Сцелидозавр
| Сцелидозавр | |
|---|---|

| |
| Сцелидозавр , слепок экземпляра Дэвида Соула BRSMG LEGL 0004, Юта. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Сафорнитишия |
| Клэйд : | † Геназаврия |
| Клэйд : | † Тиреофора |
| Род: | † Сцелидозавр Оуэн , 1859 г. |
| Разновидность: | † С. Харрисони
|
| Биномиальное имя | |
| † Сцелидозавр Харрисони Оуэн, 1861 г.
| |
Сцелидозавр ( / ˌ s ɛ l ɪ d oʊ ˈ s ɔːr ə s / ; с предполагаемым значением «ящерица конечностей», от греческого skelis / σκελίς, означающего «говяжье ребро», и sauros / σαυρος, означающего «ящерица») [ 1 ] — род травоядных панцирных птицетазовых динозавров из юрского Англии периода .
Сцелидозавр жил в раннем юрском периоде , от синемурийского до плинсбахского периода, около 191 миллиона лет назад . Этот род и родственные ему роды в то время обитали на суперконтиненте Лавразия . Его окаменелости были найдены в формации Чармутского аргиллита недалеко от Чармута в Дорсете , Англия, и эти окаменелости известны своей превосходной сохранностью. Сцелидозавр считается одним из самых ранних полноценных динозавров. [ 2 ] и является одним из наиболее известных динозавров Британских островов . Несмотря на это, современное описание материализовалось только в 2020 году. После первоначальных находок в 1850-х годах сравнительный анатом Ричард Оуэн назвал и описал Scelidosaurus в 1859 году. Только один вид , Scelidosaurus harrisonii, названный Оуэном в 1861 году, сегодня считается действительным, хотя еще один вид было предложено в 1996 году.
Сцелидозавр имел длину около 4 метров (13 футов). Это было в основном четвероногое животное, питавшееся низкими кустистыми растениями, части которых откусывались за небольшую вытянутую голову для переработки в большую кишку. Сцелидозавр был легкобронирован, защищен длинными горизонтальными рядами килевидных овальных щитков, которые тянулись вдоль шеи, спины и хвоста.
в этой группе, одного из старейших известных и наиболее «примитивных» тиреофоров , Точное место Scelidosaurus было предметом споров на протяжении почти 150 лет. Этому не способствовали ограниченные дополнительные знания о ранней эволюции панцирных динозавров. Сегодня большинство данных свидетельствуют о том, что Scelidosaurus является наиболее производным из известных базальных тиреофоров и тесно связан либо с Ankylosauria , либо с Stegosauria + Ankylosauria.
Описание
[ редактировать ]Размер и поза
[ редактировать ]
Взрослый сцелидозавр был довольно маленьким по сравнению с большинством более поздних нептичьих динозавров, но в ранней юре это был вид среднего размера. Некоторые ученые оценили длину в 4 метра (13 футов). [ 3 ] В 2010 году Грегори С. Пол дал длину тела 3,8 метра (12,5 футов) и вес 270 кг (600 фунтов). [ 4 ] Сцелидозавр был четвероногим , его задние конечности длиннее передних. Возможно, он поднимался на задние лапы, чтобы пощипать листву с деревьев, но его руки были относительно длинными, что указывало на то, что он в основном находился на четвероногих ногах. [ 5 ] Тропа из гор Святого Креста в Польше показывает скелидозавра, похожего на животное, идущего на двух ногах, что намекает на то, что сцелидозавр , возможно, был более опытным в прямохождении на двух ногах, чем считалось ранее. [ 6 ]
Отличительные черты
[ редактировать ]Первый современный диагноз был поставлен Дэвидом Брюсом Норманом в 2020 году. В первой статье Норман представил аутапоморфии , уникальные производные признаки черепа. Передние кости морды, предчелюстные кости, имеют общее центральное шероховатое расширение и при жизни имеют небольшой верхний клюв. Носовая кость имеет на своей верхней внешней стороне фасетку, касающуюся внутренней стороны восходящей ветви предчелюстной кости. Анторбитальное окно представляет собой бобовидное углубление, нижний край которого образован острым гребнем. Центральный теменный гребень крыши черепа образован двумя параллельными гребнями, разделенными узким впадиной по средней линии. Крыша полости носа образована специальными пластинками над сошниками, называемыми «эпивомерами». Эпиптеригоидная кость имеет форму небольшой конической вертикальной структуры, основание которой соединяется с верхней стороной крыловидной кости посредством латеральной плоской поверхности. Базиокципиталь имеет на нижних сторонах крупные косые фасетки. Опистотик имеет расширенную ножку с гранями на нижней стороне. Удлиненные эпистилоидные кости выступают косо назад и вниз, от задней части черепа. Небольшая шпорообразная структура на верхнем крае парокципитального отростка окружает поствисочное окно. Задняя часть черепа по верхнему краю сращена с парой крупных изогнутых роговидных остеодерм. На нижней челюсти имеется лишь небольшой экзостоз, ограниченный угловой частью, без прикрепленной остеодермы. [ 7 ]
Череп
[ редактировать ]
Голова сцелидозавра была небольшой, длиной около двадцати сантиметров, вытянутой формы. Череп был низким при виде сбоку и треугольным при виде сверху, длиннее, чем ширина , как у более ранних птицетазовых животных . Морда, в основном образованная носовыми костями , сверху была плоской. У сцелидозавра все еще было пять пар фенестров (отверстий в черепе), которые наблюдались у базальных птицетазовых животных: за исключением ноздрей и глазниц, которые присутствуют у всех базальных динозавров, fenestra antorbitalis , а также верхние и нижние височные отверстия не были закрыты и не заросли, как у сцелидозавра. многие более поздние бронированные формы. Фактически, верхние височные окна были очень большими, образуя заметные круглые отверстия в верхней части задней части черепа, служившие местами прикрепления мощных мышц, закрывавших нижнюю челюсть. Глазница в передней части была слегка затемнена надбровной дугой, которую воспринимали как предлобную кость . В 2020 году Норман пришел к выводу, что это сросшаяся глазная кость . [ 7 ] Позади нее верхний край глазницы образовывался надглазничной костью . Исследование Сюзанны Мейдмент и др. пришло к выводу, что молодые экземпляры показывают, что эта кость представляла собой слияние трех элементов: одного спереди, другого сзади и третьего на внутренней стороне. [ 8 ]
, Предчелюстная кость образующая кончик морды, была короткой и не имела предзубных костей ; на кончике толстой нижней челюсти был обнаружен костный стержень нижнего клюва, поэтому роговой клюв, который, как предполагается, присутствует у всех птицетазовых, вероятно, был очень короткий. Его зубы были длиннее и более треугольными при взгляде сбоку, чем у более поздних панцирных динозавров. [ 9 ] В каждой предчелюстной кости было не менее пяти зубов, на верхней челюсти не менее девятнадцати и на зубной кости нижней челюсти не менее шестнадцати. [ 5 ] Однако количество верхнечелюстных и зубных зубов было установлено по неполному черепу одного из первых найденных экземпляров; фактическое число могло достигать примерно двух дюжин, возможно, двадцати шести для нижней челюсти. Предчелюстные зубы были несколько длиннее и загнуты назад. Кзади они постепенно приближаются к форме верхнечелюстных зубов, начиная проявлять зубчики . Коронки верхних и зубных зубов имеют зубцы по краям и вздутое основание. [ 5 ]
Восходящие ветви парных предчелюстных костей зазубривали объединенные носовые кости, тогда как у птицетазовых было обычно наоборот. Лобные кости были покрыты ореолом тонких гребней; это указывает на наличие ороговевших пластинок, как у современных черепах . топорообразной формы В передней части черепной коробки парные окостеневшие орбитосфеноиды образовывали дно обонятельных долей мозга. Череп лектотипа был поврежден палеоихтиологом, в результате чего от неба отслоилась треугольная пластинка. Эти элементы были зарисованы Норманом в семидесятые годы до инцидента и интерпретированы как части крыловидных костей, но в 2020 году он пришел к выводу, что это особые кости, покрывающие крышу носовой полости, которые он назвал «эпивомерами». Они не известны ни у одного другого животного. [ 7 ]
Посткраниальный скелет
[ редактировать ]
сцелидозавра В позвоночнике было спинных не менее шести шейных позвонков, семнадцати позвонков , четырех крестцовых позвонков и не менее тридцати пяти хвостовых позвонков. [ 5 ]
Хотя, возможно, общее количество шейных позвонков достигало семи или восьми, шея была лишь умеренно длинной. Туловище . было относительно плоским при виде сбоку, однако, несмотря на ширину брюха, оно не было сильно сжато по вертикали, как у анкилозавров, а было выше его ширины Последние три спинных позвонка не имели ребер. Отростки крестцовых позвонков соприкасались друг с другом, но не срастались в надневральную пластинку. Быстро сужающийся хвост был относительно коротким и, вероятно, составлял около половины длины тела. Хвостовые шевроны были сильно наклонены назад. Область бедра и основание хвоста были усилены большим количеством окостеневших сухожилий. [ 5 ]
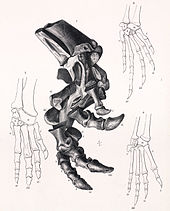
Лопатка . короткая, с умеренно расширенным верхним концом Коракоид имел круглую форму при виде сбоку. Элементы передних конечностей обычно были умеренно длинными, прямыми и толстыми. Рука известна только благодаря недавним открытиям и еще не описана. При довольно широком тазе при подвздошная кость виде сбоку прямая. Его переднее лезвие имело стержнеобразную форму и умеренно расширялось наружу, оставляя место для брюшка. Это было усилено тем, что крестцовые ребра стали длиннее спереди. Крестцовые ребра были шире в местах прикрепления к подвздошной кости, но не срастались в крестцовое ярмо. был На лобке короткий препубис . Лобковая кость была прямой, идущей параллельно прямой кости седалищной кости, уплощенной в поперечном направлении на нижнем конце. Бедренная кость при виде сбоку была прямой, при виде спереди несколько выгнута наружу. Его голова не была отделена от древка настоящей шеей. В то время как большой вертел находился примерно на одном уровне с головкой, нижний малый вертел был отделен от обоих глубокой расщелиной. На задней стороне В средней части бедренной кости имеется хорошо развитый опущенный четвертый вертел, отросток прикрепления хвостовой мышцы-ретрактора, Musculus caudofemoralis longus . Голень была несколько короче бедренной кости. Большеберцовая кость имела широкий верхний конец с кнемиальным гребнем хорошо выступающим вперед . Нижний конец большеберцовой кости также был прочным и поворачивался примерно на 70° по сравнению с верхней частью, сильно поворачивая стопу наружу. Нога была очень большой и широкой. Пятая плюсневая кость была лишь рудиментарной, но остальные четыре были крепкими. У сцелидозавра было четыре больших пальца на ногах, причем самый внутренний палец был самым маленьким. Четвертая плюсневая кость была короткой, но ее палец был длинным и располагался наружу для улучшения устойчивости. Когти были плоскими, копытообразными и загнутыми внутрь. [ 5 ]
Броня
[ редактировать ]
Наиболее очевидной особенностью сцелидозавра является его панцирь, состоящий из костных щитков , встроенных в кожу. Эти остеодермы располагались горизонтальными параллельными рядами вдоль тела животного. [ 3 ] Остеодермы сегодня встречаются в коже крокодилов , броненосцев и некоторых ящериц . Остеодермы сцелидозавра различались как по размеру, так и по форме. Большинство из них представляли собой овальные пластины меньшего или большего размера с высоким килем снаружи, самая высокая точка киля располагалась ближе к задней части. Некоторые щитки были маленькими, плоскими и полыми внутри. Более крупные килеватые щитки располагались в правильные горизонтальные ряды. Их было по три ряда вдоль каждой стороны туловища. Щитки нижнего, бокового, ряда были более коническими, чем пластинчатые остеодермы Scutellosaurus . [ 10 ] Между этими основными сериями присутствовал один или два ряда более мелких овальных килевидных щитков. Всего на хвосте было четыре ряда крупных щитков: один на верхней средней линии, один на средней линии нижней стороны и по одному на каждой стороне хвоста. Продолжались ли средние хвостовые щитки по туловищу и шее вперед, неизвестно и маловероятно для шеи, хотя сцелидозавра часто изображают таким.

На шее имелось по два ряда крупных щитков с каждой стороны. Остеодермы нижнего шейного ряда очень крупные, плоские, пластинчатые. Первые остеодермы верхних рядов шеи образовывали пару своеобразных трехконечных щитков непосредственно за головой. Эти точки, по-видимому, были соединены сухожилиями с задними суставными отростками, постзигапофизами осевого позвонка. [ 5 ] В целом щитки были крупнее в передней части туловища, остеодермы уменьшались к задней части, особенно на поверхности бедер. Мельчайшие плоские круглые щитки могли заполнить пространство между более крупными рядами остеодермы. Возможно, на плечах присутствовал ряд вертикальных остеодерм. По сравнению с более поздними анкилозаврами , сцелидозавр имел легкую броню, без сплошных пластин, шипов или тазового щита. Шершавые участки на черепе и нижней челюсти указывают на наличие оссификаций кожи.
Некоторые из последних найденных экземпляров демонстрируют частично разные остеодермы, включая щитки, у которых киль больше похож на шип или шип. У этих экземпляров также имеются небольшие рога в задних углах головы, расположенные на чешуйчатых костях . [ 11 ]
Также были обнаружены окаменелые отпечатки кожи. Между костными щитками у сцелидозавра были округлые неперекрывающиеся чешуи, как у нынешнего монстра Гила . [ 3 ] Между большими щитками в коже, возможно, были распределены очень мелкие (5-10 миллиметров [0,2-0,4 дюйма]) плоские «гранулы» кости. У более поздней Ankylosauria эти маленькие щитки, возможно, развились в более крупные щитки, слившись в мультиостеодермальный пластинчатый панцирь, наблюдаемый у таких родов, как Ankylosaurus . [ 10 ]
История обучения
[ редактировать ]Открытие, наименование и типовые образцы
[ редактировать ]
В 1850-х годах владелец карьера Джеймс Харрисон из Чармута , Западный Дорсет, Англия, нашел окаменелости в скалах Блэк-Вен между Чармутом и Лайм-Реджисом , которые добывались, возможно, в качестве сырья для производства цемента . Некоторые из них он подарил коллекционеру и хирургу на пенсии Генри Норрису. В 1858 году Норрис и Харрисон отправили несколько фрагментов костей конечностей профессору Ричарду Оуэну из Британского музея (естественной истории) в Лондоне (сегодня Музей естественной истории ). Среди них была левая бедренная кость, образец GSM 109560. В 1859 году Оуэн назвал род Scelidosaurus в статье о палеонтологии в Британской энциклопедии . [ 12 ] В тексте леммы содержался диагноз, подразумевающий, что род назван правильно и не является nomen nudum , несмотря на то, что определение было расплывчатым и экземпляры не были идентифицированы. [ 13 ] Оуэн намеревался назвать динозавра «завридом с задними конечностями», но перепутал греческое слово σκέλος, skelos , «задняя конечность», с σκελίς, skelis , «говяжье ребро». [ 14 ] [ 15 ] Название было навеяно сильным развитием задней ноги. Впоследствии Харрисон отправил Оуэну коленный сустав, коготь (GSM 109561), молодой экземпляр и череп, которые были описаны в 1861 году. Тогда типовой вид Scelidosaurus harrisonii был назван - видовое название в честь Харрисона. [ 14 ] Позже выяснилось, что череп является частью почти полного скелета, описанного Оуэном в 1863 году. [ 16 ]
Британский палеонтолог Дэвид Брюс Норман подчеркнул, насколько примечательно то, что Оуэн, который ранее утверждал, что динозавры были активными четвероногими животными, в значительной степени пренебрегал Scelidosaurus , хотя он мог служить ярким примером этой гипотезы, а его окаменелость была одним из наиболее полных найденных динозавров. в это время. Норман объяснил это чрезмерной нагрузкой Оуэна в этот период, включавшей выполнение нескольких административных функций, полемику с коллегами-учеными и изучение большого количества еще более интересных недавно открытых вымерших животных, таких как археоптерикс . [ 17 ] Норман также отметил, что Оуэн в 1861 году предложил образ жизни сцелидозавра , который сильно отличался от нынешних представлений: он должен был питаться рыбой и частично обитать в море. [ 2 ] [ 14 ]

Оуэн не указал голотип . В 1888 году Ричард Лидеккер при каталогизации окаменелостей NHMUK обозначил некоторые из фрагментов задних конечностей, описанных в 1861 году, экземпляр NHMUK PV 39496, состоящий из нижней части бедренной кости и верхней части большеберцовой и малоберцовой костей, вместе образующих коленный сустав, как типовой экземпляр, тем самым неявно выбрав его в лектотипа сцелидозавра качестве . Лидеккер не объяснил причины своего выбора; [ 18 ] возможно, его мотивировал их больший размер. К сожалению, среди окаменелостей сцелидозавра были смешаны частичные останки тероподного динозавра, и, таким образом, бедренная кость и голень принадлежали такому хищнику; это было обнаружено только в 1968 году Бернардом Ньюманом . [ 19 ] В том же году Б. Х. Ньюман предложил, чтобы выбор Лидеккера коленного сустава в качестве лектотипа был официально аннулирован Международной комиссией по зоологической номенклатуре , поскольку, по его мнению, этот сустав принадлежал к виду, родственному мегалозавру . В конце концов, после того как Ньюман уже умер, Алан Джек Чариг действительно подал запрос в 1992 году. [ 13 ] В 1994 году ICZN отреагировал положительно, в Мнении 1788, решив, что череп и скелет, образец NHMUK PV R.1111 , будут новым лектотипом сцелидозавра . [ 20 ] Коленный сустав был описан в 1995 году Сэмюэлем Уэллсом и др. неофициально отнесен к « Мерозавру », имя которого еще официально не опубликовано. [ 21 ] Скорее всего, он принадлежит какому-то представителю Coelophysoidea или Neoceratosauria . [ 22 ] Ньюман также установил и подтвердил Роджер Бенсон , что первоначальная левая бедренная кость, GSM 109560, принадлежала тероподу. [ 22 ]

Скелет нового лектотипа был обнаружен в пласте черного мергеля или слоя вудстоунских конкреций, морских отложениях формации Чармут-аргиллит , относящихся к позднему синемурийскому этапу, около 191 миллиона лет назад. [ 23 ] Он состоит из довольно полного скелета с черепом и нижними челюстями. Отсутствуют только кончик морды, основание шеи, передние конечности и хвост. Рядом со скелетом были обнаружены сотни остеодерм, многие из которых находились более или менее в исходном положении. Начиная с 1960-х годов, Рональд Краучер обрабатывал это ископаемое с использованием кислотных ванн для освобождения костей от окружающей матрицы - метод, усовершенствованный для окаменелостей Чармута. В 1992 году Чариг сообщил, что еще предстоит лечить только один блок. [ 13 ] но он умер до того, как результаты были опубликованы. Норман, который намеревался выполнить эту задачу, в 2004 году раскрыл некоторые новые анатомические детали. [ 5 ] Помимо этого, современное описание практически отсутствовало. [ 23 ] В 2020 году Норман опубликовал статьи о черепе и посткрании, приняв во внимание и более поздние находки. Выяснилось, что кислотные ванны из-за протечек серьезно ухудшили состояние костей, а дальнейшее неправильное обращение привело к их поломке и разрушению. [ 7 ]
Дополнительные образцы
[ редактировать ]
Помимо лектотипа, известны и другие окаменелости сцелидозавра . В 1888 году Лидеккер каталогизировал большое количество отдельных костей, в основном элементов конечностей и остеодерм, которые были приобретены НХМУК из коллекции Норриса. [ 18 ] Оуэн в 1861 году описал второй, частичный скелет молодого животного, который позже был добавлен в коллекцию Элизабет Филпот и сегодня зарегистрирован в музее Лайм-Реджиса как образец LYMPH 1997.37.4-10. Поскольку он был относительно большим, Оуэн предположил, что в контексте его предполагаемого морского образа жизни сцелидозавр мог быть яйцеживородящим . [ 14 ] Короткий препубис в этом экземпляре убедил ученых, что этот отросток не представляет собой основное лобковое тело, как думали некоторые, которые не могли поверить, что тонкий, обращенный назад лобок с орнитишиями гомологичен обращенному вперед гораздо более крупному лобку. лобковая кость у большинства групп рептилий. [ нужна ссылка ]
В последнее время в Чармуте были сделаны новые открытия, но не в результате коммерческих карьеров, а усилиями палеонтологов-любителей. В 1968 году был описан второй частичный скелет молодого человека, экземпляр NHMUK PV R6704. [ 24 ] об этом уже сообщалось в 1959 году. [ 25 ] Найденный геологом Джеймсом Фредериком Джексоном (1894–1966) из Чармута, он принадлежит к немного более молодому слою, пачке мергелей Стоунбарроу, датируемой ранним плинсбахом , возрастом около 190 миллионов лет. [ 23 ] В 1985 году Саймон Барнсли, Дэвид Костейн и Питер Лэнгэм раскопали частичный скелет, включая очень полный череп и отпечатки кожи. [ 26 ] который был продан Бристольскому музею , где зарегистрирован как образец BRSMG CE12785. Образец CAMSMX.39256 является частью коллекции Музея Седжвика в Кембридже . [ 23 ] Некоторые экземпляры остались неописанными, поскольку находятся в частных коллекциях. К ним относится скелет длиной 3,1 метра (десять футов), найденный Дэвидом Соулом в 2000 году, возможно, самый полный экземпляр нептичьего динозавра, когда-либо обнаруженный на Британских островах. Теперь известны все элементы скелета. [ 23 ] Находки Соле отличаются от лектотипа деталями доспеха и могут представлять собой отдельный таксон или отражать половой диморфизм . [ 11 ] В 2020 году Норман это опроверг. [ 7 ]

Между 1980 и 2000 годами три окаменелости были обнаружены на пляже недалеко от Гоббинса в Северной Ирландии палеонтологом Роджером Бирном. Точное геологическое происхождение ни одного из образцов не сообщается, но очень темная окраска образцов указывает (путем сравнения с морскими окаменелостями в других местах Северной Ирландии), что они происходят из пород группы Лиас , вероятно, из зоны Планорбис или Зона Pre-planorbis аргиллитовой формации Ватерлоо . В число образцов входят BELUM K3998, фрагмент проксимального отдела бедренной кости, обнаруженный в январе 1980 года; BELUM K12493, фрагмент диафиза большеберцовой кости, обнаруженный в апреле 1981 г.; и BELUM K2015.1.54, небольшой пятиугольный объект, обнаруженный в 2000 году. Гистолог Робин Рид признал первый экземпляр динозавровым из-за его текстуры и структуры костей и сообщил о нем в 1989 году, подозревая, что он принадлежал сцелидозавру или подобному животному. Затем Бирн опознал образец большеберцовой кости как динозавра, используя аналогичные идентификаторы; На основании ассоциации предполагалось, что эти два экземпляра произошли от одного и того же животного. По той же логике пятиугольный образец затем был принят за остеодерму сцелидозавра. [ 27 ]
Эти ирландские экземпляры, наряду с другими, обнаруженными коллекционером окаменелостей Уильямом Греем где-то в конце 19 или начале 20 века, были официально изучены Майклом Дж. Симмсом и его коллегами, и исследование было опубликовано по ним в журнале Proceedings of the Geists' Association в декабре. 2021. Отнесение бедренного фрагмента было подтверждено: он явно принадлежит к птицетазовым особям, а по размеру и морфологии очень похож на Scelidosaurus и в отличие от близкого родственника Scutellosaurus . Однако большеберцовая кость была интерпретирована как принадлежащая неопределенному неотероподу , пятиугольный объект — как простой кусок базальта, напоминающий окаменелость, а образец Грея — как принадлежащий ихтиозавру . Бедренная кость скелидозавра и большая берцовая кость теропода — единственные известные останки динозавров из Ирландии , которая имеет скудную мезозойскую летопись окаменелостей, полностью состоящую из морских местонахождений, а образец сцелидозавра был первым, когда-либо зарегистрированным с острова. [ 27 ] Однако в 2024 году Сатчелл повторно идентифицировал фрагмент проксимального отдела бедренной кости (BELUM K3998) как останки неопределенного динозавра, не принадлежащие сцелидозавру или птицетазовым животным. [ 28 ]
В 2000 году Дэвид Мартилл и др. сообщил о сохранении мягких тканей в образце, указанном ср. Сцелидозавр сп. (то есть материал ориентировочно относился к роду Scelidosaurus , а не к какому-либо конкретному виду). Окаменелость с инвентарным номером BRSMG CF2781, находившаяся в начале 1990-х годов в уже подготовленном состоянии, была обнаружена в наследии покойного профессора Джона Чаллинора , который использовал ее для иллюстрации своих лекций. Его происхождение неизвестно. Он состоит из серии из восьми хвостовых позвонков в вырезанной плите карбонатного аргиллита, возраст которого оценивается от позднего геттанга до синемурийского этапа . Части окаменелости сохранились таким образом, что вокруг позвонков видна оболочка сохранившихся мягких тканей и видно наличие эпидермального слоя над щитками. [ 10 ] Авторы пришли к выводу, что остеодермы всех базальных панцирных динозавров были покрыты жестким, вероятно, ороговевшим слоем кожи. [ 10 ]
Дополнительные виды
[ редактировать ]
Scelidosaurus harrisonii , названный и описанный Оуэном, в настоящее время является единственным признанным видом , основанным на нескольких почти полных скелетах. Потенциальный второй вид из синемурийского возраста формации Нижний Луфэн , Scelidosaurus oehleri , был описан Дэвидом Джеем Симмонсом в 1965 году под собственным родом Tatisaurus . В 1996 году Спенсер Г. Лукас перенес его на сцелидозавра . [ 29 ] Хотя окаменелости фрагментарны, эта переоценка не была принята, и сегодня S. oehleri снова признан Tatisaurus . [ 5 ] [ 30 ]
В 1989 году щитки, обнаруженные в формации Кайента (группа Глен-Каньон) на севере Аризоны , были отнесены Кевином Падианом к Scelidosaurus sp. и использованы для определения возраста этих слоев около 199,6–196,5 миллионов лет назад. в то время еще считалось, что Scelidosaurus harrisonii относится к раннему синемурийскому периоду. [ 31 ] Эти щитки установили географическую связь между каньоном Глен в Аризоне и Европой, где окаменелости сцелидозавра . ранее были обнаружены [ 31 ] Позже учёные отвергли отнесение к сцелидозавру , так как щитки различны по форме. [ 5 ] [ 23 ] назвал новый вид S. arizonenesis . В 2014 году Роман Уланский на основе этих экземпляров [ 32 ] В 2016 году Питер Малкольм Гальтон и Кеннет Карпентер определили его как nomen dubium , вместо этого снова поместив образцы как Thyreophora indet. [ 33 ]
Классификация и филогения
[ редактировать ]

Scelidosaurus был помещен в Dinosauria Оуэном в 1861 году. В 1868/1869 году Эдвард Дринкер Коуп предложил семейство Scelidosauridae в двойной лекции, но она была опубликована только в декабре 1871 года; [ 34 ] поэтому именно Томас Генри Хаксли обоснованно назвал Scelidosauridae в 1869 году. [ 13 ] [ 35 ] В девятнадцатом веке почти любой известный тогда панцирный динозавр считался представителем Scelidosauridae. В конце двадцатого века этот термин использовался для обозначения группы «примитивных» птицетазовых, близких к предкам анкилозавров и стегозавров, таких как Scutellosaurus , Emausaurus , Lusitanosaurus и Tatisaurus . [ 3 ] Сегодня палеонтологи обычно считают Scelidosauridae парафилетическими , не образующими, таким образом, отдельной ветви или клады ; однако Бентон (2004) называет эту группу монофилетической . [ 36 ] Семья была возрождена китайским палеонтологом Дун Чжимином в его описании в 2001 году биенозавра , тиреофора, имеющего близкое родство со сцелидозавром . [ 37 ]
Сцелидозавр был птицетазом . Это был самый старый птицетаз, известный до описания геранозавра в 1911 году. [ 5 ] В течение двадцатого века его в разное время классифицировали как анкилозавра или стегозавра . Альфред фон Зиттель (1902 г.), Уильям Элджин Суинтон (1934 г.) и Роберт Эпплби и др. (1967) определили этот род как стегозавров, [ 38 ] хотя тогда это понятие охватывало все броневые формы. В статье 1968 года Ромер утверждал, что это был анкилозавр. [ 38 ] В 1977 году Ричард Талборн из Университета Квинсленда попытался реклассифицировать сцелидозавра как орнитопода, похожего на тенонтозавра или игуанодона . [ 38 ] Талборн утверждал, что сцелидозавр был легко сложенным двуногим динозавром, приспособленным для бега. Теории Тулборна об этом роде, выдвинутые в 1977 году, с тех пор были отвергнуты.
Эти дебаты все еще продолжаются; в настоящее время сцелидозавр считается либо более близким родственником анкилозавридам, чем стегозавридам, и, как следствие, настоящим анкилозавром, [ 9 ] [ 39 ] или базально по отношению к расколу анкилозавра и стегозавра. [ 5 ] Классификация стегозавров вышла из моды, но ее можно увидеть в старых книгах о динозаврах. [ 40 ] Кладистический анализ всегда обнаруживал базальное положение Scelidosaurus вне Eurypoda . [ 23 ]
Положение сцелидозавра согласно кладистическому исследованию 2011 года показано на этой кладограмме: [ 41 ]
В монографии сцелидозаврах 2022 о Дэвида Нормана года были обнаружены другие взаимоотношения между тиреофорами: стегозаврия является наиболее базовой группой, а сцелидозавр наиболее тесно связан с анкилозаврией. [ 42 ]
| Тирофора |
| ||||||||||||||||||||||||
Ископаемые находки тиреофоров, более базальных, чем Scelidosaurus, редки. Более «примитивный» Scutellosaurus , также найденный в Аризоне, был более ранним родом, который был факультативно двуногим. Во Франции были найдены следы возможного раннего панцирного динозавра, жившего примерно 195 миллионов лет назад. [ 43 ] Предки этих базальных тиреофоров произошли от ранних птицетазовых, подобных лесотозавру, в позднем триасе. [ 5 ]
Палеобиология
[ редактировать ]
Диета
[ редактировать ]Как и большинство других тиреофоров, сцелидозавр известен как травоядный . Однако в то время как некоторые более поздние группы птицетазовых обладали зубами, способными перемалывать растительный материал, у сцелидозавра были меньшие и менее сложные зубы листовидной формы, подходящие для обрезки растительности, а челюсти могли совершать только вертикальные движения из-за короткого челюстного сустава. [ 3 ] Пол Барретт пришел к выводу, что сцелидозавра кормили с помощью системы прокола-раздавливания зубов с точным, но простым движением челюсти вверх и вниз, при котором пища раздавливалась между внутренней стороной верхних зубов и внешней. стороны нижних зубов, при этом зубы фактически не касаются друг друга, о чем свидетельствуют очень длинные вертикальные фасетки износа только на нижних зубах. [ 44 ] В этом аспекте он напоминал стегозаврид , которые также имели примитивные зубы и простые челюсти. [ 45 ] Его диета состояла бы из папоротников или хвойных деревьев , поскольку травы появились только в конце мелового периода , после того как сцелидозавр уже давно вымер.
Еще одно сходство со стегозаврами — узкая голова, что может указывать на избирательный рацион, состоящий из высококачественных кормов. Однако Барретт отметил, что для животного размером со сцелидозавра , с большим кишечником, обеспечивающим эффективную ферментацию, потребление легкоусвояемой пищи с высокой энергетической ценностью было менее важным, чем для более мелких животных, которые часто критически зависят от нее. [ 44 ] Норман пришел к выводу, что сцелидозавр питался низкой кустарниковой растительностью высотой до одного метра. Подъем на задних конечностях мог бы увеличить зону кормления по вертикали и, возможно, был анатомически возможен, но Норман сомневался, что это важная часть его поведения. [ 5 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Лидделл и Скотт (1980). Греко-английский лексикон, сокращенное издание. Издательство Оксфордского университета, Оксфорд, Великобритания. ISBN 0-19-910207-4
- ^ Jump up to: а б Норман, Дэвид (2001). « Сцелидозавр , самый ранний полный динозавр» в книге «Бронированные динозавры» , стр. 3–24. Блумингтон: Издательство Университета Индианы. ISBN 0-253-33964-2 .
- ^ Jump up to: а б с д и Ламберт Д. (1993). Самая лучшая книга о динозаврах . Дорлинг Киндерсли, Нью-Йорк, 110–113. ISBN 1-56458-304-X
- ^ Пол, GS, 2010, Принстонский полевой справочник по динозаврам , Princeton University Press, стр. 217
- ^ Jump up to: а б с д и ж г час я дж к л м н Норман, Д.Б., Уитмер, Л.М. и Вейшампель, Д.Б. (2004). «Базальная тиреофора». В Вейшампеле Д.Б., Додсоне П. и Осмолской Х. (ред.). Динозаврия, 2-е издание . Калифорнийский университет прессы. стр. 335–342. ISBN 978-0-520-24209-8 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Герлинский, Грегард (1999). «Следы крупного тиреофора из ранней юры Польши» (PDF) . Acta Palaeontologica Polonica . 44 . Проверено 27 июля 2016 г.
- ^ Jump up to: а б с д и Норман, Дэвид Б. (2020). «Scelidosaurus harrisonii из ранней юры Дорсета, Англия: анатомия черепа». Зоологический журнал Линнеевского общества . 188 (1): 1–81. doi : 10.1093/zoolinnean/zlz074 .
- ^ Мейдмент, SCR, Порро, Л.Б., 2010, «Гомология глазных яблок и происхождение супраорбитальных оссификаций у птицетазовых динозавров», Lethaia , 43 : 95-111.
- ^ Jump up to: а б Казлев, М. Алан (2007). «Ornithischia: Ankylosauromorpha». Архивировано 2 марта 2007 года в Wayback Machine Palaeos. Проверено 11 февраля 2007 г.
- ^ Jump up to: а б с д Мартилл, Д.М., Баттен, DJ, и Лойделл, Д.К. (2000). Новый экземпляр динозавра-тиреофора ср. Сцелидозавр с сохранением мягких тканей. Палеонтология , Том. 43, часть 3, 2000, стр. 549–559. два : 10.1111/j.0031-0239.2000.00139.x
- ^ Jump up to: а б Нэйш, Д. и Мартилл, Д.М., 2007, «Динозавры Великобритании и роль Лондонского геологического общества в их открытии: базальные динозаврии и заурисхии», Журнал Геологического общества, Лондон , 164 : 493–510.
- ^ Оуэн, Р., 1859, «Палеонтология», В: Британская энциклопедия, издание 8, том 17, с. 150
- ^ Jump up to: а б с д Чариг, А.Дж. и Ньюман, Б.Х.†, 1992, « Scelidosaurus harrisonii Owen, 1861 (Reptilia, Ornithischia): предлагаемая замена в неподходящем лектотипе», Бюллетень зоологической номенклатуры , 49 : 280–283
- ^ Jump up to: а б с д Р. Оуэн, 1861, Монография об ископаемых динозаврах (Scelidosaurus harrisonii, Owen) из Нижнего Лиаса, часть I. Монографии о британских ископаемых рептилиях из оолитовых формаций 1 стр. 14
- ^ Джордж К. Стейскал, 1970, «О грамматике имен, образованных с помощью -scelus, -sceles, -scelis и т. д.», Proceedings of the Biological Society of Washington 84 (2): 7-12
- ^ Р. Оуэн, 1863, Монография ископаемых рептилий лиасовых формаций. Часть 2. Монография об ископаемых динозаврах ( Scelidosaurus harrisonii Owen) Нижнего Лиаса . Монографии Палеонтографического общества. Часть 2 . стр. 1-26
- ^ Норман, Д.Б., 2000, «Профессор Ричард Оуэн и важный, но забытый динозавр Scelidosaurus harrisonii », «Историческая биология» , 14 : 235–253.
- ^ Jump up to: а б Лидеккер Р., 1888 г., Каталог ископаемых рептилий и амфибий в Британском музее. Часть 1. Включающая отряды Ornithosauria, Crocodilia, Dinosauria, Squamata, Rhynchocephalia и Proterosauria. Британский музей (естественная история)
- ^ Ньюман, Б.Х. (1968) Юрский динозавр Scelidosaurus harrisoni , Оуэн. Палеонтология 11 (1), 40-3.
- ^ Международная комиссия по зоологической номенклатуре, 1994, « Scelidosaurus harrisonii Owen, 1861 (Reptilia, Ornithischia): заменен лектотип», Бюллетень зоологической номенклатуры 51 : 288
- ^ Пикеринг, С., 1995, Парк Юрского периода: несанкционированные еврейские фракталы в филопатрии. Проект «Фрактальное масштабирование в динозаврологии» , 2-е исправленное издание. Капитола, Калифорния. 478 стр.
- ^ Jump up to: а б Бенсон, Р., 2010, «Остеология Magnosaurus nethercombensis (Dinosauria, Theropoda) из байоса (средняя юра) Соединенного Королевства и повторное исследование древнейших записей о столбняках», Журнал систематической палеонтологии 8 (1) : 131-146
- ^ Jump up to: а б с д и ж г Барретт, П. М. и Мейдмент, SCR, 2011, «Динозавры Дорсета: Часть III, птицетазовые динозавры (Dinosauria, Ornithischia) с дополнительными комментариями о зауроподах», Труды Дорсетского естественной истории и археологического общества 132 : 145–163
- ^ Риксон, А.Е., 1968, «Развитие останков небольшого сцелидозавра из узелка Лиаса», Museums Journal , 67 : 315–321.
- ^ Делар, Дж. Б., 1959, «Мезозойские рептилии Дорсета: Часть вторая», Труды Дорсетского общества естественной истории и археологии, за 1958 год 80 : 52-90
- ^ Энсом, ПК, 1989, «Новые останки скелидозавра из Нижнего Лиаса Дорсета», Труды Дорсетского естественной истории и археологического общества 110 : 165 и 167
- ^ Jump up to: а б Симмс, Майкл Дж.; Смит, Роберт Ш.; Мартиалл, Дэвид М.; Коллинз, Патрик С.; Бирн, Роджер (2021). «Первый динозавр остался в Ирландии» . Труды Общества геологов . 132 (6): 771–779. Бибкод : 2021ПрГА..132..771С . дои : 10.1016/j.pgeola.2020.06.005 . S2CID 228811170 .
- ^ Сатчелл, КГ (2024 г.). «Переоценка остатков сцелидозавра из Ирландии и важность идентификации на основе апоморфии». Труды Общества геологов . 135 (3): 349–351. дои : 10.1016/j.pgeola.2024.03.002 .
- ^ Лукас С.Г. (1996). Тиреофорный динозавр Scelidosaurus из нижнеюрской формации Луфэн, Юньнань, Китай. стр. 81-85, в Моралесе, М. (ред.), Континентальная юра . Бюллетень 60 Музея Северной Аризоны.
- ^ Норман, Дэвид Б.; Батлер, Ричард Дж .; Мейдмент, Сюзанна CR (2007). «Пересмотр статуса и сходства птицетазовых динозавров Tatisaurus oehleri Simmons, 1965» (PDF) . Зоологический журнал Линнеевского общества . 150 (4): 865–874. дои : 10.1111/j.1096-3642.2007.00301.x . Архивировано из оригинала (PDF) 27 октября 2021 года . Проверено 20 сентября 2019 г.
- ^ Jump up to: а б Падиан, К. (1989). «Присутствие динозавра Scelidosaurus указывает на юрский возраст формации Кайента (группа Глен-Каньон, северная Аризона)». Геология . май 1989 г., т. 17; нет. 5; п. 438-441
- ^ Уланский, Р.Э., 2014. Эволюция стегозавров (Dinosauria; Ornithischia). Динология, 35 с. [СКАЧАТЬ PDF] http://dinoweb.narod.ru/Ulansky_2014_Stegosaurs_evolution.pdf .
- ^ Гальтон, Питер М. и Карпентер, Кеннет, 2016, «Пластинчатый динозавр Stegosaurus longispinus Gilmore, 1914 (Dinosauria: Ornithischia; верхняя юра, запад США), типовой вид Alcovasaurus n. gen.», Новый ежегодник геологии и палеонтологии - Трактаты 279 (2): 185-208
- ^ Э.Д. Коуп, 1871, Краткий обзор вымерших батрахий, рептилий и авесов Северной Америки. Труды Американского философского общества, новая серия 14 , стр. 252.
- ^ Хаксли, TH 1869. «О динозаврах триаса, с наблюдениями по классификации динозавров», Nature , 1 : 146.
- ^ Бентон, MJ (2004). Палеонтология позвоночных (Третье изд.). Издательство Блэквелл. ISBN 978-0-632-05637-8
- ^ Дун Чжимин (2001). «Примитивный панцирный динозавр из бассейна Луфэн, Китай» . В Танке, Даррен Х.; Карпентер, Кеннет (ред.). Жизнь мезозойских позвоночных . Издательство Университета Индианы. стр. 237–243 . ISBN 0-253-33907-3 .
- ^ Jump up to: а б с Тулборн, Р.А. (1977)Взаимоотношения динозавров нижней юры Scelidosaurus harrisonii . Журнал палеонтологии. июль 1977 г.; т. 51; нет. 4; п. 725-739
- ^ Карпентер, Кеннет (2001). «Филогенетический анализ анкилозаврии». В Карпентер, Кеннет (ред.). Бронированные динозавры . Издательство Университета Индианы. стр. 455–480. ISBN 978-0-253-33964-5 .
- ^ Кольбер, Эдвин Х. (1965). Эпоха рептилий . Нью-Йорк: WW Norton & Company, Inc., с. 128. ИСБН 978-0-486-29377-6 .
- ^ Ричард С. Томпсон, Джолион К. Пэриш, Сюзанна С.Р. Мейдмент и Пол М. Барретт, 2011, «Филогения анкилозавровых динозавров (Ornithischia: Thyreophora)», Журнал систематической палеонтологии 10 (2): 301–312
- ^ Норман, Дэвид Б. (1 января 2021 г.). «Scelidosaurus harrisonii (Dinosauria: Ornithischia) из ранней юры Дорсета, Англия: биология и филогенетические связи» . Зоологический журнал Линнеевского общества . 191 (1): 1–86. doi : 10.1093/zoolinnean/zlaa061 . ISSN 0024-4082 .
- ^ Ле Лефф, Дж., Локли, М., Мейер, К., и Пети, Ж.-П. (1999). Открытие тиреофорных путей в геттанге центральной Франции. ЧР акад. наук. Париж 2 328, 215-219
- ^ Jump up to: а б Барретт, премьер-министр (2001). «Износ зубов и возможное движение челюстей Scelidosaurus harrisoni Owen и обзор механизмов питания у других тиреофорных динозавров». В Карпентер, Кеннет (ред.). Бронированные динозавры . Издательство Университета Индианы. стр. 25–52. ISBN 978-0-253-33964-5 .
- ^ Гальтон ПМ; Апчерч П. (2004). «Стегозаврия». В базе данных Вейшампель; Додсон П; Осмольска Х (ред.). Динозаврия (2-е изд.). Издательство Калифорнийского университета. п. 361. ИСБН 978-0-520-24209-8 .
Внешние ссылки
[ редактировать ]- Анкилозавроморфа на Древе Жизни
- Сцелидозавриды