятся к речной озерно -аргиллитово - песчаниковой толще в середине пачки Лифуа отложений бассейна Манда в Рухуху Танзании. На основании сравнения с более изученной фауной четвероногих подзоны С Cynognathus зоны комплекса Южной Африки относят эту пачку к анизийскому этапу раннего среднего триаса . [ 1 ]
Описание
[ редактировать ]
Череп
[ редактировать ]очень ограничен Материал черепа Nundasuchus ; У голотипа сохранились только правая нижняя челюсть и правая крыловидная кость (которая составляла часть нёба). Однако даже эти фрагменты информативны. Крыловидная мышца выпуклая, если смотреть сверху, и вогнутая снизу. Эта вентральная (нижняя) вогнутость проявляется в виде глубокой впадины, ограниченной задним (задним) и медиальным (внутренним) гребнями. Часть крыловидного отростка, которая должна была располагаться вдоль средней линии неба, необычна по сравнению с другими архозаврами из-за наличия зубов. Зубы эти были маленькими и округлыми, располагались в три ряда параллельно средней линии рта. Крыловидные зубы обычно отсутствуют у архозавров, хотя известно, что некоторые авеметатарзалии ( Eudimorphodon , Eoraptor , Eodromaeus ) и еще один вероятный псевдозух ( Turfanosuchus ) обладали ими. [ 1 ]
Челюсть типична по сравнению с челюстью большинства хищных архозавров: низкая и длинная, с зазубренными ножевидными зубами, расположенными в глубоких лунках. Имелось не менее 14 зубов, причем зубы в середине челюсти (около девятого зуба) были меньше, чем зубы в передней и задней части челюсти. Нижний край боковой (внешней) стороны челюсти был покрыт небольшими продольными бороздками - особенность, по-видимому, уникальная для Nundasuchus среди ранних архозавров. Менее необычно наличие нескольких рядов ямок на латеральной поверхности и крупных борозд на медиальной поверхности. Надугольная кость в задней части челюсти образует глубоко вогнутый челюстной сустав, которому предшествует продольный гребень (уникальный для этого рода), а за ним следует большая вертикально ориентированная костная пластинка. [ 1 ]
Позвонки и остеодермы
[ редактировать ]Два не полностью сохранившихся шейных позвонка были амфицелозными (вогнутыми как спереди, так и сзади) с большой латеральной ямкой возле основания нервной дуги . Вогнутые плеврапофизы (нижние фасетки ребер) располагались вблизи нижнего переднего края позвонков, а выпуклые диапофизы (верхние фасетки ребер) располагались выше. Нижний край каждого позвонка имел большой киль, наиболее глубокий ближе к задней части. Изолированные шейные нервные отростки были похожи на таковые у Batrachotomus , расширяясь к верхним передним кончикам в структуры в форме сердца, если смотреть сверху. [ 1 ]
Большинство спинных (задних) позвонков были высокими, амфицелозными и имели пару вентральных килей, прилегающих к средней линии, разделенных неглубокой бороздкой. Эти вентральные кили похожи на кили необычной водной архозавриформы Vancleavea , хотя и не столь выражены. Учитывая, что Nundasurus не является близким родственником Vancleavea , его брюшные кили, вероятно, были примером конвергентной эволюции, и их можно считать уникальной чертой по сравнению с другими архозаврами. Фасетки ребер были короткими, располагались высоко на позвонках у основания нервных дуг. От каждого диапофиза наружу расходятся три гребня; один соединяется с парапофизом, другой соединяется с основным телом позвонка, а последний достигает постзигапофизов (задних позвоночных суставов). Одной примечательной особенностью позвонков Nundasuchus является возможное наличие сочленений гипосфен-гипантрум , как сообщается в оригинальном описании. [ 1 ] Однако исследование 2018 года поставило под сомнение эту интерпретацию, утверждая, что предполагаемые сочленения на самом деле были ошибочно идентифицированными компонентами зигапофизов. [ 6 ] Как и в случае с шейным отделом позвоночника, нервные отростки спинных позвонков расширяются наружу, образуя плоские структуры, которые иногда называют «столами позвоночника». Некоторые спинные кости возле бедра теряют характерный вентральный киль, вместо этого приобретая дополнительные ямки возле граней ребер. [ 1 ]
Крестец , вероятно, был образован двумя несросшимися позвонками, соединенными с массивными крестцовыми ребрами. Фасетки ребер в сочетании с крестцовыми ребрами огромны, но недостаточно велики, чтобы уничтожить ямки в основании нервной дуги. Каждая реберная грань первого крестцового позвонка имела отчетливое верхнее и нижнее соединение с трубчатым первым крестцовым ребром. Второе крестцовое ребро более плоское и наклонено по диагонали, так что его сужающийся задний край расположен выше переднего края. имеется бороздка По внешнему краю этого ребра, примерно там, где оно соединяется с подвздошной костью, . Крестец в целом похож на крестец рауизуха Stagonosuchus , за исключением более тонких нервных отростков. [ 1 ]
Хвостовые позвонки возле бедра были высокими и в целом похожи на спинные позвонки, за исключением того, что их зигапофизы были наклонены вверх под большим углом. Далее по хвосту они приобретают шевроны и срастаются с хвостовыми рёбрами, а также в целом становятся меньше и проще. Спинные ребра были двуглавыми, с вогнутыми и выпуклыми суставами, соответствующими выпуклым и вогнутым реберным граням позвонков. Около позвонков они были уплощены, а на концах имели небольшие фасетки для прикрепления к плотно расположенным гастралиям (брюшным ребрам). [ 1 ]
сохранилось также несколько групп остеодерм (пластинчатых костных щитков) У Nundasuchus . Отдельные остеодермы имели слегка орнаментированную форму сердца или листа, с закругленными краями и сужающимися передними кончиками. Они похожи на таковые у большинства других псевдозухий, а также на родственников архозавров, таких как Euparkeria , но менее вытянуты, чем у некоторых из этих таксонов, а имеют примерно такую же ширину, как и длину. Большинство остеодерм, отнесенных к Nundasuchus, представляют собой тонкие кости с заостренными передними кончиками, тонким продольным гребнем и значительным перекрытием. Также присутствует редкий «толстый» морфотип с меньшим перекрытием, небольшим холмообразным гребнем и более закругленными передними кончиками. По крайней мере, остеодермы «тонкого» типа образовывали несколько продольных рядов, тянущихся вдоль позвоночника животного. Если смотреть сверху, эти ряды не были бы симметричными с каждой стороны животного. Вместо этого они были «в шахматном порядке», при этом левый и правый ряды были слегка смещены друг от друга. По оценкам, на каждые два позвонка присутствовало пять остеодерм. [ 1 ]
Грудной пояс и передние конечности
[ редактировать ]Лопатка . (лопатка) была весловидной и несколько маленькой В нижней части лопатки находится гленоид (плечевая впадина), направленный назад от заднего края кости. Эта общая область является самой широкой частью кости, если смотреть спереди, и находится под самой тонкой частью, если смотреть сбоку. Непосредственно над гленоидом находится небольшая ямка, вероятно, место начала трехглавой мышцы. Между тем, передний край лопатки имеет отчетливый гребень, вероятно, акромиальный отросток. Лопатка Nundasuchus отличается от продвинутых псевдозухий небольшой ямкой для трицепса, а также от фитозавров из-за отчетливого акромиона. Нижний край лопатки соединяется с полукруглой клювовидной костью. Задняя часть клювовидного отростка отходит назад и изгибается вверх, образуя нижнюю половину гленоида, но не смещена от впадины отчетливой выемкой. Однако, если смотреть сбоку, под гленоидом имеется широкая бороздка, как и у этозавров. и Постозух . Перед гленоидом отверстие, известное как клювовидное отверстие, пронизывает клювовидный отросток, и, что является особенностью, уникальной для Nundasuchus , это отверстие окружено несколькими выступающими структурами. другие фрагменты грудного пояса, такие как межключица и ключицы Сохранились и . Они покрыты бороздками, с тонкими, похожими на лезвия краями. Межключица имеет форму ложки, вогнутая сверху и выпуклая снизу, хотя истинная форма ее неизвестна, так как задняя часть отломана. В целом он больше похож на паракрокодиломорфов, чем на фитозавров. [ 1 ]
Единственной частью передней конечности, известной для этого рода, была плечевая кость (плечевая кость). Медиальный край его был вогнутым, а латеральный — почти прямым, как у фитозавров. В связи с этим дельтопекторальный гребень (большая уплощенная структура вблизи головки плечевой кости ) ориентирован скорее вперед, чем латерально. И головка плечевой кости, и верхушка нижнего конца кости были покрыты глубокими бороздками. Чуть выше нижней конечности было углубление, а сразу латеральнее этого углубления находилась отчетливая эктепикондилярная борозда. [ 1 ]
Тазовый пояс и задние конечности
[ редактировать ]Единственной сохранившейся частью тазового пояса (бедра), не считая крестца, была левая лобковая кость. Эта кость была характерно маленькой у Nundasuchus , составлявшей всего около 30% длины бедренной кости. Это сравнимо с фитозаврами, этозаврами и ранними авеметатарсалиями, но в отличие от состояния других архозавров. Области, где лобковая кость соприкасалась с другими костями бедра, сломаны, но, очевидно, лобковая кость имела такие особенности, как вставленное запирательное отверстие и место прикрепления окружающей мышцы под вертлужной впадиной (тазовая впадина). Основная часть лобковой кости не сильно расширена в большинстве направлений, но медиальный край имеет пластинчатое внутреннее расширение, известное как лобковый фартук, которое контактировало с соответствующим расширением на правой лобковой кости. Этот лобковый фартук выпуклый, если смотреть спереди. [ 1 ]
Бедренная кость (бедренная кость) имеет сигмовидную форму (S-образную форму), головка бедренной кости слегка вывернута внутрь, а дистальные мыщелки слегка вывернуты наружу. Это создает угол в 45 градусов между этими двумя концами кости, как и у большинства еврокоподов, за исключением динозавров. Как и у других еврокоподов, головка бедренной кости имеет несколько отчетливых тубер (шишек), один на передне-латеральной поверхности кости, а другой - на задне-медиальной поверхности. У Нундазухуса также мог быть третий бугор на медиальной поверхности головы, как у архозавров , но эта часть кости повреждена, поэтому ее невозможно оценить с уверенностью. Головка бедренной кости также имеет на верхней поверхности бороздку, как у паракрокодиломорфов. Далее вниз по заднемедиальной поверхности диафиза находится длинный острый гребень, известный как четвертый вертел . Около 40% передней части стержня занимает клубень, покрытый бороздками. Эта шишка, вероятно, была точкой прикрепления подвздошно-феморальной мышцы, которая помогала стабилизировать бедро. В дистальной части бедренной кости имеется большой crista tibiofibularis, верхнее продолжение латеральный мыщелок . И криста, и медиальный мыщелок большие и сужаются к закругленным вершинам. Кроме того, латеральные и медиальные мыщелки имеют небольшое углубление на своих концах, причем углубление медиального мыщелка, вероятно, соответствует бедренно -большеберцовой мышце. [ 1 ]
Проксимальная часть большеберцовой кости (внутренняя большеберцовая кость) имеет примерно ромбовидную форму в поперечном сечении, с закругленными латеральными и медиальными туберами, а также низким и нечетким направленным вперед гребнем, известным как кнемиальный гребень. Боковой клубень крупнее и более вогнутый, как у орнитозухид. Помимо этих тубер, проксимальная часть голени преимущественно выпуклая. Однако позади бокового клубня имеется отчетливая выемка. Вероятно, это место прикрепления внутреннего сгибателя большеберцовой мышцы. Дистальная часть гладкая, овальной формы в поперечном сечении и «согнута», если смотреть сзади. Малоберцовая кость (наружная большеберцовая кость) имеет сигмовидную форму, с уплощенной медиальной поверхностью, большим гребнем подвздошно -берцовой мышцы на латеральной поверхности и наклоненной вперед дистальной поверхностью. [ 1 ]
Стопа состояла из пяти плюсневых костей, прикрепленных к фалангам (костям пальцев ног). Самая маленькая плюсневая кость была самой внутренней (I), при этом II плюсневая кость была длиннее, а III длиннее любой из них. Хотя IV плюсневая кость неполная, по оценкам, она длиннее III. Это было бы весьма необычно среди рептилий триаса, из которых только Proterosuchus обладает этой характеристикой. I плюсневая кость несколько сплющена по диагонали, ее край, обращенный к остальной части стопы, расположен выше внутреннего края. На внутреннем крае также присутствует гребень. Сочленение с голеностопным суставом выпуклое и расширенное к остальной части стопы. II плюсневая кость имеет в поперечном сечении форму параллелограмма с «губой» на верхнем крае, принимающей первую плюсневую кость. На III плюсневой кости самый большой дистальный сустав, а на IV — самый маленький, что указывает на то, что третий палец ноги был более массивным, чем четвертый. Пятая плюсневая кость имеет крючковидную форму с двумя отчетливыми проксимальными сочленениями малоберцовой кости и четвертым дистальным сочленением лодыжки. Фаланги в целом крепкие, но фаланги пятого пальца были длиннее и имели форму песочных часов, прикрепленные к грубой текстуре. когтевой (коготь). В целом кости стопы напоминают кости престозухов и фитозавров. [ 1 ]
Лодыжка
[ редактировать ]
Голеностопный сустав в основном образован двумя костями: пяточной костью на внешней стороне и таранной костью на внутренней стороне. Эти кости и их суставы ясно демонстрируют отличительные черты группы Crurotarsi, в которую входят зухи, фитозавры и, вероятно, авеметатарзалии. Например, место прикрепления пяточной кости к таранной кости вогнутое, а пяточная кость также имеет цилиндрическое расширение с расширяющимися краями, известное как пяточный бугор. Пяточный бугор направлен примерно на 45 градусов между боком и назад, подобно фитозаврам и неархозавровым эвкрокоподам, но в отличие от большинства других таких, у которых пяточный бугор ориентирован более назад. С другой стороны, длина пяточного бугра умеренная, похожая на таковую у типичных псевдозухий, а не на гораздо более удлиненную структуру фитозавров. На боковой стороне клубня имеется заметная «подушечка», которая, по-видимому, уникальна для Nundasurus . Верхняя поверхность пяточной кости соединяется с малоберцовой костью выпуклым бочкообразным суставом. Это также похоже на фитозавров и зухов, но в отличие от орнитозухид, у которых этот сустав более куполообразный. И малоберцовая, и астрагаларная фасетки пяточной кости представляют собой сплошную поверхность, в отличие от таких костей. [ 1 ]
Верхняя поверхность таранной кости имеет треугольную фасетку для малоберцовой кости, а также более крупную фасетку овальной формы для большеберцовой кости. Фасетка большеберцовой кости разделена на две впадины слабой выпуклостью, хотя это сгибание несколько невыражено по сравнению со сгибанием зухий и авеметатарсалий. Пяточная фасетка имеет форму плоской поверхности, покрывающей выпуклый «колышек», что является особенностью голеностопных суставов. Колышек развит слабо, больше похож на таковой у фитозавров, чем у других таких животных. Передний край таранной кости имеет большую вогнутую поверхность (астрагаларная впадина), лежащую над небольшой выпуклостью (астрагаларный шарик). Астрагаларный шар примерно так же хорошо развит, как и у этозавра Longosurus . [ 1 ]
Этимология
[ редактировать ]Nundasurus был впервые описан и назван Стерлингом Дж. Несбиттом, Кристианом А. Сидором, Кеннетом Д. Ангельчиком, Роджером М. Х. Смитом и Линдой А. Цуджи в 2014 году , типовым видом является Nundasuchus songeaensis . Родовое название происходит от на языке суахили слова nunda , что означает « хищник », а также от греческого soukhos , египетского бога-крокодила. Новое видовое название относится к столице провинции Сонгеа , расположенной недалеко от места сбора останков, на что указывает латинский суффикс -ensis , означающий «от». [ 1 ]
Классификация
[ редактировать ]Первоначальное описание Nundasurus, Nesbitt et al. (2014) проверили его филогенетическое положение, используя два наиболее полных доступных набора филогенетических данных ранних архозавров . Первым использованным набором данных была версия Брусатте и др. 2010 года. филогении архозавров [ 7 ] который был обновлен Батлером и др. (2011) во время исследования анатомии Ctenosauriscus . [ 8 ] Второй набор данных представлял собой более полное и широко признанное исследование взаимоотношений архозавров, созданное Несбиттом в 2011 году. [ 9 ] Оба анализа показали, что Нундазух был псевдозухом , то есть архозавром, более близким к крокодилам, чем к динозаврам. Эта идентификация была в первую очередь основана на чертах лодыжки, которые были общими у Nundasuchus и псевдозухий. Брусатте и др. В наборе данных (2010) он оказался самым базальным псевдозухием (хотя для группы использовалось название «Crurotarsi»), менее направленным к короне (дальше от крокодилов), чем даже фитозавры . [ 1 ]
Несбитт и др. Применение (2014) набора данных Nesbitt (2011) поместило Nundasuchus в более высокое положение по сравнению с его положением в Brusatte et al. результаты. Несбитт (2011) считал, что крокодиломорфы (предки крокодилов) произошли от таксонов, традиционно называемых « рауизухаями ». Поскольку Rauisuria обычно не включает крокодиломорфов и, следовательно, были бы парафилетическими результаты , Несбитт решил отказаться от этого названия, заменив его монофилетической кладой Paracrocodylomorpha . Сама эта клада родственна швейцарскому суциану Ticinosuchus, прежде всего, на основании сходства седалищной кости. [ 9 ] Когда Nundasurus был добавлен в набор данных, он был помещен в качестве сестринского таксона кладе Ticinosurus + Paracrocodylomorpha. Это оправдывалось тремя чертами: сердцевидными «столбиками позвоночника», бороздкой на проксимальной стороне головки бедренной кости и «шахматным» остеодермом. [ 1 ]
Несбитт и др. (2014) поставили под сомнение эти результаты в связи с тем, что Nundasurus обладает многими плезиоморфными чертами архозавров (т.е. типичными для очень базальных архозавров), которые кажутся аутапоморфиями (уникальными особенностями), если он действительно является членом Pseudosuria . К ним относятся небные зубы, короткий лобок и не столь сильно отклоненный назад пяточный бугор. Таким образом, они провели дальнейший анализ при определенных ограничениях взаимоотношений, таких как принуждение Nundasuchus находиться за пределами Archosauria или ближе к Phytosauria. Этот анализ выявил небольшие изменения в отношениях (кроме самих ограничений), но серьезные изменения в признаках, связанных с этими отношениями, изменение признаков, которые считались диагностическими для Pseudosuria, Archosauria и других клад . Таким образом, неясно, является ли Nundasuchus зухием, имеющим сходство с гораздо более базальными архозаврами, как предполагают два филогенетических анализа, или архозавроформой , тесно связанной с Archosauria, с чертами, сходящимися с морфологией более продвинутых паракрокодиломорфов. Однако поддержка размещения Nundasurus действительно ослабевает по мере удаления от Paracrocodylomorpha. Результаты неограниченного анализа Nesbitt et al. (2014) с использованием набора данных Nesbitt (2011) упрощены ниже (отношения внутри выделенных жирным шрифтом клад не показаны). [ 1 ]

В 2016 году Nundasurus был включен в исследование архозавроморфов, проведенное Мартином Эскуррой , и подвергся филогенетическому анализу, аналогичному по размеру и охвату Несбитту (2011). В отличие от результатов Nesbitt et al. (2014), Ezcurra (2016) отнесли Nundasuria к базальным псевдозухиям, хотя и не настолько базальным, как Phytosauria. Он также обсудил три особенности, которые эти авторы использовали для обоснования размещения Nundasuchus рядом с Paracrocodylomorpha. Например, считалось, что столы позвоночника в форме сердца имели беспорядочное распространение в пределах Pseudosuria: у некоторых таксонов (таких как грацилизухиды ) они отсутствовали, а у других (например, у фитозавров и Batrachotomus ) они были только в определенных позвонках. Аналогичным образом, шахматные дорсальные остеодермы были обнаружены только у Nundasuchus и Gracilisuchus , причем ни один из выбранных паракрокодиломорфов не имел этого признака. Он признал, что бороздка на головке бедренной кости согласуется с гипотезой о Nundasurus к паракрокойломорфам, поскольку этот признак также обнаружен у близости Престозух и Батрахотомус . [ 3 ]
Однако Эскурра также признал, что его анализ был больше сосредоточен на базальных архозавроморфах и архозавриформах, а не на настоящих архозаврах, и поэтому он может быть не совсем точным для групп, обращенных к короне, таких как псевдозухи. Он отметил, что некоторые таксоны, имеющие решающее значение для результатов Несбитта (2011), такие как Ticinoзарух и Заурозух , были исключены из его исследования. Эти роды были паракрокодиоморфами, которые обладали расположенными в шахматном порядке остеодермами, сердцевидными позвоночными пластинами и бороздкой на головке бедренной кости, что оправдывало близкое родство с Nundasuchus . В 2018 году еще один ранний паракрокодиломорф — Mandasurus долгожданное официальное описание получил . У этого рода также были эти три особенности, определенные Nesbitt et al. (2014). В описание был включен филогенетический анализ, основанный главным образом на Несбитте (2011), но также включающий новые данные из описания Nundasuchus , а также недавнего исследования грацилизухид. Результаты показали, что либо Nundasurus , либо gracilisuchids были сестринскими таксонами Paracrocodylomorpha. Тицинозух , хотя и не смог точно определить, какой из них ближе. [ 10 ]
Обновленная версия набора данных Nesbitt (2011), опубликованная Da-Silva et al . (2018) поместили Нундазухов в основание Псевдозухий. В это исследование вошли многие недавние изменения и дополнения к методологии Несбитта, хотя его результаты больше напоминали результаты Эзкурры (2016), а не Несбитта (2014). [ 4 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и Несбитт, Стерлинг Дж.; Сидор, Кристиан А.; Ангельчик, Кеннет Д.; Смит, Роджер М.Х.; Цудзи, Линда А. (ноябрь 2014 г.). «Новый архозавр из пластов Манда (анисий, средний триас) на юге Танзании и его значение для оптимизации состояния признаков в Archosauria и Pseudosuria». Журнал палеонтологии позвоночных . 34 (6): 1357–1382. Бибкод : 2014JVPal..34.1357N . дои : 10.1080/02724634.2014.859622 . S2CID 129558756 .
- ^ «Nundasuchus songeaensis: в Танзании обнаружена новая триасовая рептилия | Палеонтология | Sci-News.com» . Последние научные новости | Sci-News.com . 21 января 2015 г.
- ^ Jump up to: а б Эскурра, Мартин Д. (28 апреля 2016 г.). «Филогенетические взаимоотношения базальных архозавроморфов с акцентом на систематике протерозуховых архозавроформ» . ПерДж . 4 : е1778. дои : 10.7717/peerj.1778 . ISSN 2167-8359 . ПМЦ 4860341 . ПМИД 27162705 .
- ^ Jump up to: а б Лусио Роберто-Да-Сильва; Родриго Темп Мюллер; Марко Аурелио Галло из Франции; Сержиу Фуртаду Кабрейра; Сержио Диас-Да-Сильва (2018). «Впечатляющий скелет гигантского высшего хищника Prestosuchus chiniquensis (Pseudosuria: Loricata) из триаса на юге Бразилии, с физиологическими данными» . Историческая биология: Международный журнал палеобиологии . 32 (7): 1–20. дои : 10.1080/08912963.2018.1559841 . S2CID 92517047 .
- ^ Гриффин, Эндрю (21 января 2015 г.). «Огромная рептилия, похожая на крокодила, бродила по Земле до того, как динозавры захватили ее» . Независимый . Проверено 21 января 2015 г.
- ^ Стефаник, Кэндис М.; Несбитт, Стерлинг Дж. (14 февраля 2018 г.). «Осевой скелет Poposaurus langstoni (Pseudosuchia: Poposauroidea) и его значение для эволюции добавочного межпозвонкового сочленения у псевдозуховых архозавров» . ПерДж . 6 : е4235. дои : 10.7717/peerj.4235 . ISSN 2167-8359 . ПМЦ 5816584 . ПМИД 29472991 .
- ^ Брусатте, Стивен Л.; Бентон, Майкл Дж.; Дезохо, Джулия Б.; Лангер, Макс К. (15 марта 2010 г.). «Филогения высшего уровня Archosauria (Tetrapoda: Diapsida)» . Журнал систематической палеонтологии . 8 (1): 3–47. Бибкод : 2010JSPal...8....3B . дои : 10.1080/14772010903537732 . hdl : 20.500.11820/24322ff3-e80e-45f2-8d53-d35fd104195c . ISSN 1477-2019 . S2CID 59148006 .
- ^ Батлер, Ричард Дж.; Брусатте, Стивен Л.; Райх, Майк; Несбитт, Стерлинг Дж.; Шох, Райнер Р.; Хорнунг, Ян Дж. (14 октября 2011 г.). «Парусная рептилия Ctenosauriscus из последнего раннего триаса Германии, а также время и биогеография излучения ранних архозавров» . ПЛОС ОДИН . 6 (10): e25693. Бибкод : 2011PLoSO...625693B . дои : 10.1371/journal.pone.0025693 . ISSN 1932-6203 . ПМК 3194824 . ПМИД 22022431 .
- ^ Jump up to: а б Несбитт, SJ (2011). «Ранняя эволюция архозавров: взаимоотношения и происхождение основных клад» . Бюллетень Американского музея естественной истории . 352 : 1–292. дои : 10.1206/352.1 . hdl : 2246/6112 . S2CID 83493714 .
- ^ Батлер, Ричард Дж.; Несбитт, Стерлинг Дж.; Чариг, Алан Дж.; Гауэр, Дэвид Дж.; Барретт, Пол М. (29 ноября 2017 г.). « Mandasuchus tanyauchen , gen. et sp. nov., псевдозуховый архозавр из слоев Манда (? Средний триас) Танзании» (PDF) . Журнал палеонтологии позвоночных . 37 (суп1): 96–121. Бибкод : 2017JVPal..37S..96B . дои : 10.1080/02724634.2017.1343728 . ISSN 0272-4634 . S2CID 90164051 .
