Барионикс
| Барионикс Временной диапазон: баррем , | |
|---|---|
 | |
| Реконструированный скелет в Национальном музее природы и науки , Токио. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Спинозавриды |
| Подсемейство: | † Барионихины |
| Род: | † Барионикс Чариг и Милнер , 1986 г. |
| Разновидность: | † Б. Уокери |
| Биномиальное имя | |
| † Барионикс ходери Чариг и Милнер, 1986 г. | |
Барионикс ( / ˌ b ær i ˈ ɒ n ɪ k s / ) — род - теропод динозавров , живших на барремском этапе раннего мелового периода , около 130–125 миллионов лет назад. Первый скелет был обнаружен в 1983 году в глиняной яме Smokejack в графстве Суррей , Англия, в отложениях формации Weald Clay и стал голотипом Baryonyx walkeri , названного палеонтологами Аланом Дж. Чаригом и Анжелой К. Милнер в 1986 году . Родовое название Baryonyx означает «тяжелый коготь» и намекает на очень большой коготь животного на указательном пальце; Видовое название Walkeri - относится к его первооткрывателю, коллекционеру окаменелостей любителю Уильяму Дж. Уокеру. Образец голотипа является одним из наиболее полных скелетов теропод из Великобритании (и остается наиболее полным спинозавридом ), и его открытие привлекло внимание средств массовой информации. Образцы, позже обнаруженные в других частях Соединенного Королевства и Иберии, также были отнесены к этому роду, хотя многие из них с тех пор были переведены в новые роды.
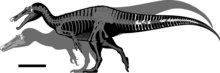
Образец голотипа, который, возможно, не был полностью выращен, по оценкам, имел длину от 7,5 до 10 метров (25 и 33 фута) и весил от 1,2 до 2 метрических тонн (от 1,3 до 2,2 коротких тонн ; от 1,2 до 2,0 длинных) . тонн ). У барионикса была длинная, низкая и узкая морда, которую сравнивали с мордой гавиала . Кончик морды расширен в стороны в форме розетки . Позади верхней челюсти была выемка, которая вписывалась в нижнюю челюсть (которая в той же области загибалась вверх). у него был треугольный гребень На вершине носовых костей . У барионикса было большое количество мелко зазубренных конических зубов, самые большие из которых располагались спереди. Шея приняла S-образную форму, а нервные отростки ее спинных позвонков увеличились в высоту спереди назад. Один удлиненный нервный отросток указывает на то, что у него мог быть горб или гребень в центре спины. У него были крепкие передние конечности с одноименным когтем на первом пальце длиной около 31 сантиметра (12 дюймов).
В настоящее время признанный членом семейства Spinosauridae , барионикса родство было неясным, когда он был обнаружен. Некоторые исследователи предположили, что suchosauruscultridens является старшим синонимом (более старым названием) и чтоuchomimus tenerensis принадлежит к тому же роду; последующие авторы держали их отдельно. Барионикс был первым динозавром-тероподом, который, как было продемонстрировано, был рыбоядным (рыбоядным), о чем свидетельствует рыбья чешуя в области желудка экземпляра-голотипа. Он также мог быть активным хищником более крупной добычи и падальщиком , поскольку в нем также находились кости молодых игуанодонтидов . Существо ловило и перерабатывало добычу в первую очередь с помощью передних конечностей и больших когтей. Барионикс , возможно, вел полуводный образ жизни и сосуществовал с другими динозаврами-тероподами, орнитоподами и зауроподами , а также с птерозаврами , крокодилами , черепахами и рыбами в речной среде.
История открытия
[ редактировать ]
В январе 1983 года сантехник и коллекционер окаменелостей -любителей Уильям Дж. Уокер исследовал глиняную яму Smokejack , глиняную яму в формации Уилд-Клей недалеко от Окли в графстве Суррей, Англия. Он нашел камень, в котором обнаружил большой коготь, но, собрав его дома, обнаружил, что кончик когтя отсутствует. Уокер вернулся на то же место в яме несколько недель спустя и нашел недостающую деталь после часа поисков. Он также нашел кость фаланги и часть ребра . Позже зять Уокера привез коготь в Лондонский музей естественной истории , где его исследовали палеонтологи Алан Дж. Чариг и Анджела К. Милнер , которые идентифицировали его как принадлежавшего тероподу динозавру- . [1] [2] В феврале палеонтологи обнаружили на этом месте еще фрагменты костей, но весь скелет удалось собрать только в мае и июне из-за погодных условий в яме. [3] [2] Команда из восьми сотрудников музея и нескольких волонтеров раскопала 2 метрические тонны (2,2 коротких тонны ; 2,0 длинных тонны ) горной породы в 54 блоках за трехнедельный период. Уокер подарил коготь музею, а компания Ockley Brick (владельцы ямы) передала в дар остальную часть скелета и предоставила оборудование. [3] [4] [2] Эту местность исследовали на протяжении 200 лет, но подобных останков ранее не находили. [5] [2]
Большая часть собранных костей была заключена в алевролитовые конкреции, окруженные мелким песком и илом, а остальные лежали в глине. Кости были разъединены и разбросаны на площади 5 на 2 метра (16,4 на 6,6 фута), но большинство из них находились недалеко от своего естественного положения. Положение некоторых костей было нарушено бульдозером , а некоторые были сломаны механическим оборудованием еще до того, как их собрали. [3] [1] [6] Подготовка образца была затруднена из-за твердости алевролитовой матрицы и присутствия сидерита ; Была предпринята попытка получения кислоты, но большая часть матрицы была удалена механически. Чтобы извлечь все кости из породы, потребовалось шесть лет почти постоянной подготовки, и в конце концов стоматологические инструменты и воздушные молотки пришлось использовать под микроскопом. Образец составляет около 65 процентов скелета и состоит из частичных костей черепа, включая предчелюстные кости (первые кости верхней челюсти); левая верхняя челюсть (вторая кость верхней челюсти); обе носовые кости; левая слезная кость ; левая префронтальная кость ; левый заглазничный ; черепная коробка, включая затылок ; обе зубные кости (передние кости нижней челюсти); различные кости задней части нижней челюсти; зубы; шейные (шея), спинные (спина) и хвостовые (хвост) позвонки ; ребра; грудина ; обе лопатки (лопатки); оба коракоида ; обе плечевые кости (кости плеча); левая лучевая и локтевая кости (кости предплечья); кости пальцев и когти (когтевые кости); бедренные кости; верхний конец левого бедра (бедренная кость) и нижний конец правого; правая малоберцовая кость (голени); и кости стопы, включая ногтевую. [3] [1] [7] [2] Первоначальный номер экземпляра был BMNH R9951, но позже он был переименован в NHMUK PV R9951. [1] [8]



В 1986 году Чариг и Милнер назвали новый род и вид со скелетом в качестве голотипа : Baryonyx walkeri . Родовое название происходит от древнегреческого ; βαρύς ( барыс ) означает «тяжелый» или «сильный», а ὄνυξ ( оникс ) означает «коготь» или «коготь». Видовое название дано в честь Уокера, обнаружившего этот экземпляр. В то время авторы не знали, принадлежал ли большой коготь руке или ноге (как у дромеозавров , которых тогда считали [9] ). Динозавр был представлен ранее в том же году во время лекции на конференции по систематике динозавров в Драмхеллере , Канада. Из-за продолжающейся работы над костями (на тот момент было подготовлено 70 процентов) они назвали свою статью предварительной и пообещали более подробное описание позже. Барионикс был первым крупным тероподом раннего мела, обнаруженным к тому времени где-либо в мире. [1] [6] До открытия барионикса последней значимой находкой теропода в Соединенном Королевстве был Eustreptospondylus в 1871 году, а в интервью 1986 года Чариг назвал барионикс «лучшей находкой века» в Европе. [3] [4] Барионикс широко освещался в международных средствах массовой информации, и журналисты прозвали его «Когти», каламбурив в названии фильма « Челюсти » . Его открытие стало предметом документального фильма BBC 1987 года , а слепок скелета выставлен в Музее естественной истории в Лондоне. В 1997 году Чариг и Милнер опубликовали монографию, подробно описывающую скелет голотипа. [3] [5] [10] Образец голотипа остается наиболее полностью известным скелетом спинозаврид . [11]
Назначенные образцы
[ редактировать ]Окаменелости из других частей Великобритании и Иберии , в основном отдельные зубы, впоследствии были отнесены к бариониксу или подобным животным. [3] отдельные зубы и кости с острова Уайт , в том числе кости рук, о которых сообщалось в 1998 году, и позвонок, о котором сообщили палеонтологи Стив Хатт и Пенни Ньюбери в 2004 году. К этому роду отнесены [12] Фрагмент верхней челюсти из Ла-Риохи, Испания , был приписан бариониксу палеонтологами Луисом И. Виерой и Хосе Анхелем Торресом в 1995 году. [13] (хотя палеонтолог Томас Р. Хольц и его коллеги в 2004 году высказали предположение, что он мог принадлежать Зухомиму ). [14] заглазничных, чешуйчатых костей В 1999 году остатки , зубов, позвонков, пястных костей (кости рук) и фаланг из месторождения Салас-де-лос-Инфантес в провинции Бургос , Испания, были отнесены к незрелому бариониксу (хотя некоторые из этих элементов неизвестны в голотип) палеонтолога Каролины Фуэнтес Видарте и ее коллег. [15] [16] следы динозавров Также было высказано предположение, что возле Бургоса принадлежат бариониксу или подобному тероподу. [17]

В 2011 году был приписан экземпляр (каталогизированный как ML1190 в Museu da Lourinhã ) из формации Папо-Секо в Бока-ду-Чапим , Португалия, с фрагментарной зубной костью, зубами, позвонками, ребрами, тазовыми костями, лопаткой и фаланговой костью. к Бариониксу палеонтологом Октавио Матеусом и его коллегами - наиболее полные иберийские останки животного. Элементы скелета этого экземпляра также представлены в более полном голотипе (который был аналогичного размера), за исключением позвонков средней части шеи. [18] В 2018 году палеонтолог Томас М. С. Арден и его коллеги обнаружили, что португальский скелет не принадлежал бариониксу , поскольку передняя часть его зубной кости не была сильно перевернута. [19] Этот экземпляр был положен в основу нового рода Iberospinus Матеусом и Дарио Эстравис-Лопес в 2022 году. [20] Многочисленные исследования показали, что дополнительные останки спинозаврид из Иберии могут принадлежать к таксонам , отличным от Baryonyx , таким как Vallibonavenatrix и Protathlitis , или могут быть неопределенными. [21] [22] [23] [24] В статье палеонтолога Эрика Исасменди и его коллег, вышедшей в 2024 году, были рассмотрены летописи окаменелостей спинозаврид Иберии и сделан вывод, что ни один образец оттуда не может быть отнесен к бариониксу . Они переместили образец, ранее отнесенный к Baryonyx, из Ла-Риохи в новый род Riojavenatrix . [25]
В 2021 году палеонтолог Крис Т. Баркер и его коллеги описали два новых рода спинозавридов из уэссекской формации острова Уайт, Ceratosurhops и Riparovenator (последний назвал R. milnerae в честь Милнер за ее вклад в исследование спинозаврид) и заявили, что спинозавриды материал оттуда, который ранее приписывался современному бариониксу, мог принадлежать другим таксонам. [26] Эти образцы ранее были отнесены к Бариониксу в тезисах конференции 2017 года. [27] Баркер и его коллеги заявили, что признание образцов формации Уэссекс как новых родов делает присутствие там барионикса неоднозначным, и поэтому большая часть ранее отнесенного изолированного материала из супергруппы Уэлдена является неопределенной. [26] Исследование изолированного зуба, проведенное Баркером и его коллегами в 2023 году, показало, что он и другие зубы из супергруппы Уилдена, которые ранее были отнесены к бариониксу, вероятно, не принадлежат к этому роду, исходя из их морфологии и возраста. [28]
Возможные синонимы
[ редактировать ]

В 2003 году Милнер отметил, что некоторые зубы в Музее естественной истории, ранее идентифицированные как принадлежащие к родам Сухозавр (первый названный спинозаврид) и Мегалозавр, вероятно, принадлежали бариониксу . [7] [29] Типовой вид Сухозавра в , S.cultridens , был назван биологом Ричардом Оуэном в 1841 году на основании зубов, обнаруженных геологом Гидеоном А. Мантеллом Тилгейт -Форест , Сассекс . Первоначально Оуэн думал, что зубы принадлежали крокодилу ; ему еще предстояло назвать группу Dinosauria, что произошло в следующем году. Второй вид, S. girardi , был назван палеонтологом Анри Эмилем Соважем в 1897 году на основе фрагментов челюсти и зуба из Бока-ду-Чапим, Португалия. В 2007 году палеонтолог Эрик Бюффето считал зубы S. girardi очень похожими на зубы Baryonyx (и S.cultridens ), за исключением более сильного развития канавок коронки зуба (или «ребер»; продольных гребней), предполагая, что останки принадлежали к тому же роду. Баффето согласился с Милнером в том, что зубы S.cultridens почти идентичны зубам B.walkeri , но с более ребристой поверхностью. Первый таксон мог быть старшим синонимом второго (поскольку он был опубликован первым), в зависимости от того, были ли различия внутри таксона или между разными таксонами. По мнению Бюффето, поскольку голотип S.cultridens представляет собой одиночный зуб, а B. walkeri — скелет, было бы практичнее сохранить новое название. [30] [31] [32] В 2011 году Матеус и его коллеги согласились, что Сухозавр был тесно связан с бариониксом , но отнесли оба вида к прежнему роду nomina dubia (сомнительные названия), поскольку их голотипические экземпляры не считались диагностическими (отсутствовали отличительные особенности) и не могли быть однозначно отождествлены с другими видами. таксоны. [18] Баркер и его коллеги согласились с этим в 2023 году. [28]

В 1997 году Чариг и Милнер отметили, что две фрагментарные морды спинозаврид из формации Эльраз в Нигере (о которых сообщил палеонтолог Филипп Таке в 1984 году) были настолько похожи на барионикс , что они считали их принадлежащими к неопределенному виду этого рода (несмотря на их большую численность). более молодой аптский геологический возраст). [3] В 1998 году эти окаменелости легли в основу рода и вида Cristatusaurus lapparenti , названного Таке и палеонтологом Дейлом Расселом . [33] Позже в 1998 году палеонтолог Пол Серено и его коллеги назвали новый род и вид suchomimus tenerensis на основе более полных окаменелостей из формации Эльраз. В 2002 году немецкий палеонтолог Ханс-Дитер Сьюс и его коллеги предположили, что Зухомимус tenerensis был достаточно похож на Baryonyx walkeri , чтобы считаться видом в пределах того же рода (как B. tenerensis ), и что Зухомимус был идентичен Cristatusaurus . [34] Милнер согласился, что материал из Нигера неотличим от барионикса в 2003 году. [7] В тезисах конференции 2004 года Хатт и Ньюберри поддержали синонимию, основанную на большом позвонке теропода с острова Уайт, который они приписали животному, тесно связанному с бариониксом и зухомимом . [35] Более поздние исследования сохранили разделение барионикса и зухомима , тогда как кристатузавр было высказано предположение, что либо является nomen dubium , либо, возможно, отличается от обоих. [18] [36] [37] [38] [39] 2017 года В обзорном документе палеонтолога Карлоса Роберто А. Кандейро и его коллег говорится, что эти дебаты носили скорее семантический, чем научный характер, поскольку общепризнано, что B. walkeri и S. tenerensis являются разными, родственными видами. [40] В 2021 году Баркер и его коллеги обнаружили, что Зухомим более связан с британскими родами Riparovenator и Ceratosurhops , чем с Baryonyx . [26]
Описание
[ редактировать ]
По оценкам, барионикс имел длину от 7,5 до 10 м (25 и 33 футов), высоту бедра 2,5 м (8,2 фута) и весил от 1,2 до 2 т (от 1,3 до 2,2 коротких тонн; от 1,2 до 2,0 длинных тонн). ). Тот факт, что элементы черепа и позвоночного столба экземпляра голотипа B. walkeri (NHM R9951), по-видимому, не окостенели (слились), предполагает, что особь не была полностью взрослой, и взрослое животное могло быть намного крупнее. (как и у некоторых других спинозавридов). С другой стороны, сросшаяся грудина экземпляра указывает на то, что он мог быть зрелым. [3] [41] [42] [43]
Череп
[ редактировать ]

Череп барионикса известен не полностью, большая часть средней и задней частей не сохранилась. По оценкам, полная длина черепа составляла 91–95 сантиметров (36–37 дюймов) на основе сравнения с черепом родственного рода suchomimus (который был на 20% больше). [3] [38] Он был удлиненным, а передние 17 см (6,7 дюйма) предчелюстных костей образовывали длинную, узкую и низкую морду ( рострум ) с плавно закругленной верхней поверхностью. [3] ( Внешние ноздри костные ноздри) были длинными, низкими и располагались далеко назад от кончика морды. Передние 13 см (5,1 дюйма) морды расширились до лопатообразной (ложкообразной) «концевой розетки », по форме напоминающей рострум современного гавиала . Передние 7 см (2,8 дюйма) нижнего края предчелюстных костей были загнуты вниз (или загнуты), тогда как передняя часть верхних челюстей была перевернута. Такая морфология привела к образованию сигмовидного или S-образного края нижнего верхнего зубного ряда, в котором зубы передней части верхней челюсти выступали вперед. Морда была особенно узкой непосредственно за розеткой; эта область получила крупные зубы нижней челюсти. Верхняя и предчелюстная кости барионикса образуют сложное сочленение , образующийся зазор между верхней и нижней челюстью известен как субростральная выемка . [3] Опущенная предчелюстная кость и нижний край сигмовидной кишки верхнего зубного ряда также присутствовали у отдаленно родственных теропод, таких как Dilophosaurus . Рыло имело обширные отверстия (отверстия), которые служили выходами кровеносных сосудов и нервов. [3] и на верхней челюсти, по-видимому, имеются пазухи . [3] [38] [42]

Барионикс имел рудиментарное вторичное небо , похожее на крокодилов, но в отличие от большинства тероподных динозавров. [44] Морщинистая (грубо морщинистая) поверхность предполагает наличие роговой подушечки на нёбе. Носовые кости были слиты, что отличало барионикса от других спинозаврид, а сагиттальный гребень над глазами, на верхней средней линии носовых костей, присутствовал . Этот гребень был треугольным, узким и острым в передней части и отличался от гребней других спинозаврид тем, что задние части заканчивались крестообразным отростком. Слезная кость перед глазом, по-видимому, имела роговой стержень, подобный тем, что наблюдались, например, у аллозавра , и отличалась от других спинозавридов тем, что была твердой и почти треугольной. Затылок был узким, парокципитальные отростки были направлены наружу горизонтально, а базиптеригоидные отростки были удлинены и опускались намного ниже базиокципитальной кости (самой нижней кости затылка). [3] [42] [45] что некоторые черепные кости Барионикса Серено и его коллеги предположили , были ошибочно идентифицированы Чаригом и Милнером, в результате чего затылок был реконструирован как слишком глубокий, и что вместо этого череп, вероятно, был таким же низким, длинным и узким, как у Зухомима . [45] Передние 14 см (5,5 дюйма) зубной кости нижней челюсти имели наклон вверх к изгибу морды. Зубная кость была очень длинной и неглубокой, с заметной меккелевой бороздкой на внутренней стороне. Нижнечелюстной симфиз , где две половины нижней челюсти соединялись спереди, был особенно коротким. Остальная часть нижней челюсти была хрупкой; задняя треть была намного тоньше передней и имела вид лезвия. Передняя часть зубной кости выгибалась наружу, чтобы вместить крупные передние зубы, и эта область образовывала нижнечелюстную часть розетки. Зубная кость, как и морда, имела множество отверстий. [3] [38]

Большинство зубов, обнаруженных на образце голотипа, не сочленялись с черепом; некоторые из них остались на верхней челюсти, а на нижней челюсти все еще оставались лишь небольшие сменные зубы. Зубы имели форму загнутых конусов, слегка приплюснутых сбоку, а кривизна их была почти равномерной. Корни были очень длинными и сужались к концам. [3] Кили (острые передний и задний края) зубов были мелко зазубрены с зубчиками спереди и сзади и простирались по всей длине коронки. Зубчиков на мм (0,039 дюйма) было от шести до восьми, что намного больше, чем у крупных теропод, таких как Торвозавр и Тираннозавр . Некоторые зубы были рифлеными, с шестью-восемью гребнями по длине внутренней стороны и мелкозернистой эмалью (самый внешний слой зубов), тогда как другие не имели канавок; их наличие, вероятно, связано с положением или онтогенезом (развитием в процессе роста). [3] [38] Внутренняя сторона каждого зубного ряда имела костную стенку. Количество зубов было большим по сравнению с большинством других теропод: от шести до семи зубов в каждой предчелюстной кости и по тридцать два в каждой зубной кости. Судя по более плотной упаковке и меньшему размеру зубных зубов по сравнению с таковыми в предчелюстной кости соответствующей длины, разница между числом зубов в верхней и нижней челюстях, по-видимому, была более выраженной, чем у других теропод. [3] Концевая розетка верхней челюсти голотипа имела тринадцать зубных альвеол (зубных лунок), шесть слева и семь справа, что свидетельствует об асимметрии количества зубов. Первые четыре верхних зуба были большими (второй и третий самые большие), а четвертый и пятый постепенно уменьшались в размерах. [3] Диаметр самого большого из них был в два раза больше самого маленького. Первые четыре альвеолы зубной кости (соответствующие кончику верхней челюсти) были самыми крупными, а остальные - более правильными по размеру. небольшие субтреугольные межзубные пластинки . Между альвеолами имелись [3] [38]
Посткраниальный скелет
[ редактировать ]

Первоначально считалось, что у него отсутствует сигмовидная кривая, типичная для теропод. [3] шея барионикса , по-видимому, имела S-образную форму, хотя и более прямую, чем у других теропод. [46] Шейные позвонки сужались к голове и постепенно становились длиннее спереди назад. Их зигапофизы (отростки, соединяющие позвонки) были плоскими, а эпипофизы (отростки, к которым прикреплялись мышцы шеи) были хорошо развиты. Аксис развитую (второй шейный позвонок) был небольшим по сравнению с размерами черепа и имел хорошо гипосфену . Нервные дуги шейных позвонков не всегда были пришиты к центрам (телам позвонков), а нервные отростки там были низкими и тонкими. Шейные ребра были короткими, похожими на крокодиловые, и, возможно, несколько перекрывали друг друга. Центры спинных позвонков спины были одинаковыми по размеру. Как и у других теропод, скелет барионикса демонстрировал пневматическость скелета , уменьшая свой вес за счет фенестров (отверстий) в нервных дугах и плевроцелей (полых впадин) в центрах (в первую очередь возле поперечных отростков ). Спереди назад нервные отростки спинных позвонков изменились с коротких и толстых на высокие и широкие. Один изолированный дорсальный нервный отросток был умеренно удлиненным и тонким, что указывает на У барионикса мог быть горб или гребень в центре спины (хотя он начал развиваться по сравнению с таковыми у других спинозавридов). Барионикс был уникальным среди спинозавридов, поскольку имел заметное сужение из стороны в сторону позвонка, который принадлежал либо крестцу , либо передней части хвоста. [3] [6] [45]

Коракоид сужается назад, если смотреть в профиль, и, что уникально для спинозавридов, соединен с лопаткой в виде колышка и выемки. Лопатки были крепкими, а кости передних конечностей короткими по сравнению с размером животного, но широкими и крепкими. Плечевая кость была короткой и толстой, ее концы широко расширены и уплощены: верхняя сторона служила для дельтопекторального гребня и прикрепления мышц, а нижняя — для сочленения с лучевой и локтевой костью. Лучевая кость была короткой, толстой и прямой, менее половины длины плечевой кости, а локтевая кость была немного длиннее. Локтевая кость имела мощный локтевой отросток и расширенный нижний конец. На руках было три пальца; на первом пальце голотипа был большой коготь размером около 31 см (12 дюймов) по изгибу. коготь был бы удлинен за счет кератиновой При жизни (роговой) оболочки. Помимо размеров, пропорции когтя были вполне типичны для теропода: он был двусторонне-симметричным , слегка сжатым, плавно закругленным и остро заостренным. По всей длине когтя проходила канавка для ножен. Остальные когти руки были намного меньше. Подвздошная кость (основная бедренная кость) таза имела выступающий надвертлужный гребень , передний отросток был тонким и вертикально расширенным, а задний отросток был длинным и прямым. Подвздошная кость также имела выступающую короткую полку и глубокую бороздку, обращенную вниз. ( Вертлужная впадина гнездо для бедренной кости) была длинной спереди назад. Седалищная кость хорошо развитый запирательный отросток (нижняя и задняя бедренная кость) имела в верхней части . Край лобковой лопатки на нижнем конце вывернут наружу, лобковая ножка не расширена. У бедренной кости не было бороздки на мыщелке малоберцовой кости, и, что уникально для спинозаврид, малоберцовая кость имела очень неглубокую ямку малоберцовой кости (впадину). [3] [42] [6] [45]
Классификация
[ редактировать ]
В оригинальном описании Чариг и Милнер [1] обнаружили, что барионикс достаточно уникален, чтобы создать новое семейство тероподовых динозавров: Baryonychidae. Они обнаружили, что барионикс не похож ни на одну другую группу теропод, и рассмотрели возможность того, что это был текодонт (группа ранних архозавров, которая теперь считается неестественной ) из-за его очевидно примитивных особенностей. [1] но отметил, что сочленение верхней и предчелюстной костей было сходным с таковым у дилофозавра . Они также отметили, что две морды из Нигера (которые позже стали основой Cristatusaurus ), отнесенные Таке в 1984 году к семейству Spinosauridae, оказались почти идентичными мордам барионикса , и вместо этого они отнесли их к Baryonychidae. [1] В 1988 году палеонтолог Грегори С. Пол согласился с Таке в том, что спинозавр , описанный в 1915 году на основе фрагментарных останков из Египта, уничтоженных во время Второй мировой войны , и барионикс были похожими и (из-за их изогнутых морд), возможно, поздно выжившими дилофозаврами. [43] Баффето также поддержал эти отношения в 1989 году. [47] В 1990 году Чариг и Милнер отвергли родство барионикса со спинозавридами , поскольку не нашли их останки достаточно похожими. [6] В 1997 году они согласились, что Baryonychidae и Spinosauridae родственны, но не согласились с тем, что первое название должно стать синонимом последнего, потому что полнота Baryonyx по сравнению со Spinosaurus сделала его лучшим типовым родом для семейства, и потому что они не нашли сходство между ними достаточно значительное. [3] Хольц и его коллеги включили Baryonychidae в список синонимов Spinosauridae в 2004 году. [14]
Открытия 1990-х годов пролили больше света на взаимоотношения барионикса и его родственников. В 1996 году морда из Марокко была отнесена к Спинозавру , а Ирритатор и Ангатурама из Бразилии (два возможных синонима) были названы. [48] [34] Кристатусавр и Зухомим были названы на основе окаменелостей из Нигера в 1998 году. В своем описании Зухомима Серено и его коллеги поместили его и барионикса в новое подсемейство Baryonychinae внутри Spinosauridae; Спинозавр и Ирритатор были отнесены к подсемейству Spinosaurinae . Baryonychinae отличались небольшими размерами и большим количеством зубов в зубной кости за концевой розеткой, глубоко килеватыми передними спинными позвонками и наличием зазубренных зубов. Spinosaurinae отличались прямыми коронками зубов без зазубрин, маленьким первым зубом в предчелюстной кости, увеличенным расстоянием между зубами в челюстях и, возможно, расположением ноздрей дальше назад и наличием глубокого паруса нервного отростка . [45] [33] Они также объединили спинозавридов и их ближайших родственников в надсемейство Spinosauroidea, но в 2010 году палеонтолог Роджер Бенсон посчитал это младшим синонимом Megalosauroidea (более старое название). [49] В тезисах конференции 2007 года палеонтолог Денвер В. Фаулер предположил, что, поскольку suchosaurus является первым названным родом в своей группе, названия клад Spinosauroidea, Spinosauridae и Baryonychinae должны быть заменены наuchosauroidea, suchosauridae и suchosaurinae, независимо от того, являются ли они название Барионикс сохранено. [29] Исследование, проведенное в 2017 году палеонтологами Маркосом А. Ф. Сейлсом и Сезаром Л. Шульцем, показало, что клада Baryonychinae не получила должной поддержки, поскольку зазубренные зубы могут быть наследственной чертой спинозавридов. [39]


Баркер и его коллеги нашли подтверждение разделения Baryonychinae-Spinosaurinae в 2021 году, и следующая кладограмма показывает положение барионикса внутри Spinosaurinae согласно их исследованию: [26]
| Спинозавриды |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




Эволюция
[ редактировать ]
Спинозавриды, по-видимому, были широко распространены от барремского до сеноманского этапов мелового периода средней , примерно от 130 до 95 миллионов лет назад, в то время как самые старые известные останки спинозаврид относятся к юре . [50] У них были общие черты, такие как длинные и узкие черепа, похожие на крокодиловые; зубы субкруглой формы, с мелкими зубцами или без них; конечная розетка морды; и вторичное небо, которое делало их более устойчивыми к скручиванию. Напротив, примитивным и типичным состоянием теропод была высокая узкая морда с клиновидными (зифодонтными) зубами с зазубренными килями. [51] Адаптации черепа спинозаврид совпадали с черепами крокодилов ; ранние представители последней группы имели черепа, похожие на типичные тероподы, позже у них появились удлиненные морды, конические зубы и вторичное небо. Эта адаптация могла быть результатом перехода от наземной добычи к рыбе. В отличие от крокодилов, посткраниальные скелеты барионихиновых спинозаврид, по-видимому, не имеют водных адаптаций. [52] [51] Серено и его коллеги предположили в 1998 году, что большой коготь большого пальца и крепкие передние конечности спинозаврид развились в средней юре, до удлинения черепа и других адаптаций, связанных с поеданием рыбы, поскольку первые черты являются общими с их родственниками -мегалозавридами . Они также предположили, что спинозаврины и барионихины разошлись до барремского периода раннего мела. [45]
Было предложено несколько теорий о биогеографии спинозавридов. Поскольку Зухомим был более тесно связан с бариониксом (из Европы), чем со спинозавром — хотя этот род также обитал в Африке — распространение спинозаврид не может быть объяснено викариантностью , возникшей в результате континентального рифта . [45] Серено и коллеги [45] предположил, что спинозавриды первоначально были распространены по суперконтиненту Пангея , но раскололись с открытием моря Тетис . Тогда спинозавры развились бы на юге (Африка и Южная Америка: в Гондване ) и барионихины на севере (Европа: в Лавразии ), причем Зухомим стал бы результатом единственного события расселения с севера на юг . [45] Баффето и тунисский палеонтолог Мохамед Уаха также предположили в 2002 году, что барионихины могли быть предками спинозавринов, которые, по-видимому, заменили первых в Африке. [53] В 2003 году Милнер предположил, что спинозавриды возникли в Лавразии во время юрского периода и распространились через Иберийский сухопутный мост в Гондвану, где они распространились . [7] В 2007 году Баффето отметил, что палеогеографические исследования показали, что Иберия находилась недалеко от Северной Африки в раннем меловом периоде, что, по его мнению, подтверждает идею Милнера о том, что Иберийский регион был ступенькой между Европой и Африкой, что подтверждается наличием барионихинов. в Иберии. Направление расселения между Европой и Африкой пока неизвестно. [30] и последующие открытия останков спинозавридов в Азии и, возможно, Австралии указывают на то, что они могли быть сложными. [18]
Кандейро и его коллеги предположили в 2017 году, что спинозавриды северной Гондваны были заменены другими хищниками, такими как абелизавроиды , поскольку нигде в мире не известно никаких определенных окаменелостей спинозавридов после сеномана. Исчезновение спинозавридов и другие сдвиги в фауне Гондваны они объяснили изменениями в окружающей среде, возможно, вызванными нарушениями уровня моря . [40] Малафайя и его коллеги заявили в 2020 году, что барионикс остается самым старым бесспорным спинозавридом, признав при этом, что более старые останки также были предварительно отнесены к этой группе. [23] Баркер и его коллеги обнаружили подтверждение европейского происхождения спинозавридов в 2021 году с их распространением в Азии и Гондване в первой половине раннего мела. В отличие от Серено, эти авторы предположили, что произошло как минимум два события расселения из Европы в Африку, которые привели к Зухомиму и африканской части Spinosaurinae. [26]
Палеобиология
[ редактировать ]
Диета и кормление
[ редактировать ]В 1986 году Чариг и Милнер предположили, что его удлиненная морда со множеством мелко зазубренных зубов указывает на то, что барионикс был рыбоядным (рыбоядным), предполагая, что он приседал на берегу реки и использовал когти, чтобы ловить рыбу из воды (аналогично современному медведь гризли ). [1] Двумя годами ранее Таке [54] отметил, что морды спинозаврид из Нигера были похожи на морды современных гавиалов и предполагали поведение, подобное цапли или аистам . [3] [1] В 1987 году биолог Эндрю Китченер оспорил рыбоядное поведение барионикса и предположил, что он был падальщиком , использовавшим свою длинную шею, чтобы питаться землей, когти, чтобы вгрызаться в тушу, и длинную морду (с далеко задвинутыми назад ноздрями). для дыхания) для исследования полостей тела. [55] Китченер утверждал, что и зубы барионикса челюсти были слишком слабыми, чтобы убивать других динозавров, и слишком тяжелыми, чтобы ловить рыбу, и у них было слишком много приспособлений для рыбоядной деятельности. [55] По мнению палеонтолога Робина Э. Рида, труп хищника мог бы расчленить хищник, а крупные животные, способные на это, например, медведи гризли, также способны ловить рыбу (по крайней мере, на мелководье). [56]


в области желудка В 1997 году Чариг и Милнер продемонстрировали прямые доказательства диетического питания голотипа B. walkeri . Он содержал первые свидетельства рыбоядности динозавра-теропода, протравленные кислотой чешую и зубы обыкновенной рыбы Scheenstia mantelli (тогда отнесенной к роду Lepidotes). [57] ), а также истертые или протравленные кости молодого игуанодонтида . Они также представили косвенные доказательства рыбоядности, такие как крокодиловые приспособления для ловли и глотания добычи: длинные узкие челюсти с «концевой розеткой», похожие на челюсти гавиала, а также опущенный кончик и выемка морды. По их мнению, эти приспособления предполагали, что барионикс ловил рыбу мелкого и среднего размера, как крокодил: хватая ее выемкой на морде (придавая зубам «колющую функцию»), наклоняя голову назад и проглатывая их с головой. [3] Более крупную рыбу разбивают клешнями. То, что зубы нижней челюсти были меньше, скученнее и многочисленнее, чем зубы верхней челюсти, возможно, помогало животному захватывать пищу. Чариг и Милнер утверждали, что барионикс в основном питался рыбой (хотя он также был активным хищником и оппортунистом-падальщиком), но он не был приспособлен для того, чтобы быть макрохищником, как Аллозавр . Они предположили, что барионикс в основном использовал свои передние конечности и большие когти, чтобы ловить, убивать и разрывать более крупную добычу. кажущийся гастролит ( желудочный камень). Вместе с образцом также был обнаружен [3] Немецкий палеонтолог Оливер Уингс в 2007 году предположил, что небольшое количество камней, обнаруженных у теропод, таких как барионикс и аллозавр, могло быть проглочено случайно. [58] В 2004 году Баффетаут и его коллеги обнаружили шейный позвонок птерозавра из Бразилии со встроенным в него зубом спинозавра, который подтвердил, что последние не были исключительно рыбоядными. [59]

проведенное палеонтологом Франсуа Террьеном и его коллегами в 2005 году Исследование лучевой теории, , не смогло реконструировать силовые профили барионикса , но обнаружило, что родственный им Зухомим использовал переднюю часть своих челюстей для захвата добычи, и предположило, что челюсти спинозаврид были адаптированы. для охоты не только на рыбу, но и на более мелкую наземную добычу. Они предполагали, что спинозавриды могли захватывать более мелкую добычу с помощью розетки зубов в передней части челюстей и добивать ее, встряхивая. Вместо этого более крупную добычу пришлось бы поймать и убить передними конечностями, а не укусом, поскольку их черепа не смогли бы противостоять изгибающему напряжению. Они также согласились, что конические зубы спинозаврид были хорошо развиты для пронзения и удержания добычи, а их форма позволяла им выдерживать изгибающие нагрузки со всех направлений. [60] в 2007 году конечных элементов Анализ методом морд , полученных с помощью компьютерной томографии, проведенный палеонтологом Эмили Дж. Рэйфилд и его коллегами показал, что биомеханика барионикса , наиболее похожа на биомеханику гавиала и отличается от биомеханики американского аллигатора и более традиционных теропод, поддерживающих рыбоядную диету для спинозавриды. Вторичное небо помогало им сопротивляться изгибу и скручиванию трубчатой морды. [44] , проведенном палеонтологами Эндрю Р. Каффом и Рэйфилдом в 2013 году В исследовании лучевой теории , сравнили биомеханику морд спинозаврид, полученных с помощью компьютерной томографии, с мордами современных крокодилов и обнаружили, что морды барионикса и спинозавра схожи по их устойчивости к изгибу и скручиванию. Было обнаружено, что барионикс обладает относительно высокой устойчивостью морды к дорсовентральному изгибу по сравнению со спинозавром и гавиалом. Авторы пришли к выводу (в отличие от исследования 2007 года), что барионикс действует иначе, чем гавиал; спинозавриды не были исключительно рыбоядными животными, и их диета определялась их индивидуальным размером. [8]
В тезисах конференции 2014 года палеонтолог Дэнни Андуза и Фаулер отметили, что медведи гризли не ловят рыбу из воды, как это было предложено для барионикса , а также исключили, что динозавр не стал бы выбрасывать голову, как цапли, поскольку шеи спинозаврид не имели сильно S-образной изогнутости, а их глаза не были хорошо расположены для бинокулярного зрения . Вместо этого они предположили, что челюсти совершали боковые движения, чтобы ловить рыбу, как у гавиала, а когти, вероятно, использовались для того, чтобы топтать и пронзать большую рыбу, после чего они манипулировали ею с помощью челюстей, подобно медведям гризли и рыбной ловле. кошки . Они не нашли зубы спинозавридов пригодными для расчленения добычи из-за отсутствия зазубрин и предположили, что они проглатывали бы добычу целиком (при этом отмечая, что они также могли использовать свои когти для расчленения). [61]
Исследование, проведенное в 2016 году палеонтологом Кристофом Хендриксом и его коллегами, показало, что взрослые спинозавры могли смещать свои нижнечелюстные ветви (половинки нижней челюсти) в сторону, когда челюсть была опущена, что позволяло глотку (отверстие, соединяющее рот с пищеводом расширять ). . Это челюстное сочленение похоже на то, которое наблюдается у птерозавров и современных пеликанов , и оно также позволяло спинозавридам глотать крупную добычу, такую как рыба и другие животные. Они также сообщили, что некоторые возможные окаменелости португальского барионикса были обнаружены связанными с изолированными зубами игуанодона , и перечислили это вместе с другими подобными ассоциациями как подтверждение оппортунистического пищевого поведения спинозавров. [38] Другое исследование, проведенное в 2016 году палеонтологом Роменом Вулло и его коллегами, показало, что челюсти спинозавридов сходились с челюстями щуки и морского угря ; у этих рыб также есть челюсти, сжатые из стороны в сторону (тогда как челюсти крокодилов сжаты сверху вниз), удлиненная морда с «концевой розеткой» с увеличенными зубами и выемка за розеткой с меньшими зубами. Такие челюсти, вероятно, развились для хватания добычи в водной среде при слабом освещении и, возможно, помогали в обнаружении добычи. [62]
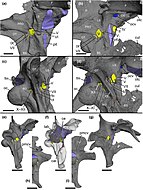

Исследование, проведенное Баркером и его коллегами в 2023 году на основе компьютерной томографии черепных коробок бариониксов и цератозухопов, показало, что анатомия мозга этих барионихинов аналогична анатомии мозга других неманирапториформных теропод . Их нейросенсорные способности, такие как слух и обоняние (обоняние), были обычными, а стабилизация взгляда менее развита, чем у спинозавринов, поэтому их поведенческая адаптация, вероятно, была сопоставима с адаптацией других крупных наземных теропод. Это говорит о том, что их переход от наземных гиперхищников к полуводным «универсалам» в ходе эволюции не потребовал существенной модификации их мозга и сенсорных систем. Это могло означать, что спинозавриды были либо предварительно адаптированы для обнаружения и поимки водной добычи, либо что их переход к полуводному образу жизни потребовал лишь модификации костей, связанных со ртом. подразумевают Значения коэффициента энцефализации рептилий , что когнитивные способности и поведенческая сложность барионихинов не сильно отличались от таковых у других особей. базальные тероподы. [63]
Движение и полуводные привычки
[ редактировать ]В своем первоначальном описании Чариг и Милнер не считали барионикса водным животным (из-за того, что его ноздри расположены по бокам морды — далеко от кончика — и формы посткраниального скелета), но полагали, что он способен плавают, как и большинство наземных позвоночных. [1] Они предположили, что удлиненный череп, длинная шея и сильные плечевые кости барионикса указывают на то, что это животное было факультативным четвероногим , уникальным среди теропод. [1] В своей статье 1997 года они не нашли для этого скелетной опоры, но утверждали, что передние конечности были бы достаточно сильными для четвероногого положения, и он, вероятно, поймал бы водную добычу, присев — или на четвереньках — возле (или в) воде. [3] Повторное описание спинозавра в 2014 году , проведенное палеонтологом Низаром Ибрагимом и его коллегами на основе новых останков, позволило предположить, что это было четвероногое животное, исходя из его переднего центра массы тела . Авторы обнаружили, что четвероногое положение маловероятно для барионикса , поскольку более известные ноги близкого родственника Зухомима не поддерживают эту позу. [52]
В 2017 году палеонтологи Дэвид Э. Хоун и Хольц выдвинули гипотезу, что гребни на голове спинозаврид, вероятно, использовались для демонстрации сексуального характера или угроз. Авторы также отметили, что (как и у других теропод) не было оснований полагать, что передние конечности барионикса были способны пронироваться (пересекая лучевую и локтевую кости предплечья, чтобы поворачивать кисть) и тем самым давать ей возможность отдыхать. или ходить по его ладоням. Возможно, было возможно опираться на передние конечности или использовать их для передвижения (о чем свидетельствуют следы отдыхающего теропода), но если бы это было нормой, передние конечности, вероятно, продемонстрировали бы адаптацию к этому. Кроме того, Хон и Хольц предположили, что передние конечности спинозавридов не кажутся оптимальными для ловли добычи, а вместо этого кажутся похожими на передние конечности роющих животных. Они предположили, что способность копать могла бы пригодиться при раскапывании гнезд, поиске воды или добыче некоторых видов добычи. Хон и Хольц также полагали, что спинозавриды бродили и ныряли в воду, а не погружались в воду из-за скудности их водных адаптаций. [11]


Исследование, проведенное в 2010 году палеонтологом Роменом Амио и его коллегами, предположило, что спинозавриды были полуводными, на основании изотопного состава кислорода зубов спинозаврид со всего мира по сравнению с таковым у других теропод и современных животных. Спинозавриды, вероятно, проводили большую часть дня в воде, как крокодилы и гиппопотамы , и имели диету, аналогичную первым; оба были оппортунистическими хищниками. Поскольку у большинства спинозавридов, по-видимому, нет анатомических приспособлений к водному образу жизни, авторы предположили, что погружение в воду было средством терморегуляции, сходным с таковым у крокодилов и гиппопотамов. Спинозавриды, возможно, также обратились к водной среде обитания и рыбоядному питанию, чтобы избежать конкуренции с крупными наземными тероподами. [64] В 2016 году Сейлс и его коллеги статистически исследовали распределение окаменелостей спинозаврид, абелизаврид и кархародонтозаврид и пришли к выводу, что спинозавриды имеют наибольшую поддержку ассоциации с прибрежной палеосредой. Спинозавриды, по-видимому, также населяли внутренние территории (их распространение там сравнимо с кархародонтозавридами), что указывает на то, что они, возможно, были более универсальными, чем обычно думают. [65]
В 2017 году Сейлс и Шульц согласились, что спинозавриды были полуводными и частично рыбоядными, основываясь на особенностях черепа, таких как конические зубы, сжатая из стороны в сторону морда и втянутые ноздри. Они интерпретировали тот факт, что гистологические данные указывают на то, что некоторые спинозавриды были более наземными, чем другие, как отражение разделения экологических ниш между ними. Поскольку у некоторых спинозавридов ноздри меньше, чем у других, их обонятельные способности, по-видимому, были меньшими, как у современных рыбоядных животных, и вместо этого они могли использовать другие чувства (такие как зрение и механорецепция ) при охоте на рыбу. Обоняние, возможно, было более полезным для спинозавридов, которые также питались наземной добычей, такой как барионихины. [39] Исследование, проведенное палеонтологом Маттео Фаббри и его коллегами в 2022 году, показало, что у барионикса были плотные кости, которые позволили бы ему нырять под воду. Такая же адаптация была выявлена у родственного Spinosaurus , и они, как полагают, были подводными собирателями, нырявшими за водной добычей, в то время как Зухомим был лучше адаптирован к неныряющему образу жизни по сравнению с представленным анализом. Это открытие также демонстрирует уникальный и экологически несопоставимый образ жизни спинозаврид: более виды с полыми костями предпочитают охотиться на мелководье. [66] [67] [68]
Палеосреда
[ редактировать ]
Глинистая формация Уилд состоит из отложений от готерива (нижняя глина Уилд) до барремского (верхняя глина Уилд) возраста , возрастом около 130–125 миллионов лет. Первоначальный образец барионикса был найден в последнем, в глине, представляющей собой неморскую стоячую воду, которая была интерпретирована как речная или илистая среда с мелководьем, лагунами и болотами . [3] В раннем меловом периоде район Уилда в графствах Суррей, Сассекс и Кент был частично покрыт большим с пресной и солоноватой водой озером Уилден . Две большие реки осушали северную территорию (там, где сейчас находится Лондон), впадая в озеро через дельту реки ; Англо-Парижский бассейн находился на юге. Климат здесь был субтропический , похожий на современный средиземноморский регион . Поскольку глиняный карьер Smokejack состоит из разных стратиграфических уровней , обнаруженные там ископаемые таксоны не обязательно относятся к современности. [3] [69] [70] Динозавры из этой местности включают орнитопод Mantellisaurus , Iguanodon и мелких зауроподов . [71] Другие позвоночные животные из глины Уилда включают крокодилов, птерозавров, ящериц (таких как Dorsetisaurus ), амфибий, акул (таких как Hybodus ) и костистых рыб (включая Scheenstia ). [72] [73] Были идентифицированы представители десяти отрядов насекомых, включая Valditermes , Archisphex и Pterinoblattina . Другие беспозвоночные включают остракоды , изоподы , конхостраки и двустворчатые моллюски . [74] [75] Растения Weichselia и водная травянистая Bevhalstia были обычными. Другие найденные растения включают папоротники , хвощи , плауны и хвойные деревья . [76] [77]
Другие динозавры из формации Уэссекс на острове Уайт, где, возможно, обитал барионикс , включают тероподов Riparovenator , Ceratosurhops , Neovenator , Eotyrannus , Aristosurus , Thecocoelurus , Calamospondylus и Ornithodesmus ; орнитоподы игуанодон , гипсилофодон и вальдозавр ; зауроподы Ornithopsis , Eucamerotus и Chondrosteosaurus ; и анкилозавр Полакантус . [78] [26] Баркер и его коллеги заявили в 2021 году, что идентификация двух дополнительных спинозавридов из супергруппы Уилдена, Riparovenator и Ceratosurhops , имеет значение для потенциального экологического разделения внутри Spinosauridae, если бы они и барионикс были современниками и взаимодействовали. Они предупредили, что вполне возможно, что формации Аппер-Уилд-Клей и Уэссекс, а также известные из них спинозавриды были разделены во времени и на расстоянии. [26]
Обычно считается, что крупные хищники встречаются с небольшим таксономическим разнообразием в любой области из-за экологических требований, однако многие мезозойские комплексы включают двух или более симпатрических теропод, сопоставимых по размеру и морфологии, и это, по-видимому, также имело место для спинозавридов. Баркер и его коллеги предположили, что высокое разнообразие спинозавридов в данной области могло быть результатом экологических обстоятельств, приносящих пользу их нише. Хотя обычно считалось, что только идентифицируемые анатомические особенности, связанные с разделением ресурсов, допускают сосуществование крупных теропод, Баркер и его коллеги отметили, что это не исключает того, что сходные и близкородственные таксоны могут сосуществовать и перекрываться в экологических требованиях. Возможное разделение ниш может быть во времени (сезонное или ежедневное), в пространстве (между местами обитания в одних и тех же экосистемах) или в зависимости от условий, а также они могут быть разделены по выбору среды обитания в пределах своих регионов (которые могут варьироваться в зависимости от условий). климат). [26]
Тафономия
[ редактировать ]
Чариг и Милнер представили возможный сценарий, объясняющий тафономию (изменения во время распада и окаменения) образца голотипа B. walkeri . [3] Мелкозернистые отложения вокруг скелета, а также тот факт, что кости были найдены близко друг к другу (элементы черепа и передних конечностей на одном конце раскопа, а элементы таза и задних конечностей на другом), указывают на то, что окружающая среда была спокойной. в момент захоронения, а водные течения не унесли тушу далеко — возможно, потому, что вода была мелкой. Место, где погиб экземпляр, похоже, было подходящим для рыбоядного животного. Возможно, он ловил рыбу и собирал мусор на илистой равнине, завязнув в болоте, прежде чем умер и был похоронен. Поскольку кости хорошо сохранились и не имели следов погрызов, падальщики, судя по всему, не трогали труп (что позволяет предположить, что он быстро был покрыт осадком). [3]
Разъединение костей могло быть результатом разложения мягких тканей. Части скелета, похоже, подверглись выветриванию в разной степени, возможно, из-за изменения уровня воды или смещения отложений (обнажая части скелета). Кости пояса и конечностей, зубная кость и ребро были сломаны до окаменения, возможно, из-за того, что их затоптали крупные животные во время захоронения. Большая часть хвоста, по-видимому, была потеряна до окаменения, возможно, из-за сбора мусора или сгнила и уплыла. Ориентация костей указывает на то, что туша лежала на спине (возможно, слегка наклоненной влево, правой стороной вверх), что может объяснить, почему все нижние зубы выпали из лунок, а некоторые верхние зубы остались на месте. . [3] [2]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л Чариг, Эй Джей; Милнер, AC (1986). « Барионикс , новый замечательный динозавр-теропод». Природа . 324 (6095): 359–361. Бибкод : 1986Natur.324..359C . дои : 10.1038/324359a0 . ПМИД 3785404 . S2CID 4343514 .
- ^ Перейти обратно: а б с д и ж Психойос, Л.; Ноббер, Дж. (1994). Охота на динозавров . Лондон: Касселл. стр. 176–179 . ISBN 978-0679431244 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах Чариг, Эй Джей; Милнер, AC (1997). « Baryonyx walkeri , рыбоядный динозавр из Уэлдена в графстве Суррей» . Бюллетень Лондонского музея естественной истории . 53 : 11–70.
- ^ Перейти обратно: а б Эдвардс, Д.Д. (1986). «Ископаемый коготь открывает новое генеалогическое древо». Новости науки . 130 (23): 356. дои : 10.2307/3970849 . JSTOR 3970849 .
- ^ Перейти обратно: а б Муди, RTJ; Нэйш, Д. (2010). «Алан Джек Чариг (1927–1997): обзор его научных достижений и роли в мире исследований ископаемых рептилий». Геологическое общество, Лондон, специальные публикации . 343 (1): 89–109. Бибкод : 2010ГСЛСП.343...89М . дои : 10.1144/SP343.6 . S2CID 129586311 .
- ^ Перейти обратно: а б с д и Чариг, Эй Джей; Милнер, AC (1990). «Систематическое положение Baryonyx walkeri в свете реклассификации Теропод Готье» . В Карпентере, К.; Карри, Пи Джей (ред.). Систематика динозавров: перспективы и подходы . Кембридж: Издательство Кембриджского университета. стр. 127–140. дои : 10.1017/CBO9780511608377.012 . ISBN 978-0-521-43810-0 .
- ^ Перейти обратно: а б с д Милнер, AC (2003). «Рыбоядные тероподы: краткий обзор систематики, биологии и палеобиогеографии спинозавров». Actas de las II Jornadas Internacionales Sobre Paleontologýa de Dinosaurios y Su Entorno : 129–138.
- ^ Перейти обратно: а б Манжета, Арканзас; Рэйфилд, Э.Дж. (2013). Фарке, Эндрю А. (ред.). «Механика питания спинозавровых теропод и современных крокодилов» . ПЛОС ОДИН . 8 (5): e65295. Бибкод : 2013PLoSO...865295C . дои : 10.1371/journal.pone.0065295 . ПМЦ 3665537 . ПМИД 23724135 .
- ^ Норман, Д.Б. (1985). «Дромеозавриды». Иллюстрированная энциклопедия динозавров: оригинальный и убедительный взгляд на жизнь в царстве динозавров . Нью-Йорк: Crescent Books. стр. 57–58. ISBN 978-0-517-46890-6 .
- ^ Остерлов, Э. (2018). «Развенчание мифов о динозаврах и заблуждений о кино» . www.nhm.ac.uk. Проверено 20 октября 2018 г.
- ^ Перейти обратно: а б Хон, DWE; Хольц, ТР (2017). «Век спинозавров – обзор и пересмотр спинозавридов с комментариями по их экологии» . Acta Geologica Sinica — английское издание . 91 (3): 1120–1132. Бибкод : 2017AcGlS..91.1120H . дои : 10.1111/1755-6724.13328 . S2CID 90952478 .
- ^ Клэбби, С.М. (2005). « Барионикс Чариг и Милнер 1986» . ДиноВайт . Проверено 12 октября 2015 г.
- ^ Виера, Л.И.; Торрес, Дж. А. (1995). «Присутствие Baryonyx walkeri (Saurischia, Theropoda) в пустоши Ла-Риоха (Испания)» . Munibe Ciencias Naturales (на испанском языке). 47 : 57–61. ISSN 0214-7688 .
- ^ Перейти обратно: а б Хольц, ТР; Мольнар, Р.Э.; Карри, Пи Джей (2004). «Базальная столбняк». В Вейшампеле, Д.Б.; Додсон, П.; Осмольская, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 71–110. ISBN 978-0-520-24209-8 .
- ^ Видарте, CF; Кальво, ММ; Мейджиде, М.; Слева, Лос-Анджелес; Монтеро, Д.; Перес, Г.; Торсида, Ф.; Уриен, В.; Фуэнтес, FM; Фуэнтес, ММ (2001). «Ископаемые останки барионикса (Dinosauria, Theropoda) в раннем мелу Салас-де-лос-Инфантес (Бургос, Испания)». Материалы Первой международной конференции по палеонтологии динозавров и их окружающей среды. Салас-де-лос-Инфантес, Бургос (на испанском языке): 349–359.
- ^ Переда-Субербиола, X.; Руис-Оменьака, Дж.И.; Канудо, Дж.И.; Торсида, Ф.; Санс, Дж. Л. (2012). «Фауна динозавров раннего мела (валанжин-альб) Испании» . В Годфруа, П. (ред.). Берниссар Динозавры . Издательство Университета Индианы. стр. 389–390. ISBN 978-0-253-00570-0 .
- ^ Перес-Лоренте, Ф. (2015). Следы динозавров и тропы Ла- Риохи Жизнь прошлого. Индиана: Издательство Университета Индианы. п. 325. ИСБН 978-0-253-01515-0 .
- ^ Перейти обратно: а б с д Мэтью, О.; Араужо, Р.; Натарио, К.; Кастанинья, Р. (2011). «Новый экземпляр тероподного динозавра барионикса из раннего мела Португалии и таксономическая достоверность Сухозавра » (PDF) . Зоотакса . 2827. 2827 : 54–68. дои : 10.11646/zootaxa.2827.1.3 .
- ^ Арден, ТМС; Кляйн, КГ; Зухри, С.; Лонгрич, Северная Каролина (2018). «Водная адаптация черепа хищных динозавров (Theropoda: Spinosauridae) и эволюция водных привычек спинозавра». Меловые исследования . 93 : 275–284. Бибкод : 2019CrRes..93..275A . дои : 10.1016/j.cretres.2018.06.013 . S2CID 134735938 .
- ^ Матеус, Октавиус; Эстравис-Лопес, Дарио (2022). «Новый динозавр-теропод из раннего мела (баррема) мыса Эспишель, Португалия: значение для эволюции спинозаврид» . ПЛОС ОДИН . 17 (2): e0262614. Бибкод : 2022PLoSO..1762614M . дои : 10.1371/journal.pone.0262614 . ISSN 1932-6203 . ПМЦ 8849621 . ПМИД 35171930 .
- ^ Малафайя, Э.; Гасулла, Дж. М.; Эскасо, Ф.; Нарваес, И.; Санс, Дж.Л.; Ортега, Ф. (2018). «Новые спинозавриды (Theropoda, Megalosauroidea) остаются из формации Арсиллас-де-Морелла (верхний баррем) в Морелле, Испания». Меловые исследования . 92 : 174–183. Бибкод : 2018CrRes..92..174M . дои : 10.1016/j.cretres.2018.08.006 . S2CID 134572693 .
- ^ Перейти обратно: а б Малафайя, Э.; Гасулла, Дж. М.; Эскасо, Ф.; Нарваес, И.; Ортега, Ф. (2020). «Обновление летописи окаменелостей спинозаврид (Dinosauria: Theropoda) из нижнего мела Пиренейского полуострова: распространение, разнообразие и история эволюции». Журнал иберийской геологии . 46 (4): 431–444. Бибкод : 2020JIbG...46..431M . дои : 10.1007/s41513-020-00138-9 . S2CID 222149842 .
- ^ Сантос-Кубедо, Андрес; де Сантистебан, Шарль; Поса, Бегонья; Месегер, Сержи (2023). «Новый вид динозавров-спинозаврид из раннего мела Синкторреса (Испания)» . Научные отчеты . 13 (1): 6471. Бибкод : 2023NatSR..13.6471S . дои : 10.1038/s41598-023-33418-2 . ПМЦ 10195869 . ПМИД 37202441 .
- ^ Исасменди, Эрик (2024). «Увеличение количества теропод в Европе: новый базальный спинозаврид из группы Энсизо бассейна Камероса (Ла-Риоха, Испания). Эволюционные последствия и палеобиоразнообразие». Зоологический журнал Линнеевского общества . XX : 34. doi : 10.1093/zoolinnean/zlad193 .
- ^ Перейти обратно: а б с д и ж г час Баркер, Коннектикут; Хон, DWE; Нэйш, Д.; Кау, А.; Локвуд, JAF; Фостер, Б.; Кларкин, CE; Шнайдер, П.; Гостлинг, Нью-Джерси (2021 г.). «Новые спинозавриды из формации Уэссекс (ранний мел, Великобритания) и европейское происхождение спинозавридов» . Научные отчеты . 11 (1): 19340. Бибкод : 2021NatSR..1119340B . дои : 10.1038/s41598-021-97870-8 . ПМЦ 8481559 . ПМИД 34588472 .
- ^ Мунт, MC; Блэквелл, Г.; Кларк, Дж.; Фостер, Б. (2017). «Новые находки динозавров-спинозавридов из формации Уэссекс (группа Уэлден, ранний мел) острова Уайт». СВПКА . 65 :1. дои : 10.13140/RG.2.2.17925.86242 .
- ^ Перейти обратно: а б Баркер, Крис Т.; Нэйш, Даррен; Гостлинг, Нил Дж. (2023). «Изолированный зуб раскрывает скрытое разнообразие динозавров-спинозаврид в британской супергруппе Уилдена (нижний мел)» . ПерДж . 11 : е15453. дои : 10.7717/peerj.15453 . ПМЦ 10239232 . ПМИД 37273543 .
- ^ Перейти обратно: а б Фаулер, Д.В. (2007). «Недавно открытые барионихиновые зубы (Dinosauria: Theropoda): новые морфологические данные, расширение ареала и сходство с цератозавром » . Журнал палеонтологии позвоночных . 27 (3): 3.
- ^ Перейти обратно: а б Баффето, Э. (2007). «Спинозавридный динозавр барионикс (Saurischia, Theropoda) в раннем мелу Португалии» (PDF) . Геологический журнал . 144 (6): 1021–1025. Бибкод : 2007GeoM..144.1021B . дои : 10.1017/S0016756807003883 . S2CID 130212901 .
- ^ Хендрикс, К.; Матеус, О.; Араужо, Р. (2015). «Предлагаемая терминология зубов теропод (Dinosauria, Saurischia)» . Журнал палеонтологии позвоночных . 35 (5): e982797. Бибкод : 2015JVPal..35E2797H . дои : 10.1080/02724634.2015.982797 . S2CID 85774247 .
- ^ Хендрикс, К. (2008). «Spinosauridae - Historique des découvertes» . spinosauridae.fr.gd (на французском языке) . Проверено 22 октября 2018 г.
- ^ Перейти обратно: а б Таке, П.; Рассел, Д.А. (1998). «Новые данные о динозаврах-спинозавридах раннего мела Сахары». Comptes Rendus de l'Académie des Sciences, Série IIA . 327 (5): 347–353. Бибкод : 1998CRASE.327..347T . дои : 10.1016/S1251-8050(98)80054-2 .
- ^ Перейти обратно: а б Сьюс, HD; Фрей, Э.; Мартилль, М.; Скотт, DM (2002). « Irritator Challengeri , спинозаврид (Dinosauria: Theropoda) из нижнего мела Бразилии». Журнал палеонтологии позвоночных . 22 (3): 535–547. doi : 10.1671/0272-4634(2002)022[0535:icasdt]2.0.co;2 . S2CID 131050889 .
- ^ Хатт, С.; Ньюбери, П. (2004). «Новый взгляд на Baryonyx walkeri (Чариг и Милнер, 1986) на основе недавней находки окаменелостей в Уилдене» . Симпозиум по палеонтологии и сравнительной анатомии позвоночных . Архивировано из оригинала 5 октября 2015 года.
- ^ Аллен, Р.; Ксайсанавонг, Т.; Ричир, П.; Хентавонг, Б. (2012). «Первый окончательный азиатский спинозаврид (Dinosauria: Theropoda) из раннего мела Лаоса». Naturwissenschaften . 99 (5): 369–377. Бибкод : 2012NW.....99..369A . дои : 10.1007/s00114-012-0911-7 . ПМИД 22528021 . S2CID 2647367 .
- ^ Бенсон, РБЖ; Каррано, Монтана; Брусатте, СЛ (2009). «Новая клада архаичных крупнотелых хищных динозавров (Theropoda: Allosauroidea), дожившая до позднего мезозоя» (PDF) . Naturwissenschaften (Представленная рукопись). 97 (1): 71–78. Бибкод : 2010NW.....97...71B . дои : 10.1007/s00114-009-0614-x . ПМИД 19826771 . S2CID 22646156 .
- ^ Перейти обратно: а б с д и ж г Хендрикс, К.; Матеус, О.; Бюффето, Э.; Эванс, Арканзас (2016). «Морфофункциональный анализ квадрата спинозаврид (Dinosauria: Theropoda) и наличие спинозавра и второго таксона спинозавров в сеномане Северной Африки» . ПЛОС ОДИН . 11 (1): e0144695. Бибкод : 2016PLoSO..1144695H . дои : 10.1371/journal.pone.0144695 . ПМК 4703214 . ПМИД 26734729 .
- ^ Перейти обратно: а б с Продажи, МАФ; Шульц, CL (2017). «Таксономия спинозавров и эволюция краниодентальных особенностей: данные из Бразилии» . ПЛОС ОДИН . 12 (11): e0187070. Бибкод : 2017PLoSO..1287070S . дои : 10.1371/journal.pone.0187070 . ПМЦ 5673194 . ПМИД 29107966 .
- ^ Перейти обратно: а б Кандейро, CRA; Брусатте, СЛ; Соуза, Алабама (2017). «Раннемеловые динозавры-спинозавры Северной Африки и Европы: летопись окаменелостей, биогеография и вымирание» . Ежегодник Института геологических наук . 40 (3): 294–302. дои : 10.11137/2017_3_294_302 . hdl : 20.500.11820/1bc143b5-5bd5-416b-bf10-dd28304d2c9b .
- ^ Терриен, Ф.; Хендерсон, DM (2007). «Мой теропод больше вашего… или нет: оцениваем размер тела теропод по длине черепа». Журнал палеонтологии позвоночных . 27 (1): 108–115. doi : 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 . S2CID 86025320 .
- ^ Перейти обратно: а б с д Пол, GS (2010). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. стр. 87–88 . ISBN 978-0-691-13720-9 .
- ^ Перейти обратно: а б Пол, GS (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. стр. 271–274 . ISBN 978-0-671-61946-6 .
- ^ Перейти обратно: а б Рэйфилд, Э.Дж.; Милнер, AC; Сюань, В.Б.; Янг, П.Г. (2007). «Функциональная морфология динозавров-крокодилоподобных спинозавров». Журнал палеонтологии позвоночных . 27 (4): 892–901. doi : 10.1671/0272-4634(2007)27[892:FMOSCD]2.0.CO;2 . S2CID 85854809 .
- ^ Перейти обратно: а б с д и ж г час я Серено, ПК; Бек, Алабама; Дютейл, Д.Б.; Гадо, Б.; Ларссон, HCE; Лион, GH; Маркот, доктор юридических наук; Раухут, OWM; Садлер, RW; Сидор, Калифорния; Варриккио, Д.Д.; Уилсон, врач общей практики; Уилсон, Дж. А. (1998). «Длиннорылый хищный динозавр из Африки и эволюция спинозаврид». Наука . 282 (5392): 1298–1302. Бибкод : 1998Sci...282.1298S . дои : 10.1126/science.282.5392.1298 . ПМИД 9812890 .
- ^ Эверс, Юго-Запад; Раухут, OWM; Милнер, AC; Макфитерс, Б.; Аллен, Р. (2015). «Переоценка морфологии и систематического положения тероподного динозавра Sigilmassasaurus из «среднего» мела Марокко» . ПерДж . 3 : е1323. дои : 10.7717/peerj.1323 . ПМЦ 4614847 . ПМИД 26500829 .
- ^ Баффето, Э. (1989). «Новые останки загадочного динозавра спинозавра из мелового периода Марокко и сходство между спинозавром и бариониксом ». Новый ежегодник по геологии и палеонтологии - ежемесячные выпуски . 1989 (2): 79–87. дои : 10.1127/njgpm/1989/1989/79 .
- ^ Рассел, Д.А. (1996). «Изолированные кости динозавров из среднего мела Тафилальта, Марокко». Бюллетень Национального музея естественной истории, Париж, раздел C. 4е серия. 18 (2–3): 349–402.
- ^ Бенсон, RBJ (2010). «Описание Megalosaurus bucklandii (Dinosauria: Theropoda) из бата Великобритании и взаимоотношения теропод средней юры» . Зоологический журнал Линнеевского общества . 158 (4): 882–935. дои : 10.1111/j.1096-3642.2009.00569.x . S2CID 84266680 .
- ^ Серрано-Мартинес, А.; Видаль, Д.; Шишио, Л.; Ортега, Ф.; Нолл, Ф. (2015). «Изолированные зубы теропод из средней юры Нигера и ранняя стоматологическая эволюция спинозавридов». Acta Palaeontologica Polonica . дои : 10.4202/app.00101.2014 . hdl : 10261/152148 . S2CID 53331040 .
- ^ Перейти обратно: а б Хольц-младший, TR (1998). «Спинозавры как подражатели крокодилам». Наука . 282 (5392): 1276–1277. дои : 10.1126/science.282.5392.1276 . S2CID 16701711 .
- ^ Перейти обратно: а б Ибрагим, Н. ; Серено, ПК; Даль Сассо, К.; Магануко, С.; Фабри, М.; Мартилл, DM; Зухри, С.; Мирвольд, Н.; Лурино, Д.А. (2014). «Полуводные адаптации гигантского хищного динозавра» . Наука . 345 (6204): 1613–1616. Бибкод : 2014Sci...345.1613I . дои : 10.1126/science.1258750 . ПМИД 25213375 . S2CID 34421257 . Дополнительная информация
- ^ Бюффето, Э.; Уаха, М. (2002). «Новый экземпляр спинозавра (Dinosauria, Theropoda) из нижнего мела Туниса с замечаниями по эволюционной истории спинозавридов» (PDF) . Бюллетень геологического общества Франции . 173 (5): 415–421. дои : 10.2113/173.5.415 . HDL : 2042/216 . S2CID 53519187 .
- ^ Таке, П. (1984). «Любопытная специализация черепа некоторых хищных динозавров мелового периода: длинная и узкая морда спинозавридов». Comptes Rendus de l’Académie des Sciences (на французском языке). 299 : 217–222.
- ^ Перейти обратно: а б Китченер, А. (1987). «Функция когтей когтей». Природа . 325 (6100): 114. Бибкод : 1987Natur.325..114K . дои : 10.1038/325114a0 . S2CID 4264665 .
- ^ Рид, REH (1987). «Когти Когти» . Природа . 325 (6104): 487. Бибкод : 1987Natur.325..487R . дои : 10.1038/325487b0 . S2CID 4259887 .
- ^ Лопес-Арбарелло, А. (2012). «Филогенетические взаимоотношения гинглимодовых рыб (Actinopterygii: Neopterygii)» . ПЛОС ОДИН . 7 (7): e39370. Бибкод : 2012PLoSO...739370L . дои : 10.1371/journal.pone.0039370 . ПМК 3394768 . ПМИД 22808031 .
- ^ Крылья, О. (2007). «Обзор функции гастролитов с последствиями для ископаемых позвоночных и пересмотренная классификация» . Acta Palaeontologica Polonica . 52 (1): 1–16.
- ^ Бюффето, Э.; Мартилль, Д.; Эскуилье, Ф. (2004). «Птерозавры как часть диеты спинозавров» . Природа . 429 (6995): 33. Бибкод : 2004Natur.429...33B . дои : 10.1038/430033а . ПМИД 15229562 . S2CID 4398855 .
- ^ Терриен, Ф.; Хендерсон, Д.; Рафф, К. (2005). «Укуси меня - биомеханические модели нижних челюстей теропод и их влияние на пищевое поведение». В Карпентер, К. (ред.). Плотоядные динозавры . Издательство Университета Индианы. стр. 179–230. ISBN 978-0-253-34539-4 .
- ^ Андуза, Д.; Фаулер, Д.В. (2014). «Как спинозавриды могли ловить рыбу? Проверка поведенческих выводов путем сравнения с современными четвероногими, питающимися рыбой» . Рефераты с программами Геологического общества Америки . 46 (5): 12.
- ^ Вулло, Р.; Аллен, Р.; Кэвин, Л. (2016). «Конвергентная эволюция челюстей спинозавридных динозавров и щучьих морских угрей» . Acta Palaeontologica Polonica . 61 . дои : 10.4202/app.00284.2016 . S2CID 54514619 .
- ^ Баркер, Крис Тиджани; Нэйш, Даррен; Тренд, Джейкоб; Михельс, Лисанн Верле; Уитмер, Лоуренс; Риджли, Райан; Рэнкин, Кэти; Кларкин, Клэр Э.; Шнайдер, Филипп; Гостлинг, Нил Дж. (2023). «Модифицированные черепа, но консервативный мозг? Палеоневрология и эндокраниальная анатомия барионихиновых динозавров (Theropoda: Spinosauridae)» . Журнал анатомии . 242 (6): 1124–1145. дои : 10.1111/joa.13837 . ПМЦ 10184548 . ПМИД 36781174 .
- ^ Амиот, Р.; Бюффето, Э.; Лекуйер, К.; Ван, X.; Будад, Л.; Дин, З.; Фурель, Ф.; Хатт, С.; Мартино, Ф.; Медейрос, Массачусетс; Мо, Дж.; Саймон, Л.; Сутитхорн, В.; Свитман, С.; Тонг, Х.; Чжан, Ф.; Чжоу, З. (2010). «Изотопные доказательства полуводного образа жизни спинозавридных теропод». Геология . 38 (2): 139–142. Бибкод : 2010Geo....38..139A . дои : 10.1130/G30402.1 .
- ^ Продажи, МАФ; Ласерда, МБ; Хорн, БЛД; де Оливейра, IAP; Шульц, CL; Биби, Ф. (2016). «Х» материи: проверка связи между палеосредой и тремя кладами теропод» . ПЛОС ОДИН . 11 (2): e0147031. Бибкод : 2016PLoSO..1147031S . дои : 10.1371/journal.pone.0147031 . ПМЦ 4734717 . ПМИД 26829315 .
- ^ Фаббри, Маттео; Навалон, Гильермо; Бенсон, Роджер Б.Дж.; Пол, Диего; О'Коннор, Цзинмай; Бхуллар, Бхарт-Анжан С.; Эриксон, Грегори М.; Норелл, Марк А.; Оркнейские острова, Эндрю; Ламанна, Мэтью С.; Зухри, Самир; Беккер, Жюстин; Эмке, Аманда; Даль Сассо, Криштиану; Бинделлини, Габриэле; Магануко, Симоне; Аудиторе, Марко; Ибрагим, Низар (23 марта 2022 г.). «Подводное кормление хищных динозавров» . Природы . 603 (7903): 852–857. Бибкод : 2022Natur.603..852F . дои : 10.1038/s41586-022-04528-0 . ISSN 1476-4687 . ПМИД 35322229 . S2CID 247630374 .
- ^ «У спинозавра были кости, похожие на пингвиньи, что было признаком подводной охоты» . Наука . 23 марта 2022 г. Архивировано из оригинала 23 марта 2022 г.
- ^ Музей, Поле (23 марта 2022 г.). «Плотные кости позволяли спинозавру охотиться под водой, как показывают исследования» . физ.орг .
- ^ Рэдли, доктор медицинских наук; Аллен, П. (2012). «Южный английский Уэлден (неморской нижний мел): обзор палеосреды и палеоэкологии». Труды Ассоциации геологов . 123 (2): 382–385. Бибкод : 2012ПрГА..123..382Р . дои : 10.1016/j.pgeola.2011.12.005 .
- ^ Рэдли, Джей Ди (2006). «Путеводитель по Уилдену I: Суббассейн Уилда». Геология сегодня . 22 (3): 109–118. Бибкод : 2006GeolT..22..109R . дои : 10.1111/j.1365-2451.2006.00563.x . S2CID 129247105 .
- ^ Удары, WT (1998). «Обзор динозавров нижнего и среднего мела Англии» . Бюллетени Музея естественной истории Нью-Мексико . 14 : 29–38.
- ^ Кук, Э.; Росс, Эй Джей (1996). «Стратиграфия, седиментология и палеонтология глины Нижнего Уилда (готерив) на заводе Keymer Tileworks, Западный Суссекс, южная Англия». Труды Ассоциации геологов . 107 (3): 231–239. Бибкод : 1996PrGA..107..231C . дои : 10.1016/S0016-7878(96)80031-9 .
- ^ Милнер, AC; Эванс, SE (1998). «Первое сообщение об земноводных и ящерицах из Уилдена (нижний мел) в Англии» . Бюллетени Музея естественной истории Нью-Мексико . 14 : 173–176.
- ^ Най, Э.; Файст-Буркхардт, С.; Хорн, диджей; Росс, Эй Джей; Уиттакер, Дж. Э. (2008). «Палеосреда, связанная с частичным скелетом игуанодона из глины Верхнего Уилда (баррем, ранний мел) на кирпичном заводе Smokejacks (Окли, Суррей, Великобритания), на основе палиноморфов и остракод». Меловые исследования . 29 (3): 417–444. Бибкод : 2008CrRes..29..417N . дои : 10.1016/j.cretres.2008.01.004 .
- ^ Кук, Э. (1997). «Седиментология и тафономия позвоночных костеносных фаций из Clockhouse Rock Store (Уилд-Клэй, поздний готерив), Кэпел, Суррей, Великобритания». Труды Ассоциации геологов . 108 (1): 49–56. Бибкод : 1997ПрГА..108...49С . дои : 10.1016/S0016-7878(97)80005-3 .
- ^ Росс, Эй Джей; Кук, Э. (1995). «Стратиграфия и палеонтология глины Верхнего Уилда (баррем) на кирпичном заводе Smokejacks, Окли, Суррей, Англия» (PDF) . Меловые исследования . 16 (6): 705–716. Бибкод : 1995CrRes..16..705R . дои : 10.1006/cres.1995.1044 .
- ^ Баттен, диджей (1998). «Палеоэкологические последствия растений, насекомых и других микрокаменелостей с органическими стенками в формации Уилд-Клей (нижний мел) на юго-востоке Англии». Меловые исследования . 19 (3–4): 279–315. Бибкод : 1998CrRes..19..279B . дои : 10.1006/cres.1998.0116 .
- ^ Мартилл, DM; Хатт, С. (1996). «Возможные зубы барионихидных динозавров из формации Уэссекс (нижний мел, баррем) острова Уайт, Англия». Труды Ассоциации геологов . 107 (2): 81–84. Бибкод : 1996ПрГА..107...81М . дои : 10.1016/S0016-7878(96)80001-0 .
Внешние ссылки
[ редактировать ]- Музей естественной истории – « Барионикс : открытие удивительного рыбоядного динозавра» – четырехминутное видео, представленное Анжелой К. Милнер.















![]() Эта статья была отправлена в WikiJournal of Science на внешнюю академическую рецензию в 2018 году ( отчеты рецензентов ). Обновленный контент был реинтегрирован на страницу Википедии по лицензии CC-BY-SA-3.0 ( 2018 ). Проверенная версия записи: Майкл Бек; и др. (2019). «Барионикс» (PDF) . Викижурнал науки . 2 (1): 3. дои : 10.15347/WJS/2019.003 . ISSN 2470-6345 . Викиданные Q63252951 .
Эта статья была отправлена в WikiJournal of Science на внешнюю академическую рецензию в 2018 году ( отчеты рецензентов ). Обновленный контент был реинтегрирован на страницу Википедии по лицензии CC-BY-SA-3.0 ( 2018 ). Проверенная версия записи: Майкл Бек; и др. (2019). «Барионикс» (PDF) . Викижурнал науки . 2 (1): 3. дои : 10.15347/WJS/2019.003 . ISSN 2470-6345 . Викиданные Q63252951 .
- Статьи Википедии, опубликованные в рецензируемой литературе
- Статьи Википедии, опубликованные в WikiJournal of Science
- Статьи, рецензируемые внешними экспертами
- Статьи Википедии, опубликованные в рецензируемой литературе (W2J)
- Спинозавриды
- Монотипические роды динозавров
- Барремская жизнь
- Раннемеловые динозавры Европы.
- Меловая Англия
- Окаменелости Англии
- Ископаемые таксоны описаны в 1986 г.
- Таксоны, названные Аланом Дж. Чаригом
- Таксоны, названные Анджелой Милнер