Альбертозаврины
| Альбертозаврины Временной диапазон: поздний мел , | |
|---|---|
 | |
| альбертозавра Отливка черепа | |
 | |
| горгозавра Череп | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Суперсемейство: | † Тираннозавроидеа |
| Клэйд : | † Пантираннозаврия |
| Клэйд : | † Эутираннозаврия |
| Семья: | † Тираннозавриды |
| Подсемейство: | † Альбертозаврины Карри и др. , 2003 г. |
| Типовой вид | |
| † Саркофаг Альбертозавра | |
| Роды [1] [2] [3] | |
Альбертозаврины , или динозавры подсемейства Albertosaurinae , жили в позднем мелу США и Канады. Подсемейство было впервые использовано Филипом Дж. Карри , Йорном Х. Хурумом и Каролем Сабатом как группа динозавров -тираннозаврид . Первоначально он был определен как «( Альбертозавр + Горгозавр )», включая только два рода . Группа является сестринской кладой Tyrannosaurinae . В 2007 году было обнаружено, что группа также содержала Малеевозавра , часто называемого синонимом Тарбозавра . Однако эта классификация не была принята, и Малеевозавр до сих пор считается молодым тарбозавром или тираннозавром .
Описание
[ редактировать ]Альбертозаврины — крупные тираннозавриды легкого телосложения . По сравнению с тираннозавринами , они более стройные, имеют более короткие и плоские черепа, более короткие подвздошные кости и пропорционально более длинные голени . Альбертозаврины и тираннозаврины имели руки примерно одинаковой длины, за исключением тарбозавра , у которого для своего размера руки были короткими. [1]

Альбертозавр был меньше некоторых других тираннозаврид, таких как Тарбозавр и Тираннозавр . Типичные взрослые особи альбертозавра и горгозавра имели длину от 8 до 9 м (от 26 до 30 футов). [4] [5] в то время как редкие особи альбертозавра могли вырасти до более 10 м (33 футов) в длину. [6] Несколько независимых оценок массы, полученных разными методами, позволяют предположить, что взрослый альбертозавр весил от 1,3 тонны (1,4 коротких тонны; 1,3 длинных тонны) до 2 тонн (2,2 коротких тонны; 2,0 длинных тонны). [7] [8] [9] [10] Оценки горгозавра выше, около 2,5 тонны (2,8 коротких тонны; 2,5 длинных тонны). [11] [10] хотя существуют более высокие оценки - около 2,8–2,9 тонны (3,1–3,2 коротких тонны; 2,8–2,9 длинных тонны). [12] [9]
Все тираннозавриды, включая альбертозаврин, имели схожий внешний вид тела. Типичные для теропод тираннозавриды были двуногими и уравновешивали тяжелую голову и туловище длинным хвостом. Однако руки тираннозаврид были чрезвычайно маленькими для своего размера тела и сохранили только два пальца. Ноги были длинными и заканчивались четырехпалой ступней. Первый палец, называемый Hallux , был коротким, и только три других касались земли, причем третий (средний) палец был длиннее остальных. [5] Альбертозавр , возможно, мог достигать скорости ходьбы 14–21 километров в час (8–13 миль в час). [13] По крайней мере, для молодых особей высокая скорость бега вполне вероятна. [14]
Классификация и систематика
[ редактировать ]Подсемейство было впервые использовано Филипом Дж. Карри , Йорном Х. Хурумом и Каролем Сабатом как группа динозавров -тираннозаврид . Первоначально он был определен как «( Альбертозавр + Горгозавр )», включая только два рода . Эта группа является сестринской кладой Tyrannosaurinae . [1] В 2007 году было обнаружено, что группа также содержала Малеевозавра , часто называемого синонимом Тарбозавра . [15] Однако эта классификация не была принята, и Малеевозавр до сих пор считается молодым тарбозавром или тираннозавром . [16] Эта классификация не была принята. [2] [3]
Альбертозавр — представитель семейства теропод Tyrannosauridae , подсемейства Albertosaurinae. Его ближайшим родственником является немного более старый Gorgosaurus libratus (иногда называемый Albertosaurus libratus ; см. ниже). [1] Эти два вида — единственные описанные альбертозаврины, но могут существовать и другие неописанные виды. [17] Томас Хольц обнаружил, что аппалачиозавр был альбертозавром в 2004 году. [5] но в его более поздней неопубликованной работе он находится недалеко от тираннозаврид. [18] по согласованию с другими авторами. [19] Другим крупным подсемейством тираннозаврид является Tyrannosaurids , включая такие роды, как Daspletosaurus , Tarbosaurus и Tyrannosaurus . [1] [17]
Отношения горгозавра и альбертозавра
[ редактировать ]Близкое сходство между Gorgosaurus libratus и Albertosaurus sarcophagus на протяжении многих лет побудило многих экспертов объединить их в один род. Альбертозавр был назван первым, поэтому по соглашению ему отдается приоритет перед названием Горгозавр , которое иногда считают его младшим синонимом . Уильям Диллер Мэтью и Барнум Браун усомнились в различии этих двух родов еще в 1922 году. [20] Gorgosaurus libratus был официально переименован в Albertosaurus (как Albertosaurus libratus ) Дейлом Расселом в 1970 году. [4] и многие последующие авторы последовали его примеру. [19] [21] Объединение этих двух видов значительно расширяет географический и хронологический диапазон рода Albertosaurus . Другие эксперты считают эти два рода отдельными. [5] Канадский палеонтолог Фил Карри существует столько же анатомических различий утверждает, что между альбертозавром и горгозавром , сколько между дасплетозавром и тираннозавром , которые почти всегда держатся отдельно. Он также отмечает, что неописанные тираннозавриды, обнаруженные на Аляске , в Нью-Мексико и других местах Северной Америки, могут помочь прояснить ситуацию. [17] Грегори С. Пол предположил, что Gorgosaurus libratus является предком саркофага Альбертозавра . [22]
Филогения
[ редактировать ]Albertosaurinae базальное подсемейство тираннозаврид . — Они были признаны в 2014 году при анализе нового рода Nanuqsaurus , производного тираннозавра . Было обнаружено, что Albertosaurinae включает только Альбертозавра и Горгозавра . [3] альбертозавра и горгозавра . Большинство классификаций разделяют [2] [3] как и должно быть по мнению Карри. [17] Приведенная ниже кладограмма была найдена в ходе анализа Нануксавра Энтони Фиорилло и Рональдом Тыкоски . [3]
Палеобиология
[ редактировать ]Сосуществование горгозавра и дасплетозавра
[ редактировать ]На средних стадиях формирования парка динозавров горгозавр жил рядом с более редким видом тираннозавров, дасплетозавром . Это один из немногих примеров сосуществования двух родов тираннозаврид. Хищники одинакового размера в современных гильдиях хищников разделены на разные экологические ниши из-за анатомических, поведенческих или географических различий, которые ограничивают конкуренцию. Дифференциация ниш между тираннозавридами Парка динозавров недостаточно изучена. [23] В 1970 году Дейл Рассел выдвинул гипотезу, что более распространенный горгозавр активно охотился на быстроногих гадрозавров , в то время как более редкие и доставляющие больше беспокойства цератопсы и анкилозавры (рогатые и тяжелобронированные динозавры ) были оставлены на усмотрение более массивного дасплетозавра . [4] Однако экземпляр дасплетозавра ( OTM 200) из современной формации Ту Медисин в Монтане сохраняет переваренные останки молодого гадрозавра в области кишечника. [24] еще одно костное ложе содержит останки трех дасплетозавров и останки как минимум пяти гадрозавров. [1]
В отличие от некоторых других групп динозавров, ни один из этих родов не был более распространен на возвышенностях или понижениях, чем другой. [23] Однако горгозавр чаще встречается в северных формациях, таких как Парк динозавров, а виды дасплетозавра более распространены на юге. Та же картина наблюдается и у других групп динозавров. Цератопсы-хасмозавры и гадрозаврины-гадрозаврины также более распространены в формации Ту Медисин в Монтане и на юго-западе Северной Америки в кампанском периоде, в то время как центрозаврины и ламбеозаврины доминируют в северных широтах. Хольц предположил, что эта закономерность указывает на общие экологические предпочтения между тираннозавринами , хасмозаврами и гадрозавринами . В конце поздней маастрихтской стадии тираннозаврины , такие как Tyrannosaurus rex , гадрозаврины, такие как Edmontosaurus annectens , и хасмозаврины, как трицератопсы, были широко распространены по всей западной части Северной Америки, в то время как ламбеозаврины были редки, а альбертозаврины и центрозаврины вымерли такие . [5]
Рост
[ редактировать ]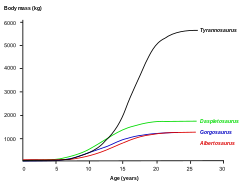
Грегори Эриксон и его коллеги изучили рост и историю жизни тираннозаврид костей с помощью гистологии , которая может определить возраст экземпляра на момент его смерти. Кривую роста можно построить, отобразив на графике возраст различных людей в зависимости от их размеров. Тираннозавриды росли на протяжении всей своей жизни, но в течение примерно четырех лет после продолжительной ювенильной фазы претерпевали огромные скачки роста. Половая зрелость могла закончить эту фазу быстрого роста, после чего рост у взрослых животных значительно замедлился. [7] Изучая пять экземпляров горгозавра различных размеров, Эриксон рассчитал максимальную скорость роста около 50 кг (110 фунтов) в год во время фазы быстрого роста, что медленнее, чем у тираннозавров, таких как дасплетозавр и тираннозавр , но сравнимо с альбертозавром . [7]
Горгозавр провел почти половину своей жизни в ювенильной фазе, прежде чем всего за несколько лет раздулся почти до максимального размера. [7] Это, наряду с полным отсутствием хищников, промежуточных по размеру между огромными взрослыми тираннозавридами и другими мелкими тероподами, позволяет предположить, что эти ниши могли быть заполнены молодыми тираннозавридами. Эта закономерность наблюдается у современных варанов Комодо , чьи детеныши начинаются с древесных насекомоядных животных и постепенно превращаются в массивных высших хищников, способных уничтожать крупных позвоночных животных. [5]
В процессе роста утолщение морфологии зубов изменилось настолько, что, если бы связь молодых и взрослых скелетов на костном ложе Сухого острова не была доказана их принадлежность к одному и тому же таксону, зубы молодых особей, вероятно, были бы идентифицированы статистическим анализом как зубы молодых особей. другой вид. [25]
История жизни
[ редактировать ]Большинству известных особей альбертозавра на момент смерти было 14 лет и больше. [26] Однако самый молодой из известных особей - двухлетний подросток, а самый старший - 28-летний взрослый человек. Двухлетний малыш был обнаружен в костном пласте Сухого острова , весил около 50 кг (110 фунтов) и имел длину около 2 м (6,6 футов). 28-летний экземпляр из того же карьера имел длину 10 м (33 фута). [6] Молодые животные редко встречаются в виде окаменелостей по нескольким причинам, в основном из-за ошибки сохранности , когда более мелкие кости более молодых животных с меньшей вероятностью сохранились в результате окаменения, чем более крупные кости взрослых, а также из-за ошибки коллекционирования, когда меньшие окаменелости с меньшей вероятностью будут замечены. коллекционерами на местах. [26] Молодые альбертозавры относительно крупные для молодых животных, но их останки все еще редки в летописи окаменелостей по сравнению с взрослыми особями. Было высказано предположение, что это явление является следствием истории жизни , а не предвзятостью, и что окаменелости молодых альбертозавров редки, потому что они просто не умирали так часто, как взрослые особи. [6]
Гипотеза истории жизни альбертозавра постулирует, что вылупившиеся птенцы умерли в большом количестве, но не сохранились в летописи окаменелостей из-за их небольшого размера и хрупкой конструкции. Всего через два года молодые особи стали крупнее, чем любой другой хищник в регионе, за исключением взрослого альбертозавра , и имели большую скорость ног, чем большинство их жертв. Это привело к резкому снижению уровня их смертности и соответствующей редкости ископаемых останков. Уровень смертности удвоился в возрасте двенадцати лет, возможно, в результате физиологических потребностей фазы быстрого роста, а затем снова удвоился с наступлением половой зрелости в возрасте от четырнадцати до шестнадцати лет. Этот повышенный уровень смертности продолжался на протяжении всей взрослой жизни, возможно, из-за высоких физиологических потребностей деторождения, включая стресс и травмы, полученные во время внутривидовой конкуренции за партнеров и ресурсы, и, в конечном итоге, постоянно усиливающиеся эффекты старения . Более высокий уровень смертности среди взрослых может объяснить их более частое сохранение. Очень крупные животные были редкостью, потому что немногие особи выживали достаточно долго, чтобы достичь таких размеров. Высокий уровень младенческой смертности, за которым следует снижение смертности среди молоди и внезапное увеличение смертности после половой зрелости, при этом очень немногие животные достигают максимального размера, - это закономерность, наблюдаемая у многих современных крупных млекопитающих, включая слонов. Африканский буйвол и носорог . Та же картина наблюдается и у других тираннозаврид. Сравнение с современными животными и другими тираннозавридами подтверждает эту гипотезу истории жизни , но предвзятость в летописи окаменелостей все еще может играть большую роль, тем более что более двух третей всех экземпляров альбертозавра известны из одного местоположения. [6] [27] Особи альбертозавра и горгозавра растут наиболее быстро в течение четырехлетнего периода, пока им не исполнится примерно 16 лет. [6]
Поведение пакета
[ редактировать ]Костное ложе Сухого острова, обнаруженное Барнумом Брауном и его командой, содержит останки 26 альбертозавров , что является наибольшим количеством особей, найденных в одном местонахождении среди крупных меловых теропод, и вторым по величине среди всех крупных динозавров-тероподов после комплекса аллозавров в Кливленде. Карьер динозавров Ллойда в штате Юта . Группа, судя по всему, состоит из одного очень старого взрослого человека, восьми взрослых в возрасте от 17 до 23 лет, семи суб-взрослых, переживающих фазы быстрого роста в возрасте от 12 до 16 лет, и шести подростков в возрасте от 2 до 11 лет, которые еще не достигла фазы роста. [6]

Почти отсутствие останков травоядных животных и аналогичная степень сохранности, характерная для многих особей в карьере, где находился костяной пласт альбертозавра, привели Карри к выводу, что это место не было ловушкой для хищников, как смоляные ямы Ла Бреа в Калифорнии , и что все сохранившиеся животные умер одновременно. Карри утверждает, что это свидетельство поведения стаи. [14] Другие ученые настроены скептически, отмечая, что животные могли быть согнаны вместе из-за засухи, наводнения или по другим причинам. [6] [26] [29]
Существует множество свидетельств стадного поведения травоядных динозавров, включая цератопсов и гадрозавров . [30] Однако лишь изредка на одном и том же месте встречается такое количество хищников-динозавров. Маленькие тероподы, такие как дейноних , [31] Целофиз и Мегапнозавр ( Syntarsus rhodesiensis ) [32] были обнаружены в скоплениях, как и более крупные хищники, такие как аллозавр и мапузавр . [33] Есть некоторые свидетельства стадного поведения и у других тираннозаврид. Фрагментарные останки более мелких особей были найдены рядом со «Сью», тираннозавром, установленным в Полевом музее естественной истории в Чикаго , а костное ложе в формации Ту Медисин в Монтане содержит по крайней мере три экземпляра дасплетозавра, сохранившиеся вместе с несколькими гадрозаврами. [34] Эти результаты могут подтвердить данные о социальном поведении альбертозавра , хотя некоторые или все из вышеперечисленных мест могут представлять собой временные или неестественные скопления. [1] Другие предполагают, что вместо социальных групп, по крайней мере, некоторые из этих находок представляют собой толпу туш, похожую на драконов Комодо некоторых хищников , где агрессивная конкуренция приводит к гибели и каннибализации . [26] Несмотря на то, что альбертозавры обычно встречаются стаями, нет никаких свидетельств группового поведения у близкого родственника горгозавра . [34] [35]
См. также
[ редактировать ]Ссылки
[ редактировать ]Сноски
[ редактировать ]- ^ Jump up to: а б с д и ж г Карри, Хурум и Сабат, 2003 г.
- ^ Jump up to: а б с Лоуэн и др. 2013 .
- ^ Jump up to: а б с д и Фиорилло и Тыкоски 2014 .
- ^ Jump up to: а б с Рассел 1970 .
- ^ Jump up to: а б с д и ж Хольц 2004 .
- ^ Jump up to: а б с д и ж г Эриксон и др. 2006 год .
- ^ Jump up to: а б с д Эриксон и др. 2004 .
- ^ Кристиансен и Мука 2004 .
- ^ Jump up to: а б Кампионе, Николас Э.; Эванс, Дэвид С.; Браун, Калеб М.; Каррано, Мэтью Т. (2014). «Оценка массы тела у двуногих, не являющихся птицами, с использованием теоретического преобразования в пропорции шилоподий четвероногих» . Методы экологии и эволюции . 5 (9): 913–923. дои : 10.1111/2041-210X.12226 . S2CID 84317234 .
- ^ Jump up to: а б Бенсон, РБЖ; Кампионе, Национальная школа экономики; Каррано, Монтана; Мэннион, Полиция; Салливан, К.; Апчерч, П.; Эванс, округ Колумбия (2014). «Скорость эволюции массы тела динозавров указывает на 170 миллионов лет устойчивых экологических инноваций в линии стволовых птиц» . ПЛОС Биология . 12 (5): e1001853. дои : 10.1371/journal.pbio.1001853 . ПМК 4011683 . ПМИД 24802911 .
- ^ Сибахер 2001 .
- ^ Терриен и Хендерсон 2007 .
- ^ Тулборн 1982 .
- ^ Jump up to: а б Карри 2000 .
- ^ Савельев и Алифанов 2007 .
- ^ Карри 2002 .
- ^ Jump up to: а б с д Карри 2003 .
- ^ Хольц 2005 .
- ^ Jump up to: а б Карр, Уильямсон и Швиммер, 2005 г.
- ^ Мэтью и Браун 1922 .
- ^ Пол 1988 .
- ^ Пол 2010 .
- ^ Jump up to: а б Фарлоу и Пианка 2002 .
- ^ Варриккио 2001 .
- ^ Бакли, Лиза Г.; Ларсон, Дерек В.; Райхель, Мириам; Самман, Таня (2010). «Количественная оценка вариаций зубов в пределах одной популяции саркофага альбертозавра (Theropoda: Tyrannosauridae) и значение для идентификации изолированных зубов тираннозаврид». Канадский журнал наук о Земле . 47 (9): 1227–1251. Бибкод : 2010CaJES..47.1227B . дои : 10.1139/e10-029 .
- ^ Jump up to: а б с д Роуч и Бринкман, 2007 .
- ^ Эриксон и др. 2010 .
- ^ Танке, Даррен Х. (2011). «История событий и мероприятий популярной культуры, связанных с альбертозавром , в Канаде». Бюллетень Палеонтологического общества Альберты . 26 (2): 12–30.
- ^ Эберт, Дэвид А.; и МакКри, Ричард Т. (2001). «Были ли крупные тероподы стадными?». Журнал палеонтологии позвоночных . 21 (Приложение к 3 – Тезисы статей, 61-е ежегодное собрание Общества палеонтологии позвоночных): 46A. дои : 10.1080/02724634.2001.10010852 . S2CID 220414868 . (опубликован только аннотация)
- ^ Хорнер, Джон Р. (1997). "Поведение". В Карри, Филип Дж .; Падиан, Кевин (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 45–50. ISBN 978-0-12-226810-6 .
- ^ Максвелл, В. Десмонд; Остром, Джон Х. (1995). «Тафономия и палеобиологические последствия ассоциаций тенонтозавра и дейнониха ». Журнал палеонтологии позвоночных . 15 (4): 707–712. Бибкод : 1995JVPal..15..707M . дои : 10.1080/02724634.1995.10011256 .
- ^ Раат, Майкл А. (1990). «Морфологическая изменчивость мелких теропод и ее значение в систематике: данные Syntarsus rhodesiensis ». В Карпентере, Кеннет ; Карри, Филип Дж. (ред.). Систематика динозавров: подходы и перспективы . Кембридж: Издательство Кембриджского университета. стр. 91–105. ISBN 978-0-521-43810-0 .
- ^ Кориа, Родольфо А .; И Карри, Филип Дж. (2006). «Новый кархародонтозаврид (Dinosauria, Theropoda) из верхнего мела Аргентины» (PDF) . Геодиверситас . 28 (1): 71–118. Архивировано из оригинала (PDF) 26 марта 2009 года . Проверено 3 мая 2009 г.
- ^ Jump up to: а б Карри и др. 2005 .
- ^ Танке и Карри 1998 .
Библиография
[ редактировать ]- Карр, Томас Д.; Уильямсон, Томас Э.; Швиммер, Дэвид Р. (2005). «Новый род и вид тираннозавроидов из позднемеловой (средний кампан) формации Демополис в Алабаме». Журнал палеонтологии позвоночных . 25 (1): 119–143. doi : 10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2 . ISSN 0272-4634 . S2CID 86243316 .
- Кристиансен, П .; Фаринья, РА (2004). «Массовое предсказание динозавров-теропод». Историческая биология . 16 (2–4): 85–92. Бибкод : 2004HBio...16...85C . дои : 10.1080/08912960412331284313 . S2CID 84322349 .
- Карри, Пи Джей (2000). «Возможные доказательства стадного поведения тираннозаврид» (PDF) . Гайя . 15 : 271–277. Архивировано из оригинала (PDF) 26 марта 2009 г.
- Карри, Пи Джей (2002). «Аллометрический рост тираннозаврид (Dinosauria: Theropoda) верхнего мела Северной Америки и Азии» (аннотация) . Канадский журнал наук о Земле . 40 (4): 651–665. Бибкод : 2003CaJES..40..651C . дои : 10.1139/e02-083 .
- Карри, Пи Джей (2003). «Краниальная анатомия тираннозаврид позднего мела Альберты» (PDF) . Acta Palaeontologica Polonica . 48 (2): 191–226.
- Карри, Пи Джей ; Хурум, Дж. Х. ; Сабат, К. (2003). Хольц, Томас Р. мл. ; Осмольска, Гальшка (ред.). «Строение черепа и эволюция динозавров-тираннозаврид» (PDF) . Acta Palaeontologica Polonica . 48 (2): 227–234.
- Карри, Пи Джей ; Трекслер, Д .; Коппельхус, EB ; Уикс, К .; Мерфи, Н. (2005). «Необычное костное ложе тираннозаврид, состоящее из нескольких особей, в формации Ту Медисин (поздний мел, кампан) в Монтане (США)». В Карпентере, Кеннет Дж. (ред.). Плотоядные динозавры . Блумингтон: Издательство Университета Индианы. стр. 313–324. ISBN 978-0-253-34539-4 .
- Эриксон, генеральный директор ; Маковицкий, Пи Джей ; Карри, Пи Джей ; Норелл, Массачусетс ; Йерби, ЮАР ; Брочу, Калифорния (2004). «Гигантизм и сравнительные параметры жизненного цикла динозавров-тираннозаврид» (PDF) . Природа . 430 (7001): 772–775. Бибкод : 2004Natur.430..772E . дои : 10.1038/nature02699 . ПМИД 15306807 . S2CID 4404887 . Архивировано из оригинала (PDF) 6 октября 2011 г. Проверено 16 апреля 2014 г. (Ошибка: дои : 10.1038/nature16487 , PMID 26675726 , Часы втягивания )
- Эриксон, генеральный директор ; Карри, Пи Джей ; Иноуе, BD ; Винн, А.А. (2006). «Таблицы жизни тираннозавров: пример биологии популяции нептичьих динозавров» (PDF) . Наука . 313 (5784): 213–217. Бибкод : 2006Sci...313..213E . дои : 10.1126/science.1125721 . ПМИД 16840697 . S2CID 34191607 . Архивировано из оригинала (PDF) 18 июля 2010 г.
- Эриксон, генеральный директор ; Карри, Пи Джей ; Иноуе, BD ; Винн, А.А. (2010). «Пересмотренная таблица смертности и кривая выживаемости саркофага альбертозавра на основе совокупности массовой гибели на Сухом острове». Канадский журнал наук о Земле . 47 (9): 1269–1275. Бибкод : 2010CaJES..47.1269E . дои : 10.1139/E10-051 .
- Фарлоу, Джо ; Пианка, скорая помощь (2002). «Перекрытие размеров тела, разделение среды обитания и требования к жизненному пространству наземных позвоночных хищников: последствия для палеоэкологии крупных динозавров-теропод» (PDF) . Историческая биология . 16 (1): 21–40. Бибкод : 2002HBio...16...21F . дои : 10.1080/0891296031000154687 . S2CID 18114585 .
- Фиорилло, Арканзас ; Тыкоски, Р.С. (2014). Додсон, Питер (ред.). «Маленький новый тираннозавр с вершины мира» . ПЛОС ОДИН . 9 (3): e91287. Бибкод : 2014PLoSO...991287F . дои : 10.1371/journal.pone.0091287 . ПМЦ 3951350 . ПМИД 24621577 .
- Хольц, TRJr. (20 сентября 2005 г.). «RE: Конференция по бурпи (ДЛИННЫЙ)» . Список рассылки динозавров . Проверено 18 июня 2007 г.
- Хольц, TRJr. (2004). «Тираннозавроидея» . В Вейшампеле, Дэвид Б .; Додсон, Питер ; Осмольска, Гальшка (ред.). Динозаврия (Второе изд.). Беркли: Издательство Калифорнийского университета. стр. 111–136. ISBN 978-0-520-24209-8 .
- Лоуэн, Массачусетс ; Ирмис, РБ ; Сертич, JJW ; Карри, Пи Джей ; Сэмпсон, SD (2013). Эванс, Дэвид С. (ред.). «Эволюция динозавров-тиранов отслеживает подъем и падение позднемеловых океанов» . ПЛОС ОДИН . 8 (11): е79420. Бибкод : 2013PLoSO...879420L . дои : 10.1371/journal.pone.0079420 . ПМЦ 3819173 . ПМИД 24223179 .
- Мэтью, Вашингтон ; Браун, Б. (1922). «Семейство Deinodontidae с уведомлением о новом роде из позднего мела Альберты» (PDF) . Бюллетень Американского музея естественной истории . 46 (6): 367–385. Архивировано из оригинала (PDF) 16 апреля 2014 г.
- Пол, GS (1988). Хищные динозавры мира Нью-Йорк: Симос и Шустер. стр. 100-1 464 . ISBN 978-0-671-61946-6 .
- Пол, GS (2010). Принстонский полевой справочник по динозаврам . Принстон: Издательство Принстонского университета. п. 105 . ISBN 978-0-691-13720-9 .
- Роуч, Британская Колумбия ; Бринкман, Д.Т. (2007). «Переоценка совместной охоты и стадности Deinonychus antirhopus и других нептичьих динозавров-тероподов». Бюллетень Музея естественной истории Пибоди . 48 (1): 103–138. doi : 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 . S2CID 84175628 .
- Рассел, Д.А. (1970). «Тираннозавры позднего мела Западной Канады» (Обзор) . Публикации Национального музея естественных наук по палеонтологии . 1 (1–6): 1–34. АСИН B0006C6VM8 .
- Савельев С.В. ; Алифанов, В.Р. (2007). Курочкин, Евгений Н. (ред.). «Новое исследование мозга хищного динозавра Tarbosaurus bataar (Theropoda, Tyrannosauridae)» (PDF) . Палеонтологический журнал . 41 (3): 281–289. Бибкод : 2007PalJ...41..281S . дои : 10.1134/S0031030107030070 . ISSN 0031-0301 . S2CID 53529148 .
- Сибахер, Ф. (2001). «Новый метод расчета аллометрических соотношений длины и массы динозавров» (PDF) . Журнал палеонтологии позвоночных . 21 (1): 51–60. CiteSeerX 10.1.1.462.255 . doi : 10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2 . ISSN 0272-4634 . S2CID 53446536 .
- Танке, DH ; Карри, Пи Джей (1998). «Кусание головы динозавров-теропод: палеопатологические данные» (PDF) . Гайя . 15 : 167–184. Архивировано из оригинала (PDF) 27 февраля 2008 г.
- Терриен, Флорида ; Хендерсон, DM (2007). «Мой теропод больше вашего… или нет: оценка размера тела теропод по длине черепа» (PDF) . Журнал палеонтологии позвоночных . 27 (1): 108–115. doi : 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 . ISSN 0272-4634 . S2CID 86025320 .
- Талборн, РА (1982). «Скорость и походка динозавров». Палеогеография, Палеоклиматология, Палеоэкология . 38 (3–4): 227–256. Бибкод : 1982PPP....38..227T . дои : 10.1016/0031-0182(82)90005-0 .
- Варриккио, диджей (2001). «Содержимое кишечника мелового тираннозаврид: значение для пищеварительного тракта тероподных динозавров» (PDF) . Журнал палеонтологии . 75 (2): 401–406. doi : 10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2 . ISSN 0022-3360 . S2CID 86113170 .
Внешние ссылки
[ редактировать ]- «Альбертозаврины» . Fossilworks: вход в базу данных палеобиологии.














