Анкилозавр
| Анкилозавр Временной диапазон: поздний мел ( маастрихт ), | |
|---|---|
 | |
| Слепок черепа анкилозавра (AMNH 5214) спереди, Музей Скалистых гор. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Тиреофора |
| Клэйд : | † Анкилозаврия |
| Семья: | † Анкилозавриды |
| Подсемейство: | † Анкилозаврины |
| Племя: | † Анкилозаврини |
| Род: | † Анкилозавр Браун , 1908 год. |
| Разновидность: | † А. магнитентрис |
| Биномиальное имя | |
| † Анкилозавр магнивентрис Браун, 1908 год. | |
Анкилозавр [номер 1] — род панцирных динозавров . Его окаменелости были найдены в геологических образованиях, относящихся к самому концу мелового периода , около 68–66 миллионов лет назад, на западе Северной Америки, что делает его одним из последних нептичьих динозавров . Он был назван Барнумом Брауном в 1908 году; он монотипичен и содержит только A. Magniventris . Родовое название означает «сросшаяся» или «согнутая ящерица», а видовое название означает «большой живот». На сегодняшний день раскопано несколько экземпляров, но полный скелет так и не был обнаружен. Хотя другие представители Ankylosauria представлены более обширным ископаемым материалом, Ankylosaurus часто считают архетипическим членом своей группы, несмотря на некоторые необычные особенности.
Вероятно, самый крупный из известных анкилозавридов , Анкилозавр , по оценкам, имел длину от 6 до 8 метров (20 и 26 футов) и весил от 4,8 до 8 метрических тонн (от 5,3 до 8,8 коротких тонн). Оно было четвероногим , с широким, крепким телом. У него был широкий и низкий череп с двумя рогами, направленными назад от затылка, и двумя рогами под ними, направленными назад и вниз. В отличие от других анкилозавров, его ноздри были обращены вбок, а не вперед. Передняя часть челюстей была покрыта клювом, а дальше за ним располагались ряды маленьких листовидных зубов. Он был покрыт панцирными пластинами, или остеодермами, с костяными полукольцами, закрывающими шею, и имел большую булаву на конце хвоста. Кости черепа и других частей тела срастались, увеличивая их прочность, и эта особенность послужила источником названия рода.
Анкилозавр — член семейства Ankylosauridae, а его ближайшими родственниками являются Anodontosaurus и Euoplocephalus . Считается, что анкилозавр был медлительным животным, способным при необходимости совершать быстрые движения. Его широкая морда указывает на то, что это был неселективный браузер . Пазухи и носовые камеры на морде, возможно, предназначались для поддержания теплового и водного баланса или могли играть роль в вокализации. Считается, что хвостовая булава использовалась для защиты от хищников или во внутривидовых боях . Экземпляры анкилозавра были обнаружены в формациях Хелл-Крик , Лэнс , Сколлард , Френчмен и Феррис , но, по-видимому, он был редкостью в своей среде обитания. Хотя он жил рядом с анкилозавром- нодозавром , их ареалы и экологические ниши , по-видимому, не пересекались, и анкилозавр , возможно, населял горные районы. Анкилозавр также жил рядом с такими динозаврами, как тираннозавр , трицератопс и эдмонтозавр .
История открытия
[ редактировать ]
В 1906 году экспедиция Американского музея естественной истории под руководством американского палеонтолога Барнума Брауна обнаружила типовой экземпляр Ankylosaurus magniventris (AMNH 5895) в формации Хелл-Крик , недалеко от Гилберт-Крик, штат Монтана . Экземпляр (найденный коллекционером Питером Кайсеном) состоял из верхней части черепа, двух зубов, части плечевого пояса, шейных, спинных и хвостовых позвонков, ребер и более тридцати остеодерм (броневых пластин). Браун научно описал это животное в 1908 году; Общее название происходит от греческих слов αγκυλος ankulos («изогнутый» или «кривой»), относящихся к медицинскому термину анкилоз , жесткости, возникающей в результате слияния костей черепа и тела, и σαυρος sauros («ящерица»). . Название можно перевести как «сросшаяся ящерица», «жесткая ящерица» или «изогнутая ящерица». Название вида , Magniventris, происходит от Magnus латинского ( «большой») и латинского типового venter («живот»), что указывает на большую ширину тела животного. [2] [3] [4]

Реконструкция скелета, сопровождавшая описание 1908 года, восстановила недостающие части аналогично стегозавру , и Браун сравнил результат с вымершим панцирным млекопитающим глиптодоном . [2] В отличие от современных изображений, реконструкция Брауна, похожая на стегозавра, показала крепкие передние конечности, сильно изогнутую спину, таз с зубцами, выступающими вперед из подвздошной и лобковой кости, а также короткий свисающий хвост без хвостовой булавы , который был неизвестен в то время. время. Браун также реконструировал броневые пластины параллельными рядами, идущими по спине; эта договоренность была чисто гипотетической. Реконструкция Брауна стала очень влиятельной, а реставрации животного на основе его диаграммы были опубликованы только в 1980-х годах. [5] [6] [7] Брауна в 1908 году В обзоре описания Анкилозавра американский палеонтолог Сэмюэл Венделл Уиллистон раскритиковал реконструкцию скелета как основанную на слишком небольшом количестве останков и заявил, что Анкилозавр был просто синонимом рода Stegopelta , который Уиллистон назвал в 1905 году. Уиллистон также заявил, что что реконструкция скелета родственного полакантуса, проведенная венгерским палеонтологом Францем Нопчей, была лучшим примером того, как анкилозавры могли появиться в жизни. [8] Заявление о синонимии не было принято другими исследователями, и теперь эти два рода считаются разными. [9]
Браун собрал 77 остеодерм во время раскопок экземпляра тираннозавра в формации Ланс в Вайоминге в 1900 году. Он упомянул эти остеодермы (образец AMNH 5866) в своем описании анкилозавра, но думал, что они вместо этого принадлежали тираннозаврам . Палеонтолог Генри Фэйрфилд Осборн также выразил эту точку зрения, когда в 1905 году он описал образец тираннозавра как ныне синонимичный род Dynamosaurus. Более поздние исследования показали, что они похожи на экземпляры анкилозавра ; похоже, что Браун сравнил их с некоторыми остеодермами Euoplocephalus , которые были ошибочно занесены в каталог как принадлежащие Ankylosaurus . AMNH [10] [11]

В 1910 году другая экспедиция AMNH под руководством Брауна обнаружила экземпляр анкилозавра (AMNH 5214) в формации Сколлард на берегу реки Ред-Дир в Альберте, Канада. Этот образец включал в себя полный череп, нижние челюсти, первую и единственную хвостовую булаву, известную для этого рода, а также ребра, позвонки, кости конечностей и доспехи. В 1947 году американские коллекционеры окаменелостей Чарльз М. Штернберг и Т. Поттер Чамни собрали череп и нижнюю челюсть (образец CMN 8880, ранее NMC 8880). В 1 километре ( 5 ⁄ 8 миль) к северу от места, где был найден экземпляр 1910 года. Это самый крупный из известных черепов анкилозавра , но он местами поврежден. Часть хвостовых позвонков (образец CCM V03) была обнаружена в 1960-х годах в русле реки Паудер , штат Монтана, части формации Хелл-Крик. Помимо этих пяти неполных экземпляров, было обнаружено множество других изолированных остеодерм и зубов. [12] [10]
В 1990 году американский палеонтолог Уолтер П. Кумбс отметил, что зубы двух черепов, отнесенных к A. magniventris, в некоторых деталях отличались от зубов голотипа, и хотя он выразил «внимательное искушение» назвать новый вид Ankylosaurus для от этого он воздержался, поскольку диапазон изменчивости вида не был полностью задокументирован. Он также предположил, что два зуба, связанные с экземпляром голотипа, возможно, ему не принадлежали, поскольку они были обнаружены в матрице внутри носовых камер. [13] Американский палеонтолог Кеннет Карпентер в 2004 году признал зубы принадлежащими A. magniventris и что все экземпляры принадлежали к одному и тому же виду, отметив, что зубы других анкилозавров сильно различаются. [10]
Большинство известных экземпляров анкилозавров не были подробно описаны с научной точки зрения, хотя некоторые палеонтологи планировали сделать это, пока Карпентер не переописал этот род в 2004 году. [10] В 2017 году канадские палеонтологи Виктория М. Арбор и Джордан Мэллон переописали род в свете новых открытий анкилозавров, включая элементы голотипа, которые ранее не упоминались в литературе (например, части черепа и шейные полукольца). . Они пришли к выводу, что, хотя анкилозавр является самым известным представителем своей группы, он был причудливым по сравнению с родственными анкилозаврами и, следовательно, не был репрезентативным представителем группы. Несмотря на свою известность, он известен по гораздо меньшему количеству останков, чем его ближайшие родственники. [12]
Описание
[ редактировать ]
Анкилозавр был крупнейшим известным динозавром -анкилозавром и, возможно, самым крупным анкилозавридом. [12] В 2004 году Карпентер подсчитал, что человек с самым большим из известных черепов (образец CMN 8880), который 64.5 centimeters (2 ft 1+1⁄2 in) long and 74.5 cm (2 ft 5+1⁄4 in) wide, was about 6.25 m (20 ft 6 in) long and had a hip height of about 1.7 m (5 ft 7 in). The smallest-known skull (specimen AMNH 5214) is 55.5 cm (1 ft 9+3⁄4 in) long and Его ширина составляла 64,5 см (2 фута 1 + 1 ⁄ 2 дюйма), и, по оценкам Карпентера, его длина составляла около 5,4 м (17 футов 9 дюймов) и высота в бедрах около 1,4 м (4 фута 7 дюймов). [10] Английский палеонтолог Роджер Б. Дж. Бенсон и его коллеги оценили вес AMNH 5214 в 4,78 метрических тонны (5,27 коротких тонны) в 2014 году. [14]
В 2017 году на основе сравнений с более полными анкилозавринами Арбор и Мэллон оценили длину От 7,56 до 9,99 м (от 24 футов 9 + 1 ⁄ 2 дюйма до 32 футов 9 + 1 ⁄ дюйма ) для CMN 8880 и от 6,02 до 7,95 м (от 19 футов 9 дюймов до 26 футов 1 дюйм) для AMNH 5214. Хотя последнее — самый маленький экземпляр анкилозавра , его череп все же крупнее, чем у любых других анкилозавров. Несколько других анкилозавров достигали около 6 м (20 футов) в длину. Поскольку позвонки AMNH 5214 ненамного больше, чем у других анкилозавров, Арбор и Мэллон сочли свою оценку верхнего диапазона почти 10 метров (33 фута) для большого анкилозавра слишком длинной и вместо этого предложили длину 8 м (26 футов). . Арбор и Мэллон оценили вес AMNH 5214 в 4,78 т (5,27 коротких тонны) и предварительно оценили вес CMN 8880 в 7,95 т (8,76 коротких тонн). [12]
Череп
[ редактировать ]
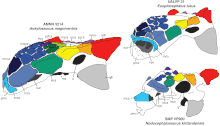
Три известных черепа анкилозавра различаются различными деталями; Считается, что это результат тафономии (изменений, происходящих во время распада и окаменения останков) и индивидуальных вариаций . Череп был низким и треугольной формы, ширина его превышала длину; задняя часть черепа была широкой и низкой. Череп имел широкий клюв на предчелюстных костях . Орбиты (глазницы) были от почти круглых до слегка овальных и не были обращены прямо вбок, поскольку череп суживался вперед. Черепная коробка была короткой и прочной, как и у других анкилозавров. Гребни над глазницами сливались в верхние чешуйчатые рога (их форма описана как « пирамидальная »), направленные назад в стороны от задней части черепа. Гребень и рог, вероятно, изначально были отдельными элементами, как это видно у родственных пинакозавров и эвоплоцефалов . Ниже верхних рогов скуловые имелись рога, направленные назад и вниз. Рога, возможно, изначально представляли собой остеодермы, сросшиеся с черепом. Чешуйчатые черепные орнаменты на поверхности черепов анкилозавров называются « caputegulae » и являются результатом ремоделирования самого черепа. Это привело к стиранию швов между элементами черепа, что характерно для взрослых анкилозавров. Рисунок caputegulum черепа у разных экземпляров был разным, хотя некоторые детали являются общими. Головки названы в соответствии с их положением на черепе, а у анкилозавра они включают относительно большую шестиугольную (или ромбовидную) носовую головку в передней части морды между ноздрями, которая имела лореальные головки с каждой стороны, передняя и задняя надглазничные головки над каждой глазницей и гребень затылочных головок в задней части черепа. [10] [12] [15]

Морда анкилозавра была уникальной среди анкилозавров и претерпела «экстремальную» трансформацию по сравнению с его родственниками. Морда была изогнутой и усеченной спереди, а ноздри были эллиптической формы и были направлены вниз и наружу, в отличие от всех других известных анкилозаврид, у которых они были обращены наклонно вперед или вверх. Кроме того, ноздри не были видны спереди, потому что пазухи были расширены по бокам предчелюстных костей в большей степени, чем у других анкилозавров. Большие лореальные головки - ремни, боковые остеодермы морды - полностью закрывали увеличенное отверстие ноздрей, придавая луковичный вид. Ноздри также имели внутринариальную перегородку , которая отделяла носовой ход от пазухи. На каждой стороне морды было по пять пазух, четыре из которых переходили в верхнечелюстную кость. Носовые полости (или камеры) анкилозавра были удлинены и разделены перегородкой по средней линии, которая разделяла внутреннюю часть морды на две зеркальные половины. Носовые камеры имели два отверстия, в том числе хоаны (внутренние ноздри), а воздухоносный проход имел петлеобразную форму. [10] [12] Верхнечелюстные кости расширялись в стороны, создавая впечатление выпуклости, что могло быть связано с расположенными внутри пазухами. На верхних челюстях был гребень, который, возможно, служил местом прикрепления мясистых щек; Наличие щек у птицетазовых является спорным, но у некоторых нодозавров были броневые пластины, закрывающие область щек, которые, возможно, были встроены в плоть. [10]
Образец AMNH 5214 имеет 34–35 зубных альвеол (зубных лунок) на верхней челюсти. Зубные ряды на верхних челюстях этого экземпляра имеют длину около 20 сантиметров (7,9 дюйма). У каждой альвеолы было отверстие сбоку замещающий зуб (отверстие), где можно было увидеть . По сравнению с другими анкилозаврами нижняя челюсть анкилозавра была низкой по сравнению с ее длиной, а при взгляде сбоку зубной ряд был почти прямым, а не изогнутым. Нижние челюсти полностью сохранились только у самого маленького экземпляра (AMNH 5214) и имеют длину около 41 сантиметра (16 дюймов). Такую же длину имеет неполная нижняя челюсть самого крупного экземпляра (CMN 8880). AMNH 5214 имеет 35 зубных альвеол в левой зубной кости () и 36 в правой, всего 71. Предзубная кость кончика нижней челюсти пока не обнаружена. [10] Как и у других анкилозавров, у анкилозавра были маленькие пластинчатые (листовидные) зубы, сжатые вбок. [13] Зубы в основном были выше ширины и очень маленькими; их размер по отношению к черепу означал, что челюсти анкилозавра могли вместить больше зубов, чем у других анкилозавров. Зубы самого большого черепа анкилозавра в абсолютном смысле меньше, чем у самого маленького черепа. Некоторые зубы сзади в зубном ряду изогнуты назад, а коронки зубов обычно с одной стороны более плоские, чем с другой. [10] анкилозавра Зубы имеют диагностическое значение и их можно отличить от зубов других анкилозаврид по их гладким сторонам. Зубчики . крупные, их количество от шести до восьми на передней части зуба и от пяти до семи сзади [10] [16]
Посткраниальный скелет
[ редактировать ]
Структура большей части скелета анкилозавра , включая большую часть таза , хвоста и ступней, до сих пор неизвестна. [10] Оно было четвероногим , и его задние конечности были длиннее передних. [17] У экземпляра-голотипа длина лопатки (лопатки) составляет 61.5 cm (2 ft 1⁄4 in) long and was fused with the coracoid (a rectangular bone connected to the lower end of the scapula). It also had entheses (connective tissue) for various muscle attachments. The humerus (upper arm bone) of AMNH 5214 was short, very broad and about 54 cm (1 ft 9+1⁄2 in) long. The femur (thigh bone), also from AMNH 5214, was 67 см (2 фута 2 + 1 ⁄ дюйма ) в длину и очень прочный. Хотя ступни анкилозавра известны не полностью, на задних ступнях, вероятно, было три пальца, как и у продвинутых анкилозаврид. [10]
Шейные позвонки имели широкие нервные отростки, высота которых увеличивалась по направлению к телу. Передняя часть нервных отростков имела хорошо развитые энтезы, что было распространено среди взрослых динозавров и указывает на наличие крупных связок , которые помогали поддерживать массивную голову. Спинные позвонки имели центры (или тела ), короткие по сравнению с их шириной, а их нервные отростки были короткими и узкими. Спинные позвонки были плотно расставлены, что ограничивало движение спины вниз. Нервные отростки имели окостеневшие (превратившиеся в кости) сухожилия , которые также перекрывали некоторые позвонки. Ребра последних четырех задних позвонков были слиты с диапофизами и парапофизами (структурами, соединяющими ребра с позвонками), и грудная клетка в этой части тела была очень широкой. Центры хвостовых позвонков были слегка амфицелозными, то есть вогнутыми с обеих сторон. [10]
Броня
[ редактировать ]
Характерной особенностью анкилозавра был его панцирь, состоящий из бугорков и костных пластин, известных как остеодермы или щитки, встроенных в кожу. Они не были обнаружены в сочленениях, поэтому их точное расположение на теле неизвестно, хотя выводы можно сделать на основе родственных животных, и были предложены различные конфигурации. Остеодермы варьировались от От 1 сантиметра ( 1 ⁄ дюйма ) в диаметре до 35,5 см (14 дюймов) в длину и различной формы. Остеодермы анкилозавра обычно были тонкостенными и полыми на нижней стороне. По сравнению с Euoplocephalus остеодермы анкилозавра были более гладкими. Пространство между более крупными, вероятно, занимало множество более мелких остеодерм и косточек, как и у других анкилозаврид. Остеодермы, покрывающие тело, были очень плоскими, хотя и с низким килем по одному краю. Напротив, у нодозаврид Edmontonia были высокие кили, простирающиеся от одного края до другого по средней линии остеодерм. У анкилозавра были остеодермы меньшего размера с килем по средней линии. [12] [10]


Как и у других анкилозаврид, у анкилозавра были шейные полукольца (броневые пластины на шее), но они известны только по фрагментам, что делает их точное расположение неопределенным. Карпентер предположил, что, если смотреть сверху, пластинки должны были быть парными, образуя перевернутую V-образную форму на шее, а промежуток срединной линии, вероятно, был заполнен небольшими косточками (круглыми костными щитками), обеспечивающими движение. Он полагал, что ширина этого бронепояса была слишком велика, чтобы его можно было разместить только на шее, и что он закрывал основание шеи и переходил на область плеч. Арбор и канадский палеонтолог Филип Дж. Карри не согласились с интерпретацией Карпентера в 2015 году и указали, что фрагменты шейного полукольца образца голотипа не совпадали друг с другом так, как предлагал Карпентер (хотя это могло быть связано с поломкой). Вместо этого они предположили, что фрагменты представляют собой остатки двух шейных полуколец, которые образовывали две полукруглые пластины доспеха вокруг верхней части шеи, как у близкородственных Анодонтозавр и Эуоплоцефал . [10] [15] Арбор и Мэллон развили эту идею, описав форму этих полуколец как «непрерывные U-образные хомуты» над верхней частью шеи, и предположили, что у Анкилозавра было шесть килевидных остеодерм с овальными основаниями на каждом полукольце. [12]
Первые остеодермы за вторым шейным полукольцом по форме были бы аналогичны остеодермам первого полукольца, а остеодермы на спине, вероятно, уменьшались в диаметре кзади. Самые крупные остеодермы, вероятно, располагались поперечными и продольными рядами на большей части тела, причем четыре или пять поперечных рядов были разделены складками кожи. Остеодермы на боках, вероятно, имели более квадратный контур, чем на спине. На боках могло быть четыре продольных ряда остеодерм. В отличие от некоторых базальных анкилозавров и многих нодозавров, анкилозавриды, по-видимому, не имели окостеневших тазовых щитков над бедрами. Некоторые остеодермы без килей, возможно, располагались над тазобедренной областью анкилозавра , как у Euoplocephalus . Анкилозавр мог иметь три или четыре поперечных ряда круглых остеодерм в области таза, которые были меньше, чем на остальной части тела, как у сколозавра . Меньшие треугольные остеодермы могли присутствовать по бокам таза. Уплощенные заостренные пластинки напоминают пластинки по бокам хвоста Saichania и, возможно, были распространены аналогичным образом на Ankylosaurus . Остеодермы с овальными килями могли располагаться на верхней стороне хвоста или на боковых сторонах конечностей. Сжатые треугольные остеодермы, обнаруженные у экземпляров анкилозавра , возможно, располагались по бокам таза или хвоста. Яйцевидные, килеватые и каплевидные остеодермы известны у анкилозавра и, возможно, располагались на передних конечностях, как и те, что известны у пинакозавра , но неизвестно, имели ли задние конечности остеодермы. [10] [12]

Хвостовая булава (или хвостовая шишка) анкилозавра состояла из двух крупных остеодерм с рядом мелких остеодерм по средней линии и двух маленьких остеодерм на кончике; эти остеодермы закрывали последний хвостовой позвонок. Поскольку известна только хвостовая булава экземпляра AMNH 5214, диапазон различий между особями неизвестен. Хвостовая булава AMNH 5214 60 cm (23+1⁄2 in) long, 49 cm (19+1⁄2 in) wide, and 19 cm (7+1⁄2 in) tall. The club of the largest specimen may have been 57 см ( 22 + 1/2 ) дюйма Ширина . Хвостовая булава анкилозавра была полукруглой, если смотреть сверху, подобно булаве Euoplocephalus и Scolosaurus , но в отличие от заостренной булавы остеодермы Anodontosaurus или узкой, удлиненной булавы Dyoplosaurus . Последние семь хвостовых позвонков образовывали «ручку» хвостовой булавы. Эти позвонки соприкасались, между ними не было хрящей, а иногда были ко-окоссифицированы, что делало их неподвижными. Окостеневшие сухожилия прикреплялись к позвонкам перед хвостовой булавой, и эти особенности в совокупности способствовали ее укреплению. Сцепленные зигапофизы (суставные отростки) и нервные отростки рукояточных позвонков имели U-образную форму, если смотреть сверху, тогда как у большинства других анкилозаврид они имеют V-образную форму, что может быть связано с тем, что рукоятка анкилозавра была шире. Большая ширина может указывать на то, что хвост анкилозавра был короче по отношению к длине его тела, чем у других анкилозаврид, или что он имел те же пропорции, но с меньшей булавой. [12] [10] [18]
Классификация
[ редактировать ]Браун считал Ankylosaurus настолько особенным, что сделал его типовым родом нового семейства Ankylosauridae, для которого характерны массивные треугольные черепа, короткие шеи, жесткая спина, широкое тело и остеодермы. Он также классифицировал Palaeoscincus (известный только по зубам) и Euoplocephalus (тогда известный только по частичному черепу и остеодермам) как часть семейства. Из-за фрагментарного состояния останков Брауну не удалось полностью различить Euoplocephalus и Ankylosaurus . Имея для сравнения лишь несколько неполных членов семейства, он полагал, что эта группа была частью подотряда Stegosauria . [2] В 1923 году Осборн придумал название Ankylosauria , тем самым поместив анкилозаврид в отдельный подотряд. [19]


Ankylosauria и Stegosauria теперь сгруппированы вместе в кладе Thyreophora . Эта группа впервые появилась в синемурийскую эпоху и просуществовала 135 миллионов лет, пока не исчезла в маастрихте . Они были широко распространены и обитали в самых разных средах. [10] [6] По мере открытия более полных экземпляров и новых родов теории о взаимосвязи анкилозавров стали более сложными, и гипотезы часто менялись между исследованиями. Помимо Ankylosauridae, Ankylosauria подразделялась на семейства Nodosauridae, а иногда и на Polacanthidae (в этих семействах отсутствовали хвостовые булавы). Анкилозавр считается частью подсемейства Ankylosaurinae (представителей которого называют анкилозаврами) внутри Ankylosauridae. [20] Анкилозавр , по-видимому, наиболее близок к анодонтозаврам и эвоплоцефалам . [21] Следующая кладограмма основана на филогенетическом анализе Ankylosaurinae, проведенном Арбором и Карри в 2015 году: [15]
Поскольку анкилозавр и другие позднемеловые североамериканские анкилозавриды были сгруппированы с азиатскими родами (в трибе, которую авторы назвали анкилозаврини ), Арбор и Карри предположили, что более ранние североамериканские анкилозавриды вымерли к позднему альбу или сеноманскому периоду среднего мела . После этого анкилозавриды повторно колонизировали Северную Америку из Азии в кампанский или туронский период позднего мела и там снова диверсифицировались , что привело к появлению таких родов, как Ankylosaurus , Anodontosaurus и Euoplocephalus . Теория объясняет 30-миллионный разрыв в летописи окаменелостей североамериканских анкилозаврид между возрастами. [15]
Палеобиология
[ редактировать ]Кормление
[ редактировать ]
Как и другие птицетазовые животные, анкилозавр был травоядным . Его широкая морда была приспособлена для неселективной с низкими проростками , подрезки [10] хотя и не в такой степени, как это наблюдается у некоторых родственных родов, особенно Euoplocephalus . [12] [22] Хотя анкилозавры, возможно, не питались волокнистыми и древесными растениями , у них могла быть разнообразная диета, включающая жесткие листья и мясистые плоды. [23] Анкилозавр, вероятно, питался обильными папоротниками и низкорослыми кустарниками . Если предположить, что он был эндотермическим , Анкилозавр съедал бы 60 килограммов (130 фунтов) папоротников в день, что соответствует количеству сухой растительности, которую съел бы большой слон . Потребности в питании могли бы быть более эффективно удовлетворены, если бы Анкилозавр ел фрукты, к которым его маленькие, похожие на бугорки зубы и форма клюва кажутся хорошо приспособленными, по сравнению, например, с Euoplocephalus . Некоторые беспозвоночные, маленькие зубы которых могли быть приспособлены для обращения с ними, также могли обеспечивать дополнительное питание. [12]
Окаменелости зубов анкилозавров имеют износ на лицевой стороне коронки, а не на кончике коронки, как у анкилозавров-нодозавров. [10] В 1982 году Карпентер приписал детенышу анкилозавра два очень маленьких зуба, происходящих из формаций Ланс и Хелл-Крик и измеряющих размер. От 3,2 до 3,3 мм ( от 1 ⁄ 8 до 17 ⁄ 128 дюймов ) в длину соответственно. Меньший зуб сильно изношен, что позволяет Карпентеру предположить, что анкилозавриды в целом или, по крайней мере, молодые особи не глотали пищу целиком, а использовали какой-то вид жевания. [16] Поскольку взрослый анкилозавр мало пережевывал пищу, он тратил в день на поиск пищи меньше времени, чем слон. [12] Учитывая ширину грудной клетки, перевариванию непережеванной пищи могло способствовать ферментация в задней кишке имеется несколько камер , как у современных травоядных ящериц, у которых в увеличенной толстой кишке . [10]

В 1969 году палеонтолог Георг Хаас пришел к выводу, что, несмотря на большие размеры черепов анкилозавров, связанная с ними мускулатура была относительно слабой. Он также считал, что движение челюсти ограничивается движениями вверх и вниз. Экстраполируя это, Хаас предположил, что анкилозавры питались относительно мягкой неабразивной растительностью. [24] Более поздние исследования Euoplocephalus показали, что у этих животных было возможно движение челюсти вперед и в стороны, причем череп мог выдерживать значительные силы. [25] Исследование окклюзии зубов (контакта между зубами) образцов анкилозавров в 2016 году показало, что способность к движению челюсти назад (палинально) развивалась независимо у разных линий анкилозавров , включая позднемеловых североамериканских анкилозаврид, таких как Ankylosaurus и Euoplocephalus . [22]
втянутое положение ноздрей анкилозавра Арбор и Мэллон в 2017 году сравнили с таковыми у роющих червячных ящериц и слепых змей , и хотя это, вероятно, не было роющим животным, морда Анкилозавра может указывать на поведение, связанное с перемещением земли. Эти факторы, а также низкая скорость образования зубов у анкилозавров по сравнению с другими птицетазами указывают на то, что анкилозавр мог быть всеядным (питался как растительной, так и животной пищей). Он также может (или альтернативно) выкопать землю в поисках корней и клубней . [12] Исследование, проведенное палеонтологом Антонио Балеллом и его коллегами в 2023 году, показало, что североамериканские анкилозавриды позднего мелового периода (включая анкилозавра ) имели челюсти с низким механическим преимуществом, тогда как челюсти более ранних родственников были от высокого до умеренного. Эти поздние анкилозавриды также имели окклюзию зубов и сложный двухфазный механизм челюстей - черты, общие с некоторыми нодозавридами позднего мела, но вместо этого у них были челюсти с высокими механическими преимуществами. Это указывает на то, что, хотя эти две группы сходились по некоторым характеристикам, нодозавры имели более высокую относительную силу укуса , что предполагает различную механику челюстей и разделение рациона между ними. [26]
Воздушное пространство и чувства
[ редактировать ]
В 1977 году палеонтолог Тереза Марьянска предположила, что сложные пазухи и носовые полости анкилозавров, возможно, облегчали вес черепа, содержали носовую железу или служили камерой для голосового резонанса . [10] [27] Карпентер отверг эти гипотезы, утверждая, что четвероногие животные издают звуки через гортань , а не через ноздри, и что снижение веса было минимальным, поскольку промежутки составляли лишь небольшой процент объема черепа. Он также считал наличие железы маловероятной и отмечал, что пазухи, возможно, не имели какой-либо конкретной функции. [10] Также было высказано предположение, что дыхательные пути использовались для обработки вдыхаемого воздуха, как у млекопитающих, на основании наличия и расположения специализированных костей . [27]
Исследование носовых ходов Euoplocephalus , проведенное в 2011 году палеонтологом Тецуто Мияшитой и его коллегами, подтвердило их функцию как системы баланса тепла и воды, отметив обширную систему кровеносных сосудов и увеличенную площадь поверхности слизистой оболочки (используемой для тепло- и водообмена в современном мире). животные). Исследователи также поддержали идею о том, что петли действуют как резонансная камера, сравнимая с удлиненными носовыми ходами сайгаков и петлеобразными трахеями журавлей и лебедей . Реконструкции внутреннего уха предполагают адаптацию к слуху на низких частотах, например, к низким резонансным звукам, которые, возможно, производятся носовыми ходами. Они оспаривали возможность того, что образование петель связано с обонянием (обонянием), поскольку обонятельная область смещается в стороны от основных дыхательных путей. [28]
По мнению Карпентера, форма носовых камер анкилозавра указывает на то, что поток воздуха был однонаправленным (проходя через легкие во время вдоха и выдоха), хотя в задней носовой камере он также мог быть двунаправленным, при этом воздух направлялся мимо обонятельных долей . [10] Увеличенная обонятельная область анкилозаврид свидетельствует о хорошо развитом обонянии. [28] Хотя втягивание ноздрей назад наблюдается у водных животных и животных с хоботком , такая возможность маловероятна для анкилозавра , поскольку ноздри имеют тенденцию уменьшаться, а предчелюстная кость расширяется. Кроме того, хотя широко расставленные ноздри могли обеспечивать стереообоняние (когда каждая ноздря ощущает запахи с разных направлений), как это было предложено для лосей , об этой особенности мало что известно. [12] Положение орбит Анкилозавра предполагает некоторое стереоскопическое зрение . [10]
Движения конечностей
[ редактировать ]
Реконструкция мускулатуры передних конечностей анкилозавра, сделанная Кумбсом в 1978 году, предполагает, что передние конечности выдерживали большую часть веса животного и были приспособлены для приложения больших усилий на передние лапы, возможно, для сбора пищи. Кроме того, Кумбс предположил, что анкилозавры могли быть способными землекопами, хотя копытообразная структура кисти имела ограниченную роющую активность. Анкилозавры, вероятно, были медлительными и медлительными животными. [29] [30] хотя при необходимости они могли быть способны к быстрым движениям. [17]
Рост
[ редактировать ]Чешуйчатые рога самого крупного экземпляра анкилозавра тупее, чем у самого маленького экземпляра, что также характерно для Euoplocephalus , и это может представлять собой онтогенетическую вариацию (связанную с развитием роста). [12] Исследования экземпляров пинакозавров разного возраста показали, что в ходе онтогенетического развития ребра юных анкилозавров срастались с их позвонками. Передние конечности значительно увеличились в прочности, в то время как задние конечности не стали больше по сравнению с остальной частью скелета, что еще раз свидетельствует о том, что руки несли большую часть веса. В шейных полукольцах на подлежащем костном переплете образовались выросты, соединяющие его с подлежащими остеодермами, которые одновременно срослись друг с другом. [31] На черепе средние костные пластинки сначала окостенели на рыле и заднем крае, причем окостенение постепенно распространяется к средним областям. На остальной части тела окостенение прогрессировало от шеи назад в направлении хвоста. [32]
Оборона
[ редактировать ]
Остеодермы анкилозаврид были тонкими по сравнению с остеодермами других анкилозавров и, по-видимому, были укреплены беспорядочно расположенными подушечками коллагеновых волокон. Структурно подобные волокнам Шарпи , они были встроены непосредственно в костную ткань, что является уникальной особенностью анкилозаврид. Это обеспечило бы анкилозавридам броню, которая была бы одновременно легкой и очень прочной, устойчивой к поломке и проникновению зубов хищников. [33] Пальпебральные кости над глазами, возможно, обеспечивали им дополнительную защиту. [34] Карпентер предположил в 1982 году, что сильно васкуляризированный панцирь, возможно, также играл роль в терморегуляции, как и у современных крокодилов . [35]
Хвостовая булава Анкилозавра, по-видимому, была активным защитным оружием, способным нанести удар, способный сломать кости нападавшему. Сухожилия хвоста были частично окостеневшими и не очень эластичными, что позволяло передавать большую силу дубине, когда она использовалась в качестве оружия. [10] В 1979 году Кумбс предположил, что несколько мышц задних конечностей могли контролировать покачивание хвоста и что сильные толчки дубинки могли сломать плюсневые кости крупных теропод . [30]

Исследование 2009 года показало, что анкилозавриды могли поворачивать хвосты на 100 градусов в стороны, а булавы, в основном губчатые, имели пониженный момент инерции и были эффективным оружием. Исследование также показало, что хвостовые булавы взрослых анкилозаврид были способны ломать кости, а у молодых - нет. Несмотря на возможность размахивания хвостом, исследователи не смогли определить, использовали ли анкилозавриды свои булавы для защиты от потенциальных хищников, во внутривидовых боях или для того и другого. [36] Другие исследования обнаружили доказательства того, что анкилозавриды использовали свои хвостовые булавы для внутривидовых боев. У одного экземпляра Tarchia были обнаружены признаки травм как в области таза, так и в области хвоста, а клюшка оказалась асимметричной, что является признаком износа от ударов. [37]
В 1993 году Тони Талборн предположил, что хвостовая булава анкилозаврид в первую очередь действовала как приманка для головы, поскольку он считал хвост слишком коротким и негибким, чтобы обеспечить эффективный досягаемость; «фиктивная голова» могла заманить хищника близко к хвосту, где его можно было ударить. [38] Карпентер отверг эту идею, поскольку форма хвостовой булавы сильно различается у анкилозаврид, даже принадлежащих к одному и тому же роду. [10]
Палеосреда
[ редактировать ]
Анкилозавр существовал между 68 и 66 миллионами лет назад, на заключительном, или маастрихтском, этапе позднего мелового периода. Это был один из последних родов динозавров, появившихся перед мел-палеогеновым вымиранием . Типовой образец взят из формации Хелл-Крик в Монтане, тогда как другие экземпляры были обнаружены в формациях Ланс и Феррис в Вайоминге, формации Сколлард в Альберте и формации Френчмен в Саскачеване, все из которых датируются концом мелового периода. . [39] [40] [12]
Окаменелости анкилозавра редки в отложениях , из которых он известен, и распространение его останков предполагает, что он был экологически редким или ограничивался возвышенностями формаций, где вероятность его окаменения была бы меньшей, чем прибрежные низменности. . Другой анкилозавр, нодозавр, называемый Edmontonia sp., также встречается в тех же образованиях, но, по мнению Карпентера, ареалы двух родов, похоже, не пересекаются. Их останки пока не обнаружены в тех же местах, и нодозавр, по-видимому, населял низменности. Более узкая морда нодозавра предполагает, что у него был более избирательный рацион, чем у анкилозавра , что еще раз указывает на экологическое разделение , независимо от того, перекрывался ли их ареал или нет. [12] [10]
Из-за низкого центра тяжести Анкилозавр не смог бы сбивать деревья, как это делают современные слоны. Он также был неспособен жевать кору и, следовательно, вряд ли практиковал снятие коры. Взрослый анкилозавр , похоже, не собирался в группы (хотя некоторые анкилозавры, похоже, собирались в молодые группы). Поэтому маловероятно, что Анкилозавр смог изменить ландшафт своей экосистемы так, как это делают слоны; Вместо этого гадрозавриды могли выполнять роль « инженера экосистемы ». [12]
Образования, в которых были обнаружены окаменелости анкилозавра , представляют собой различные участки западного берега Западного внутреннего морского пути, разделяющего западную и восточную часть Северной Америки в меловой период, широкую прибрежную равнину, простирающуюся на запад от морского пути до недавно образовавшихся Скалистых гор . Эти образования состоят в основном из песчаника и аргиллита , которые относятся к пойменной среде. [41] [42] [43] В регионах, где были обнаружены анкилозавры и другие позднемеловые анкилозавры, был теплый субтропический / умеренный климат , который был муссонным , с редкими дождями, тропическими штормами и лесными пожарами . [22] В формации Хелл-Крик поддерживались многие виды растений, в первую очередь покрытосеменные , реже хвойные , папоротники и саговники . Обилие ископаемых листьев, найденных в десятках различных мест, указывает на то, что эта территория была в основном засажена небольшими деревьями. [44] Анкилозавр делил свою среду обитания с другими динозаврами, включая цератопсидов Triceratops и Torosaurus , гипсилофодонта Thescelosaurus , гадрозаврида Edmontosaurus , неопределенного нодозавра, пахицефалозавра Pachycephalosaurus и тероподов Struthiomimus , Ornithomimus , Pectinodon и Tyrannosaurus . [40] [45]
Культурное значение
[ редактировать ]
В 2004 году Карпентер отметил, что анкилозавр стал архетипическим членом своей группы и самым известным анкилозавром в популярной культуре , возможно, благодаря реконструкции животного в натуральную величину, представленной на Всемирной выставке 1964 года в Нью-Йорке. [10] Арбор и Мэллон назвали Анкилозавра «культовым» динозавром в 2017 году и отметили, что скульптура Всемирной выставки, а также фреска американского художника Рудольфа Заллингера 1947 года «Эпоха рептилий» и другие более поздние популярные изображения изображали Анкилозавра с хвостовой булавой. после первого открытия этой функции в 1910 году. [12]
На многих традиционных популярных изображениях Анкилозавр изображен сидящим на корточках и с огромной хвостовой дубинкой, волочащейся по земле. Современные реконструкции показывают животное с более вертикальным положением конечностей и хвостом, приподнятым над землей. Точно так же большие шипы, выступающие вбок из тела (похожие на шипы нодозаврид-анкилозавров), присутствуют на многих традиционных изображениях, но не известны у анкилозавра . самого [5] Доспехи Анкилозавра часто отождествляли с доспехами Эдмонтонии (ранее называвшейся Палеосцинк ); Помимо того, что Анкилозавр был изображен с шипами, Эдмонтония также была изображена с хвостовой булавой, похожей на Анкилозавр (особенность, которой не было у нодозавридов), в том числе на фреске американского художника Чарльза Р. Найта 1930 года. [12] Анкилозавр был показан во франшизе « Парк Юрского периода» , где они изображены атакующими хвостами и бегущими - способности, которые палеонтологи раскритиковали как маловероятные. [46] [47] [48]
См. также
[ редактировать ]Ссылки
[ редактировать ]Примечания
- ^ Произносится / ˌ æ ŋ k ə l oʊ ˈ s ɔːr / ə s ANG -kə -loh- SOR -es [1]
Цитаты
- ^ «Анкилозавр» . Словарь Merriam-Webster.com .
- ^ Jump up to: а б с Браун, Б. (1908). «Ankylosauridae, новое семейство панцирных динозавров верхнего мела». Бюллетень Американского музея естественной истории . 24: 187–201. hdl : 2246/1435 .
- ^ Крейслер, Б. (7 июля 2003 г.). «Руководство по переводу и произношению Dinosauria A» . Архивировано из оригинала 18 августа 2010 года . Проверено 3 сентября 2010 г.
- ^ Лидделл, ХГ; Скотт, Р. (1980) [1871]. Греко-английский лексикон (сокращенное изд.). Издательство Оксфордского университета . п. 5 . ISBN 978-0-19-910207-5 .
- ^ Jump up to: а б Перенасыщение, Д.Ф. (1997). «Анкилозавр» . Динозавры, энциклопедия . Издательство McFarland & Company, Inc. стр. 141–143 . ISBN 978-0-375-82419-7 .
- ^ Jump up to: а б Кумбс, В. (1978). «Семейства птицетазовых динозавров отряда Ankylosauria» (PDF) . Журнал палеонтологии . 21 (1): 143–170. Архивировано из оригинала (PDF) 8 июля 2015 года . Проверено 8 июля 2015 г.
- ^ Нэйш, Д. (2009). Великие открытия динозавров . Лондон: A&C Black Publishers LTD. стр. 58–59. ISBN 978-1408119068 .
- ^ Уиллистон, Юго-Запад (1908 г.). «Обзор: Анкилозавриды» . Американский натуралист . 42 (501): 629–30. дои : 10.1086/278987 . JSTOR 2455817 .
- ^ Карпентер, К. (2001). «Глава 21: Филогенетический анализ анкилозавров». В Карпентер, К. (ред.). Бронированные динозавры . Издательство Университета Индианы. стр. 454–83. ISBN 978-0-253-33964-5 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление Карпентер, К. (2004). «Переописание Ankylosaurus magniventris Brown 1908 (Ankylosauridae) из верхнего мела западных внутренних районов Северной Америки». Канадский журнал наук о Земле . 41 (8): 961–86. Бибкод : 2004CaJES..41..961C . дои : 10.1139/e04-043 .
- ^ Осборн, HF (1905). «Тираннозавр и другие плотоядные динозавры мелового периода». Бюллетень АМНХ . 21 (14): 259–265. hdl : 2246/1464 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в Арбур, ВМ; Мэллон, Джей Си (2017). «Необычная краниальная и посткраниальная анатомия архетипического анкилозавра Ankylosaurus magniventris » . ГРАНИ . 2 (2): 764–794. doi : 10.1139/facets-2017-0063 .
- ^ Jump up to: а б Кумбс, В. (1990). «Зубы и систематика анкилозавров». В Карпентере, К.; Карри, Пи Джей (ред.). Систематика динозавров: подходы и перспективы . Издательство Кембриджского университета. стр. 269–79. ISBN 978-0-521-43810-0 .
- ^ Бенсон, РБЖ; Кампионе, Невада; Каррано, Монтана; Мэннион, Полиция; Салливан, К.; и др. (2014). «Скорость эволюции массы тела динозавров указывает на 170 миллионов лет устойчивых экологических инноваций в линии стволовых птиц» . ПЛОС Биол . 12 (5): e1001853. дои : 10.1371/journal.pbio.1001853 . ПМК 4011683 . ПМИД 24802911 .
- ^ Jump up to: а б с д Арбур, ВМ; Карри, Пи Джей (2015). «Систематика, филогения и палеобиогеография динозавров-анкилозаврид». Журнал систематической палеонтологии . 14 (5): 1–60. дои : 10.1080/14772019.2015.1059985 . S2CID 214625754 .
- ^ Jump up to: а б Карпентер, К. (1982). «Детеныши динозавров из позднемеловых формаций Ланс и Хелл-Крик и описание нового вида теропод» . Геология Скалистых гор . 20 (2): 123–134.
- ^ Jump up to: а б Кумбс, WP (1978). «Теоретические аспекты беговых адаптаций динозавров». Ежеквартальный обзор биологии . 53 (4): 393–418. дои : 10.1086/410790 . S2CID 84505681 .
- ^ Арбур, ВМ; Карри, Пи Джей (2015). «Хвостовые булавы динозавров-анкилозаврид развились путем постепенного приобретения ключевых особенностей» . Журнал анатомии . 227 (4): 514–23. дои : 10.1111/joa.12363 . ПМК 4580109 . ПМИД 26332595 .
- ^ Осборн, HF (1923). «Два динозавра нижнего мела Монголии». Новитаты Американского музея (95): 1–10. HDL : 2246/3267 .
- ^ Томпсон, РС; Пэриш, JC; Мейдмент, ЮКР; Барретт, премьер-министр (2012). «Филогения анкилозавровых динозавров (Ornithischia: Thyreophora)». Журнал систематической палеонтологии . 10 (2): 301–312. Бибкод : 2012JSPal..10..301T . дои : 10.1080/14772019.2011.569091 . S2CID 86002282 .
- ^ Арбур, ВМ; Карри, Пи Джей; Бадамгарав, Д. (2014). «Анкилозавридные динозавры верхнемеловых баруунгойотской и нэмегтской свит Монголии» Зоологический журнал Линнеевского общества . 172 (3): 631–652. дои : 10.1111/zoj.12185 .
- ^ Jump up to: а б с Оси, А.; Прондвай, Э.; Мэллон, Дж.; Бодор, скорая помощь (2016). «Разнообразие и конвергенция в эволюции пищевых адаптаций анкилозавров (Dinosauria: Ornithischia)» . Историческая биология . 29 (4): 539–570. дои : 10.1080/08912963.2016.1208194 . S2CID 55372674 .
- ^ Хилл, Р.В.; Д'Эмик, доктор медицины; Бевер, Г.С.; Норелл, Массачусетс (2015). «Сложный гиобранхиальный аппарат мелового динозавра и древность параглоссалий птиц» . Зоологический журнал Линнеевского общества . 175 (4): 892–909. дои : 10.1111/zoj.12293 .
- ^ Хаас, Г. (1969). «О челюстной мускулатуре анкилозавров». Американский музей Novitates (2399): 1–11. hdl : 2246/2609 .
- ^ Рыбчинский, Н.; Викариус, МК (2001). «Глава 14: Свидетельства сложного движения челюстей у позднемеловых анкилозаврид, Euoplocephalus tutus (Dinosauria: Thyreophora)». В К. Карпентере (ред.). Бронированные динозавры . Издательство Университета Индианы. стр. 299–317. ISBN 978-0-253-33964-5 .
- ^ Баллелл, Антонио; Май, Бохао; Бентон, Майкл Дж. (2023). «Дивергентные стратегии в краниальной биомеханике и экологии питания анкилозавровых динозавров» . Научные отчеты . 13 (1): 18242. Бибкод : 2023NatSR..1318242B . дои : 10.1038/s41598-023-45444-1 . ПМК 10600113 . ПМИД 37880323 .
- ^ Jump up to: а б Марьянская, Т. (1977). «Анкилозавриды (Динозавры) из Монголии» (PDF) . Польская палеонтология . 37 : 85–151. Архивировано из оригинала (PDF) 12 июля 2020 г. Проверено 25 июля 2015 г.
- ^ Jump up to: а б Мияшита, Т.; Беседка В.М.; Витмер Л.М.; Карри, Пи Джей (2011). «Внутренняя морфология черепа панцирного динозавра Euoplocephalus, подтвержденная рентгеновской компьютерной томографической реконструкцией» (PDF) . Журнал анатомии . 219 (6): 661–75. дои : 10.1111/j.1469-7580.2011.01427.x . ПМК 3237876 . ПМИД 21954840 . Архивировано из оригинала (PDF) 24 сентября 2015 г.
- ^ Кумбс, В. (1978). «Мышцы передних конечностей анкилозавров (рептилий, орнитиший)». Журнал палеонтологии . 52 (3): 642–57. JSTOR 1303969 .
- ^ Jump up to: а б Кумбс, В. (1979). «Остеология и миология задних конечностей у Ankylosauria (Reptillia, Ornithischia)». Журнал палеонтологии . 53 (3): 666–84. JSTOR 1304004 .
- ^ Бернс, М; Туманова Т; Карри, П. (2015). «Посткрания молодого Pinacosaurus grangeri (Ornithischia: Ankylosauria) из верхнемеловой формации Алагтеэг, Алаг Теег, Монголия: значение для онтогенетической аллометрии анкилозавров». Журнал палеонтологии . 89 (1): 168–182. Бибкод : 2015JPal...89..168B . дои : 10.1017/jpa.2014.14 . S2CID 130610291 .
- ^ Карри, Пи Джей; Бадамгарав, Д.; Коппельхус, Э.Б.; Сиссонс, Р.; Викариус, МК (2011). «Руки, ноги и поведение пинакозавра (Dinosauria: Ankylosauridae)» (PDF ) Польский палеонтологический акт . 56 (3): 489–504. дои : 10.4202/app.2010.0055 . S2CID 129291148 .
- ^ Шайер, ТМ; Сандер, премьер-министр (2004). «Гистология остеодерм анкилозавров: значение для систематики и функций». Журнал палеонтологии позвоночных . 24 (4): 874–93. doi : 10.1671/0272-4634(2004)024[0874:hoaoif]2.0.co;2 . JSTOR 4524782 . S2CID 86333501 .
- ^ Кумбс В. (1972). «Костяное веко Euoplocephalus (Reptilia, Ornithischia)». Журнал палеонтологии . 46 (5): 637–50. JSTOR 1303019 . .
- ^ Карпентер, К. (1982). «Реконструкция скелета и кожного панциря Euoplocephalus tutus (Ornithischia: Ankylosauridae) из позднемеловой формации Олдмана в Альберте». Канадский журнал наук о Земле . 19 (4): 689–97. Бибкод : 1982CaJES..19..689C . дои : 10.1139/e82-058 .
- ^ Арбур, В.М. (2009). «Оценка силы удара хвостовой булавой динозавров-анкилозаврид» . ПЛОС ОДИН . 4 (8): е6738. Бибкод : 2009PLoSO...4.6738A . дои : 10.1371/journal.pone.0006738 . ПМК 2726940 . ПМИД 19707581 .
- ^ Пак Дж.И., Ли Ю.Н., Кобаяши Ю., Джейкобс Л.Л., Барсболд Р., Ли Х.Дж., Ким Н., Сон К.Ю., Полцин М.Дж. (2021). «Новый анкилозаврид из верхнемеловой немегтовой формации Монголии и значение для палеоэкологии панцирных динозавров» . Научные отчеты . 11 (1): Артикул 22928. Цифровой код : 2021NatSR..1122928P . дои : 10.1038/s41598-021-02273-4 . ПМЦ 8616956 . ПМИД 34824329 .
- ^ Талборн, Т. (1993). «Мимикрия у анкилозавридных динозавров». Записи Южно-Австралийского музея . 27: 151–58.
- ^ Викариус, МК; Марьянская, Т.; Вейшампель, Д.Б. (2004). «Анкилозаврия». В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия . Издательство Калифорнийского университета. стр. 363–92. ISBN 978-0-520-24209-8 .
- ^ Jump up to: а б Вейшампель, Д.Б.; Барретт, премьер-министр; Кориа, РА; ЛеЛёфф, Дж.; Сюй С.; Чжао X.; Сахни, А.; Гомани, ЕМП; Ното, CR (2004). «Распространение динозавров». В Вейшампеле, Д.Б.; Додсон, П.; Осмольская, Х. (ред.). Динозаврия (2-я) . Издательство Калифорнийского университета. стр. 517–606. ISBN 978-0-520-24209-8 .
- ^ Лофгрен, Д.Ф. (1997). «Формация Хелл-Крик». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . Академическая пресса. стр. 302–03. ISBN 978-0-12-226810-6 .
- ^ Брейтаупт, Б.Х. (1997). «Лансовый строй». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . Академическая пресса. стр. 394–95. ISBN 978-0-12-226810-6 .
- ^ Эберт, Д.А. (1997). «Эдмонтон Групп». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . Академическая пресса. стр. 199–204. ISBN 978-0-12-226810-6 .
- ^ Джонсон, КР (1997). «Флора Хелл-Крик». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . Академическая пресса. стр. 300–02. ISBN 978-0-12-226810-6 .
- ^ Бигелоу, П. «Меловые фаунистические фации Хелл-Крик; поздний маастрихт» . Архивировано из оригинала 26 декабря 2009 года . Проверено 24 марта 2014 г.
- ^ «Мы спросили палеонтолога, насколько точен «Мир Юрского периода» на самом деле…» Wired UK . 2015 . Проверено 29 августа 2023 г.
- ^ Ваксман, Оливия Б. (2018). «Настоящая научная история динозавров Парка Юрского периода » . Время . Проверено 29 августа 2023 г.
- ^ Дорш, Рита (2022). «Все, что франшиза « Парка Юрского периода» ошибается в палеонтологии » Лупер . Проверено 29 августа 2023 г.
Внешние ссылки
[ редактировать ]
- Анкилозавриды
- Позднемеловые динозавры Северной Америки.
- Ископаемые таксоны описаны в 1908 году.
- Таксоны, названные Барнумом Брауном
- Ланс фауна
- Фауна Хелл-Крик
- Фауна Сколларда
- Палеонтология в Монтане
- Палеонтология в Вайоминге
- Палеонтология в Альберте
- Маастрихтские роды
- Орнитишские роды
- Монотипические роды динозавров
- Тиреофоры Северной Америки
- Позднемеловые тиреофоры