Альбертозавр
| Альбертозавр | |
|---|---|
 | |
| Установлен слепок в Общественном музее Милуоки. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Тираннозавриды |
| Подсемейство: | † Альбертозаврины |
| Род: | † Альбертозавр Осборн , 1905 год. |
| Разновидность: | † А. саркофаг |
| Биномиальное имя | |
| † Саркофаг Альбертозавра Осборн, 1905 год. | |
| Синонимы | |
| |
Альбертозавр ( / æ l ˌ b ɜːr t ə ˈ s ɔːr ə s / ; означает «ящерица Альберты») — род крупных тероподов-тираннозаврид - динозавров , которые жили на северо-западе Северной Америки в период от раннего до среднего маастрихта позднего мелового периода. около 71 миллиона лет назад . Типовой вид , A. sarcophagus , по-видимому, был ограничен территорией современной канадской провинции Альберта , в честь которой и назван род, хотя неопределенный вид («ср. Albertosaurus sp.») был обнаружен в Коррале де Формации Энмедио и Паккард в Мексике . [1] Ученые расходятся во мнениях относительно содержания рода, а некоторые признают Gorgosaurus libratus вторым видом. [2] [3]
Будучи тираннозавридом, альбертозавр был двуногим хищником с короткими руками, двупалыми руками и массивной головой с десятками больших острых зубов, сильным обонянием, мощным бинокулярным зрением и силой укуса, сокрушающей кости. Возможно, он даже был высшим хищником в своей местной экосистеме . Хотя Альбертозавр , безусловно, был большим для теропода, он все же был намного меньше своего более крупного и известного родственника Тираннозавра рекса , вырастая до 8–9 метров (26–30 футов) в длину и веся 1,7–3,0 метрических тонны (1,9–3,3 метра). короткие тонны).
С момента первого открытия в 1884 году были обнаружены окаменелости более 30 особей, которые дают ученым более подробные знания об анатомии альбертозавра, чем те, которые доступны для большинства других тираннозаврид. Обнаружение 26 особей на одном конкретном участке свидетельствует о стадном поведении и позволяет проводить исследования онтогенеза и популяционной биологии . Это практически невозможно с менее известными динозаврами, потому что их останки более редки и фрагментарны по сравнению с останками альбертозавра .
История открытия
[ редактировать ]Мы
[ редактировать ]
Альбертозавр был назван Генри Фэйрфилдом Осборном в одностраничной заметке в конце его описания тираннозавра рекса в 1905 году . [4] Его тезкой является Альберта , канадская провинция, основанная в том же году, когда были найдены первые останки. Родовое название также включает греческое слово σαυρος / sauros , что означает «ящерица», которое является наиболее распространенным суффиксом в именах динозавров. Типовой вид — Albertosaurus sarcophagus , а видовое название происходит от древнегреческого термина σαρκοφάγος ( саркофагос ), означающего «плотоядный», и имеющего ту же этимологию , что и погребальный контейнер , с которым он разделяет свое название, которое представляет собой комбинацию слов греческие слова σαρξ/ sarx («плоть») и φαγειν / фагеин («есть»). [5] Науке известно более 30 экземпляров всех возрастов. [6] [7]
Ранние открытия
[ редактировать ]
Типовой образец представляет собой частичный череп, собранный 9 июня 1884 года в обнажении формации Хорсшу-Каньон на берегу реки Ред-Дир в Альберте. Его обнаружила экспедиция Геологической службы Канады под руководством известного геолога Джозефа Берра Тиррелла . Из-за отсутствия специального оборудования почти целый череп удалось сохранить лишь частично. В 1889 году коллега Тиррелла Томас Чесмер Уэстон обнаружил неподалеку неполный череп меньшего размера, связанный с некоторым скелетным материалом. [8] Эти два черепа были отнесены к ранее существовавшему виду Laelaps incrassatus Эдвардом Дринкером Коупом в 1892 году. [9] Хотя название Laelaps было связано с родом клещей и было изменено на Dryptosaurus в 1877 году Отниэлем Чарльзом Маршем , Коуп упорно отказывался признавать новое имя, созданное его главным соперником. Однако Лоуренс Ламбе использовал название Dryptosaurus incrassatus вместо Laelaps incrassatus, когда подробно описывал останки в 1903 и 1904 годах. [10] [11] эта комбинация была впервые предложена Оливером Перри Хэем в 1902 году. [12]
Вскоре Осборн отметил, что D. incrassatus был основан на родовых зубах тираннозаврид, поэтому два черепа из каньона Подкова не могут быть с уверенностью отнесены к этому виду. Черепа из каньона Подкова также заметно отличались от останков D. aquilunguis , типового вида Dryptosaurus , поэтому Осборн дал им новое название Albertosaurus sarcophagus в 1905 году. Он не описывал останки сколь-нибудь подробно, сославшись на полное описание Ламбе, сделанное годом ранее. . [5] Оба экземпляра, голотип CMN 5600 и паратип CMN 5601, хранятся в Канадском музее природы в Оттаве . К началу двадцать первого века возникли некоторые опасения, что из-за поврежденного состояния голотипа альбертозавр может быть nomen dubium , который можно использовать только для самого типового экземпляра, поскольку другие окаменелости не могут быть надежно отнесены к нему. Однако в 2010 году Томас Карр установил, что голотип, паратип и сопоставимые более поздние находки имеют одну общую уникальную черту, или аутапоморфию . Наличие увеличенного пневматического отверстия на заднем крае небной кости доказывает, что Albertosaurus является действительным таксоном . [13]
Костный пласт Сухого острова
[ редактировать ]
11 августа 1910 года американский палеонтолог Барнум Браун обнаружил останки большой группы альбертозавров в другом карьере на берегу реки Ред-Дир. Из-за большого количества костей и ограниченного времени группа Брауна не собрала все образцы, но позаботилась о том, чтобы собрать останки всех особей, которых они смогли идентифицировать в костном пласте . Среди костей, хранящихся в Американского музея естественной истории коллекциях в Нью-Йорке, есть семь наборов правых плюсневых костей , а также две изолированные кости пальцев ног, которые не соответствовали ни одной из плюсневых костей по размеру. Это указывало на присутствие в карьере как минимум девяти особей. Палеонтолог Филип Дж. Карри из Королевского Тиррелловского музея палеонтологии заново открыл костное ложе в 1997 году и возобновил полевые исследования на этом участке, который сейчас расположен на территории провинциального парка Драй-Айленд-Баффало-Джамп . [14] Дальнейшие раскопки, проводившиеся с 1997 по 2005 год, обнаружили останки еще 13 особей разного возраста, в том числе миниатюрного двухлетнего ребенка и очень старого человека, длина которого оценивается более 10 метров (33 фута). Ни один из этих людей не известен по полным скелетам, и большинство из них представлены останками в обоих музеях. [6] [15] Раскопки продолжались до 2008 года, когда минимальное количество присутствующих особей было установлено в 12 (на основании сохранившихся элементов, встречающихся в скелете только один раз) и в 26, если учитывать зеркальные элементы, различающиеся по размерам в зависимости от онтогенеза . Всего было обнаружено 1128 костей альбертозавра , что является крупнейшей концентрацией окаменелостей крупных теропод, известных из мелового периода. [16]
Другие открытия
[ редактировать ]
В 1911 году Барнум Браун, во время второго года работы Американского музея естественной истории в Альберте, обнаружил фрагментарный частичный череп альбертозавра у реки Ред-Дир возле Толмен-Бридж (образец AMNH 5222). [17]
Уильям Паркс описал новый вид в 1928 году, Albertosaurus arctunguis , на основе частичного скелета без черепа, который был раскопан Гасом Линдбладом и Ральфом Хорнеллом возле реки Ред-Дир в 1923 году. [18] но с 1970 года этот вид считается идентичным A. sarcophagus . [19] Образец Паркса (ROM 807) хранится в Королевском музее Онтарио в Торонто . [8]
не С 1926 по 1972 год окаменелостей альбертозавра было обнаружено, но с тех пор количество находок увеличилось. Помимо костного пласта на Сухом острове, еще шесть черепов и скелетов были обнаружены в Альберте и хранятся в различных канадских музеях. Экземпляр RTMP 81.010.001 был найден в 1978 году палеонтологом-любителем Морисом Стефанюком. RTMP 85.098.001 был найден Стефанюком 16 июня 1985 года. RTMP 86.64.001 был найден в декабре 1985 года. RTMP 86.205.001 был найден в 1986 году. RTMP 97.058.0001 был найден в 1996 году, а затем есть CMN 11315. К сожалению, ни один из этих скелетов не был найден с целыми черепами. [8] Окаменелости также были обнаружены в американских штатах Монтана, Нью-Мексико, Вайоминг и Миссури , но есть сомнения, что они принадлежат A. sarcophagus и могут даже не принадлежать к роду Albertosaurus . [20] [7]
Два экземпляра «cf Albertosaurus «.sp» были найдены в Мексике ( формация Паккард и формация Коррал-де-Энмедио ). [1]
Горгозавр либратус
[ редактировать ]
В 1913 году палеонтолог Чарльз Х. Штернберг обнаружил еще один скелет тираннозавра в немного более старой формации Парка динозавров в Альберте. Лоуренс Ламбе назвал этого динозавра Gorgosaurus libratus в 1914 году. [21] Другие экземпляры позже были найдены в Альберте и американском штате Монтана . Не найдя существенных различий для разделения двух таксонов (в основном из-за отсутствия хорошего материала черепа альбертозавра ), Дейл Рассел объявил название Gorgosaurus младшим синонимом Albertosaurus . , который был назван первым, а G. libratus был переименован в Albertosaurus libratus в 1970 году Видовое различие сохранялось из-за разницы в возрасте. Это дополнение расширило временной ареал рода Albertosaurus ранее на несколько миллионов лет, а его географический ареал на юг - на сотни километров. [19]
В 2003 году Филип Дж. Карри , воспользовавшись гораздо более обширными находками и общим увеличением анатомических знаний теропод, сравнил несколько черепов тираннозаврид и пришел к выводу, что эти два вида более различны, чем считалось ранее. Поскольку эти два вида являются сестринскими таксонами , они более тесно связаны друг с другом, чем с любым другим видом тираннозаврид. Признавая это, Карри, тем не менее, рекомендовал сохранить Альбертозавра и Горгозавра как отдельные роды, поскольку он пришел к выводу, что они не более похожи, чем Дасплетозавр и Тираннозавр , которые почти всегда разделены. В дополнение к этому, несколько экземпляров альбертозаврина были обнаружены на Аляске и в Нью-Мексико . Карри предположил, что ситуация с альбертозавром и горгозавром может быть прояснена, как только они будут полностью описаны. [7] Большинство авторов последовали рекомендации Карри. [20] [15] [22] но некоторые этого не сделали. [23]
Другие виды
[ редактировать ]В 1930 году Анатолий Николаевич Рябинин назвал Albertosaurus pericolosus по зубу из Китая, который, вероятно, принадлежал тарбозавру . [24] В 1932 году Фридрих фон Хюне переименовал Dryptosaurus incrassatus , не считавшееся сомнительным именем им , в Albertosaurus incrassatus . [25] Поскольку он отождествил горгозавра с альбертозавром , в 1970 году Рассел также переименовал Gorgosaurus sternbergi (Matthew & Brown 1922) в Albertosaurus sternbergi и Gorgosaurus lancensis (Gilmore 1946) в Albertosaurus lancensis . [19] Первый вид сегодня рассматривается как юная форма Gorgosaurus libratus , а второй считается либо идентичным тираннозавру , либо представляющим отдельный род Nanotyrannus . В 1988 году Грегори С. Пол основал Albertosaurus megagracilis на небольшом скелете тираннозавра, экземпляр LACM 28345, из формации Хелл-Крик в Монтане. [26] он был переименован в Динотираннус . В 1995 году [27] но теперь считается, что это молодой тираннозавр рекс . [28] Также в 1988 году Пол переименовал Alectrosaurus olseni (Gilmore 1933) в Albertosaurus olseni , [26] но это не нашло всеобщего признания. В 1989 году Gorgosaurus novojilovi (Малеев 1955) был переименован Брин Мэдер и Робертом Брэдли в Albertosaurus novojilovi . [29]
В двух случаях виды, основанные на действительном материале об альбертозаврах, были переведены в другой род, Deinodon . В 1922 году Уильям Диллер Мэтью переименовал A. sarcophagus в Deinodon sarcophagus . [30] В 1939 году немецкий палеонтолог Оскар Кун переименовал A. arctunguis в Deinodon arctunguis . [31]
Описание
[ редактировать ]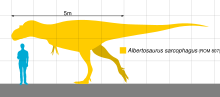
Альбертозавр был довольно крупным двуногим хищником, но меньшего размера, чем Тарбозавр и Тираннозавр рекс . Типичные взрослые особи альбертозавра имели длину до 8–9 м (26–30 футов). [19] [20] [3] и весил от 1,7 до 3,0 метрических тонн (от 1,9 до 3,3 коротких тонны) по массе тела. [32] [3] [33] [2]
Альбертозавр имел сходный внешний вид тела со всеми другими тираннозавридами, с горгозавром в частности . Типичный для теропода, альбертозавр был двуногим и уравновешивал свою большую тяжелую голову и туловище длинным мускулистым хвостом. Однако передние конечности тираннозаврид были чрезвычайно маленькими для своего размера тела и сохранили только два функциональных пальца, причем второй был длиннее первого. Ноги были длинными и заканчивались четырехпалой стопой, на которой первый палец, hallux , был очень коротким и не достигал земли. Третий палец был длиннее остальных. [20] Альбертозавр , возможно, мог развивать скорость ходьбы 14–21 км/час (8–13 миль/час). [34] По крайней мере, для молодых особей высокая скорость бега вполне вероятна. [14]
два отпечатка кожи альбертозавра Известны , и оба имеют чешую . Было обнаружено одно пятно, связанное с некоторыми желудочными ребрами и отпечатком длинной неизвестной кости, что указывает на то, что пятно находится на животе. Чешуя галечная, постепенно становится крупнее и имеет несколько шестиугольную форму. Также сохранились две более крупные масштабные шкалы, расположенные на расстоянии 4,5 см друг от друга. [35] что делает Альбертозавра , наряду с Карнотавром , единственными известными тероподами с сохранившейся чешуей. [36] Еще один отпечаток кожи — с неизвестной части тела. Эти чешуйки маленькие, ромбовидные и расположены рядами. [35]
Череп и зубы
[ редактировать ]
Массивный череп альбертозавра . , располагавшийся на мускулистой короткой S-образной шее, у самых крупных взрослых особей имел длину около 1 метра (3,3 фута) [28] Широкие отверстия в черепе, называемые фенестрами , обеспечивали место для мест прикрепления мышц и органов чувств , что уменьшало его общий вес. Его длинные челюсти содержали с обеих сторон 58 или более зубов банановой формы. Более крупные тираннозавриды обладали меньшим количеством зубов, а у горгозавра их было 62. В отличие от большинства теропод, альбертозавр и другие тираннозавриды были гетеродонтными , с зубами различной формы в зависимости от их положения во рту. Предчелюстные зубы на кончике верхней челюсти, по четыре с каждой стороны, были намного меньше остальных, более плотно упакованы и имели D-образную форму в поперечном сечении . [20] Как и в случае с тираннозавром рексом , верхнечелюстные (щёчные) зубы альбертозавра были адаптированы в общей форме, чтобы противостоять боковым силам, оказываемым борющимся животным-жертвой. Сила укуса альбертозавра была менее огромной, однако максимальная сила со стороны задних зубов достигала 3413 ньютонов. [37] Над глазами располагались короткие костяные гребни, которые, возможно, были ярко окрашены при жизни и, возможно, использовались, в частности, самцами во время ухаживания для привлечения партнера. [38]

В 2001 году Уильям Аблер заметил, что альбертозавра зубцы напоминают трещину в зубе, оканчивающуюся круглой пустотой, называемой ампулой. [39] Зубы тираннозавров использовались в качестве опоры для отрывания плоти от тела, поэтому, когда тираннозавр тянул кусок мяса, напряжение могло привести к тому, что по зубу распространились чисто похожие на трещины зазубрины. [39] Однако наличие ампулы распределяло эти силы по большей площади поверхности и уменьшало риск повреждения зуба при нагрузке. [39] Наличие разрезов, заканчивающихся пустотами, имеет параллели в человеческой инженерии. Производители гитар используют надрезы, оканчивающиеся пустотами, чтобы, как описывает Аблер, «придать чередующиеся области гибкости и жесткости» древесине, над которой они работают. [39] Использование сверла для создания своего рода «ампулы» и предотвращения распространения трещин через материал также используется для защиты поверхностей самолетов. [39] Аблер продемонстрировал, что стержень из плексигласа с надрезами, называемыми «пропилами», и просверленными отверстиями был более чем на 25% прочнее, чем брусок с регулярными надрезами. [39] В отличие от тираннозавров, более древние хищники, такие как фитозавры и диметродон , не имели приспособлений, предотвращающих распространение трещинообразных зазубрин их зубов под воздействием сил питания. [39]
Классификация и систематика
[ редактировать ]Альбертозавр — член семейства теропод Tyrannosauridae, а именно подсемейства Albertosaurinae. Его ближайшим родственником является немного более старый Gorgosaurus libratus (иногда называемый Albertosaurus libratus ; см. ниже). [40] Эти два вида — единственные описанные альбертозаврины, но могут существовать и другие неописанные виды. [7] Томас Хольц обнаружил, что аппалачиозавр был альбертозавром в 2004 году. [20] но в его более поздней неопубликованной работе он отнесен к базальному эотираннозавру, находящемуся недалеко от тираннозаврид, [41] по согласованию с другими авторами. [23]
Другое крупное подсемейство тираннозаврид — Tyrannosaurinae, в которое входят такие представители, как Daspletosaurus , Tarbosaurus и Tyrannosaurus . По сравнению с более крепкими тираннозавринами, альбертозаврины имели стройное телосложение, с пропорционально меньшими черепами и более длинными костями голеней ( голени ) и ступней ( плюсневые кости и фаланги ). [28] [40]

Ниже представлена кладограмма тираннозаврид, основанная на филогенетическом анализе, проведенном Loewen et al. в 2013 году. [42]
Палеобиология
[ редактировать ]Модель роста
[ редактировать ]
Большинство возрастных категорий альбертозавров представлены в летописи окаменелостей . костей Используя гистологию , часто можно определить возраст отдельного животного на момент смерти, что позволяет оценить темпы роста и сравнить его с другими видами. Самый молодой из известных альбертозавров — двухлетний малыш, обнаруженный в костном пласте Сухого острова. Он весил около 50 килограммов (110 фунтов ) и имел длину чуть более 2 метров (6,6 футов). 10-метровому экземпляру (33 фута) из того же карьера 28 лет, это самый старый и крупный из известных экземпляров. Когда экземпляры промежуточного возраста и размера наносятся на график, получается S -образная кривая роста, при этом наиболее быстрый рост происходит в четырехлетний период, заканчивающийся примерно на шестнадцатом году жизни, такая же закономерность наблюдается и у других тираннозаврид. Скорость роста на этом этапе составляла 122 килограмма (269 фунтов) в год при весе взрослого человека 1,3 тонны . Другие исследования показали, что более высокий вес взрослых особей может повлиять на величину темпов роста, но не на общую картину. Тираннозавриды по размерам сходны с Альбертозавр имел аналогичные темпы роста, хотя гораздо более крупный тираннозавр рекс рос почти в пять раз быстрее (601 килограмм [1325 фунтов] в год) на пике своего развития. [6] Окончание фазы быстрого роста предполагает наступление половой зрелости у альбертозавра , хотя рост продолжался более медленными темпами на протяжении всей жизни животных. [6] [15] Половое созревание при активном росте, по-видимому, является общей чертой среди маленьких детей. [43] и большой [44] динозавров, а также крупных млекопитающих, таких как люди и слоны . [44] Такая закономерность относительно раннего полового созревания разительно отличается от закономерности у птиц, у которых половая зрелость задерживается до тех пор, пока они не перестанут расти. [22] [44]
В процессе роста утолщение морфологии зубов изменилось настолько, что, если бы ассоциация молодых и взрослых скелетов на костном ложе Сухого острова не доказала их принадлежность к одному и тому же таксону, зубы молодых особей, вероятно, были бы идентифицированы с помощью статистического анализа как другого вида. [45]
История жизни
[ редактировать ]
Большинство известных особей альбертозавра на момент смерти были в возрасте 14 лет и старше. Молодые животные редко окаменевают по нескольким причинам, главным образом из-за предвзятости сохранности , когда более мелкие кости более молодых животных с меньшей вероятностью сохранялись в результате окаменения, чем более крупные кости взрослых, и из-за предвзятости коллекционирования, когда коллекционеры с меньшей вероятностью заметят более мелкие окаменелости. в поле. [46] Молодые альбертозавры относительно велики для молодых животных, но их останки все еще редки в летописи окаменелостей по сравнению с взрослыми особями. Было высказано предположение, что это явление является следствием истории жизни , а не предвзятостью, и что окаменелости молодых альбертозавров редки, потому что они просто не умирали так часто, как взрослые особи. [6]
Гипотеза истории жизни альбертозавра постулирует, что вылупившиеся птенцы умерли в большом количестве, но не сохранились в летописи окаменелостей из-за их небольшого размера и хрупкой конструкции. Всего через два года молодые особи стали крупнее любого другого хищника в регионе, за исключением взрослого альбертозавра , и более быстроногими, чем большинство их жертв. Это привело к резкому снижению уровня их смертности и соответствующей редкости ископаемых останков. Уровень смертности удвоился в возрасте двенадцати лет, возможно, в результате физиологических потребностей фазы быстрого роста, а затем снова удвоился с наступлением половой зрелости в возрасте от четырнадцати до шестнадцати лет. Этот повышенный уровень смертности продолжался на протяжении всей взрослой жизни, возможно, из-за высоких физиологических потребностей деторождения, включая стресс и травмы, полученные во время внутривидовой конкуренции за партнеров и ресурсы, а также из-за возможных, постоянно усиливающихся эффектов старения . Более высокий уровень смертности среди взрослых может объяснить их более частое сохранение. Очень крупные животные были редкостью, потому что немногие особи выживали достаточно долго, чтобы достичь таких размеров. Высокий уровень младенческой смертности, за которым следует снижение смертности среди молоди и внезапное увеличение смертности после половой зрелости, при этом очень немногие животные достигают максимального размера, - это закономерность, наблюдаемая у многих современных крупных млекопитающих, включая слонов. Африканский буйвол и носорог . Та же картина наблюдается и у других тираннозаврид. Сравнение с современными животными и другими тираннозавридами подтверждает эту гипотезу истории жизни, но предвзятость в летописи окаменелостей все еще может играть большую роль, тем более что более двух третей всех экземпляров альбертозавра известны из одной и той же местности. [6] [22] [47]
Поведение пакета
[ редактировать ]
Костное ложе Сухого острова, обнаруженное Барнумом Брауном и его командой, содержит останки 26 альбертозавров . Это наибольшее количество особей, найденных в одном местонахождении среди крупных теропод мелового периода, и второй по величине среди всех крупных динозавров-тероподов после группы аллозавров в динозавре Кливленда-Ллойда. Карьер в штате Юта . Группа, судя по всему, состоит из одного очень старого взрослого человека, восьми взрослых в возрасте от 17 до 23 лет, семи суб-взрослых, находящихся в фазе быстрого роста в возрасте от 12 до 16 лет, и шести подростков в возрасте от 2 до 11 лет. которые еще не достигли фазы роста. [6]
Практически полное отсутствие останков травоядных животных и сходное состояние сохранности, характерное для многих особей в карьере, где добывали кости альбертозавра, привели Карри к выводу, что это место не является ловушкой для хищников, такой как смоляные ямы Ла Бреа в Калифорнии , и что все сохранившиеся животные погибли одновременно. Карри утверждает, что это свидетельство поведения стаи. [14] Другие ученые настроены скептически, отмечая, что животные могли быть согнаны вместе из-за засухи, наводнения или по другим причинам. [6] [46] [48]

Существует множество свидетельств стадного поведения травоядных динозавров, включая цератопсов и гадрозавров . [49] Однако лишь изредка на одном и том же месте встречается такое количество хищников-динозавров. Маленькие тероподы, такие как дейноних. [50] и Целофизис , [51] были обнаружены в скоплениях, как и более крупные хищники, такие как аллозавр и мапузавр . [52] Есть некоторые свидетельства стадного поведения и у других тираннозаврид: фрагментарные останки более мелких особей были найдены рядом со « Сью », тираннозавром, установленным в Полевом музее естественной истории в Чикаго , а костное ложе в формации Ту-Медисин в Монтане содержит по крайней мере три экземпляра дасплетозавра сохранились вместе с несколькими гадрозаврами. [53] Эти результаты могут подтвердить данные о социальном поведении альбертозавра , хотя некоторые или все из вышеперечисленных мест могут представлять собой временные или неестественные скопления. [14] Другие предполагают, что вместо социальных групп, по крайней мере, некоторые из этих находок представляют собой толпу туш, похожую на драконов Комодо некоторых хищников , где агрессивная конкуренция приводит к гибели и даже каннибализации . [46] О доказательствах каннибализма позже сообщили в 2024 году Коппок и Карри. [54]
Карри также размышлял о охотничьих привычках альбертозавра . Пропорции ног более мелких особей были сопоставимы с пропорциями ног орнитомимид , которые, вероятно, были одними из самых быстрых динозавров. Молодые альбертозавры, вероятно, были столь же быстроногими или, по крайней мере, быстрее, чем их добыча. Карри предположил, что младшие члены стаи, возможно, были ответственны за то, что подгоняли свою добычу к взрослым, которые были крупнее и мощнее, но при этом медленнее. [14] Молодые особи, возможно, также вели другой образ жизни, чем взрослые, заполняя ниши хищников между огромными взрослыми особями и более мелкими современными тероподами, самые крупные из которых были на два порядка меньше, чем взрослый альбертозавр по массе. [20] Похожая ситуация наблюдается у современных варанов Комодо: вылупившиеся детеныши начинают жизнь как мелкие насекомоядные, а затем вырастают и становятся доминирующими хищниками на своих островах. [55] Однако, поскольку сохранение поведения в летописи окаменелостей встречается крайне редко, эти идеи нелегко проверить. В 2010 году Карри, хотя и по-прежнему поддерживал гипотезу охотничьей стаи, признал, что концентрация могла быть вызвана другими причинами, такими как медленно поднимающийся уровень воды во время продолжительного наводнения. [56]
Палеопатология
[ редактировать ]
В 2009 году исследователи выдвинули гипотезу, что отверстия с гладкими краями, обнаруженные в ископаемых челюстях динозавров -тираннозаврид , таких как Альбертозавр , были вызваны паразитом, похожим на Trichomonas Gallinae , который заражает птиц. [57] Они предположили, что тираннозавриды передавали инфекцию, кусая друг друга, и что инфекция нарушала их способность есть. [57]
В 2001 году Брюс Ротшильд и другие опубликовали исследование, в котором изучались доказательства стрессовых переломов и отрывов сухожилий у теропод- динозавров, а также их последствия для их поведения. Они обнаружили, что только одна из 319 костей стопы альбертозавра , проверенных на наличие стрессовых переломов, на самом деле имела их, и ни одна из четырех костей рук не имела их. Ученые обнаружили, что стрессовые переломы «значительно» реже встречались у альбертозавра, чем у карнозавра аллозавра . [58] ROM 807, голотип A. arctunguis (теперь называемый A. sarcophagus ), имел глубокое отверстие размером 2,5 на 3,5 см (0,98 на 1,38 дюйма) в лопатке подвздошной кости , хотя описатель вида не признал это патологией. В образце также обнаружен экзостоз четвертой левой плюсневой кости. два из пяти экземпляров саркофага альбертозавра с плечевыми суставами имели патологические повреждения. В 1970 году Дейл Рассел сообщил, что [59]
состоянии здоровья популяции альбертозавров В 2010 году сообщалось о на Сухом острове. У большинства экземпляров не было обнаружено никаких признаков заболевания. На трех фалангах стопы присутствовали странные костные шпоры, состоящие из аномальных оссификаций сухожилий, так называемых энтезофитов , но причина их неизвестна. Два ребра и брюшное ребро имели признаки перелома и заживления. У одного взрослого экземпляра на левой нижней челюсти была колотая рана, а также зажившие и незажившие следы укусов. Низкое количество отклонений выгодно отличается от состояния здоровья популяции майюнгазавра , у которой в 2007 году было установлено, что у 19% особей наблюдались костные патологии. [60]
Палеоэкология
[ редактировать ]
Большинство окаменелостей саркофага альбертозавра известно из верхней части формации Каньон Подкова в Альберте. Эти более молодые образования этой геологической формации датируются ранним маастрихтом позднего мелового периода, примерно 70–68 миллионов лет назад. Непосредственно под этим образованием находится сланец Медвежья Лапа , морское образование, представляющее собой часть Западного внутреннего морского пути . Внутреннее море отступало по мере того, как климат охлаждался, а уровень моря падал к концу мелового периода, обнажая таким образом сушу, которая ранее находилась под водой. Однако это не был гладкий процесс, и морской путь периодически поднимался, охватывая части региона по всему каньону Подкова, а затем, наконец, полностью отступал в последующие годы. Из-за изменения уровня моря в формации Каньон Подкова представлено множество различных сред, включая прибрежные и прибрежные морские среды обитания, а также прибрежные среды обитания, такие как лагуны , устья рек и приливные отмели . Многочисленные угольные пласты представляют собой древние торфяные болота . Как и большинство других окаменелостей позвоночных из этой формации, останки альбертозавра находят в отложениях, отложенных в дельтах и поймах крупных рек во второй половине времен каньона Подкова. [61]
Фауна формации Каньон Подкова хорошо известна, поскольку окаменелости позвоночных, в том числе динозавров, очень распространены. акулы , скаты , осетры , боуфины , щуки , аспидоринхиусы Ихтиофауну составляли . Среди млекопитающих были многобугорковые и сумчатый Didelphodon . Морской плезиозавр Leurospondylus был обнаружен в морских отложениях в каньоне Подкова, в то время как пресноводная среда была населена черепахами , Champsosaurus и крокодилами, такими как Leidyosuchus и Stangerochampsa . В фауне доминируют динозавры, особенно гадрозавры, составляющие половину всех известных динозавров. К ним относятся роды Edmontosaurus , Saurolophus и Hypacrosaurus . Цератопсы и орнитомимиды также были очень распространены, вместе составляя еще треть известной фауны. Наряду с гораздо более редкими анкилозаврами и пахицефалозаврами , все эти животные могли стать добычей разнообразных плотоядных теропод, включая троодонтид , дромеозаврид и ценагнатидов . [61] [62] Среди останков альбертозавра кости небольшого теропода Albertonykus . в костном пласте Сухого острова были найдены [63] Взрослые альбертозавры были высшими хищниками в своей среде обитания, а промежуточные ниши, возможно, были заполнены молодыми альбертозаврами . [61]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Внесен в список «ср. Albertosaurus sp.» «Коррал Де Энмедио и формации Паккард, группа Каболлона, Сонора, Мексика» у Салливана и Лукаса (2006). Страница 16.
- ^ Перейти обратно: а б Ларсон, П.; Карпентер, К., ред. (2008). Тираннозавр рекс , Король-Тиран (Жизнь прошлого) . Издательство Университета Индианы. п. 310. ИСБН 9780253350879 .
- ^ Перейти обратно: а б с Пол, GS (2016). Принстонский полевой справочник по динозаврам (второе изд.). Издательство Принстонского университета. п. 112. ИСБН 978-0-691-16766-4 .
- ^ Осборн, HF (31 декабря 1905 г.). «Тираннозавр и другие плотоядные динозавры мелового периода» . Бюллетень Американского музея естественной истории . 21 : 259–265. дои : 10.5281/zenodo.1038222 .
- ^ Перейти обратно: а б Осборн, Генри Ф. (1905). « Тираннозавр и другие хищные динозавры мелового периода» (PDF) . Бюллетень Американского музея естественной истории . 21 (3): 259–265. дои : 10.1111/j.1468-5965.2007.00735_17.x . hdl : 2246/1464 .
- ^ Перейти обратно: а б с д и ж г час Эриксон, Грегори М.; Карри, Филип. Дж.; Иноуе, Брайан Д.; Винн, Элис А. (2006). «Таблицы жизни тираннозавров: пример биологии популяции нептичьих динозавров» (PDF) . Наука . 313 (5784): 213–217. Бибкод : 2006Sci...313..213E . дои : 10.1126/science.1125721 . ПМИД 16840697 . S2CID 34191607 . Архивировано из оригинала (PDF) 18 июля 2010 года . Проверено 29 августа 2010 г.
- ^ Перейти обратно: а б с д Карри, Филип Дж. (2003). «Краниальная анатомия тираннозаврид позднего мела Альберты» (PDF) . Acta Palaeontologica Polonica . 48 (2): 191–226. Архивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Перейти обратно: а б с Танке, Даррен Х.; Карри, Филип Дж. (2010). «История открытия альбертозавра в Альберте, Канада». Канадский журнал наук о Земле . 47 (9): 1197–1211. Бибкод : 2010CaJES..47.1197T . дои : 10.1139/e10-057 .
- ^ Коуп, Эдвард Д. (1892). «На черепе динозавра Laelaps incrassatus Cope». Труды Американского философского общества . 30 : 240–245.
- ^ Ламбе, LM (1903). «На нижней челюсти Dryptosaurus incrassatus (Cope)». Оттавский натуралист . 17 : 134.
- ^ Ламбе, Лоуренс М. (1904). «О Dryptosaurus incrassatus (Cope) из Эдмонтонской серии Северо-Западной территории» . Вклад в канадскую палеонтологию . 3 :1–27 . Проверено 29 августа 2010 г.
- ^ Хэй, Оливер Перри , 1902 г. Библиография и каталог ископаемых позвоночных Северной Америки . Бюллетень Геологической службы США, № 117, государственная типография. стр. 868
- ^ Карр, Томас Д. (2010). «Таксономическая оценка типовой серии саркофага альбертозавра и идентичность тираннозаврид (Dinosauria, Coelurosauria) в костном пласте альбертозавра из формации каньона Подкова (кампан-маастрихт, поздний мел)» . Канадский журнал наук о Земле . 47 (9): 1213–1226. Бибкод : 2010CaJES..47.1213C . дои : 10.1139/e10-035 .
- ^ Перейти обратно: а б с д и Карри, Филип Дж . (1998). «Возможные доказательства стадного поведения тираннозаврид» (PDF) . Гайя . 15 : 271–277. Архивировано из оригинала (PDF) 26 марта 2009 г. Проверено 3 мая 2009 г. (не печаталось до 2000 г.)
- ^ Перейти обратно: а б с д Эриксон, Грегори М.; Маковицкий, Питер Дж.; Карри, Филип Дж.; Норелл, Марк А.; Йерби, Скотт А.; Брошу, Кристофер А. (2004). «Гигантизм и сравнительные параметры жизненного цикла динозавров-тираннозаврид» (PDF) . Природа . 430 (7001): 772–775. Бибкод : 2004Natur.430..772E . дои : 10.1038/nature02699 . ПМИД 15306807 . S2CID 4404887 . Архивировано из оригинала (PDF) 6 октября 2011 года . Проверено 29 августа 2010 г. (Ошибка: дои : 10.1038/nature16487 , PMID 26675726 , Часы втягивания )
- ^ Эберт, Дэвид А.; Карри, Филип Дж. (2010). «Стратиграфия, седиментология и тафономия костного пласта Альбертозавра (верхняя часть формации Хорсшу-Каньон; Маастрихт), южная Альберта, Канада». Канадский журнал наук о Земле . 47 (9): 1119–1143. Бибкод : 2010CaJES..47.1119E . дои : 10.1139/e10-045 .
- ^ Карпентер, К., 1992, «Тираннозавриды (динозавры) Азии и Северной Америки», В: Н. Матер и П.-Дж. Чен (ред.) Аспекты неморской геологии мелового периода . China Ocean Press, Пекин, Китай, стр. 250–268.
- ^ Паркс, Уильям А. (1928). « Albertosaurus arctunguis , новый вид тераподных динозавров из Эдмонтонской формации Альберты». Исследования Университета Торонто, геологическая серия . 25 : 1–42.
- ^ Перейти обратно: а б с д Рассел, Дейл А. (1970). «Тираннозавры из позднего мела западной Канады». Публикации Национального музея естественных наук по палеонтологии . 1 :1–34.
- ^ Перейти обратно: а б с д и ж г Хольц, Томас Р. (2004). «Тираннозавроида». В Вейшампеле, Дэвид Б .; Додсон, Питер ; Осмольска, Гальшка (ред.). Динозаврия (Второе изд.). Беркли: Издательство Калифорнийского университета. стр. 111–136. ISBN 978-0-520-24209-8 .
- ^ Ламбе, Лоуренс М. (1914). «О новом роде и виде хищных динозавров из формации Белли-Ривер в Альберте, с описанием черепа Stephanosaurus Marginatus из того же горизонта». Оттавский натуралист . 28 : 13–20.
- ^ Перейти обратно: а б с Риклефс, Роберт Э. (2007). «Старение тираннозавра» . Письма по биологии . 3 (2): 214–217. дои : 10.1098/rsbl.2006.0597 . ПМЦ 2375931 . ПМИД 17284406 .
- ^ Перейти обратно: а б Карр, Томас Д.; Уильямсон, Томас Э.; Швиммер, Дэвид Р. (2005). «Новый род и вид тираннозавроидов из позднемеловой (средний кампан) формации Демополис в Алабаме» . Журнал палеонтологии позвоночных . 25 (1): 119–143. doi : 10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2 . S2CID 86243316 .
- ^ Рябинин А. Н. (1930). «[На русском] «О возрасте и фауне динозавровых отложений на Амуре». Воспоминания Русского минералогического общества . 59 (2): 41–51.
- ^ Фон Хюэне, Ф. , 1932. Ископаемый отряд рептилий Saurischia: его развитие и история . Монография по геологии и палеонтологии, части I и II, сер. Я, 4: 1–361
- ^ Перейти обратно: а б Пол, Грегори С. (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. ISBN 978-0-671-61946-6 .
- ^ Ольшевский, Георгий. (1995). «(Происхождение и эволюция тираннозаврид.)». Кёрюгаку Сайзенсен (Dino Frontline) (на японском языке). 9 : 92–119.
- ^ Перейти обратно: а б с Карри, Филип Дж. (2003). «Аллометрический рост тираннозаврид (Dinosauria: Theropoda) из верхнего мела Северной Америки и Азии» (PDF) . Канадский журнал наук о Земле . 40 (4): 651–665. Бибкод : 2003CaJES..40..651C . дои : 10.1139/e02-083 .
- ^ Мадер, Б.; Брэдли, Р. (1989). «Переописание и пересмотренный диагноз синтипов монгольского тираннозавра Electrosaurus olseni ». Журнал палеонтологии позвоночных . 9 (1): 41–55. Бибкод : 1989JVPal...9...41M . дои : 10.1080/02724634.1989.10011737 .
- ^ Мэтью, штат Вашингтон; Браун, Б. (1922). «Семейство Deinodontidae с уведомлением о новом роде из мелового периода Альберты». Бюллетень Американского музея естественной истории . 46 (6): 367–385.
- ^ Кун, О. , 1939 Saurischia - Каталог окаменелостей I, Animalia, Часть 87 . 'с-Грейвенхейдж, В. Джанк, 1939, 124 стр.
- ^ Кристиансен, Пер; Фаринья, Ричард А. (2004). «Массовое предсказание динозавров-теропод». Историческая биология . 16 (2–4): 85–92. дои : 10.1080/08912960412331284313 . S2CID 84322349 .
- ^ Кампионе, Невада; Эванс, округ Колумбия; Браун, СМ; Каррано, Монтана (2014). «Оценка массы тела у двуногих, не являющихся птицами, с использованием теоретического преобразования в пропорции шилоподий четвероногих» . Методы экологии и эволюции . 5 (9): 913–923. Бибкод : 2014MEcEv...5..913C . дои : 10.1111/2041-210X.12226 . hdl : 10088/25281 .
- ^ Талборн, Ричард А. (1982). «Скорость и походка динозавров». Палеогеография, Палеоклиматология, Палеоэкология . 38 (3–4): 227–256. Бибкод : 1982PPP....38..227T . дои : 10.1016/0031-0182(82)90005-0 .
- ^ Перейти обратно: а б Белл, Фил; Э. Кампионе, Николас; Скотт Персонс, Вт; Дж. Карри, Филип; Ларсон, Питер; Танке, Даррен; Т. Баккер, Роберт (1 июня 2017 г.). «Покровы тираннозавроидов демонстрируют противоречивые закономерности гигантизма и эволюции перьев» . Письма по биологии . 13 (6): 20170092. doi : 10.1098/rsbl.2017.0092 . ПМЦ 5493735 . ПМИД 28592520 .
- ^ Хендрикс, Кристоф; Белл, Фил Р.; Питтман, Майкл; Милнер, Эндрю Р.К.; Куэста, Елена; О'Коннор, Цзинмай; Лоуэн, Марк; Карри, Филип Дж.; Матеус, Октавио; Кэй, Томас Г.; Делькур, Рафаэль (июнь 2022 г.). «Морфология и распределение чешуи, кожных оссификаций и других неперьевых покровных структур у нептичьих динозавров-тероподов» . Биологические обзоры . 97 (3): 960–1004. дои : 10.1111/brv.12829 . ISSN 1464-7931 . ПМИД 34991180 . S2CID 245820672 .
- ^ Райхель, Мириам (2010). «Гетеродонтность саркофага альбертозавра и тираннозавра рекса : биомеханические последствия, выведенные с помощью трехмерных моделей». Канадский журнал наук о Земле . 47 (9): 1253–1261. Бибкод : 2010CaJES..47.1253R . дои : 10.1139/e10-063 .
- ^ «Альбертозавр». В: Додсон, Питер; Бритт, Брукс; Карпентер, Кеннет; Форстер, Кэтрин А.; Джилетт, Дэвид Д.; Норелл, Марк А.; Ольшевский, Георгий; Пэрриш, Дж. Майкл; и Вейшампель, Дэвид Б. Эпоха динозавров . Линкольнвуд, Иллинойс: Publications International, Ltd., 1993. стр. 106–107. ISBN 0-7853-0443-6 .
- ^ Перейти обратно: а б с д и ж г Аблер, В.Л. 2001. Модель зубцов тираннозавра, сделанная с помощью пропила и сверла. п. 84–89. В: Жизнь мезозойских позвоночных . Под ред. Танке, Д.Х., Карпентер, К., Скрепник, М.В., Издательство Индианского университета.
- ^ Перейти обратно: а б Карри, Филип Дж .; Хурум, Йорн Х; Сабат, Кароль (2003). «Строение черепа и эволюция в филогении тираннозаврид» (PDF) . Acta Palaeontologica Polonica . 48 (2): 227–234. Архивировано (PDF) из оригинала 26 марта 2009 г. Проверено 3 мая 2009 г.
- ^ Хольц, Томас Р. (20 сентября 2005 г.). «RE: Конференция по бурпи (ДЛИННЫЙ)» . Проверено 18 июня 2007 г.
- ^ Лоуэн, Массачусетс ; Ирмис, РБ ; Сертич, JJW ; Карри, Пи Джей ; Сэмпсон, SD (2013). Эванс, Дэвид С. (ред.). «Эволюция динозавров-тиранов отслеживает подъем и падение позднемеловых океанов» . ПЛОС ОДИН . 8 (11): е79420. Бибкод : 2013PLoSO...879420L . дои : 10.1371/journal.pone.0079420 . ПМЦ 3819173 . ПМИД 24223179 .
- ^ Эриксон, Грегори М.; Карри Роджерс, Кристи; Варриккио, Дэвид Дж.; Норелл, Марк; Сюй, Син (2007). «Модели роста высиживающих динозавров показывают сроки половой зрелости нептичьих динозавров и происхождение птичьего состояния» . Письма по биологии . 3 (5): 558–561. дои : 10.1098/rsbl.2007.0254 . ПМК 2396186 . ПМИД 17638674 .
- ^ Перейти обратно: а б с Ли, Эндрю Х.; Вернинг, Сара (2008). «Половая зрелость растущих динозавров не соответствует моделям роста рептилий» . ПНАС . 105 (2): 582–587. Бибкод : 2008PNAS..105..582L . дои : 10.1073/pnas.0708903105 . ПМК 2206579 . ПМИД 18195356 .
- ^ Бакли, Лиза Г.; Ларсон, Дерек В.; Райхель, Мириам; Самман, Таня (2010). «Количественная оценка вариаций зубов в пределах одной популяции саркофага альбертозавра (Theropoda: Tyrannosauridae) и значение для идентификации изолированных зубов тираннозаврид». Канадский журнал наук о Земле . 47 (9): 1227–1251. Бибкод : 2010CaJES..47.1227B . дои : 10.1139/e10-029 .
- ^ Перейти обратно: а б с Роуч, Брайан Т.; Бринкман, Дэниел Т. (2007). «Переоценка совместной охоты и стадности Deinonychus antirhopus и других нептичьих динозавров-тероподов». Бюллетень Музея естественной истории Пибоди . 48 (1): 103–138. doi : 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 . S2CID 84175628 .
- ^ Эриксон, Грегори М.; Карри, Филип Дж.; Иноуе, Брайан Д.; Винн, Элис А. (2010). «Пересмотренная таблица смертности и кривая выживаемости саркофага альбертозавра на основе совокупности массовой гибели на Сухом острове». Канадский журнал наук о Земле . 47 (9): 1269–1275. Бибкод : 2010CaJES..47.1269E . дои : 10.1139/e10-051 .
- ^ Эберт, Дэвид А.; МакКри, Ричард Т. (2001). «Были ли крупные тероподы стадными?». Журнал палеонтологии позвоночных . 21 (Приложение к 3 – Тезисы статей, 61-е ежегодное собрание Общества палеонтологии позвоночных): 46A. дои : 10.1080/02724634.2001.10010852 . S2CID 220414868 . (опубликован только аннотация)
- ^ Хорнер, Джон Р. (1997). "Поведение". В Карри, Филип Дж .; Падиан, Кевин (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 45–50. ISBN 978-0-12-226810-6 .
- ^ Максвелл, В. Десмонд; Остром, Джон Х. (1995). «Тафономия и палеобиологические последствия ассоциаций тенонтозавра и дейнониха ». Журнал палеонтологии позвоночных . 15 (4): 707–712. Бибкод : 1995JVPal..15..707M . дои : 10.1080/02724634.1995.10011256 .
- ^ Раат, Майкл А. (1990). «Морфологическая изменчивость мелких теропод и ее значение в систематике: данные Syntarsus rhodesiensis ». В Карпентере, Кеннет ; Карри, Филип Дж. (ред.). Систематика динозавров: подходы и перспективы . Кембридж: Издательство Кембриджского университета. стр. 91–105. ISBN 978-0-521-43810-0 .
- ^ Кориа, Родольфо А .; Карри, Филип Дж. (2006). «Новый кархародонтозаврид (Dinosauria, Theropoda) из верхнего мела Аргентины» (PDF) . Геодиверситас . 28 (1): 71–118. Архивировано из оригинала (PDF) 26 марта 2009 г. Проверено 3 мая 2009 г.
- ^ Карри, Филип Дж .; Трекслер, Дэвид; Коппельхус, Ева Б .; Уикс, Келли; Мерфи, Нейт (2005). «Необычное костное ложе тираннозаврид, состоящее из нескольких особей, в формации Ту Медисин (поздний мел, кампан) в Монтане (США)». В Карпентер, Кеннет (ред.). Плотоядные динозавры . Блумингтон: Издательство Университета Индианы. стр. 313–324. ISBN 978-0-253-34539-4 .
- ^ Коппок, К.; Карри, Пи Джей (2024). «Дополнительный материал саркофага альбертозавра (Tyrannosauridae, Albertosaurinae) из костного пласта Данека в Эдмонтоне, Альберта, Канада, со свидетельствами каннибализма». Канадский журнал наук о Земле . 61 (3): 401–407. doi : 10.1139/cjes-2023-0055 . S2CID 266279287 .
- ^ Ауффенберг, Уолтер (2000). Поведенческая экология комодского варана . Гейнсвилл: Университетское издательство Флориды. ISBN 978-0-8130-0621-5 .
- ^ Карри, Филип Дж.; Эберт, Дэвид А. (2010). «О стадном поведении альбертозавра ». Канадский журнал наук о Земле . 47 (9): 1277–1289. Бибкод : 2010CaJES..47.1277C . дои : 10.1139/e10-072 .
- ^ Перейти обратно: а б Вольф, Юэн Д.С.; Солсбери, Стивен В.; Хорнер, Джон Р.; Варриккио, Дэвид Дж. (2009). Хансен, Деннис Маринус (ред.). «Обычная птичья инфекция поразила динозавров-тиранов» . ПЛОС ОДИН . 4 (9): е7288. Бибкод : 2009PLoSO...4.7288W . дои : 10.1371/journal.pone.0007288 . ПМЦ 2748709 . ПМИД 19789646 .
- ^ Ротшильд, Б., Танке, Д.Х., и Форд, Т.Л., 2001, Стрессовые переломы теропод и разрывы сухожилий как ключ к активности: В: Жизнь мезозойских позвоночных, под редакцией Танке, Д.Х., и Карпентера, К., Издательство Индианского университета. , с. 331–336.
- ^ Молнар, Р.Э., 2001, Палеопатология теропод: обзор литературы: В: Жизнь мезозойских позвоночных, под редакцией Танке, Д.Х., и Карпентера, К., Indiana University Press, стр. 337–363.
- ^ Белл, Фил Р. (2010). «Палеопатологические изменения в популяции саркофага альбертозавра из верхнемеловой формации каньона Подкова в Альберте, Канада». Канадский журнал наук о Земле . 47 (9): 1263–1268. Бибкод : 2010CaJES..47.1263B . дои : 10.1139/e10-030 .
- ^ Перейти обратно: а б с Эберт, Дэвид А. (1997). «Эдмонтон Групп». В Карри, Филип Дж .; Падиан, Кевин (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 199–204. ISBN 978-0-12-226810-6 .
- ^ Ларсон, Дерек В.; Бринкман, Дональд Б.; Белл, Фил Р. (2010). «Наборы фауны из верхней части формации Подкова-Каньон, ранний маастрихтский комплекс с прохладным климатом из Альберты, с особым упором на костное ложе саркофага Альбертозавра ». Канадский журнал наук о Земле . 47 (9): 1159–1181. Бибкод : 2010CaJES..47.1159L . дои : 10.1139/e10-005 .
- ^ Лонгрич, Николас Р.; Карри, Филип Дж. (2009). « Alvarezsauridae , новый альвареззавр (Dinosauria: Theropoda) из раннего маастрихта Альберты, Канада: значение для систематики и экологии Alvarezsauridae». Меловые исследования . 30 (1): 239–252. Бибкод : 2009CrRes..30..239L . дои : 10.1016/j.cretres.2008.07.005 .
Внешние ссылки
[ редактировать ] СМИ, связанные с альбертозавром, на Викискладе?
СМИ, связанные с альбертозавром, на Викискладе?














