Мегалозавриды
| Мегалозавриды | |
|---|---|
 | |
| Скелет Torvosaurus tanneri , Музей древней жизни. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Клэйд : | † Мегалозаврия |
| Семья: | † Мегалозавриды Хаксли , 1869 г. |
| Типовой вид | |
| † Мегалозавр бакландии Мантель , 1827 г. | |
| Подгруппы | |
| |
| Синонимы | |
| |
Megalosauridae — монофилетическое семейство плотоядных динозавров - теропод в группе Megalosauroidea . Появившись в средней юре , мегалозавриды были среди первых крупных видов крупных тероподных динозавров. [1] Это была относительно примитивная группа базальных столбняков , содержащая два основных подсемейства, Megalosaurinae и Afrovenatorinae, а также базальный род Eustreptospondylus , неопределенный таксон, который отличается от обоих подсемейств. [2]
Определяющий мегалозаврид, Megalosaurus bucklandii , был впервые назван и описан в 1824 году Уильямом Баклендом после многочисленных находок в Стоунсфилде, Оксфордшир , Великобритания. Мегалозавр был первым официально описанным динозавром и послужил основой для создания клады Dinosauria. Это также один из крупнейших известных хищных динозавров средней юры : наиболее хорошо сохранившаяся бедренная кость имеет длину 805 мм и предполагаемую массу тела около 943 кг. [3] Megalosauridae в основном была признана европейской группой динозавров на основании окаменелостей, найденных во Франции и Великобритании, но окаменелости показывают, что эта группа также встречается в Северной Америке, Африке, Южной Америке и, возможно, Азии. [4] [5] [6]
Семейство Megalosauridae было впервые определено Томасом Хаксли в 1869 году, однако на протяжении всей истории оно оспаривалось из-за его роли « корзины для мусора » для многих частично описанных динозавров или неопознанных останков. [7] В первые годы палеонтологии большинство крупных теропод были сгруппированы вместе и до 48 видов включались в кладу Megalosauria , базальную кладу Megalosauridae. Со временем большинство этих таксонов были помещены в другие клады, а параметры Megalosauridae существенно сузились. Тем не менее, остаются некоторые разногласия по поводу того, следует ли считать Megalosauridae отдельной группой, и динозавры этого семейства остаются одними из наиболее проблемных таксонов во всей Dinosauria. [5] [7] Некоторые палеонтологи, такие как Пол Серено в 2005 году, проигнорировали эту группу из-за ее шаткой основы и отсутствия выясненной филогении. Однако недавние исследования Каррано, Бенсона и Сэмпсона систематически проанализировали всех базальных столбняков и определили, что Megalosauridae должны существовать как отдельное семейство. Они, как правило, были тесно связаны с семейством Spinosauridae .
Описание
[ редактировать ]Размер тела
[ редактировать ]
Как и другие столбняки, мегалозавриды — плотоядные тероподы, характеризующиеся крупными размерами и прямохождением . В частности, мегалозавриды обладают особенно гигантскими размерами: некоторые представители семейства весят более одной тонны . Со временем появляются свидетельства увеличения размеров семьи. Базальные мегалозавриды ранней юры имели меньшие размеры тела, чем появившиеся в поздней средней юре. Из-за этого увеличения размера с течением времени Megalosauridae, по-видимому, следуют закономерности увеличения размеров, аналогичной таковой у других теропод гигантского размера, таких как Spinosauridae . [2] Эта закономерность соответствует правилу Коупа — постулату палеонтолога Эдварда Коупа об эволюционном увеличении размеров тела. [5] [8]
Анатомические характеристики
[ редактировать ]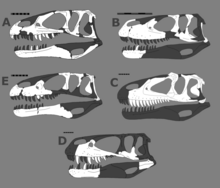
Одной из однозначных синапоморфий Megalosauridae является более низкий и длинный череп с соотношением длины к высоте 3:1. Кроме того, типичная крыша черепа, как правило, гораздо менее украшена, чем у других столбняков, а гребни или рога либо очень малы, либо полностью отсутствуют. У мегалозаврид также есть головки бедренных костей с ориентацией 45 градусов между переднемедиальной и полностью медиальной частью. Megalosauridae также определяются следующими уникальными однозначными синапоморфиями: [1] [2]
- Плечевой дельтопекторальный гребень, заканчивающийся примерно на середине плечевого стержня.
- Отсутствие малоберцового переднелатерального бугорка.
- Ноздри , которые доходят до предчелюстных зубов, однако часть предчелюстной кости перед ноздрями длиннее, чем часть под ними; угловатый кончик рыла (угол между передним и альвеолярным краями <70 градусов).
- Медиальные отверстия на квадратной кости, прилегающие к мыщелкам нижней челюсти.
- Плевроцелозные ямки на крестцовых позвонках .
- Косая связочная борозда на задней поверхности головки бедренной кости неглубокая.
Megalosaurinae (все мегалозавриды, более близкие к Megalosaurus, чем к Afrovenator ) характеризуются умеренным (0,5–2,0) соотношением высоты и длины предчелюстной кости ниже уровня ноздрей по сравнению с другими мегалозавридами, которые имеют более низкое соотношение и, следовательно, менее высокую морду. кончик.
Afrovenatorinae (все мегалозавриды, более близкие к Afrovenator , чем к Megalosaurus) характеризуются квадратным передним краем предглазничной ямки и широко открытой по средней линии лобково-ишиадной пластинкой.
Морфология зубов
[ редактировать ]
Стоматологические данные часто используются для дифференциации различных теропод и для дальнейшей информации кладистической филогении . Морфология зубов и маркеры эволюции зубов склонны к гомоплазии и исчезают или появляются вновь на протяжении всей истории. Однако у мегалозаврид есть несколько специфических состояний зубных протезов, которые отличают их от других базальных теропод. Одним из заболеваний зубов, присутствующих у Megalosauridae, являются множественные морщины эмали возле килей, острого края или зубчатого ряда зуба. Орнаментированные зубы и хорошо выраженная поверхность эмали также характеризуют базальные мегалозавриды. Орнамент и хорошо выраженная поверхность появляются у ранних мегалозавридов, но исчезают у производных мегалозавридов, что позволяет предположить, что это состояние было потеряно с течением времени по мере роста мегалозавридов. [9]
Классификация
[ редактировать ]Историческая классификация
[ редактировать ]
С момента создания семейства многие экземпляры, найденные в полевых условиях, были ошибочно отнесены к мегалозавридам. Например, большинство крупных хищников, обнаруженных примерно через столетие после того, как Megalosaurus bucklandii был назван Megalosaurus bucklandii, были отнесены к Megalosauridae. Мегалозавр был первой палеонтологической находкой такого рода, когда Уильям Бакленд обнаружил гигантскую бедренную кость и назвал ее в 1824 году, еще до появления термина «динозаврия». [1] При первоначальном определении вид M. bucklandii анатомически основывался на различных разобщенных костях, найденных в карьерах вокруг деревни Стоунсфилд, Великобритания. Некоторые из этих ранних находок включали правую зубную кость с хорошо сохранившимся зубом, ребрами , костями таза и крестцовыми позвонками . [10] Поскольку ранние палеонтологи и исследователи обнаружили больше костей динозавров в окрестностях, они отнесли их все к M. bucklandii , поскольку на тот момент в истории это был единственный названный и описанный динозавр. Поэтому вид изначально был описан и классифицирован по массе, возможно, неродственных признаков. [3]
Современная палеонтология впервые начала подходить к проблемному кладистическому разделению мегалозаврид в начале 20 века. Фридрих фон Хюне разделил плотоядных теропод, которые все были сгруппированы в широкую категорию мегалозавридов, на два отдельных семейства: более крупных, более гигантских размеров и меньших, более легкого телосложения теропод. Эти две группы были названы целурозаврией и пахиподозаврией соответственно. Позже Хуэне различил плотоядных и травоядных динозавров в Pachypodosauria, поместив мясоедов в новую группу Carnosauria . [2]
По мере того, как было обнаружено больше информации о базальных тероподах и их филогенетических характеристиках, современные палеонтологи начали сомневаться в правильности названия этой группы. В 2005 году палеонтолог Пол Серено отказался от использования клады Megalosauridae из-за ее неоднозначной ранней истории в пользу названия Torvosauridae. [11] Сегодня принято считать, что мегалозавриды существовали, по крайней мере, как группа базальных столбняков, поскольку у них больше производных таксонов, чем у цератозавров. [2] и что название Megalosauridae должно обозначать эту группу. Megalosauridae также имеет приоритет перед Torvosauridae в соответствии с правилами МКЗН, регулирующими фамилии. [11]
Филогения
[ редактировать ]
Megalosauridae были впервые филогенетически определены в 1869 году Томасом Хаксли, но многие годы использовались как клада «мусорной корзины». В 2002 году Ронан Аллен переопределил кладу после того, как обнаружил на северо-западе Франции полный череп мегалозавра вида Poekilopleuron . Используя персонажей, описанных в этом исследовании, Аллен определил Megalosauridae как динозавров, включая Poekilopleuron valesdunesis , ныне известного как Dubreuillosaurus , Torvosaurus , Afrovenator , и всех потомков их общего предка. Аллен также определил два таксона в пределах Megalosauridae: Torvosaurinae определялся как все Megalosaurinae, более тесно связанные с Torvosaurus, чем с Poekilopleuron и Afrovenator , а Megalosaurinae определялся как все те, которые более тесно связаны с Poekilopleuron . [3] Megalosauridae также попадает в базальную кладу Megalosauroidea, которая также содержит Spinosauridae . Однако многие таксоны все еще весьма нестабильны и не могут быть со стопроцентной уверенностью отнесены к одной кладе. Например, Eustreptospondylus и Streptospondylus , хотя они оба определены как Megalosauridae, часто исключаются для создания более стабильных кладограмм, поскольку они не относятся к определенной подгруппе. [1] [2] Представленная здесь кладограмма соответствует Benson ( 2010) и Benson et al. (2010). [12] [13]
| Мегалозавриды | |

Затем, в 2012 году, Каррано, Бенсон и Сэмпсон провели гораздо более масштабный анализ столбняков и определили Мегалозаврию в более широком смысле как кладу, содержащую Мегалозавра , Спинозавра и всех его потомков. Другими словами, Megalosauria — это группа, состоящая из двух семейств Megalosauridae и его близкого родственника Spinosauridae. В рамках этой новой кладограммы Megalosauridae получили новое подсемейство Afrovenatorinae, которое включало всех мегалозаврид, более тесно связанных с Afrovenator , чем с Megalosaurus .
Каррано, Бенсон и Сэмпсон также включили различные мегалозавриды, которые ранее были исключены из кладограмм в их исследовании 2012 года, такие как Duriavenator и Wiehenvenator у Megalosaurinae и Magnosaurus , Leshansaurus и Piveteausaurus у Afrovenatorinae. [2]
Scuirumimus albersodoerferi , небольшой теропод, описанный в 2012 году и сохранивший протоперья , первоначально считался молодым мегалозавроидом. Это привело к убеждению, что у мегалозавридов могли быть перья. [14] Однако последующие анализы отнесли Sciurumimus к базальному целурозавру . [15] и несколько предполагаемых синапоморфий мегалозавроидов, о которых сообщалось в первоначальном описании, являются общими с базальными целурозаврами. [16]
В 2016 году с помощью филогенетического анализа было обнаружено, что Wiehenvenator Megalosauridae как сестринский таксон Torvosaurus относится к . Ниже представлена кладограмма, основанная на филогенетическом анализе, проведенном Раухутом и др., Показывающая родство Wiehenvenator . [17]
| Мегалозавриды | |
В 2019 году Раухут и Пол описали Asfaltovenator vialidadi , базальный аллозавроид, демонстрирующий мозаику примитивных и производных особенностей, наблюдаемых у Tetanurae . что традиционные Megalosauroidea представляют собой базальную ступень карнозавров Их филогенетический анализ показал , , парафилетическую по отношению к Allosauroidea . Это сделало бы Megalosauridae семейством карнозавров. [18]



Палеоэкология
[ редактировать ]
Было высказано предположение, что мегалозавриды были хищниками или падальщиками, населяющими прибрежную среду. средней юрского периода, Следы предположительно оставленные мегалозавридами, были найдены в долине Мейос в Португалии. В середине юрского периода это место должно было представлять собой приливную равнину, обнаженную во время отлива на краю лагуны. В отличие от большинства прибрежных троп, которые параллельны береговой линии и, вероятно, оставлены мигрирующими животными, тропы долины Мейос были перпендикулярны побережью, причем подавляющее большинство из них было ориентировано в сторону лагуны. Это указывает на то, что мегалозавриды, которые должны были оставить эти следы, приблизились к приливной отмели после отступления прилива. [19]
Это указывает на то, что мегалозавриды могли искать трупы морских существ, оставленных отступающими приливами. Другая возможность состоит в том, что мегалозавриды были рыбоядными и приближались к берегу для охоты за рыбой. Спинозавриды, которые были близкими родственниками мегалозаврид, имели многочисленные приспособления к рыбоядному и полуводному образу жизни, поэтому такой образ жизни подтверждается филогенетическими данными. Зубы акулы, фрагменты хрящей и гастролиты были зарегистрированы как содержимое желудка Poekilopleuron . И этот род, и Dubreuillosaurus были обнаружены в отложениях, также сохранивших корни мангровых зарослей, что является дополнительным доказательством прибрежной среды обитания. Тем не менее, это не исключает возможности того, что мегалозавриды питались и наземной добычей. [20]
Палеогеография
[ редактировать ]
Виды, входящие в состав Megalosauridae, были обнаружены на каждом современном континенте , относительно поровну распределенные между участками на Гондвана и Лавразия суперконтинентах . Результаты палеогеографии показывают, что Megalosauridae в основном обитали в средней и поздней юре , что позволяет предположить, что они вымерли на границе юры и мела 145 миллионов лет назад. [10] Зубы из берриасско - валанжинского возраста формации Бахада-Колорада в Аргентине предполагают, что эта группа, возможно, сохранилась до раннего мела в Южной Америке. [21]
Глобальное распространение этих хищных теропод происходило в два этапа. Во-первых, радиация произошла во время распада Пангеи в ранней юре , около 200 миллионов лет назад. Когда между суперконтинентом возникло море Тетис , мегалозавроиды распространились на две половины Пангеи. Второй этап радиации произошел в средней и поздней юре, от 174 до 145 миллионов лет назад, у аллозавроидов и целурозавров . Megalosauridae, похоже, вымерли в конце этого периода времени. [3]
Останки мегалозаврид на протяжении всей истории находили в различных частях мира. Например, Megalosauridae содержит самый примитивный эмбрион теропода , когда-либо обнаруженный, из раннего титона Португалии, 152 миллиона лет назад (млн лет назад). Кроме того, различные открытия окаменелостей мегалозаврид были датированы байосом - келловейским периодом в Англии и Франции (168–163 млн лет назад), средней юрской Африкой (около 170 млн лет назад), позднеюрским Китаем (163–145 млн лет назад) и титонским периодом Северной Америки (около 150 млн лет назад). [10] Совсем недавно мегалозавриды были обнаружены в формации Тиурарен в Нигере, что еще раз доказывает, что эти базальные столбняки подверглись воздействию глобальной радиации. [5] позднеюрского периода Зубы из формации Такуарембо в Уругвае и формации Тендагуру в Танзании указывают на присутствие крупного мегалозавра, вероятно, Торвозавра. [6]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Бенсон, RBJ (2010). «Описание Megalosaurus bucklandii (Dinosauria: Theropoda) из бата Великобритании и взаимоотношения теропод средней юры» . Зоологический журнал Линнеевского общества . 158 (4): 882–935. дои : 10.1111/j.1096-3642.2009.00569.x .
- ^ Перейти обратно: а б с д и ж г Каррано, Мэтью Т. (2012). «Филогения столбняков (Dinosauria: Theropoda)». Журнал систематической палеонтологии . 10 (2): 211–300. дои : 10.1080/14772019.2011.630927 . S2CID 85354215 .
- ^ Перейти обратно: а б с д Аллен, Ронан (2002). «Открытие мегалозавра (Dinosauria, Theropoda) в среднем бате Нормандии (Франция) и его значение для филогении базальных столбняков». Журнал палеонтологии позвоночных . 22 (3): 548–563. doi : 10.1671/0272-4634(2002)022[0548:DOMDTI]2.0.CO;2 . S2CID 85751613 .
- ^ Рэйфилд, Э.Дж. (2011). «Структурные характеристики черепов столбнячных теропод с упором на Megalosauridae, Spinosauridae и Carcharodontosauridae». Исследования ископаемых четвероногих. Специальные статьи по палеонтологии .
- ^ Перейти обратно: а б с д Серрано-Мартинес, Алехандро (февраль 2015 г.). «Новые остатки теропод из формации Тиурарен (? Средняя юра, Нигер) и их влияние на эволюцию зубов у базальных столбняков». Труды Ассоциации геологов . 126 : 107–118. дои : 10.1016/j.pgeola.2014.10.005 .
- ^ Перейти обратно: а б Сото, Матиас; Ториньо, Пабло; Переа, Дэниел (март 2020 г.). «Крупноразмерный мегалозаврид (Theropoda, Tetanurae) из поздней юры Уругвая и Танзании» . Журнал южноамериканских наук о Земле . 98 : 102458. Бибкод : 2020JSAES..9802458S . дои : 10.1016/j.jsames.2019.102458 . S2CID 213672502 .
- ^ Перейти обратно: а б Бенсон, RBJ (2008). «Таксономический статус Megalosaurus bucklandii (Dinosauria, Theropoda) из средней юры Оксфордшира, Великобритания» . Палеонтология . 51 (2): 419–424. дои : 10.1111/j.1475-4983.2008.00751.x .
- ^ Хоун, Д. (1 января 2005 г.). «Фокус исследования: эволюция больших размеров: как работает правило Коупа?» (PDF) . Тенденции в экологии и эволюции . 20 (1): 4–6. дои : 10.1016/j.tree.2004.10.012 . ПМИД 16701331 .
- ^ Эриксон, Г. (1995). «Расщепление килей на зубах тираннозаврид и последствия их развития». Журнал палеонтологии позвоночных . 15 (2): 268–274. дои : 10.1080/02724634.1995.10011229 .
- ^ Перейти обратно: а б с Хендрикс, К. (2015). «Обзор открытий и классификации нептичьих теропод». Журнал палеонтологии позвоночных Паларха .
- ^ Перейти обратно: а б Серено, ПК (7 ноября 2005 г.). «Стеблевая архозаврия». ТаксонПоиск .
- ^ Бенсон, RBJ (2010). «Описание Megalosaurus bucklandii (Dinosauria: Theropoda) из бата Великобритании и взаимоотношения теропод средней юры» . Зоологический журнал Линнеевского общества . 158 (4): 882–935. дои : 10.1111/j.1096-3642.2009.00569.x .
- ^ Бенсон, РБЖ, Каррано, МТ и Брусатте, СЛ (2010). «Новая клада архаичных крупнотелых хищных динозавров (Theropoda: Allosauroidea), дожившая до позднего мезозоя». Naturwissenschaften . 97 (1): 71–78. Бибкод : 2010NW.....97...71B . дои : 10.1007/s00114-009-0614-x . ПМИД 19826771 . S2CID 22646156 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) Вспомогательная информация - ^ Раухут, Оливер В.М. (2012). «Исключительно сохранившийся молодой мегалозавроидный динозавр-теропод с нитчатым покровом из поздней юры Германии» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (29): 11746–11751. Бибкод : 2012PNAS..10911746R . дои : 10.1073/pnas.1203238109 . ПМК 3406838 . ПМИД 22753486 .
- ^ Годфруа, Паскаль; Кау, Андреа; Донг-Ю, Ху; Эскулье, Франсуа; Вэньхао, Ву; Дайк, Гарет (29 мая 2013 г.). «Юрский птичий динозавр из Китая раскрывает раннюю филогенетическую историю птиц». Природа . 498 (7454): 359–362. Бибкод : 2013Natur.498..359G . дои : 10.1038/nature12168 . ISSN 1476-4687 . ПМИД 23719374 . S2CID 4364892 .
- ^ Хартман, Скотт; Мортимер, Микки; Уол, Уильям Р.; Ломакс, Дин Р.; Липпинкотт, Джессика; Лавлейс, Дэвид М. (10 июля 2019 г.). «Новый паравианский динозавр из поздней юры Северной Америки подтверждает позднее приобретение птичьего полета» . ПерДж . 7 : е7247. дои : 10.7717/peerj.7247 . ISSN 2167-8359 . ПМК 6626525 . ПМИД 31333906 .
- ^ Раухут, Оливер В.М., Хюбнер, Том Р. и Лансер, Клаус-Петер, 2016, «Новый динозавр-теропод-мегалозавриды из поздней средней юры (келловея) северо-западной Германии: последствия для эволюции теропод и круговорота фауны в Юрский период», Palaeontologia Electronica 19.2.26A: 1–65.
- ^ Раухут, Оливер В.М.; Пол, Диего (11 декабря 2019 г.). «Вероятный базальный аллозавроид из ранней среднеюрской формации Каньядон Асфальто в Аргентине подчеркивает филогенетическую неопределенность столбнячных тероподных динозавров» . Научные отчеты . 9 (1): 18826. Бибкод : 2019NatSR...918826R . дои : 10.1038/s41598-019-53672-7 . ISSN 2045-2322 . ПМК 6906444 . ПМИД 31827108 .
- ^ Раззолини, Новелла Л.; Омс, Иволга; Кастанера, Диего; Вила, Бернат; Сантос, Ванда Фариа — два; Галобарт, Ангел (19 августа 2016 г.). «Ихнологические свидетельства пересечения динозаврами-мегалозаврами среднеюрских приливных отмелей» . Научные отчеты . 6 (1): 31494. Бибкод : 2016NatSR...631494R . дои : 10.1038/srep31494 . ISSN 2045-2322 . ПМЦ 4990902 . ПМИД 27538759 .
- ^ Аллен, Ронан (2005). «Посткраниальная анатомия мегалозавра Dubreuillosaurus valesdunensis (Dinosauria Theropoda) из средней юры Нормандии, Франция» (PDF) . Журнал палеонтологии позвоночных . 25 (4): 850–858. doi : 10.1671/0272-4634(2005)025[0850:tpaotm]2.0.co;2 . JSTOR 4524511 . S2CID 130620389 .
- ^ Канале, Хуан И.; Апестегия, С.; Галлина, Пенсильвания; Джанекини, Ф.А.; Халуза, А. (март 2017 г.). «Самые старые тероподы из бассейна Неукен: разнообразие хищных динозавров из формации Бахада-Колорада (нижний мел: берриас-валанжин), Неукен, Аргентина» . Меловые исследования . 71 : 63–78. дои : 10.1016/j.cretres.2016.11.010 . hdl : 11336/109121 .














