Дромеозавриды
| Дромеозавриды Временной диапазон:
| |
|---|---|

| |
| Коллекция ископаемых скелетов дромеозаврид. По часовой стрелке сверху слева: Deinonychus antirhopus (тяжело сложенный эвдромеозавр), Buitreraptor gonzalezorum (длиннорылый неэнлагиин), Velociraptor mongoliensis (небольшой велоцирапторин), Microraptor gui (крылатый микрораптор), Halszkaraptor escuilliei (полуводный халшкарапторин), Zhenyuanlong suni. (длиннокрылый дромеозаврид) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Клэйд : | Паравес |
| Семья: | † Дромеозавриды Мэтью и Браун , 1922 год. |
| Типовой вид | |
| † Дромеозавр альбертенсис Мэтью и Браун, 1922 год.
| |
| Подгруппы [ 5 ] | |
| |
| Синонимы | |
| |
Dromaeosauridae ( / ˌ d r ɒ m i . ə ˈ s ɔːr ɪ d iː / ) — семейство пернатых целурозавровых - динозавров теропод . маленького и среднего размера Как правило, это были пернатые хищники , процветавшие в меловой период . Название Dromaeosauridae означает «бегущие ящерицы», от греческого δρομαῖος ( dromaîos ), что означает «бегущий на полной скорости», «быстрый», и σαῦρος ( saûros ), что означает «ящерица». В неофициальном обиходе их часто называют хищниками. [ 6 ] (в честь Велоцираптора ), термин, популяризированный фильмом « Парк Юрского периода» ; несколько родов включают термин «хищник» прямо в свое название, и популярная культура стала подчеркивать их птичий внешний вид и предполагаемое птичье поведение.
Окаменелости дромеозаврид были найдены по всему миру в Северной Америке , Европе , Африке , Азии и Южной Америке , причем некоторые окаменелости позволяют предположить, что они населяли Австралию . также [ 7 ] Самые ранние окаменелости тела известны с раннего мела (145-140 миллионов лет назад), и они сохранились до конца мела ( маастрихтский этап, 66 млн лет назад), просуществовав до мел-палеогенового вымирания . Присутствие дромеозавридов уже в средней юре было предположено открытием изолированных ископаемых зубов, хотя никаких окаменелостей тела дромеозаврид этого периода обнаружено не было. [ 8 ] [ 9 ]
Описание
[ редактировать ]Техническая диагностика
[ редактировать ]Дромеозаврид диагностируют по следующим признакам: короткие Т-образные лобные кости , образующие ростральную границу надвисочного окна ; каудолатеральный нависающий выступ чешуйчатой кости ; латеральный отросток квадратной кости , который контактирует с квадратно-скуловой костью ; приподнятые, стебельчатые, парапофизы на спинных позвонках , видоизмененный педальный палец II; шевроны и презигапофизы хвостовых позвонков удлинены и охватывают несколько позвонков; наличие подсуставной ямки на клювовидном отростке . [ 10 ]
Размер и общее строение
[ редактировать ]

Дромеозавриды были динозаврами малого и среднего размера, длиной от 1,5–2,07 метра (4,9–6,8 футов) (в случае велоцираптора ) до почти 6 м (20 футов) или более (у ютараптора , дакотараптора и ахиллобатора ). [ 11 ] [ 12 ] Большие размеры, по-видимому, развивались среди дромеозаврид как минимум дважды; один раз среди дромеозавринов Utahraptor и Achillobator , и снова среди unenlagiines ( Austroraptor , длина которого составляла 5–6 м (16–20 футов)). Возможная третья линия гигантских дромеозаврид представлена изолированными зубами, найденными на Уайт острове в Англии . Зубы принадлежат животному размером с дромеозаврина ютараптора , но, судя по форме зубов, они принадлежат велоцирапторинам. [ 13 ] [ 14 ]
Характерное строение тела дромеозаврид помогло возродить теории о том, что динозавры могли быть активными, быстрыми и тесно связанными с птицами. Роберта Баккера Иллюстрация к монографии Джона Острома 1969 года. [ 15 ] показ дромеозаврида дейнониха в быстром темпе, является одной из самых влиятельных палеонтологических реконструкций в истории. [ 16 ] Строение тела дромеозаврид включает относительно большой череп, зазубренные зубы, узкую морду (исключение составляют производные дромеозаврины ) и глаза, обращенные вперед, что указывает на некоторую степень бинокулярного зрения. [ 17 ]

Дромеозавриды, как и большинство других теропод, имели умеренно длинную S-образную шею, а туловище было относительно коротким и глубоким. Как и у других манирапторов , у них были длинные руки, которые у некоторых видов можно было сложить вокруг тела, и относительно большие руки с тремя длинными пальцами (средний палец был самым длинным, а первый палец — самым коротким), заканчивающимися большими когтями. [ 10 ] Структура бедра дромеозаврид отличалась характерным большим лобковым башмаком, выступающим под основанием хвоста. На ногах дромеозаврид на втором пальце ноги был большой загнутый назад коготь. Их хвосты были тонкими, с длинными низкими позвонками без поперечного отростка и нервных отростков после 14-го хвостового позвонка. [ 10 ] Оссифицированные крючковидные отростки ребер обнаружены у нескольких дромеозаврид. [ 18 ] [ 19 ] [ 20 ]
Ступня
[ редактировать ]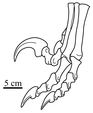
Как и другие тероподы, дромеозавриды были двуногими; то есть они ходили на задних ногах. Однако, хотя большинство теропод ходили, касаясь земли тремя пальцами ног, окаменелые следы ног подтверждают, что многие ранние паравианские группы, включая дромеозаврид, удерживали второй палец над землей в чрезмерно вытянутом положении, при этом только третий и четвертый пальцы несли вес животное. Это называется функциональной дидактилией. [ 21 ] Увеличенный второй палец имел необычно большой, изогнутый, серповидный (серповидный, альтернативно дрепаноидный ) коготь (удерживаемый над землей или «втягивающийся» при ходьбе), который, как полагают, использовался для захвата добычи и лазания по деревьям (см. «Функция когтя» ниже). Особенно клинкообразной эта клешня была у крупнотелых хищных эвдромеозавров . [ 22 ] Один из возможных видов дромеозаврид, Balaur Bondoc , также обладал первым пальцем ноги, который сильно видоизменился параллельно со вторым. И первый, и второй пальцы на каждой ноге B. Bondoc также были втянуты и имели увеличенные серповидные когти. [ 23 ]
Хвост
[ редактировать ]
Дромеозавриды имели длинные хвосты. Большинство хвостовых позвонков имели костные стержнеобразные отростки (так называемые презигапофизы), а также у некоторых видов костные сухожилия. В своем исследовании Дейнониха Остром предположил, что эти особенности придавали хвосту жесткость, так что он мог сгибаться только у основания, и тогда весь хвост мог двигаться как единый жесткий рычаг. [ 15 ] Однако один хорошо сохранившийся экземпляр Velociraptor mongoliensis (IGM 100/986) имеет сочлененный хвостовой скелет, изогнутый горизонтально в длинной S-образной форме. Это говорит о том, что в жизни хвост мог изгибаться из стороны в сторону со значительной степенью гибкости. [ 24 ] Было высказано предположение, что этот хвост использовался в качестве стабилизатора или противовеса во время бега или в воздухе; [ 24 ] у Микрораптора на конце хвоста сохранился удлиненный ромбовидный веер из перьев. Возможно, он использовался в качестве аэродинамического стабилизатора и руля направления во время планирования или полета с двигателем (см. «Полет и планирование» ниже). [ 25 ]
Перья
[ редактировать ]

Существует большое количество свидетельств того, что дромеозавриды были покрыты перьями . У некоторых окаменелостей дромеозаврид сохранились длинные перистые перья на кистях и руках ( ремигах ) и хвосте ( ректрисах ), а также более короткие, похожие на пух перья, покрывающие тело. [ 26 ] [ 27 ] Другие окаменелости, не сохранившие реальных отпечатков перьев, все еще сохраняют связанные с ними шишки на костях предплечий, где при жизни могли бы прикрепляться длинные перья крыльев. [ 28 ] В целом этот рисунок перьев очень похож на археоптерикса . [ 26 ]
Первым известным дромеозавридом с убедительными доказательствами наличия перьев был Sinornithosaurus , о котором сообщили из Китая Сюй и др. в 1999 году. [ 27 ] Многие другие окаменелости дромеозаврид были обнаружены с перьями, покрывающими их тела, а некоторые - с полностью развитыми оперенными крыльями. У микрораптора даже есть признаки наличия второй пары крыльев на задних лапах. [ 26 ] Хотя прямые отпечатки перьев возможны только в мелкозернистых отложениях, некоторые окаменелости, найденные в более грубых породах, свидетельствуют о перьях по наличию выступов игл - точек крепления перьев крыльев, которыми обладают некоторые птицы. У дромеозавридов Rahonavis и Velociraptor были обнаружены перья, что указывает на то, что у этих форм были перья, несмотря на отсутствие отпечатков. В свете этого наиболее вероятно, что даже более крупные наземные дромеозавриды имели перья, поскольку даже нелетающие птицы сегодня сохраняют большую часть своего оперения, а известно, что относительно крупные дромеозавриды, такие как велоцираптор , сохранили перистые перья. [ 28 ] [ 29 ] Хотя некоторые ученые предположили, что более крупные дромеозавриды частично или полностью утратили свое изолирующее покрытие, обнаружение перьев у экземпляров велоцирапторов было названо доказательством того, что все члены семейства сохранили перья. [ 28 ] [ 30 ]
Совсем недавно открытие Чжэньюаньлуна установило наличие полной оперенной шерсти у относительно крупных дромеозаврид. Кроме того, у животного наблюдаются пропорционально большие аэродинамические перья на крыльях, а также веерообразный хвост, обе эти неожиданные черты могут помочь понять покровы крупных дромеозаврид. [ 31 ] Дакотараптор — еще более крупный вид дромеозаврид с признаками перьев, хотя и косвенными в виде перьевых выступов. [ 32 ] хотя другие исследователи считают этот таксон химерой, поскольку даже динозавровые элементы с предполагаемыми диагностическими признаками дромеозавров также относятся к ценугнатидам и орнитомимозаврам . [ 33 ] [ 34 ]
Классификация
[ редактировать ]Отношения с птицами
[ редактировать ]
Дромеозавриды имеют много общих черт с ранними птицами (клады Avialae или Aves ). Точная природа их отношений с птицами подверглась тщательному изучению, и гипотезы об этих отношениях изменились по мере того, как стало доступно большое количество новых данных. Еще в 2001 году Марк Норелл и его коллеги проанализировали большое исследование окаменелостей целурозавров и пришли к предварительному выводу, что дромеозавриды были наиболее тесно связаны с птицами, а троодонтиды были более отдаленной внешней группой. Они даже предположили, что Dromaeosauridae могли быть парафилетическими по отношению к Avialae. [ 35 ] В 2002 году Хван и его коллеги использовали работу Норелла и др. , включая новые признаки и более качественные ископаемые свидетельства, чтобы определить, что птиц (авиалан) лучше считать двоюродными братьями дромеозаврид и троодонтид . [ 11 ] Палеонтологи сходятся во мнении, что пока недостаточно доказательств, чтобы определить, могли ли какие-либо дромеозавриды летать или планировать, или произошли ли они от предков, которые могли это делать. [ 36 ]
Альтернативные теории и нелетаемость
[ редактировать ]Дромеозавриды настолько похожи на птиц, что заставили некоторых исследователей утверждать, что их лучше отнести к птицам. Во-первых, поскольку у них были перья, дромеозавриды (наряду со многими другими целурозавровыми динозаврами-тероподами) являются «птицами» в соответствии с традиционными определениями слова «птица» или «авес», основанными на наличии перьев. Однако другие ученые, такие как Лоуренс Уитмер , утверждают, что называть теропода, такого как каудиптерикс, птицей, потому что у него есть перья, это слово может выйти за рамки любого полезного значения. [ 37 ]

По крайней мере две школы исследователей предположили, что дромеозавриды на самом деле могли произойти от летающих предков. Гипотезы о летающем предке дромеозаврид иногда называют «Птицы пришли первыми» (BCF). Георгия Ольшевского обычно называют первым автором BCF. [ 38 ] В своей работе Грегори С. Пол указал на многочисленные особенности скелета дромеозаврид, которые он интерпретировал как свидетельство того, что вся группа произошла от летающих предков-динозавров, возможно, от животного, подобного археоптериксу . В этом случае более крупные дромеозавриды были вторично нелетающими, как современный страус . [ 29 ] В 1988 году Пол предположил, что дромеозавриды на самом деле могут быть более тесно связаны с современными птицами, чем с археоптериксами . Однако к 2002 году Пол назвал дромеозаврид и археоптерикса ближайшими родственниками друг друга. [ 39 ]
В 2002 году Хван и др. обнаружили, что Микрораптор был самым примитивным дромеозавридом. [ 11 ] Сюй и его коллеги в 2003 году назвали базальное положение микрораптора , а также особенности перьев и крыльев доказательством того, что предки дромеозаврид могли планировать. В этом случае более крупные дромеозавриды были бы вторично наземными животными, потеряв способность планировать на более позднем этапе своей эволюционной истории. [ 26 ]
Также в 2002 году Стивен Черкас описал Cryptovolans , хотя это, вероятно, младший синоним Microraptor . Он неточно реконструировал окаменелость, имеющую только два крыла, и таким образом утверждал, что дромеозавриды были летающими животными, а не пассивными планерами. Позже он выпустил пересмотренную реконструкцию, соответствующую реконструкции Microraptor. [ 40 ]
Другие исследователи, такие как Ларри Мартин , предположили, что дромеозавриды, как и все манирапторы, вообще не были динозаврами. Мартин на протяжении десятилетий утверждал, что птицы не связаны с манирапторами, но в 2004 году он изменил свою позицию, согласившись, что эти двое являются близкими родственниками. Однако Мартин считал, что манирапторы были вторично нелетающими птицами и что птицы произошли не от динозавров, а скорее от нединозавровых архозавров. [ 41 ]

В 2005 году Майр и Питерс описали анатомию очень хорошо сохранившегося экземпляра археоптерикса и определили, что его анатомия больше похожа на нептичьих теропод, чем предполагалось ранее. В частности, они обнаружили, что у археоптерикса были примитивные небные кости , неперевернутый большой палец и сверхразгибаемый второй палец. Их филогенетический анализ дал противоречивый результат: Confuciusornis был ближе к Microraptor, чем к Archeopteryx , что сделало Avialae парафилетическим таксоном. Они также предположили, что предки паравиан были способны летать или планировать, а дромеозавриды и троодонтиды были вторично нелетающими (или утратили способность планировать). [ 43 ] [ 44 ] Корф и Батлер раскритиковали эту работу по методологическим соображениям. [ 45 ]
Вызов всем этим альтернативным сценариям возник, когда Тернер и его коллеги в 2007 году описали нового дромеозаврида Махакалу , который, как они обнаружили, является самым базальным и самым примитивным представителем Dromaeosauridae, более примитивным, чем Microraptor . У Махакалы были короткие руки и он не умел планировать. Тернер и др. также предположили, что полет развился только у Avialae, и эти два факта позволяют предположить, что предки дромеозаврид не могли планировать или летать. Основываясь на этом кладистическом анализе, Махакала предполагает, что предковое состояние дромеозаврид не является летальным . [ 46 ] Однако в 2012 году расширенное и пересмотренное исследование, включающее самые последние находки дромеозаврид, выявило Xiaotingia археоптериксоподобную как самого примитивного члена клады Dromaeosauridae, что, по-видимому, позволяет предположить, что самые ранние представители клады могли быть способны к полету. [ 47 ]
Таксономия
[ редактировать ]Авторство семейства Dromaeosauridae приписывают Уильяму Диллеру Мэтью и Барнуму Брауну , которые в 1922 году возвели его как подсемейство (Dromaeosaurinae) семейства Deinodontidae , содержащее только новый род Dromaeosaurus . [ 48 ]
Подсемейства Dromaeosauridae часто меняют свое содержание на основе нового анализа, но обычно состоят из следующих групп. Ряд дромеозавридов не были отнесены к какому-либо конкретному подсемейству, часто потому, что они слишком плохо сохранились, чтобы их можно было с уверенностью отнести к филогенетическому анализу (см. Раздел «Филогения» ниже), или они неопределенны и отнесены к разным группам в зависимости от методологии, используемой в разных статьях. . Самое основное известное подсемейство дромеозаврид — Halszkaraptorinae, группа причудливых существ с длинными пальцами и шеей, большим количеством мелких зубов и возможным полуводным образом жизни. [ 49 ] Другая загадочная группа, Unenlagiinae, является наиболее плохо поддерживаемым подсемейством дромеозаврид, и вполне возможно, что некоторые или все ее члены принадлежат за пределами Dromaeosauridae. [ 50 ] [ 51 ] Более крупные наземные представители, такие как Buitreraptor и Unenlagia, демонстрируют сильную адаптацию к полету, хотя они, вероятно, были слишком большими, чтобы «взлететь». Один из возможных членов этой группы, Рахонавис , очень маленький, с хорошо развитыми крыльями, на которых видны выступы игл (точки крепления маховых перьев), и весьма вероятно, что он мог летать. Следующая по примитивности клада дромеозавридов — микрораптории. В эту группу входят многие из самых маленьких дромеозавридов, приспособленных к жизни на деревьях. Все известные отпечатки кожи дромеозаврид происходят из этой группы, и все они демонстрируют обширный перьевой покров и хорошо развитые крылья. Как и уненлагиины, некоторые виды могли быть способны к активному полету. Самая развитая подгруппа дромеозаврид, Eudromaeosauria, включает коренастые и коротконогие роды, которые, вероятно, были охотниками из засад. В эту группу входят Velociraptorinae, Dromaeosaurinae, а в некоторых исследованиях и третья группа: Saurornitholestinae. Подсемейство Velociraptorinae традиционно включает Velociraptor , Deinonychus и Saurornitholestes , и хотя открытие Tsaagan оказало поддержку этой группе, включение Deinonychus , Saurornitholestes и нескольких других родов все еще остается неопределенным. Dromaeosaurinae обычно состоит из видов среднего и гигантского размера, обычно с черепами коробчатой формы (другие подсемейства обычно имеют более узкую морду). [ 5 ]





Следующая классификация различных родов дромеозаврид соответствует таблице, представленной Holtz, 2011, если не указано иное. [ 5 ]
- Семейство дромеозавриды.
- Нутеты
- Пампа Раптор
- Варираптор
- Пирораптор
- Чжэньюаньлун
- Даурлонг
- Подсемейство Halszkaraptorinae. [ 49 ]
- Подсемейство Unenlagiinae.
- Подсемейство Microraptorinae.
- Узел Эудромеозаврия [ 22 ]
- Дейноних
- Динеобеллатор
- вектираптор
- Подсемейство Saurornitholestinae.
- Подсемейство Velociraptorinae.
- Подсемейство Dromaeosaurinae.
Филогения
[ редактировать ]Dromaeosauridae были впервые определены Полом Серено как 1998 клада в году как наиболее инклюзивная естественная группа, содержащая Dromaeosaurus , но не троодона , орнитомима или пассера . Различные «подсемейства» также были переопределены как клады, обычно определяемые как все виды, более близкие к тезке группы, чем к Dromaeosaurus или любым тезкам других субклад (например, Маковицкий определил кладу Unenlagiinae как все дромеозавриды, более близкие к Unenlagia). чем велоцираптор ). Microraptoria — единственный субклад дромеозаврид, не преобразованный из подсемейства. Сентер и его коллеги специально придумали название без суффикса подсемейства -inae , чтобы избежать предполагаемых проблем с созданием традиционного таксона семейной группы, если будет обнаружено, что группа находится за пределами собственно дромеозаврид. [ 53 ] Серено предложил пересмотренное определение подгруппы, содержащей Microraptor, чтобы гарантировать, что она попадает в состав Dromaeosauridae, и выделил подсемейство Microraptorinae, приписав его Senter et al. , хотя это использование появилось только в его онлайн-базе данных TaxonSearch и официально не публиковалось. [ 54 ] Обширный кладистический анализ, проведенный Turner et al. (2012) также подтвердили монофилию Dromaeosauridae. [ 55 ]
Приведенная ниже кладограмма соответствует анализу 2015 года, проведенному DePalma et al. используя обновленные данные Рабочей группы Theropod. [ 32 ]
| Дромеозавриды | |


Другая кладограмма, построенная ниже, соответствует филогенетическому анализу, проведенному в 2017 году Cau et al. используя обновленные данные Рабочей группы Theropod в своем описании Halszkaraptor . [ 49 ]
| Дромеозавриды | |



Палеобиология
[ редактировать ]Чувства
[ редактировать ]Сравнение склеральных колец нескольких дромеозаврид ( Микрораптор , Синорнитозавр и Велоцираптор ) и современных птиц и рептилий показывает, что некоторые дромеозавриды (включая Микрораптора и Велоцираптора ), возможно, были ночными хищниками, в то время как Синорнитозавр предположительно вел катемерный образ жизни (активен в течение дня в короткие интервалы). [ 56 ] Однако открытие радужного оперения у Microraptor поставило под сомнение вывод о ночном образе жизни этого рода, поскольку не известно, чтобы современные птицы с радужным оперением вели ночной образ жизни. [ 57 ]
Исследования обонятельных луковиц дромеозавридов показывают, что они имели такое же соотношение обонятельных способностей для своего размера, как и другие нептичьи тероподы и современные птицы с острым обонянием, такие как тираннозавриды и стервятники-индейки , что, вероятно, отражает важность обонятельного чувства в повседневная деятельность дромеозаврид, такая как поиск еды. [ 58 ] [ 59 ]
Кормление
[ редактировать ]Было обнаружено, что питание дромеозаврид типично для целурозавровых теропод с характерным методом кормления «прокол и вытягивание». Исследования характера стирания зубов дромеозаврид, проведенные Анжеликой Торисес и др. указывают на то, что зубы дромеозаврид имеют схожий характер износа с зубами тираннозаврид и троодонтид. Однако микроизнос на зубах показал, что дромеозавриды, вероятно, предпочитали более крупную добычу, чем троодонтиды, с которыми они часто делили свою среду обитания. Такая диетическая дифференциация, вероятно, позволила им обитать в одной и той же среде. То же исследование также показало, что дромеозавриды, такие как Dromaeosaurus и Saurornitholestes (два дромеозаврида, проанализированные в исследовании), вероятно, включали в свой рацион кости и были лучше приспособлены к борьбе с добычей, в то время как троодонтиды, оснащенные более слабыми челюстями, охотились на более мягких животных и объекты добычи, такие как как беспозвоночные и падаль. [ 60 ]
Функция когтя
[ редактировать ]

В настоящее время существуют разногласия по поводу функции увеличенного «серповидного когтя» на втором пальце ноги. Когда Джон Остром описал его для Дейнониха в 1969 году, он интерпретировал коготь как похожее на лезвие рубящее оружие, очень похожее на клыки некоторых саблезубых кошек , которые используются мощными ударами, чтобы разрезать добычу. Адамс (1987) предположил, что коготь использовался для потрошения крупных цератопсовых динозавров. [ 61 ] Интерпретация серповидного когтя как орудия убийства применялась ко всем дромеозавридам. Однако Мэннинг и др. утверждали, что вместо этого коготь служил крючком, реконструируя кератиновую оболочку с эллиптическим поперечным сечением вместо ранее предполагаемой формы перевернутой капли. [ 62 ] По интерпретации Мэннинга, коготь на втором пальце ноги будет использоваться как средство для лазания при покорении более крупной добычи, а также как колющее оружие.
Остром сравнил Дейнониха со страусом и казуаром . Он отметил, что этот вид птиц может нанести серьезную травму большим когтем на втором пальце ноги. [ 15 ] Казуар имеет когти длиной до 125 миллиметров (4,9 дюйма). [ 63 ] Остром процитировал Гиллиарда (1958), сказавшего, что они могут отрубить руку или выпотрошить человека. [ 64 ] Кофрон (1999 и 2003) изучил 241 задокументированное нападение казуаров и обнаружил, что один человек и две собаки были убиты, но нет доказательств того, что казуары могут потрошить или расчленить других животных. [ 65 ] [ 66 ] Казуары используют свои когти, чтобы защитить себя, атаковать угрожающих животных, а также в агонистических проявлениях, таких как демонстрация угрозы с поклоном. [ 63 ] У сериемы также есть увеличенный коготь на втором пальце ноги, и он использует его, чтобы разрывать мелкую добычу и проглатывать. [ 67 ]
Филип Мэннинг и его коллеги (2009) попытались проверить функцию серповидных когтей и когтей аналогичной формы на передних конечностях. Они проанализировали биомеханику того, как напряжения и деформации будут распределяться вдоль когтей и конечностей, используя рентгеновскую визуализацию для создания трехмерной контурной карты когтя передней конечности велоцираптора . Для сравнения они проанализировали строение клешни современной хищной птицы — филина . Они обнаружили, что, судя по способу воздействия на коготь, они идеально подходят для лазания. Ученые обнаружили, что заостренный кончик когтя является колющим и захватывающим инструментом, а изогнутое и расширенное основание когтя помогает равномерно передавать стрессовые нагрузки. Команда Мэннинга также сравнила кривизну «серповидного когтя» дромеозаврид с кривизной современных птиц и млекопитающих. Предыдущие исследования показали, что степень кривизны когтей соответствует образу жизни животного: животные с сильно изогнутыми когтями определенной формы склонны к лазанию, а более прямые когти указывают на наземный образ жизни. Серповидные когти дромеозаврид У дейнонихов кривизна составляет 160 градусов, что вполне соответствует возможностям лазающих животных. Изученные ими когти передних конечностей также находились в пределах кривизны подъема. [ 68 ]
Палеонтолог Питер Маковицки прокомментировал исследование команды Мэннинга, заявив, что маленькие примитивные дромеозавриды (такие как Microraptor ), вероятно, лазали по деревьям, но это лазание не объясняет, почему позже гигантские дромеозавриды, такие как Achillobator, сохраняли сильно изогнутые когти, когда они были слишком велики, чтобы лазить по деревьям. Маковицки предположил, что гигантские дромеозавриды, возможно, приспособили коготь исключительно для захвата добычи. [ 69 ]

В 2009 году Фил Сентер опубликовал исследование пальцев ног дромеозаврид и показал, что диапазон их движений совместим с раскапыванием прочных гнезд насекомых. Сентер предположил, что мелкие дромеозавриды, такие как Rahonavis и Buitreraptor, были достаточно малы, чтобы быть частично насекомоядными , в то время как более крупные роды, такие как Deinonychus и Neuquenraptor, могли использовать эту способность для ловли позвоночных жертв, обитающих в гнездах насекомых. Однако Сентер не проверял, способствует ли сильная кривизна когтей дромеозаврид такой деятельности. [ 70 ]
В 2011 году Денвер Фаулер и его коллеги предложили новый метод, с помощью которого дромеозавриды могли ловить более мелкую добычу. Эта модель, известная как модель хищничества «хищник-хищник» (RPR), предполагает, что дромеозавриды убивали свою добычу способом, очень похожим на ныне живущих хищных птиц-ястребов : прыгая на свою добычу, прижимая ее под весом своего тела и крепко сжимая его большими серповидными когтями. Как и акципитриды, дромеозавриды затем начали питаться живым животным, пока оно в конечном итоге не умерло от потери крови и недостаточности органов. Это предложение основано прежде всего на сравнении морфологии и пропорций ступней и ног дромеозаврид с несколькими группами современных хищных птиц с известным хищническим поведением. Фаулер обнаружил, что ступни и ноги дромеозаврид больше всего напоминают ноги орлов и ястребов , особенно с точки зрения наличия увеличенной второй клешни и аналогичного диапазона хватательных движений. Однако короткая плюсна и сила стопы были бы больше похожи на таковые у совы . Метод хищничества RPR согласуется с другими аспектами анатомии дромеозаврид, такими как их необычный зубной ряд и морфология рук. Руки, которые могли оказывать большую силу, но, вероятно, были покрыты длинными перьями, возможно, использовались в качестве стабилизаторов для равновесия, когда они находились на вершине борющегося животного-жертвы, вместе с жестким уравновешивающим хвост. Челюсти дромеозаврид, которые Фаулер и его коллеги считали сравнительно слабыми, были бы полезны для поедания добычи живьем, но не так полезны для быстрого и мощного уничтожения добычи. Совместная работа этих хищнических адаптаций может также иметь значение для возникновения взмахов крыльями у паравианов . [ 71 ] [ 72 ]

В 2019 году Питер Бишоп реконструировал скелет ноги и мускулатуру дейнониха , используя трехмерные модели мышц , сухожилий и костей . Добавив математические модели и уравнения, Бишоп смоделировал условия, которые обеспечили бы максимальную силу на кончике серповидного когтя и, следовательно, наиболее вероятную функцию. Среди предлагаемых способов использования серповидных когтей: удар ногой с целью разрезать, порезать или выпотрошить добычу; для захвата добычи за бока; пирсинг с помощью веса тела; атаковать жизненно важные участки добычи; сдерживать добычу; внутри- или межвидовая конкуренция ; и выкапывание добычи из укрытий. Результаты, полученные Бишопом, показали, что положение приседа увеличивает силу когтей, однако эти силы остаются относительно слабыми, что указывает на то, что когти недостаточно сильны, чтобы их можно было использовать для рубящих ударов. Серповидные когти, скорее, использовались не для рубящих ударов, а для согнутых ног, например, для удержания добычи и нанесения ударов по ней с близкого расстояния. Эти результаты согласуются с боевых динозавров Образец , на котором велоцираптор и протоцератопс сцепились в бою, причем первый сжимает другого когтями в позе невытянутых ног. Несмотря на полученные результаты, Бишоп считал, что возможности серповидного когтя могли варьироваться в пределах таксона, учитывая, что среди дромеозаврид Адазавр имел необычно меньшую серповидную клешню, сохраняющую характерный гинглимоид - структуру, разделенную на две части - и сверхрастяжимую суставную поверхность. предпоследняя фаланга. Он не мог ни подтвердить, ни оставить без внимания тот факт, что педальный палец II мог потерять или сохранить свою функциональность. [ 73 ]
Исследование, проведенное Джанечини и др . в 2020 году, также показывает, что велоцирапторины, дромеозаврины и другие эвдромеозавры Лавразии сильно отличались по своим локомотивам и методам убийства от унэнлагииновых дромеозавридов Гондваны. Более короткая вторая фаланга второго пальца стопы позволяла генерировать повышенную силу этим пальцем, что в сочетании с более короткой и широкой плюсной и заметной выраженной шарнирной морфологией суставных поверхностей плюсневых костей и фаланг, возможно, позволило эвдромеозаврам проявлять большую силу захвата, чем уненлагиинам, что позволило более эффективно подавлять и убивать крупную добычу. Для сравнения, у унэнлагииновых дромеозаврид были более длинные и тонкие субарктометатарзусы и менее выраженные шарнирные суставы - черта, которая, возможно, давала им большую способность к передвижению и позволяла развивать большую скорость. Кроме того, более длинная вторая фаланга второго пальца позволяла unenlagiines быстро двигать вторыми пальцами ног, чтобы охотиться на более мелкие и неуловимые виды добычи. Эти различия в двигательной и хищнической специализации, возможно, были ключевой особенностью, которая повлияла на пути эволюции, сформировавшие обе группы дромеозавров в северном и южном полушариях. [ 74 ]
Групповое поведение
[ редактировать ]
Окаменелости дейнониха были обнаружены небольшими группами рядом с останками травоядного тенонтозавра , более крупного птицетазового динозавра. Это было интерпретировано как свидетельство того, что эти дромеозавриды охотились слаженными стаями, как некоторые современные млекопитающие . [ 76 ] Однако не все палеонтологи нашли доказательства убедительными, и последующее исследование, опубликованное в 2007 году Роучем и Бринкманом, предполагает, что дейнонихи, возможно, на самом деле демонстрировали дезорганизованное поведение толпы. Современные диапсиды , включая птиц и крокодилов (ближайшие родственники дромеозаврид), демонстрируют минимальную длительную совместную охоту (за исключением сокола апломадо и ястреба Харриса ); вместо этого они обычно являются охотниками-одиночками, либо время от времени объединяющими свои силы, чтобы увеличить успех охоты (как иногда делают крокодилы), либо их тянет к ранее убитым тушам, где часто возникают конфликты между особями одного и того же вида. Например, в ситуациях, когда группы варанов Комодо едят вместе, самые крупные особи едят первыми и могут атаковать более мелких варанов Комодо, которые пытаются питаться; если меньшее животное умирает, его обычно съедают . Когда эту информацию применить к местам, демонстрирующим предполагаемое охотничье поведение дромеозавридов, она кажется в некоторой степени согласующейся со стратегией питания, подобной варанам Комодо. Останки скелета дейнониха , обнаруженные на этих участках, принадлежат молодым особям, с отсутствующими частями, которые, возможно, были съедены другими дейнонихами , что, по данным исследования Роуча и соавт. представлены как свидетельство против идеи о том, что животные сотрудничали в охоте. [ 77 ] Различные диетические предпочтения молодых и взрослых дейнонихов, опубликованные в 2020 году, указывают на то, что это животное не проявляло сложного кооперативного поведения, наблюдаемого у животных, охотящихся вьюком. Распространено ли это на других дромеозавров, в настоящее время неизвестно. [ 78 ] Третий возможный вариант заключается в том, что дромеозавриды не проявляли долгосрочного кооперативного поведения, но демонстрировали краткосрочное кооперативное поведение, как это наблюдается у крокодилов, которые демонстрируют как истинное сотрудничество, так и конкуренцию за добычу.
В 2001 году на участке, который некоторые считают ловушкой для хищников из зыбучих песков, было обнаружено несколько экземпляров ютараптора в возрасте от взрослой особи до крошечного детеныша длиной три фута. Некоторые считают это свидетельством семейного охотничьего поведения; однако весь блок песчаника еще не открыт, и исследователи не уверены, погибли ли животные одновременно или нет. [ 79 ]
В 2007 году ученые описали первый известный обширный путь дромеозаврид в Шаньдуне, Китай. В дополнение к подтверждению гипотезы о том, что серповидный коготь удерживался втянутым от земли, след (проложенный крупным видом размером с ахиллобатора ) показал свидетельства того, что шесть особей примерно одинакового размера двигались вместе вдоль береговой линии. Особи находились на расстоянии примерно одного метра друг от друга, двигались в одном направлении и шли довольно медленно. Авторы статьи, описывающей эти следы, интерпретировали следы как свидетельство того, что некоторые виды дромеозаврид жили группами. Хотя следы явно не отражают охотничье поведение, по мнению авторов, нельзя исключать идею о том, что группы дромеозаврид могли охотиться вместе. [ 21 ]
Летать и планировать
[ редактировать ]
Предплечья дромеозаврид, по-видимому, хорошо приспособлены к сопротивлению скручивающим и изгибающим напряжениям, связанным с взмахами и скольжением. [ 80 ] а способность летать или планировать была предложена как минимум для пяти видов дромеозаврид. Первый, Rahonavis ostromi (первоначально классифицировался как птичья птица, но в более поздних исследованиях оказался дромеозавридом). [ 17 ] [ 81 ] ), возможно, был способен к полету с приводом , о чем свидетельствуют его длинные передние конечности со следами прикрепления игольчатых выступов к длинным прочным маховым перьям. [ 82 ] Передние конечности Рахонависа были более мощными, чем у Археоптерикса , и свидетельствуют о том, что они имели прочные связки, необходимые для машущего полета. Луис Кьяппе пришел к выводу, что с учетом этих адаптаций Рахонавис, вероятно, мог летать, но был бы более неуклюжим в воздухе, чем современные птицы. [ 83 ]
Другой вид дромеозаврид, Microraptor gui , возможно, был способен планировать, используя хорошо развитые крылья как на передних, так и на задних конечностях. Исследование Санкара Чаттерджи, проведенное в 2005 году , показало, что крылья Микрораптора функционировали как двухуровневый « биплан » и что он, вероятно, использовал фугоидный стиль планирования, при котором он запускался с насеста и пикировал вниз по U-образной кривой. , затем снова поднимите его и приземлитесь на другое дерево, при этом хвост и задние крылья помогают контролировать его положение и скорость. Чаттерджи также обнаружил, что у Микрораптора есть основные требования для поддержания горизонтального полета в дополнение к планированию. [ 25 ]
Changyuraptor yangi — близкий родственник Microraptor gui , который также считается планёром или летающим аппаратом из-за наличия четырёх крыльев и схожих пропорций конечностей. Тем не менее, это значительно более крупное животное, размером с дикую индейку , входящее в число крупнейших известных летающих мезозойских паравианов.
Другой вид дромеозаврид, Deinonychus antirhopus , может демонстрировать частичную способность к полету. У детенышей этого вида были более длинные руки и более крепкие грудные пояса, чем у взрослых, и они были похожи на те, что наблюдались у других машущих теропод, а это означает, что они, возможно, были способны летать в молодом возрасте, а затем теряли эту способность по мере роста. [ 84 ]
Возможность того, что Sinornithosaurus millenii был способен планировать или даже летать с приводом, также обсуждалась несколько раз. [ 85 ] [ 86 ] хотя никаких дальнейших исследований не проводилось.
Чжэньюаньлун сохраняет перья крыльев аэродинамической формы с особенно птичьими кроющими, в отличие от более длинных и широких кроющих форм, таких как Archeopteryx и Anchiornis , а также сросшиеся грудинные пластины. Из-за своего размера и коротких рук маловероятно, что Чжэньюаньлун был способен летать с помощью двигателя (хотя в этом отношении подчеркивается важность биомеханического моделирования). [ 31 ] ), но это может указывать на относительно близкое происхождение от летающих предков или даже на некоторую способность к планированию или бегу по наклонной поверхности с помощью крыльев .
Палеопатология
[ редактировать ]В 2001 году Брюс Ротшильд и другие опубликовали исследование, в котором изучались доказательства стрессовых переломов и отрывов сухожилий у теропод- динозавров, а также их последствия для их поведения. Поскольку стрессовые переломы вызваны повторяющейся травмой, а не единичными событиями, они с большей вероятностью могут быть вызваны регулярным поведением, чем другие типы травм. Исследователи обнаружили повреждения, подобные тем, которые вызваны стрессовыми переломами, на когтях рук дромеозавридов, одно из двух таких повреждений когтей, обнаруженных в ходе исследования. Стресс-переломы кистей имеют особое поведенческое значение по сравнению со стрессовыми переломами стоп, поскольку стресс-переломы стоп могут быть получены во время бега или во время миграции. Травмы рук, напротив, с большей вероятностью могут быть получены при контакте с борющейся добычей. [ 87 ]
Плавание
[ редактировать ]
По крайней мере, одна группа дромеозаврид, Halszkaraptorinae , членами которой являются halszkaraptorines, скорее всего, специализировалась на водном или полуводном образе жизни, имея развитые пропорции конечностей, морфологию зубов и грудную клетку, сходную с таковыми у ныряющих птиц. [ 49 ] [ 88 ] [ 89 ]
были предложены рыболовные привычки Для unenlagiine , включая сравнения с приписываемыми полуводными спинозавридами . [ 90 ] но какие-либо водные двигательные механизмы до сих пор не обсуждались.
Воспроизведение
[ редактировать ]
В 2006 году Грелле-Тиннер и Маковицки сообщили об яйце, связанном с экземпляром дейнониха . Яйцо имеет сходство с яйцами овирапторидов , и авторы интерпретировали эту связь как потенциально указывающий на высиживание. [ 91 ] Исследование, опубликованное в ноябре 2018 года Нореллом, Янгом и Виманом и др., показывает, что дейноних откладывает голубые яйца, вероятно, для их маскировки, а также для создания открытых гнезд. Другие дромеозавриды, возможно, делали то же самое, и предполагается, что они и другие динозавры-манирапторы могли быть исходной точкой для откладывания крашеных яиц и создания открытых гнезд, как это делают многие птицы сегодня. [ 92 ] [ 93 ] [ 94 ]
В популярной культуре
[ редактировать ]Велоцираптор , дромеозаврид, привлек большое внимание после того, как он был показан в Стивена Спилберга фильме 1993 года «Парк Юрского периода» . Однако размеры велоцираптора в фильме намного больше, чем у самых крупных представителей этого рода. Роберт Баккер напомнил, что Спилберг был разочарован размерами Велоцираптора и поэтому увеличил его размеры. [ 95 ] Грегори С. Пол в своей книге « Хищные динозавры мира» 1988 года также считал Deinonychus antirhopus разновидностью велоцираптора и переименовал вид Velociraptor antirhopus . [ 39 ] Это таксономическое мнение не получило широкого распространения. [ 10 ] [ 96 ] [ 97 ]
Хронология родов дромеозаврид
[ редактировать ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Хартман, Скотт; Мортимер, Микки; Уол, Уильям Р.; Ломакс, Дин Р.; Липпинкотт, Джессика; Лавлейс, Дэвид М. (10 июля 2019 г.). «Новый паравианский динозавр из поздней юры Северной Америки подтверждает позднее приобретение птичьего полета» . ПерДж . 7 : е7247. дои : 10.7717/peerj.7247 . ISSN 2167-8359 . ПМК 6626525 . ПМИД 31333906 .
- ^ Порфирий, Джон Д.; Баяно, Маттиа А.; дос Сантос, Доменика Д.; Джанекини, Федерико А.; Питтман, Майкл; Ламанна, Мэтью К. (14 июня 2024 г.). « Diuqin lechiguanae gen. et sp. nov., новый уненлагиин (Theropoda: Paraves) из формации Бахо-де-ла-Карпа (группа Неукен, верхний мел) провинции Неукен, Патагония, Аргентина» . BMC Экология и эволюция . 24 (1): 77. Бибкод : 2024BMCEE..24...77P . дои : 10.1186/s12862-024-02247-w . ISSN 2730-7182 . ПМЦ 11177497 . ПМИД 38872101 .
- ^ Аллен, Ронан; Переда-Субербиола, Хавье (2003). «Динозавры Франции». Аккаунты Палевол . 2 (1): 27–44. Бибкод : 2003CRPal...2...27A . дои : 10.1016/S1631-0683(03)00002-2 .
- ^ Аньолин, Федерико Л.; Мотта, Матиас Дж.; Бриссон Эгли, Федерико; Ло Коко, Гастон; Новас, Фернандо Э. (12 февраля 2019 г.). «Паравианская филогения и переход динозавр-птица: обзор» . Границы в науках о Земле . 6 . дои : 10.3389/feart.2018.00252 . hdl : 11336/130197 . ISSN 2296-6463 .
- ^ Перейти обратно: а б с Хольц, Томас Р. младший (2012) Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов, приложение, зима 2011 г.
- ^ Желудь, Дж. (2007). Глубокая Альберта: факты об ископаемых и раскопки динозавров . Издательство Университета Альберты. стр. 13 . дои : 10.1515/9780888648518 . ISBN 978-0-88864-481-7 .
- ^ Барретт, Пол М.; Бенсон, Роджер Б.Дж.; Рич, Томас Х.; Викерс-Рич, Патрисия (23 декабря 2011 г.). «Первый динозавр-спинозавр из Австралии и космополитизм меловой фауны динозавров» . Письма по биологии . 7 (6): 933–936. дои : 10.1098/rsbl.2011.0466 . ПМК 3210678 . ПМИД 21693488 .
- ^ Меткалф, SJ; Воган, РФ; Бентон, MJ; Коул, Дж.; Симмс, MJ; Дартналл, Д.Л. (январь 1992 г.). «Новое местонахождение микропозвоночных батского периода (средней юры) в пределах известняковой формации Чиппинг-Нортон в карьере Хорнслисоу, Глостершир». Труды Общества геологов . 103 (4): 321–342. Бибкод : 1992ПрГА..103..321М . дои : 10.1016/S0016-7878(08)80129-0 .
- ^ Уиллс, С.; Андервуд, CJ; Барретт, премьер-министр (2023). «Машинное обучение подтверждает новые данные о тероподах-манирапторах в фауне микропозвоночных Великобритании средней юры» . Статьи по палеонтологии . 9 (2). е1487. Бибкод : 2023PPal....9E1487W . дои : 10.1002/spp2.1487 .
- ^ Перейти обратно: а б с д Норелл, Массачусетс; Маковицкий, П.Дж. (2004). «Дромеозавриды». В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 196–210. ISBN 978-0-520-24209-8 .
- ^ Перейти обратно: а б с Хван, Ш.; Норелл, Массачусетс; Джи, К.; Гао, К. (2002). «Новые экземпляры Microraptor zhaoianus (Theropoda: Dromaeosauridae) из Северо-Восточного Китая». Новитаты Американского музея (3381): 44 стр. HDL : 2246/2870 .
- ^ Перл, А.; Норелл, Массачусетс; Кларк, Дж. (1999). «Новый теропод-манираптор – Achillobator giganticus (Dromaeosauridae) – из верхнего мела Бурханта, Монголия». Вклад монгольско-американского палеонтологического проекта . 101 : 1–105.
- ^ Нэйш, Д. Хатт и Мартилл, DM (2001). «Ящеротазовые динозавры: тероподы». в Мартилле Д.М. и Нэйше Д. (ред.). Динозавры острова Уайт . Палеонтологическая ассоциация, Полевые справочники по окаменелостям. 10, 242–309.
- ^ «Дино охотится сетью на редкие зубы хищника» . Новости Би-би-си . 26 апреля 2004 г.
- ^ Перейти обратно: а б с Остром, Дж. Х. (1969). «Остеология Deinonychus antirhopus , необычного теропода из нижнего мела Монтаны» (PDF) . Бюллетень Музея естественной истории Пибоди . 30 :1–165. Архивировано из оригинала (PDF) 27 июня 2010 г. Проверено 18 декабря 2010 г.
- ^ Пол, Грег (январь 2006 г.). «Рисуем динозавров» . Геотаймс .
- ^ Перейти обратно: а б Маковицкий, Питер Дж.; Апестегия, Себастьян; Аньолин, Федерико Л. (октябрь 2005 г.). «Самый ранний теропод дромеозаврид из Южной Америки». Природа . 437 (7061): 1007–1011. Бибкод : 2005Natur.437.1007M . дои : 10.1038/nature03996 . ПМИД 16222297 . S2CID 27078534 .
- ^ Аньолин, Федерико; Новас, Фернандо Э. (2013). Птичьи предки . SpringerBriefs по наукам о системе Земли. Дордрехт: Springer Нидерланды. дои : 10.1007/978-94-007-5637-3 . ISBN 978-94-007-5636-6 . S2CID 199493087 .
- ^ Кодд, Джонатан Р.; Мэннинг, Филип Л.; Норелл, Марк А; Перри, Стивен Ф (22 января 2008 г.). «Птичье дыхание у динозавров-манирапторов» . Труды Королевского общества B: Биологические науки . 275 (1631): 157–161. дои : 10.1098/rspb.2007.1233 . ISSN 0962-8452 . ПМК 2596187 . ПМИД 17986432 .
- ^ Пол, Грегори С. (1988). Хищные динозавры мира: полный иллюстрированный справочник . Нью-Йорк. ISBN 0-671-61946-2 . OCLC 18350868 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Перейти обратно: а б Ли, Рихуэй; Локли, Мартин Г.; Маковицкий, Питер Дж.; Мацукава, Масаки; Норелл, Марк А.; Харрис, Джеральд Д.; Лю, Минвэй (19 октября 2007 г.). «Поведенческие и фаунистические последствия следов дейнонихозавров раннего мела из Китая». Naturwissenschaften . 95 (3): 185–191. Бибкод : 2008NW.....95..185L . дои : 10.1007/s00114-007-0310-7 . ПМИД 17952398 . S2CID 16380823 .
- ^ Перейти обратно: а б Лонгрич, Северная Каролина; Карри, Пи Джей (2009). «Микрорапторин (Dinosauria–Dromaeosauridae) из позднего мела Северной Америки» . ПНАС . 106 (13): 5002–7. Бибкод : 2009PNAS..106.5002L . дои : 10.1073/pnas.0811664106 . ПМК 2664043 . ПМИД 19289829 .
- ^ Чики, Золтан; Времир, Матьяш; Брусатте, Стивен Л.; Норелл, Марк А. (31 августа 2010 г.). «Аберрантный островной динозавр-теропод из позднего мела Румынии» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (35): 15357–15361. Бибкод : 2010PNAS..10715357C . дои : 10.1073/pnas.1006970107 . ПМЦ 2932599 . ПМИД 20805514 .
- ^ Перейти обратно: а б Норелл, Марк А.; Маковицкий, Питер Дж. (1999). «Важные особенности скелета дромеозаврида II: информация из недавно собранных экземпляров Velociraptor mongoliensis ». Новитаты Американского музея (3282): 1–45. HDL : 2246/3025 .
- ^ Перейти обратно: а б Чаттерджи, Санкар; Темплин, Р. Джек (30 января 2007 г.). «Форма крыла биплана и летные характеристики пернатого динозавра Microraptor gui» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (5): 1576–1580. Бибкод : 2007PNAS..104.1576C . дои : 10.1073/pnas.0609975104 . ПМК 1780066 . ПМИД 17242354 .
- ^ Перейти обратно: а б с д ; Ван, Сюэвэнь; Чжан, Ду, Сянке (январь 2003 г.) Сюй, Син, Чжунхэ PDF) 421 . ( ( 6921): 335–340 . Бибкод : 2003Natur.421..335X . doi : 10.1038 . PMID 12540892 . / nature01342
- ^ Перейти обратно: а б Сюй, Син; Ван, Сяо-Линь; Ву, Сяо-Чун (сентябрь 1999 г.). «Динозавр-дромеозавр с нитевидным покровом из формации Исянь в Китае». Природа . 401 (6750): 262–266. Бибкод : 1999Natur.401..262X . дои : 10.1038/45769 . S2CID 4430574 .
- ^ Перейти обратно: а б с Тернер, А.Х.; Маковицкий, П.Дж.; Норелл, Массачусетс (21 сентября 2007 г.). «Ручки перьевого пера в динозавре-велоцирапторе» . Наука . 317 (5845): 1721. Бибкод : 2007Sci...317.1721T . дои : 10.1126/science.1145076 . ПМИД 17885130 .
- ^ Перейти обратно: а б Пол, Грегори С. (2002). Воздушные динозавры: эволюция и потеря полета у динозавров и птиц. Балтимор: Издательство Университета Джонса Хопкинса. 472 стр.
- ^ Прум, Ричард О.; Браш, Алан Х. (сентябрь 2002 г.). «Эволюционное происхождение и разнообразие перьев». Ежеквартальный обзор биологии . 77 (3): 261–295. дои : 10.1086/341993 . ПМИД 12365352 . S2CID 6344830 .
- ^ Перейти обратно: а б Лю, Цзюньчан; Брусатте, Стивен Л. (16 июля 2015 г.). «Крупный короткорукий крылатый дромеозаврид (Dinosauria: Theropoda) из раннего мела Китая и его значение для эволюции перьев» . Научные отчеты . 5 (1): 11775. Бибкод : 2015NatSR...511775L . дои : 10.1038/srep11775 . ПМК 4504142 . ПМИД 26181289 .
- ^ Перейти обратно: а б «Первый гигантский хищник (Theropoda: Dromaeosauridae) из формации Хелл-Крик» . Палеонтологический вклад . 30 октября 2015 г. doi : 10.17161/paleo.1808.18764 .
- ^ Ясински, Стивен Э.; Салливан, Роберт М.; Додсон, Питер (26 марта 2020 г.). «Новый динозавр-дромеозаврид (Theropoda, Dromaeosauridae) из Нью-Мексико и биоразнообразие дромеозаврид в конце мелового периода» . Научные отчеты . 10 (1): 5105. Бибкод : 2020NatSR..10.5105J . дои : 10.1038/s41598-020-61480-7 . ISSN 2045-2322 . ПМК 7099077 . ПМИД 32218481 .
- ^ Кау, А. (2024). «Единая основа макроэволюции хищных динозавров» (PDF) . Боллеттино делла Сосьета Палеонтологическое Итальянское . 63 (1). дои : 10.4435/BSPI.2024.08 . Архивировано из оригинала (PDF) 27 апреля 2024 г.
- ^ Норелл, М. Кларк, Дж. М., Маковицкий, П. Дж. (2001). « Филогенетические отношения между целурозавровыми тероподами. Архивировано 13 июля 2007 г. в Wayback Machine. « Новые перспективы происхождения и эволюции птиц: материалы Международного симпозиума в честь Джона Х. Острома», Йельский музей Пибоди: 49–67.
- ^ Падиан, Кевин; Дайал, Кеннет П. (2005). «Происхождение полета: могли ли «четырехкрылые» динозавры летать?» . Природа . 438 (7066): E3, обсуждение E3–4. Бибкод : 2005Natur.438E...3P . дои : 10.1038/nature04354 . ПМИД 16292258 . S2CID 4314459 .
- ^ Уитмер, Л.М. (2005) «Дебаты о птичьем происхождении; филогения, функции и окаменелости». В книге «Мезозойские птицы: над головами динозавров», стр. 3–30. ISBN 0-520-20094-2
- ^ Ольшевский, Георгий. (1994). «Сначала птицы? Теория, соответствующая фактам - эволюция рептилий в птиц». Омни , июнь 1994. Том 16 №9
- ^ Перейти обратно: а б Пол, Грегори С. (1988). Хищные динозавры мира. Нью-Йорк: Саймон и Шустер. 464 стр.
- ^ Черкас С.А., Чжан Д., Ли Дж. и Ли Ю. (2002). «Летающие дромеозавры», в Черкасе, С.Дж. (ред.): Пернатые динозавры и происхождение полета: Журнал Музея динозавров 1. Бландинг: Музей динозавров, 16–26.
- ^ Мартин, LD (2004). «Базальное архозавровое происхождение птиц» (PDF) . Акта Геологика Синика . 50 (6): 978–990.
- ^ National Geographic News - У самой ранней птицы были ноги, как у динозавра, шоу окаменелостей - Николас Бакалар, 1 декабря 2005 г., страница 2. Проверено 18 октября 2006 г.
- ^ Майр, Г.; Пол, Б; Петерс, Д.С. (2 декабря 2005 г.). «Хорошо сохранившийся образец археоптерикса с чертами теропода» (PDF) . Наука . 310 (5753): 1483–1486. Бибкод : 2005Sci...310.1483M . дои : 10.1126/science.1120331 . ПМИД 16322455 . S2CID 28611454 .
- ^ Майр, Г. (1 сентября 2006 г.). «Ответ на комментарий к статье «Хорошо сохранившийся образец археоптерикса с чертами теропода» » . Наука . 313 (5791): 1238с. Бибкод : 2006Sci...313.1238M . дои : 10.1126/science.1130964 .
- ^ Корф, Эй-Джей; Батлер, Р.Дж. (1 сентября 2006 г.). «Комментарий к «Хорошо сохранившемуся образцу археоптерикса с чертами теропода» ». Наука . 313 (5791): 1238б. Бибкод : 2006Sci...313.1238C . дои : 10.1126/science.1130800 . ПМИД 16946054 . S2CID 19859320 .
- ^ Тернер, Алан Х.; Пол, Диего; Кларк, Джулия А.; Эриксон, Грегори М.; Норелл, Марк (2007). «Базальный дромеозаврид и эволюция размеров, предшествовавшая полету птиц» . Наука . 317 (5843): 1378–1381. Бибкод : 2007Sci...317.1378T . дои : 10.1126/science.1144066 . ПМИД 17823350 .
- ^ Перейти обратно: а б Сентер, П.; Киркланд, Дж.И.; Дембье, Д.Д.; Мэдсен, С.; Тот, Н. (2012). Додсон, Питер (ред.). «Новые дромеозавриды (Dinosauria: Theropoda) из нижнего мела штата Юта и эволюция хвоста дромеозаврид» . ПЛОС ОДИН . 7 (5): e36790. Бибкод : 2012PLoSO...736790S . дои : 10.1371/journal.pone.0036790 . ПМК 3352940 . ПМИД 22615813 .
- ^ Мэтью, штат Вашингтон; Браун, Б. (1922). «Семейство Deinodontidae с уведомлением о новом роде из мелового периода Альберты». Бюллетень Американского музея естественной истории . 46 : 367–385. HDL : 2246/1300 .
- ^ Перейти обратно: а б с д Кау, А.; Бейранд, В.; Воетен, DFAE; Фернандес, В.; Таффоро, П.; Штейн, К.; Барсболд, Р.; Цогтбаатар, К.; Карри, Пи Джей; Годфруа, П. (2017). «Синхротронное сканирование выявило экоморфологию земноводных в новой кладе птицеподобных динозавров» . Природа . 552 (7685): 395–399. Бибкод : 2017Natur.552..395C . дои : 10.1038/nature24679 . ПМИД 29211712 . S2CID 4471941 .
- ^ Тернер, Алан Х.; Хван, Санни Х.; Норелл, Марк А. (2007). «Маленький теропод из Ооша, ранний мел, Байхангор, Монголия» . Новитаты Американского музея (3557): 1. doi : 10.1206/0003-0082(2007)3557[1:ASDTFS]2.0.CO;2 . hdl : 2246/5845 . S2CID 31096081 .
- ^ Аньолин, Федерико Л.; Новас, Фернандо Э. (март 2011 г.). «Уненлагиидные тероподы: являются ли они представителями Dromaeosauridae (Theropoda, Maniraptora)?» . Анналы Бразильской академии наук . 83 (1): 117–162. дои : 10.1590/s0001-37652011000100008 . ПМИД 21437379 .
- ^ Лонгрич, Николас Р.; Карри, Филип Дж. (31 марта 2009 г.). «Микрорапторин (Dinosauria–Dromaeosauridae) из позднего мела Северной Америки» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (13): 5002–5007. Бибкод : 2009PNAS..106.5002L . дои : 10.1073/pnas.0811664106 . ПМК 2664043 . ПМИД 19289829 .
- ^ Сентер, Фил; Барсболд, Р.; Бритт, Брукс Б.; Бернхэм, Дэвид Б. (2004). «Систематика и эволюция Dromaeosauridae (Dinosauria, Theropoda)». Бюллетень Музея естественной истории Гумма . 8 : 1–20.
- ^ Серено, ПК, 2005. Stem Archosauria — TaxonSearch. Архивировано 15 января 2009 г. на Wayback Machine , версия 1.0, 7 ноября 2005 г.
- ^ Тернер, Алан Х.; Маковицкий, Питер Дж.; Норелл, Марк А. (17 августа 2012 г.). «Обзор систематики дромеозаврид и филогении паравианов» . Бюллетень Американского музея естественной истории . 371 : 1–206. дои : 10.1206/748.1 . hdl : 2246/6352 . S2CID 83572446 .
- ^ Шмитц, Л.; Мотани, Р. (14 апреля 2011 г.). «Ночной образ жизни динозавров, выведенный на основании морфологии склерального кольца и орбиты». Наука . 332 (6030): 705–708. Бибкод : 2011Sci...332..705S . дои : 10.1126/science.1200043 . ПМИД 21493820 . S2CID 33253407 .
- ^ Ли, Цюаньго; Гао, К.-К.; Мэн, К.; Кларк, Дж.А.; Шоки, доктор медицины; д'Альба, Л.; Пей, Р.; Эллисон, М.; Норелл, Массачусетс; Винтер, Дж. (9 марта 2012 г.). «Реконструкция микрораптора и эволюция радужного оперения». Наука . 335 (6073): 1215–1219. Бибкод : 2012Sci...335.1215L . дои : 10.1126/science.1213780 . ПМИД 22403389 . S2CID 206537426 .
- ^ Зеленицкий, Дарла К; Терриен, Франсуа; Кобаяши, Ёсицугу (28 октября 2008 г.). «Острота обоняния теропод: палеобиологические и эволюционные последствия» . Труды Королевского общества B: Биологические науки . 276 (1657): 667–673. дои : 10.1098/rspb.2008.1075 . ПМК 2660930 . ПМИД 18957367 .
- ^ Зеленицкий, Дарла К.; Терриен, Франсуа; Риджли, Райан С.; МакГи, Аманда Р.; Уитмер, Лоуренс М. (13 апреля 2011 г.). «Эволюция обоняния нептичьих тероподов, динозавров и птиц» . Труды Королевского общества B: Биологические науки . 278 (1725): 3625–3634. дои : 10.1098/rspb.2011.0238 . ПМЦ 3203493 . ПМИД 21490022 .
- ^ Торичес, А.; Уилкинсон, Р.; Арбур, ВМ; Руис-Оменьака, Дж.И.; Карри, Пи Джей (2018). «Биомеханика прокола и вытягивания в зубах хищных целурозавровых динозавров» . Современная биология . 28 (9): 1467–1474. Бибкод : 2018CBio...28E1467T . дои : 10.1016/j.cub.2018.03.042 . ПМИД 29706515 .
- ^ Адамс, Дон (1987). «Чем они больше, тем сильнее они падают: последствия седалищной кривизны у цератопсовых динозавров». В Карри, Филип Дж.; Костер, Э. (ред.). Четвертый симпозиум по мезозойским наземным экосистемам . Драмхеллер, Канада: Музей Тиррелла. стр. 1–6.
- ^ Мэннинг, Филип Л.; Пейн, Дэвид; Пенникотт, Джон; Барретт, Пол М; Эннос, Роланд А. (11 октября 2005 г.). «Когти-убийцы динозавров или кошки для лазания?» . Письма по биологии . 2 (1): 110–112. дои : 10.1098/rsbl.2005.0395 . ПМК 1617199 . PMID 17148340 .
- ^ Перейти обратно: а б Дэвис, SJJF (2002) «Ратиты и тинамус» Oxford University Press. Нью-Йорк, США [ нужна страница ]
- ^ Жильяр, Эрнест Томас (1958). Живые птицы мира . Даблдэй. ОСЛК 681791956 . [ нужна страница ]
- ^ Кофрон, Кристофер П. (1999). «Нападения южного казуара (Casuarius casuarius johnsonii) на людей и домашних животных в Квинсленде, Австралия». Журнал зоологии . 249 (4): 375–381. дои : 10.1111/j.1469-7998.1999.tb01206.x .
- ^ Кофрон, Кристофер П. (2003). «Истории случаев нападений южного казуара в Квинсленде». Мемуары Квинслендского музея . 49 (1): 335–338.
- ^ Редфорд, Кент Х.; Петерс, Густав (1986). «Заметки по биологии и песне красноногой сериемы (Cariama cristata)». Журнал полевой орнитологии . 57 (4): 261–269. JSTOR 4513154 .
- ^ Мэннинг, Филип Л.; Маргеттс, Ли; Джонсон, Марк Р.; Уизерс, Филип Дж.; Селлерс, Уильям И.; Фолкингем, Питер Л.; Маммери, Пол М.; Барретт, Пол М.; Рэймонт, Дэвид Р. (сентябрь 2009 г.). «Биомеханика когтей динозавров-дромеозавров: применение рентгеновской микротомографии, наноиндентирования и анализа методом конечных элементов» . Анатомические записи: достижения в интегративной анатомии и эволюционной биологии . 292 (9): 1397–1405. дои : 10.1002/ar.20986 . ПМИД 19711472 .
- ^ ««Смертельные» когти велоцираптора предназначались для лазания» . Новый учёный . 203 (2725): 10 сентября 2009 г. doi : 10.1016/s0262-4079(09)62381-2 . Проверено 15 сентября 2009 г.
- ^ Сентер, Фил (2009). «Функция педали у дейнонихозавров (Dinosauria: Theropoda): сравнительное исследование» (PDF) . Бюллетень Музея естественной истории Гумма . 13 : 1–14.
- ^ Фаулер, Денвер В.; Фридман, Элизабет А.; Сканнелла, Джон Б.; Камбич, Роберт Э. (14 декабря 2011 г.). «Хищная экология дейнониха и происхождение взмахов руками у птиц» . ПЛОС ОДИН . 6 (12): e28964. Бибкод : 2011PLoSO...628964F . дои : 10.1371/journal.pone.0028964 . ПМЦ 3237572 . ПМИД 22194962 .
- ^ Чой, Чарльз (14 декабря 2011 г.). «Когти-убийцы велоцирапторов помогали им поедать добычу живьем» . ЖиваяНаука .
- ^ Бишоп, Пи Джей (2019). «Тестирование функции серповидных когтей дромеозаврид (Dinosauria, Theropoda) посредством моделирования и оптимизации скелетно-мышечной системы» . ПерДж . 7 : е7577. дои : 10.7717/peerj.7577 . ПМК 6717499 . ПМИД 31523517 .
- ^ Джанекини, Федерико А.; Эрколи, Маркос Д.; Диас-Мартинес, Игнасио (5 февраля 2020 г.). «Дифференциальные локомоторные и хищнические стратегии гондванцев и производных лавразийских дромеозаврид (Dinosauria, Theropoda, Paraves): выводы из морфометрических и сравнительно-анатомических исследований» . Журнал анатомии . 236 (5): 772–797. дои : 10.1111/joa.13153 . ПМЦ 7163733 . ПМИД 32023660 .
- ^ Мудрох, А.; Рихтер, У.; Джогер, У.; Косма, Р.; Иде, О.; Мага, А. (2011). «Дидактильные следы паравианских теропод (Maniraptora) из «средней юры Африки» . ПЛОС ОДИН . 6 (2): e1462. Бибкод : 2011PLoSO...614642M . дои : 10.1371/journal.pone.0014642 . ПМК 3038851 . ПМИД 21339816 . Рисунок 4.
- ^ Максвелл, В. Десмонд; Остром, Джон Х. (27 декабря 1995 г.). «Тафономия и палеобиологические последствия ассоциаций тенонтозавра и дейнониха ». Журнал палеонтологии позвоночных . 15 (4): 707–712. Бибкод : 1995JVPal..15..707M . дои : 10.1080/02724634.1995.10011256 .
- ^ Роуч, Брайан Т.; Бринкман, Дэниел Л. (апрель 2007 г.). «Переоценка совместной охоты стаей и стадности Deinonychus antirhopus и других нонавианских тероподных динозавров». Бюллетень Музея естественной истории Пибоди . 48 (1): 103–138. doi : 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 . S2CID 84175628 .
- ^ Фредериксон, Дж.А.; Энгель, МХ; Чифелли, РЛ (15 августа 2020 г.). «Онтогенетические изменения в рационе Deinonychus antirhopus (Theropoda; Dromaeosauridae): понимание экологии и социального поведения хищных динозавров посредством анализа стабильных изотопов». Палеогеография, Палеоклиматология, Палеоэкология . 552 : 109780. Бибкод : 2020PPP...55209780F . дои : 10.1016/j.palaeo.2020.109780 . S2CID 219059665 .
- ^ Свитек, Брайан (7 января 2015 г.). «Смертельная ловушка для динозавров в Юте обнаружила клад гигантских хищников» . Национальные географические новости . Архивировано из оригинала 30 ноября 2019 года.
- ^ Хатсон, Джоэл Д.; Хатсон, Келда Н. (25 сентября 2015 г.). «Исследование подвижности костей предплечья у Alligator Mississippiensis (Daudin, 1802) и Struthio Camelus Linnaeus, 1758 показывает, что археоптерикс и дромеозавры имеют общую адаптацию к планированию и/или взмахам крыльями» . Геодиверситас . 37 (3): 325–344. дои : 10.5252/g2015n3a3 . S2CID 55278850 . Проверено 2 декабря 2022 г.
- ^ Норелл, Марк А.; Кларк, Джеймс М.; Тернер, Алан Х.; Маковицкий, Питер Дж.; Барсболд, Ринчен; Роу, Тимоти (2006). «Новый теропод-дромеозавр из Ухаа Толгод (Омнёгов, Монголия) » Новитаты Американского музея (3545): 1. doi : 10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2 . hdl : 2246/5823 .
- ^ Форстер, Калифорния; Сэмпсон, SD; Кьяппе, LM; Краузе, Д.В. (20 марта 1998 г.). «Происхождение птиц от теропод: новые данные из позднего мела Мадагаскара». Наука . 279 (5358): 1915–1919. Бибкод : 1998Sci...279.1915F . дои : 10.1126/science.279.5358.1915 . ПМИД 9506938 .
- ^ Кьяппе, LM (2 февраля 2007 г.). Прославленные динозавры: происхождение и ранняя эволюция птиц . Сидней: UNSW Press. ISBN 978-0-471-24723-4 . [ нужна страница ]
- ^ Парсонс, Уильям Л.; Парсонс, Кристен М.; Додсон, Питер (15 апреля 2015 г.). «Морфологические вариации в онтогенезе Deinonychus antirhopus (Theropoda, Dromaeosauridae)» . ПЛОС ОДИН . 10 (4): e0121476. Бибкод : 2015PLoSO..1021476P . дои : 10.1371/journal.pone.0121476 . ПМЦ 4398413 . ПМИД 25875499 .
- ^ Чаттерджи, Санкар; Темплин, Р.Дж. (2004). «Пернатые целурозавры из Китая: новый взгляд на древесное происхождение полета птиц» . В Карри, Филип Дж.; Коппельхус, Ева Б.; Шугар, Мартин А.; Райт, Джоанна Л. (ред.). Пернатые драконы: исследования перехода от динозавров к птицам . Издательство Университета Индианы. стр. 251–281. ISBN 978-0-253-34373-4 .
- ^ Нэйш, Даррен (2012). Планета динозавров: следующее поколение гигантов-убийц . Книги Светлячка. п. 192 . ISBN 978-1-77085-049-1 .
- ^ Ротшильд, Б.; Танке, DH; Форд, ТЛ (2001). «Стрессовые переломы теропод и разрывы сухожилий как ключ к активности». В Танке, DH; Карпентер, К. (ред.). Жизнь мезозойских позвоночных . Издательство Университета Индианы. стр. 331–336.
- ^ Кау, А. (2020). «Сплан тела Halszkaraptor escuilliei (Dinosauria, Theropoda) не является переходной формой в эволюции гиперплотоядных дромеозаврид» . ПерДж . 8 : е8672. дои : 10.7717/peerj.8672 . ПМК 7047864 . ПМИД 32140312 .
- ^ Ли, С.; Ли, Ю.-Н.; Карри, Пи Джей; Сиссонс, Р.; Парк, Ж.-Ю.; Ким, С.-Х.; Барсболд, Р.; Цогтбаатар, К. (2022). «Нептичий динозавр с обтекаемым телом демонстрирует потенциальные приспособления для плавания» . Коммуникационная биология . 5 (1185): 1185. doi : 10.1038/s42003-022-04119-9 . ISSN 2399-3642 . ПМЦ 9715538 . ПМИД 36456823 .
- ^ Карри, Филип Дж.; Карабахал, Ариана Паулина (декабрь 2012 г.). «Новый экземпляр Austroraptor cabazai Novas, Pol, Canale, Porfiri и Calvo, 2008 (Dinosauria, Theropoda, Unenlagiidae) из позднего мела (маастрихта) Рио-Негро, Аргентина». Амегиниана . 49 (4): 662–667. дои : 10.5710/AMGH.30.8.2012.574 . hdl : 11336/9090 . S2CID 129058582 .
- ^ Греллет-Тиннер, Джеральд; Маковицкий, Питер (1 июня 2006 г.). «Возможное яйцо дромеозавра Deinonychus antirhopus : филогенетическое и биологическое значение». Канадский журнал наук о Земле . 43 (6): 705–719. Бибкод : 2006CaJES..43..705G . дои : 10.1139/E06-033 .
- ^ Виманн, Жасмина; Ян, Цзы-Руэй; Норелл, Марк А. (31 октября 2018 г.). «Цвет яиц динозавров имел единое эволюционное происхождение». Природа . 563 (7732): 555–558. Бибкод : 2018Natur.563..555W . дои : 10.1038/s41586-018-0646-5 . ПМИД 30464264 . S2CID 53188171 .
- ^ «Цвет яиц динозавров имел единое эволюционное происхождение» . Ноябрь 2018.
- ^ «Эволюционно говоря, динозавры складывали все цветные птичьи яйца в одну корзину» . 2018-10-31.
- ^ Баккер, Роберт Т. (1995). Раптор Красный . Нью-Йорк: Bantam Books. п. 4. ISBN 978-0-553-57561-3 .
- ^ Перес-Морено, BP; Санс, Дж.Л.; Судре, Дж.; Сиге, Б. (1994). «Динозавр-теропод из нижнего мела на юге Франции». «Динозавры и другие ископаемые рептилии Европы», Второй симпозиум Жоржа Кювье, Монбельяр; Журнал палеобиологии, специальный том . 7 : 173–188.
- ^ Карри, Пи Джей (1995). «Новая информация об анатомии и взаимоотношениях Dromaeosaurus albertensis (Dinosauria: Theropoda)». Журнал палеонтологии позвоночных . 15 (3): 576–591. Бибкод : 1995JVPal..15..576C . дои : 10.1080/02724634.1995.10011250 . ( аннотация. Архивировано 27 сентября 2007 г. в Wayback Machine )
Внешние ссылки
[ редактировать ]- Дромеозавриды: Хищники! , из Музея палеонтологии Калифорнийского университета в Беркли.
- Динозавры - полное и бесплатное онлайн-издание книги «Динозавры», написанной У. Д. Мэтью (цитируемым в этой статье под авторством семейства Dromaeosauridae) и бывшим куратором отдела палеонтологии позвоночных в Американском музее естественной истории в Нью-Йорке; Первоначально опубликовано в 1915 году.
- Dromaeosauridae , справочник по миру динозавров с подробным описанием и изображениями многих динозавров-дромеозаврид.

