Хальшкарапторины
| Хальшкарапторины Временной диапазон: поздний мел ,
| |
|---|---|
| |
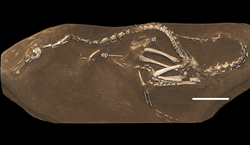
| Голотип Halszkaraptor escuilliei | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Дромеозавриды |
| Подсемейство: | † Хальшкарапторины Кау и др. , 2017 |
| Типовой род | |
| † Halszkaraptor escuilliei Кау и др. , 2017
| |
| Роды | |
| |
Halszkaraptorinae — базальное («примитивное») подсемейство Dromaeosauridae Unenlagiidae (или, возможно, ) , включающее загадочные роды Halszkaraptor , Natovenator , Mahakala и Hulsanpes . Хальшкарапторины окончательно известны только из позднемеловых отложений Азии , в частности в Монголии . Исследователи спорят о том, вела ли эта группа полуводный образ жизни.
История открытия
[ редактировать ]
Первые известные останки гальшкарапторинов были найдены в отложениях песчаника в 1970 году во время польско-монгольской экспедиции на формации Барун-Гойот в пустыне Гоби . Позже в 1982 году они были описаны польским палеонтологом Хальшкой Осмолской и использованы в качестве голотипа для нового рода и вида Hulsanpes perlei в честь монгольского палеонтолога Алтангерела Перле . Хотя родство этих окаменелостей не было до конца понятно, их предварительно интерпретировали как принадлежащие к Deinonychosauria . [2] В 1992 году окаменелости, похожие на дромеозаврид, были обнаружены в местности Тугрикен Шир формации Джадохта и описаны как новый вид в 2007 году Аланом Х. Тернером и его командой. Этот новый таксон, Mahakala omnogovae , считался примитивным дромеозавридом, главным образом, на основании пропорций его тела. [3] В 2015 году Королевский бельгийский институт естественных наук получил небольшой образец окаменелости теропода из частных коллекций Англии и Японии. Образец в результате переговоров между бывшим институтом «Элдония» и властями Монголии был наконец возвращен в Институт палеонтологии и геологии Монгольской академии наук . Согласно документам, связанным с экземпляром, он был незаконно вывезен из джадохтинской свиты (местонахождение Ухаа-Толгод; однако осадки могут свидетельствовать о том, что, наоборот, он был извлечен из близлежащего местонахождения Байн-Дзак) в течение неизвестного года и в конечном итоге попал в частные коллекции. Образец был официально описан в 2017 году Андреа Кау с коллегами, назвав род и вид Halszkaraptor escuilliei в честь Хальшки Осмольской и Франсуа Эскуилье , палеонтологов, которые сделали возможными переговоры о возвращении выловленного браконьером экземпляра в Монголию. С названием Halszkaraptor было также создано подсемейство Halszkaraptorinae, включающее этот таксон и его родственников, а именно Хулсанпес и Махакала . [4]
Описание
[ редактировать ]
Хальшкарапторины были относительно небольшими тероподами, достигавшими длины, сравнимой с длиной современных уток . [3] [4] Их диагностируют по длинным шеям, проксимальным каудальным позвонкам с ориентированными суставными отростками (выступы позвонка совпадают с соседним позвонком) и выступающими зигапофизарными пластинками (костными пластинками, образующими задние стенки каждого позвонка), уплощенной локтевой кости с острым задний край, подвздошная кость с полкообразным надвертельным (над вертелом бедренной кости) отростком, пястной стержень III кости в поперечном направлении такой же толщины, как у II пястной кости, задне-дистальная поверхность на диафизе бедренной кости с удлиненной ямкой, ограниченной латеральным гребнем и проксимальная половина III плюсневой кости не сужена и заметно выпукла. спереди . [4]
Классификация
[ редактировать ]Halszkaraptorinae определяется как наиболее инклюзивная клада, в которую входят Halszkaraptor escuilliei , но не Dromaeosaurus albertensis , Unenlagia comahuensis , Saurornithoides mongoliensis или Vultur gryphus . [4] основана Приведенная ниже кладограмма на филогенетическом анализе, проведенном в 2022 году Lee et al . в их описании Натовенатора . [5]


Палеобиология
[ редактировать ]
В 2017 году сравнение окаменелостей Хальшкараптора с костями современных крокодилов выявило и водоплавающих птиц доказательства полуводного образа жизни. Кау и его коллеги предположили, что хальшкарапторины были обязательными двуногими на суше, а также пловцами , которые использовали передние конечности, чтобы проталкиваться через воду , и использовали свою длинную шею для добычи пищи . Такие черты позволили им эксплуатировать как наземные, так и водные палеоэкосистемы. [4] Исследование, проведенное Це, Миллером и Питтманом в 2024 году, посвященное морфологии черепа и силе укуса различных дромеозаврид, обнаружило, что у Хальскараптора был быстрый укус, не подходящий для кормления рыбоядными животными, как предполагалось ранее, исходя из морфологии его черепа, и вместо этого предположили, что это было насекомоядное животное, которое охотился на мелких беспозвоночных, возможно, в условиях низкой освещенности (ночью или в мутной воде), поскольку он, вероятно, обладал исключительным среди дромеозавридов зрением при слабом освещении из-за его относительно большого размера орбиты. [6]
Другие исследователи либо не согласились, либо просто последовали интерпретации Кау как полуводного динозавра. В 2019 году Браунштейн утверждал, что особенности Хальшкараптора напрямую не подтверждают его способность плавать. Он также предположил, что этот динозавр может быть базальным дромеозавром с переходными чертами. [7] хотя год спустя Кау опроверг свои утверждения. [8] В 2021 году Хоун и Хольц отметили, что, поскольку Хальшкараптор и многие современные водные птицы без уплощенных когтей считаются полуводными, наличие уплощенных когтей, таких как у спинозавра, не обязательно означает, что животное умеет плавать; они не предложили своего собственного взгляда на потенциальную способность этого динозавра плавать. [9] В 2022 году Фаббри и его коллеги выступили против полуводной экологии Хальшкараптора , отметив, что у него низкая плотность костей — черта, не наблюдаемая у полуводных животных. [10] В ответ Кау отметил в своем блоге, что лебеди также имеют низкую плотность костей, но при этом имеют приспособления для полуводного питания. [11]
Ссылки
[ редактировать ]- ^ Ясински, Стивен Э.; Салливан, Роберт М.; Додсон, Питер (26 марта 2020 г.). «Новый динозавр-дромеозаврид (Theropoda, Dromaeosauridae) из Нью-Мексико и биоразнообразие дромеозаврид в конце мелового периода» . Научные отчеты . 10 (1): 5105. Бибкод : 2020NatSR..10.5105J . дои : 10.1038/s41598-020-61480-7 . ISSN 2045-2322 . ПМК 7099077 . ПМИД 32218481 .
- ^ Осмольская, Х. (1982). «Hulsanpes perlei ngnsp. (Deinonychosauria, Saurischia, Dinosauria) из верхнемеловой формации Барун Гойот в Монголии». Новогодний ежегодник по геологии и палеонтологии, ежемесячные выпуски . 1982 (7): 440–448.
- ^ Jump up to: а б Тернер, А.Х.; Пол, Д.; Кларк, Дж.А.; Эриксон, генеральный директор; Норелл, Массачусетс (2007). «Базальный дромеозаврид и эволюция размеров, предшествовавшая полету птиц» . Наука . 317 (5843): 1378–1381. Бибкод : 2007Sci...317.1378T . дои : 10.1126/science.1144066 . ПМИД 17823350 .
- ^ Jump up to: а б с д и Кау, А.; Бейранд, В.; Воетен, DFAE; Фернандес, В.; Таффоро, П.; Штейн, К.; Барсболд, Р.; Цогтбаатар, К.; Карри, Пи Джей; Годфруа, П. (2017). «Синхротронное сканирование выявило экоморфологию земноводных в новой кладе птицеподобных динозавров» . Природа . 552 (7685): 395–399. Бибкод : 2017Natur.552..395C . дои : 10.1038/nature24679 . ПМИД 29211712 . Дополнительная информация
- ^ Ли, С.; Ли, Ю.-Н.; Карри, Пи Джей; Сиссонс, Р.; Парк, Ж.-Ю.; Ким, С.-Х.; Барсболд, Р.; Цогтбаатар, К. (2022). «Нептичий динозавр с обтекаемым телом демонстрирует потенциальные приспособления для плавания» . Коммуникационная биология . 5 (1185). дои : 10.1038/s42003-022-04119-9 . ISSN 2399-3642 . ПМЦ 9715538 .
- ^ Це, Ю.Т.; Миллер, резюме; Питтман, М. (2024). «Морфологическое несоответствие и структурные характеристики черепа дромеозаврид играют важную роль в экологии и истории эволюции» . BMC Экология и эволюция . 24 (1). 39. Бибкод : 2024BMCEE..24...39T . дои : 10.1186/s12862-024-02222-5 . ПМК 11020771 . ПМИД 38622512 .
- ^ Браунштейн, CD (2019). « Halszkaraptor escuilliei и эволюция паравианского бауплана» . Природа . 9 (1): 16455. Бибкод : 2019НатСР...916455Б . дои : 10.1038/s41598-019-52867-2 . ПМК 6848195 . ПМИД 31712644 .
- ^ Кау, А. (2020). «Сплан тела Halszkaraptor escuilliei (Dinosauria, Theropoda) не является переходной формой в эволюции гиперплотоядных дромеозаврид» . ПерДж . 8 : е8672. дои : 10.7717/peerj.8672 . ПМК 7047864 . ПМИД 32140312 .
- ^ Отточить; Хольц, Томас Р. младший (2021). «Оценка экологии спинозавра: специалист по береговой линии или специалист по водным преследователям?» . Электронная палеонтология . 24 (1): а03. дои : 10.26879/1110 . hdl : 1903/28570 .
- ^ Фаббри, Маттео; Навалон, Гильермо; Бенсон, Роджер Б.Дж.; Пол, Диего; О'Коннор, Цзинмай; Бхуллар, Бхарт-Анджан С.; Эриксон, Грегори М.; Норелл, Марк А.; Оркнейские острова, Эндрю; Ламанна, Мэтью С.; Зухри, Самир; Беккер, Жюстин; Эмке, Аманда; Даль Сассо, Криштиану; Бинделлини, Габриэле; Магануко, Симоне; Аудиторе, Марко; Ибрагим, Низар (23 марта 2022 г.). «Подводное кормодобывание среди хищных динозавров» . Природы . 603 (7903): 852–857. Бибкод : 2022Natur.603..852F . дои : 10.1038/s41586-022-04528-0 . ПМИД 35322229 . S2CID 247630374 .
- ^ Кау, Андреа (26 марта 2022 г.). «Теропода: влияние плотности костей Хальшкараптора на его экологию» . Проверено 28 марта 2023 г.

