Хальшкараптор
| Хальшкараптор | |
|---|---|
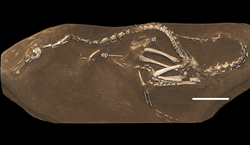 | |
| Образец голотипа | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Дромеозавриды |
| Подсемейство: | † Хальшкарапторины |
| Род: | † Хальшкараптор Кау и др. , 2017 |
| Типовой вид | |
| † Halszkaraptor escuilliei Кау и др. , 2017 | |
Хальшкараптор ( / ˈ h ɑː l ʃ k ə r æ p t ər / ; означает « Хальшки похититель ») — род динозавров водоплавающих - дромеозавридов , из Монголии живших в позднемеловой период . Он содержит только один известный вид , Halszkaraptor escuilliei .
Типовой экземпляр ( голотип ) сравнили с костями современных крокодилов и и водоплавающих птиц обнаружили свидетельства полуводного образа жизни. [1] в то время как некоторые исследователи задаются вопросом о полуводной экологии. [2] [3] Филогенетический анализ показал, что он был членом базального подсемейства Halszkaraptorinae , наряду с Mahakala и Hulsanpes .
История открытия
[ редактировать ]
Образец голотипа Халшкараптора , вероятно, произошел из формации Джадохта в Ухаа Толгоде на юге Монголии и был незаконно вывезен браконьерами в 2011 году или ранее. Окаменелость попала в Японию и Великобританию, где принадлежала нескольким коллекционерам в течение нескольких лет , пока Его приобрела компания Eldonia торговца ископаемыми Франсуа Эскулье. Он определил его как новый вид и в 2015 году отнес в Королевский бельгийский институт естественных наук в Брюсселе , показав палеонтологам Паскалю Годфруа и Андреа Кау для дальнейшей проверки. После проверки его подлинности, среди прочего путем сканирования синхротронным излучением и пучком рентгеновских лучей в Европейской установке синхротронного излучения , Кау и другие видные палеонтологи описали род в подробном исследовании, опубликованном в журнале Nature . Окаменелость была возвращена монгольским властям. [1] [4]

Голотип, MPC D-102/109 , был найден в слое оранжевого песчаника пачки Байн-Дзак формации Джадохта, датируемого поздним кампаном , возрастом около семидесяти пяти миллионов лет. Он состоит из относительно полного скелета с черепом. В 2017 году ископаемое не подвергалось дальнейшей подготовке. Работа торговцев ископаемыми на тот момент в целом обнажила левую сторону скелета. Синхротрон показал , что кости продолжались в скале и что этот кусок, вероятно, не был химерой , искусственным собранием костей разных видов, хотя верхняя часть морды была восстановлена гипсом, а некоторые элементы были повторно прикреплены к скале клей. Скелет в значительной степени сочлененный, не сжатый. Он представляет собой полувзрослую особь возрастом около года. [1]
Типовой вид Halszkaraptor escuilliei был в 2017 году назван и описан Андреа Кау, Винсентом Бейраном, Деннисом ФАЕ Вотеном, Винсентом Фернандесом, Полем Таффоро, Коэном Штайном, Ринченом Барсболдом , Хишигджавом Цогтбаатаром, Филипом Джоном Карри и Паскалем Годфруа. Родовое название сочетает в себе отсылку к покойному польскому палеонтологу Гальшке Осмольской , которая участвовала во многих экспедициях в Монголию и назвала близкородственного Hulsanpes , от латинского raptor — «грабитель». Видовое название дано в честь Эскуилье за то, что он сделал этот образец доступным науке. [1]
Описание
[ редактировать ]
Хальшкараптор был размером с крякву . Голова была около 7 см (2,8 дюйма) в длину, шея - 20 см (7,9 дюйма), спина - 13 см (5,1 дюйма) и крестец - 5 см (2,0 дюйма). Описывающие авторы указали на некоторые отличительные черты. Некоторые из них были аутапоморфиями , уникальными производными персонажами. , Предчелюстная кость передняя мордовая кость, образует уплощенную морду, занимающую 32% длины морды. Предчелюстная кость имеет одиннадцать зубов. Скуловая кость имеет палочковидную форму и ее восходящая ветвь занимает лишь десятую часть перекладины позади глазницы, не доходя до орбиты. Шея чрезвычайно удлиненная, составляет половину длины морды и крестца. Постзигапофизы , задние суставные отростки , шейных позвонков не несут эпипофизов , дополнительных отростков на их верхнем крае. сильно редуцированы В шейных позвонках нервные отростки : на втором-пятом позвонках это лишь низкие гребни, а у последующих шейных позвонков они полностью отсутствуют. Во втором-пятом шейных позвонках в норме парные постзигапофизы срослись в единый лопастный отросток. Нервные отростки хвостовых позвонков чрезвычайно укорочены: у первых трёх хвостовых позвонков они сформированы в виде невысоких бугорков, а у последующих хвостовых позвонков они полностью отсутствуют. Шевроны хвостового основания крупные, пятиугольного профиля. Первая фаланга третьего пальца составляет 47% длины третьего. пястная . [1]
Более того, присутствует уникальное сочетание черт, которые сами по себе не уникальны. Наружная костная ноздря расположена позади основной части предчелюстной кости, в месте, где она соединяется с передней ветвью верхней челюсти . Нисходящая ветвь заглазничной кости имеет палочковидную форму. Число позвонков шеи и спины насчитывает двадцать два. Лишь седьмой, восьмой и девятый шейные позвонки имеют по бокам плевроцели — пневматические впадины. Переход между основанием хвоста и средним хвостом находится на уровне седьмого-восьмого позвонка. Третий палец длиннее второго. [1]
Череп
[ редактировать ]
Морда, хотя и удлиненная, спереди расширена в поперечном направлении, образуя при виде сверху ложкообразный профиль. Он также плоский, а его ширина составляет 180% высоты. Верхний профиль при виде сбоку полый. Расширенная область состоит из относительно длинной предчелюстной кости. Эта кость изнутри выкопана системой воздушных камер. Из более крупной задней камеры сосудисто-нервные каналы пронизывают всю кость, причем не только по бокам, как в Неовенаторе , но и сверху. В этих каналах, вероятно, располагались электросенсорные органы. Каждая предчелюстная кость имеет одиннадцать зубов, что является рекордом среди всех динозавров . Тероподы обычно имеют четыре предчелюстных зуба, а предыдущий рекорд для этой группы был семь, как обнаружено у спинозавридов . У Halskaraptor предчелюстные зубы очень плотно расположены, соприкасаются друг с другом, очень удлинены, постепенно загибаются. Зубы верхней челюсти, числом от двадцати до двадцати пяти, более крепкие, изогнуты только на кончиках и расположены на большем расстоянии. Они более уплощены в поперечном направлении, имеют овальное поперечное сечение. Дентарий нижней челюсти также имеет примерно от двадцати до двадцати пяти зубов. Ноздри относительно втянуты. Они также уникальны для теропод тем, что при виде спереди наклонно ориентированы вверх. Несмотря на длину морды, основное отверстие на боковой части передней части черепа, предглазничное окно , короткое; короче, чем высокий. Задняя черепная крыша сводчатая. [1]
Посткраниальный скелет
[ редактировать ]
хальшкараптора В позвоночнике находятся десять шейных позвонков, двенадцать задних позвонков и шесть крестцовых позвонков . Сохранившиеся хвостовые позвонки включают первые двадцать хвостовых позвонков и ряд из шести от среднего хвоста. Шея очень удлиненная. Она равна 290% длины черепа и 150% длины спины. Это означает, что она составляет половину длины морды-крестца, что является самым высоким значением для всех известных мезозойских паравианов . Среди паравесов только некоторые более поздние птицы имеют пропорционально более длинную шею. Среди более базальных теропод только некоторые овирапторозавры приближаются к этому значению; даже у орнитомимозавров эта цифра никогда не превышает 40%. Длина обусловлена не большим числом позвонков, как у овирапторозаврии, а удлинением отдельных позвонков. Шестой шейный позвонок самый длинный: его длина в четыре раза превышает высоту. Шейные позвонки обычно имеют упрощенное строение, о чем свидетельствует отсутствие задних эпипофизов . Большинство из них не пневматизированы плевроцелями — впадинами, в которых располагаются дивертикулы Воздушные мешочки проникают в костные стенки. [1]

На передней части шеи нервные отростки, обычно прямоугольные пластинки, редуцированы до невысокого гребня; более того, позади они исчезли. У первых пяти шейных позвонков постзигапофизы не имеют разделительного пространства между собой и слиты в одну долю. У других базальных манирапториформ эти задние суставные отростки иногда соединены пластинкой, но в этом случае костный выступ насечен постпозвоночной ямкой, что придает вогнутый профиль на виде сверху; у Halszkaraptor эта бороздка отсутствует, профиль выпуклый. Шейные ребра короткие, не длиннее тел позвонков. Задние позвонки не пневматизированы. Хвост не усилен длинными зигапофизами или шевронами, как у производных эвдромеозавров . Основание хвоста довольно короткое, так как точка перехода к среднему хвосту, где поперечные отростки перестают существовать, находится на восьмом позвонке. Переход также очень постепенный в морфологии. Нервные отростки переднего хвоста уже сильно редуцированы: ими обладают только первые три позвонка и они сформированы в виде невысоких бугорков. [1]
Классификация
[ редактировать ]
Halszkaraptor был помещен в Dromaeosauridae новая клада Halszkaraptorinae в 2017 году . Была создана , включающая Halszkaraptor и его близких родственников Hulsanpes и Mahakala . основана Приведенная ниже кладограмма на филогенетическом анализе, проведенном в 2017 году Cau et al. используя обновленные данные Рабочей группы Theropod. Анализ показал, что Halszkaraptorinae была самой базовой известной группой дромеозаврид. Halszkaraptor занимал базальное положение в Halszkaraptorinae, как сестринская группа клады, образованной Hulsanpes и Mahakala . [1]





Палеобиология
[ редактировать ]
Андреа Кау утверждает, что у Хальшкараптора были характеристики, которые позволяли ему проводить время как в воде, так и на суше, в том числе сильные задние конечности для бега и меньшие, похожие на ласты передние конечности для плавания. Короткий хвост переместил бы центр тяжести вперед, что более полезно для плавания, чем для ходьбы. Туловище должно было держаться более вертикально, чем обычно у теропод. С этой целью существуют приспособления для улучшения разгибания задних конечностей, тазобедренного сустава и бедренной кости. У него было много острых, загнутых назад зубов во рту, длинная шея и сенсорные нейроны на морде, которые, возможно, позволяли ему улавливать вибрации в воде, что заставило ученых полагать, что он охотился на водную добычу. Для размножения ему пришлось выйти на сушу, потому что, как и всем динозаврам, ему нужно было откладывать яйца на суше. [1] Более поздний анализ, проведенный Кау, конкретно указывает на сходство с современными крохалями . Он заявил, что эти птицы, вероятно, являются ближайшими экологическими аналогами Halszkaraptor , поскольку они имеют общие черты с этим таксоном дромеозаврид, такие как длинная шея и зазубренный край морды, используемый для ловли мелкой добычи. Хотя на суше они менее активно передвигаются, принимая позу с вытянутыми бедрами, на воде они используют особую модель плавания , включая передвижение с помощью передних конечностей. Такое поведение также было предположено для Halszkaraptor и, по-видимому, поддерживает рыбоядный и водный образ жизни, аналогичный образу жизни крохалей. [4]
Другие исследователи либо не согласились с интерпретацией Кау, либо просто последовали ей. В 2019 году Браунштейн утверждал, что особенности Хальшкараптора напрямую не подтверждают его способность плавать. Он также предположил, что этот динозавр может быть базальным дромеозавром с переходными чертами. [2] хотя год спустя Кау опроверг свои утверждения. [4] В 2021 году Хоун и Хольц отметили, что, поскольку Хальшкараптор и многие современные водные птицы без уплощенных когтей считаются полуводными, наличие уплощенных когтей, таких как у спинозавра, не обязательно означает, что животное умеет плавать; они не предложили свой собственный взгляд на потенциальную способность этого динозавра плавать. [5] В 2022 году Фаббри и его коллеги выступили против полуводной экологии Хальшкараптора , отметив, что у него низкая плотность костей — черта, не наблюдаемая у полуводных животных. [3] В ответ Кау отметил в своем блоге, что лебеди также имеют низкую плотность костей, но при этом имеют приспособления для полуводного питания. [6] Исследование, проведенное Це, Миллером и Питтманом в 2024 году, посвященное морфологии черепа и силе укуса различных дромеозаврид, обнаружило, что у Хальшкараптора был быстрый укус, не подходящий для кормления рыбоядными животными, как предполагалось ранее, исходя из морфологии его черепа, и вместо этого предположили, что это было насекомоядное животное, которое охотился на мелких беспозвоночных, возможно, в условиях низкой освещенности (ночью или в мутной воде), поскольку он, вероятно, обладал исключительным среди дромеозавридов зрением при слабом освещении из-за его относительно большого размера орбиты. [7]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к Кау, А.; Бейранд, В.; Воетен, DFAE; Фернандес, В.; Таффоро, П.; Штейн, К.; Барсболд, Р.; Цогтбаатар, К.; Карри, Пи Джей; Годфруа, П. (2017). «Синхротронное сканирование выявило экоморфологию земноводных в новой кладе птицеподобных динозавров» . Природа . 552 (7685): 395–399. Бибкод : 2017Natur.552..395C . дои : 10.1038/nature24679 . ПМИД 29211712 . S2CID 4471941 . Дополнительная информация
- ^ Jump up to: а б Браунштейн, CD (2019). « Halszkaraptor escuilliei и эволюция паравианского бауплана» . Природа . 9 (1): 16455. Бибкод : 2019НатСР...916455Б . дои : 10.1038/s41598-019-52867-2 . ПМК 6848195 . ПМИД 31712644 .
- ^ Jump up to: а б Фаббри, Маттео; Навалон, Гильермо; Бенсон, Роджер Б.Дж.; Пол, Диего; О'Коннор, Цзинмай; Бхуллар, Бхарт-Анжан С.; Эриксон, Грегори М.; Норелл, Марк А.; Оркнейские острова, Эндрю; Ламанна, Мэтью С.; Зухри, Самир; Беккер, Жюстин; Эмке, Аманда; Даль Сассо, Криштиану; Бинделлини, Габриэле; Магануко, Симоне; Аудиторе, Марко; Ибрагим, Низар (23 марта 2022 г.). «Подводное кормление хищных динозавров» . Природы . 603 (7903): 852–857. Бибкод : 2022Natur.603..852F . дои : 10.1038/s41586-022-04528-0 . ПМИД 35322229 . S2CID 247630374 .
- ^ Jump up to: а б с Кау, А. (2020). «Сплан тела Halszkaraptor escuilliei (Dinosauria, Theropoda) не является переходной формой в эволюции гиперплотоядных дромеозаврид» . ПерДж . 8 : е8672. дои : 10.7717/peerj.8672 . ПМК 7047864 . ПМИД 32140312 .
- ^ Отточить; Хольц, Томас Р. младший (2021). «Оценка экологии спинозавра: специалист по береговой линии или специалист по водным преследователям?» . Электронная палеонтология . 24 (1): а03. дои : 10.26879/1110 . hdl : 1903/28570 .
- ^ Кау, Андреа (26 марта 2022 г.). «Теропода: влияние плотности костей Хальшкараптора на его экологию» . Проверено 28 марта 2023 г.
- ^ Це, Ю.Т.; Миллер, резюме; Питтман, М. (2024). «Морфологическое несоответствие и структурные характеристики черепа дромеозаврид играют важную роль в экологии и истории эволюции» . BMC Экология и эволюция . 24 (1). 39. Бибкод : 2024BMCEE..24...39T . дои : 10.1186/s12862-024-02222-5 . ПМК 11020771 . ПМИД 38622512 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Хальшкараптором, на Викискладе?
СМИ, связанные с Хальшкараптором, на Викискладе?  Данные, связанные с Хальшкараптором , в Wikispecies
Данные, связанные с Хальшкараптором , в Wikispecies - 3D skeletal model of Halszkaraptor escuilliei at Sketchfab

