Носовая раковина
| Носовая раковина / носовая раковина | |
|---|---|
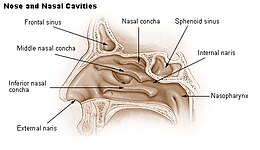
 Боковая стенка носовой полости с изображением решетчатой кости положения . (Верхний и средний — розовым, нижний — синим.) | |
 | |
| Подробности | |
| Идентификаторы | |
| латинский | носовая раковина |
| МеШ | D014420 |
| ФМА | 57456 |
| Анатомические термины кости | |

В анатомии ( носовая раковина / ˈ k ɒ n k ə / ; мн .: conchae ; / ˈ k ɒ n k iː / ; латинское слово «раковина»), также называемая носовой раковиной или раковиной , [1] [2] представляет собой длинный, узкий, изогнутый костный выступ , выступающий в дыхательные пути носа у людей и различных других животных. Раковины имеют форму вытянутой морской ракушки , что и дало им название (лат. concha от греческого κόγχη ). Раковина — любая из закрученных костей носовых ходов позвоночных губчатых . [3]
У людей раковины делят носовые дыхательные пути на четыре желобообразных воздушных прохода и отвечают за то, что вдыхаемый воздух течет устойчивым, регулярным образом вокруг максимально возможной площади поверхности слизистой оболочки носа . Являясь реснитчатой слизистой оболочкой с неглубоким кровоснабжением, слизистая оболочка носа очищает, увлажняет и согревает вдыхаемый воздух, подготавливая его для поступления в легкие .
Быстрое расширение артериолярного кровообращения в этих костях может привести к резкому повышению давления внутри в ответ на резкое охлаждение ядра тела. Боль от такого давления часто называют « замораживанием мозга » и часто связана с быстрым употреблением мороженого . Неглубокость венозного кровоснабжения слизистой оболочки способствует легкости носового кровотечения возникновения .
Структура
[ редактировать ]состоят из псевдомногослойного столбчатого мерцательного Раковины респираторного эпителия с толстым слоем васкуляризированной и эректильной железистой ткани. [4] Раковины расположены латерально в носовых полостях, закручиваются медиально и вниз, в носовые дыхательные пути. Каждая пара состоит из одной раковины с каждой стороны полости носа, разделенной перегородкой . [4]
Верхние раковины представляют собой более мелкие структуры, соединенные со средними раковинами нервными окончаниями и служат для защиты обонятельной луковицы . Верхняя раковина прикрепляется к решетчатой кости . Под верхним проходом находятся отверстия задних решетчатых пазух. [3] Устье клиновидной пазухи находится медиальнее верхней носовой раковины. [5]
Средние раковины меньше, но имеют наиболее сложную анатомию носовых раковин. Они берут начало от латерального края решетчатой пластинки решетчатой кости . Они вставляются спереди в лобный отросток верхней челюсти и сзади в перпендикулярную пластинку небной кости . [6] Различают три взаимно перпендикулярных сегмента средней носовой раковины: от проксимального к дистальному — горизонтальный сегмент ( аксиальная плоскость ), базальная пластинка ( коронковая плоскость ) и вертикальный сегмент ( сагиттальная плоскость ). Они выступают вниз над отверстиями верхнечелюстных , передних и средних решетчатых пазух и действуют как буферы, защищающие пазухи от прямого контакта с потоком носового воздуха под давлением. Большая часть вдыхаемого воздуха проходит между нижней раковиной и средним носовым ходом . [3] У человека их длина обычно равна длине мизинца .
Нижние раковины являются самыми большими носовыми раковинами, их длина у человека может достигать указательного пальца , и они отвечают за большую часть направления воздушного потока, увлажнения, нагревания и фильтрации воздуха, вдыхаемого через нос. [3]
Нижние раковины имеют классификацию 1–4 на основе системы классификации нижних раковин (известной как система классификации нижних носовых раковин ), в которой оценивается общий объем пространства дыхательных путей, занимаемый нижней раковиной. 1-я степень — 0–25% дыхательных путей, 2-я степень — 26–50% дыхательных путей, 3-я степень — 51–75% дыхательных путей и 4-я степень — 76–100% дыхательных путей. [7]
Иногда имеется пара верхних раковин , превосходящих верхние раковины. Если они присутствуют, они обычно имеют форму небольшого гребня.
Функция
[ редактировать ]Раковины составляют большую часть и необходимы слизистой оболочки носа для функционального дыхания . Они обогащены нервными рецепторами, чувствительными к давлению воздушного потока и температуре (связанными с тройничным нервом , пятым черепным нервом ), что обеспечивает огромные эректильные возможности при заложенности и снятии заложенности носа в ответ на погодные условия и меняющиеся потребности организма. [4] Кроме того, эректильная ткань претерпевает часто незамеченный цикл частичной заложенности и разгрузки, называемый носовым циклом . Приток крови к слизистой оболочке носа, в частности к венозному сплетению раковин, регулируется крылонебным ганглием и нагревает или охлаждает воздух в носу.
Носолегочный и носоторакальный рефлексы регулируют механизм дыхания посредством углубления вдоха. Под действием потока воздуха, давления воздуха в носу и качества воздуха импульсы от слизистой оболочки носа передаются тройничным нервом в дыхательные центры в стволе мозга , а генерируемая реакция передается в бронхи и , межреберные диафрагма . мышцы
Раковины также отвечают за фильтрацию , нагревание и увлажнение воздуха, вдыхаемого через нос. Из этих трех фильтрация достигается в основном другими, более эффективными средствами, такими как слизистые оболочки и реснички. Проходя над раковинами, воздух нагревается до 32–34 °C (89–93 °F), увлажняется (до 98% водонасыщения ) и фильтруется. [4]
Иммунологическая роль
[ редактировать ], Респираторный эпителий покрывающий эректильную ткань (или собственную пластинку ) раковин, играет важную роль в первой линии иммунологической защиты организма. частично Респираторный эпителий состоит из слизь продуцирующих бокаловидных клеток, . Эта выделяемая слизь покрывает носовые полости и служит фильтром, улавливая находящиеся в воздухе частицы размером более 2–3 микрометров . Респираторный эпителий также служит средством доступа для лимфатической системы , защищающей организм от заражения вирусами или бактериями. [3]
Запах
[ редактировать ]Раковины обеспечивают, в первую очередь, влажность, необходимую для сохранения нежного обонятельного эпителия , который, в свою очередь, необходим для поддержания здоровья и активности обонятельных рецепторов. Если эпителиальный слой становится сухим или раздраженным, он может перестать функционировать. Обычно это временное состояние, но со временем оно может привести к хронической аносмии . [4] Носовые раковины также увеличивают площадь поверхности внутренней части носа и, направляя и отклоняя поток воздуха через максимальную поверхность слизистой оболочки внутреннего носа, способны продвигать вдыхаемый воздух. Это, в сочетании с влажностью и фильтрацией, обеспечиваемыми раковинами, помогает переносить больше молекул запаха к более высоким и очень узким участкам носовых дыхательных путей, где расположены рецепторы обонятельного нерва. [3]
Верхние раковины полностью покрывают и защищают нервные аксоны, проходящие через решетчатую пластинку (пористую костную пластинку, отделяющую нос от мозга) в нос. Некоторые участки средних раковин также иннервируются обонятельной луковицей. Все три пары раковин иннервируются болевыми и температурными рецепторами через тройничный нерв (или пятый черепной нерв ). [4] Исследования показали, что существует сильная связь между этими нервными окончаниями и активацией обонятельных рецепторов, но науке еще предстоит полностью объяснить это взаимодействие.
Клиническое значение
[ редактировать ]Дисфункция
[ редактировать ]Большие, опухшие раковины, часто называемые в клинике носовыми раковинами, могут привести к блокировке носового дыхания. Аллергия , воздействие раздражителей окружающей среды или постоянное воспаление носовых пазух могут привести к отеку носовых раковин. Деформация носовой перегородки также может привести к увеличению носовых раковин. [8]
Лечение основной аллергии или раздражителя может уменьшить отек носовых раковин. В случаях, которые не проходят, или для лечения искривления перегородки , может потребоваться операция на носовых раковинах.
Операция
[ редактировать ]Уменьшение носовых раковин — это операция по уменьшению размера носовых раковин (обычно нижних носовых раковин ). Существуют различные методы, включая биполярную радиочастотную абляцию (также известную как сомнопластика ), электрокоагуляцию и использование инструментов из холодного оружия (например, микродебридера). Турбинэктомия — это операция по удалению носовых раковин (обычно нижних носовых раковин ).
В случае редукции носовых раковин удаляется лишь небольшое количество ткани носовых раковин, поскольку носовые раковины необходимы для дыхания. Турбинэктомию обычно назначают пациентам, у которых симптомы сохраняются, несмотря на предыдущую операцию по уменьшению носовых раковин . Риски уменьшения нижних или средних носовых раковин включают синдром пустого носа . [8] Доктор Хаузер: «Это особенно верно в случаях резекции передней нижней носовой раковины (ИТ) из-за ее важной роли во внутреннем носовом клапане». [9]
Буллезная раковина — это аномальная пневматизация средней носовой раковины, которая может препятствовать нормальной вентиляции устья пазухи и может привести к рецидивирующему синуситу . В некоторых случаях буллезную раковину можно удалить, чтобы устранить стойкие симптомы.
Другие животные
[ редактировать ]
Обычно у животных носовые раковины представляют собой извитые структуры из тонкой кости или хряща, расположенные в полости носа . Они покрыты слизистой оболочкой , которая может выполнять две функции. Они могут улучшить обоняние за счет увеличения площади, доступной для поглощения переносимых по воздуху химикатов, а также согревать и увлажнять вдыхаемый воздух, а также извлекать тепло и влагу из выдыхаемого воздуха, чтобы предотвратить высыхание легких. Обонятельные раковины имеются у всех современных четвероногих . [ нужна ссылка ] и дыхательные раковины встречаются у большинства млекопитающих и птиц.
Животные с дыхательными носовыми раковинами могут дышать быстрее, не пересушивая легкие, и, следовательно, имеют более быстрый обмен веществ. [10] Например, когда эму выдыхает, его носовые раковины конденсируют влагу из воздуха и поглощают ее для повторного использования. [11] Собаки и другие псовые обладают хорошо развитыми носовыми раковинами. [12] Эти носовые раковины обеспечивают теплообмен между мелкими артериями и венами на их верхнечелюстных поверхностях (носовые раковины, расположенные на кости верхней челюсти ) в противоточной системе теплообмена. [12] Собаки способны к длительной погоне, в отличие от кошек, устраивающих засады, и эти сложные носовые раковины играют в этом важную роль (только у кошек гораздо меньший и менее развитый набор носовых раковин). [12] Эта же сложная структура носовых раковин помогает экономить воду в засушливых условиях. [13] Сохранение воды и терморегуляторные способности этих хорошо развитых носовых раковин у собак, возможно, были решающими адаптациями, которые позволили собакам (включая как домашних собак, так и их диких доисторических предков серых волков ) выживать в суровых арктических условиях и других холодных районах Северной Евразии и Северная Америка, где одновременно очень сухо и очень холодно. [13]
У рептилий и более примитивных синапсидов есть обонятельные раковины, которые участвуют в ощущении запаха, а не в предотвращении высыхания. [14] В то время как челюстные раковины млекопитающих расположены на пути воздушного потока для сбора влаги, сенсорные носовые раковины как у млекопитающих, так и у рептилий расположены дальше назад и над носовым ходом, вдали от потока воздуха. [15] Glanosurus имеет гребни, расположенные низко в полости носа, что указывает на то, что у него были верхнечелюстные раковины, находившиеся на прямом пути воздушного потока. Нижнечелюстные раковины, возможно, не сохранились, поскольку они были либо очень тонкими, либо хрящевыми . Также была высказана вероятность того, что эти гребни связаны с обонятельным эпителием, а не с носовыми раковинами. [16] Тем не менее, возможное наличие верхнечелюстных раковин позволяет предположить, что Glanosurus мог быстро дышать, не пересушивая носовой ход, и, следовательно, мог быть эндотермом. [10] [14] [16]
Кости носовых раковин очень хрупкие и редко сохраняются в виде окаменелостей. В частности, ничего не было обнаружено у ископаемых птиц. [17] Но есть косвенные доказательства их присутствия в некоторых окаменелостях. Рудиментарные гребни, подобные тем, которые поддерживают дыхательные раковины, были обнаружены у продвинутых триасовых цинодонтов , таких как Тринаксодон и Диадемодон . Это говорит о том, что у них мог быть довольно высокий уровень метаболизма. [18] [19] [20] [21] Палеонтолог Джон Рубен и другие утверждали, что у динозавров не было обнаружено никаких свидетельств наличия носовых раковин. У всех динозавров, которые они исследовали, были носовые ходы, которые, как они утверждали, были слишком узкими и слишком короткими для размещения носовых раковин, поэтому динозавры не могли поддерживать частоту дыхания, необходимую для уровня метаболизма млекопитающих или птиц в состоянии покоя, потому что их легкие высох бы. [15] [22] [23] Однако против этого аргумента были высказаны возражения. , носовые раковины отсутствуют или очень малы У некоторых птиц, таких как бескилевые , Procellariiformes и Falconiformes . Они также отсутствуют или очень малы у некоторых млекопитающих, таких как муравьеды, летучие мыши, слоны, киты и большинство приматов, хотя эти животные полностью эндотермичны и в некоторых случаях очень активны. [24] [25] [26] [27] Кроме того, окостеневшие кости носовых раковин были идентифицированы у анкилозавридного динозавра Saichania . [28]
См. также
[ редактировать ]Дополнительные изображения
[ редактировать ]-
 Носовые раковины: заблокированы/свободны.
Носовые раковины: заблокированы/свободны. -
 Нормальный носовой КТ, передний разрез
Нормальный носовой КТ, передний разрез -
 Коронковый срез полостей носа
Коронковый срез полостей носа -
 Правый носовой дыхательный проход
Правый носовой дыхательный проход -
 Носовая оболочка
Носовая оболочка -
 Носовая раковина
Носовая раковина






Примечания
[ редактировать ]- ^ Мейнард, Роберт Льюис; Даунс, Ноэль (2019). «Носовая полость». Анатомия и гистология лабораторных крыс в токсикологии и биомедицинских исследованиях . Эльзевир. стр. 109–121. дои : 10.1016/b978-0-12-811837-5.00010-1 . ISBN 978-0-12-811837-5 .
У человека три раковины определяют ходовые отверстия: Нижний носовой ход: между нижней раковиной/раковиной и дном носовой полости; Средний проход: между средней раковиной/раковиной и нижней раковиной; Верхний проход: между верхней раковиной/раковиной и средней раковиной.
- ^ Карлсон, Брюс М. (2019). «Дыхательная система». Человеческое тело . Эльзевир. стр. 303–319. дои : 10.1016/b978-0-12-804254-0.00011-9 . ISBN 978-0-12-804254-0 . S2CID 79294211 .
- ^ Перейти обратно: а б с д и ж Анатомия человеческого тела. Архивировано 21 января 2013 г. в Wayback Machine Грей, Генри (1918). Носовая полость.
- ^ Перейти обратно: а б с д и ж Дисфункция носовых раковин: сосредоточьтесь на роли нижних носовых раковин в обструкции носовых дыхательных путей. Архивировано 22 июня 2006 г. в Wayback Machine SS Reddy и др. Презентация Гранд Раундов, UTMB, кафедра отоларингологии
- ^ Миллар, Д. Андерсон; Орланди, Ричард Р. (2006). «Естественное отверстие клиновидной пазухи всегда располагается медиальнее верхней носовой раковины» . Американский журнал ринологии . 20 (2): 180–181. дои : 10.1177/194589240602000212 . ISSN 1050-6586 . ПМИД 16686384 . S2CID 20061049 .
- ^ Ли, Хе Ён; Ким, Чан Хун; Ким, Джин Ён; Ким, Джин Кук; Сон, Ми Хён; Ян, Хи Джун; Ким, Кён Су; Чанг, Ин Хюнк; Ли, Юнг-Гвон; Юн, Джу Хон (сентябрь 2006 г.). «Хирургическая анатомия средней носовой раковины» . Клиническая анатомия . 19 (6): 493–496. дои : 10.1002/ок.20202 . ISSN 0897-3806 . ПМИД 16258979 . S2CID 5720770 .
- ^ Камачо, М.; Заги, С.; Черталь, В.; Абдуллатиф Дж.; Значит, К.; Асеведо, Дж.; Лю, С.; Брицке, SE; Кусида, Калифорния; Капассо, Р. (2014). «Система классификации нижних носовых раковин, классы от 1 до 4: разработка и валидация». Ларингоскоп . 125 (2): 296–302. дои : 10.1002/lary.24923 . ПМИД 25215619 . S2CID 34156218 .
- ^ Перейти обратно: а б Уменьшение/удаление нижней носовой раковины из Инфоцентра пазух.
- ^ Хаузер С.М. Хирургическое лечение синдрома пустого носа. Архив отоларингологии хирургии головы и шеи \ Том 133 (№ 9), сентябрь 2007 г.: 858–863.
- ^ Перейти обратно: а б Циммер, К. (1994). «Важность носов» . Обнаружить . 15 (8).
- ^ Мэлони, СК; Доусон, Ти Джей (1998). «Вентиляционная адаптация потребности в кислороде и респираторной потере воды у крупной птицы эму (Dromaius novaehollandiae) и повторное исследование вентиляционной аллометрии птиц». Физиологическая зоология . 71 (6): 712–719. дои : 10.1086/515997 . ПМИД 9798259 . S2CID 39880287 .
- ^ Перейти обратно: а б с Ван (2008) с. 88.
- ^ Перейти обратно: а б Ван (2008) с. 87.
- ^ Перейти обратно: а б Хиллениус, WJ (1994). «Торбинаты у терапсидов: доказательства позднепермского происхождения эндотермии млекопитающих». Эволюция . 48 (2): 207–229. дои : 10.2307/2410089 . JSTOR 2410089 . ПМИД 28568303 .
- ^ Перейти обратно: а б Рубен, Дж.А.; Джонс, Т.Д. (2000). «Селективные факторы, связанные с происхождением меха и перьев» . Американский зоолог . 40 (4): 585–596. дои : 10.1093/icb/40.4.585 .
- ^ Перейти обратно: а б Кемп, Т.С. (2006). «Происхождение эндотермии млекопитающих: парадигма эволюции сложной биологической структуры» . Зоологический журнал Линнеевского общества . 147 (4): 473–488. дои : 10.1111/j.1096-3642.2006.00226.x .
- ^ Уитмер, LM (август 2001 г.). «Положение ноздрей у динозавров и других позвоночных и его значение для функции носа». Наука . 293 (5531): 850–853. CiteSeerX 10.1.1.629.1744 . дои : 10.1126/science.1062681 . ПМИД 11486085 . S2CID 7328047 .
- ^ Бринк, А.С. (1955). «Исследование скелета Диадемодона ». Палеонтология Африканская . 3 :3–39.
- ^ Кемп, Т.С. (1982). Млекообразные рептилии и происхождение млекопитающих . Лондон: Академическая пресса. п. 363. ИСБН 978-0-12-404120-2 .
- ^ Хиллениус, WH (1992). «Эволюция носовых раковин и эндотермия млекопитающих». Палеобиология . 18 (1): 17–29. Бибкод : 1992Pbio...18...17H . дои : 10.1017/S0094837300012197 . JSTOR 2400978 . S2CID 89393753 .
- ^ Рубен, Дж. (1995). «Эволюция эндотермии у млекопитающих и птиц: от физиологии к окаменелостям». Ежегодный обзор физиологии . 57 : 69–95. дои : 10.1146/annurev.ph.57.030195.000441 . ПМИД 7778882 .
- ^ Рубен Дж. А., Джонс Т. Д., Гейст Н. Р. и Хиллениус У. Дж. (ноябрь 1997 г.). «Структура легких и вентиляция легких тероподовых динозавров и ранних птиц». Наука . 278 (5341): 1267–1270. Бибкод : 1997Sci...278.1267R . дои : 10.1126/science.278.5341.1267 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Рубен Дж. А., Хиллениус В. Дж., Гейст Н. Р., Лейтч А., Джонс Т. Д., Карри П. Дж., Хорнер Дж. Р. и Эспе Г. (август 1996 г.). «Метаболический статус некоторых динозавров позднего мела» (PDF) . Наука . 273 (5279): 1204–1207. Бибкод : 1996Sci...273.1204R . дои : 10.1126/science.273.5279.1204 . S2CID 84693210 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Банг, Б.Г. (1966). «Обонятельный аппарат Procellariiformes». Акта Анатомика . 65 (1): 391–415. дои : 10.1159/000142884 . ПМИД 5965973 .
- ^ Банг, Б.Г. (1971). «Функциональная анатомия обонятельной системы 23 отрядов птиц». Акта Анатомика . 79. 79 : 1–76. дои : 10.1159/isbn.978-3-318-01866-0 . ISBN 978-3-8055-1193-3 . ПМИД 5133493 .
- ^ Скотт, Дж. Х. (1954). «Теплорегулирующая функция слизистой оболочки носа». Журнал ларинологии и отологии . 68 (5): 308–317. дои : 10.1017/S0022215100049707 . ПМИД 13163588 . S2CID 32082759 .
- ^ Куломб, Х.Н., Сэм Х. Риджуэй, Ш., и Эванс, МЫ (1965). «Дыхательный водообмен у двух видов морских свиней». Наука . 149 (3679): 86–88. Бибкод : 1965Sci...149...86C . дои : 10.1126/science.149.3679.86 . ПМИД 17737801 . S2CID 38947951 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Марианская, Т. (1977). «Анкилозавриды (Динозавры) из Монголии». Польская палеонтология . 37 : 85–151.
Ссылки
[ редактировать ]- Ван, Сяомин (2008) Собаки: их ископаемые родственники и эволюционная история Издательство Колумбийского университета. ISBN 9780231509435 .