Tyrannosauridae
| Тиранозавриды Временный диапазон: поздний меховой ,
| |
|---|---|

| |
| Монтаж шести тиранозавридов, по часовой стрелке сверху слева: тиранозавр , дасплетозавр , Tarbosaurus , Gorgosaurus , Zhuchengtyrannus и Alioramus | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Динозаурия |
| Клада : | Сауриския |
| Клада : | Теропода |
| Суперсемейство: | † Тиранозавра |
| Клада : | † Pantyrannososauria |
| Клада : | † Eutyrannososauria |
| Семья: | † Tyrannososauridae Осборн , 1906 |
| Тип рода | |
| † Тиранозавр Осборн, 1905
| |
| Подгруппы [ 1 ] | |
| |
| Синонимы | |
Tyrannososauridae (или тиранозавриды , то есть « ящерицы тирана ») представляет собой с коелурозавринами которые , семейство динозавров включают в себя два подсемейства, содержащие до пятнадцати родов , включая одноименный тиранозавр . Точное количество родов является спорным, некоторые эксперты признают всего три. Все эти животные жили в конце мелового периода , и их окаменелости были найдены только в Северной Америке и Азии .
Хотя тиранозавриды происходили из более мелких предков , почти всегда были самыми крупными хищниками в соответствующих экосистемах , ставя их на вершину цепи пищевой . Самым большим видом был Tyrannosaurus rex , самый массивный известный наземный хищник, который длился более 13 метров (43 фута). [ 2 ] и в соответствии с большинством современных оценок до 8,87 метрических тонн (9,78 коротких тонн) в весе. [ 3 ] [ 4 ] Тиранозавриды были двуножными плотоядными животными с массивными черепами, наполненными большими зубами. Несмотря на их большой размер, их ноги были длинными и пропорциональными для быстрого движения. Напротив, их руки были очень маленькими, носили только две функциональные цифры .
В отличие от большинства других групп динозавров, очень полные останки были обнаружены для большинства известных тираннозавридов. Это позволило различные исследования в их биологии . Научные исследования были сосредоточены на их онтогении , биомеханике и экологии , среди других предметов.
История открытия
[ редактировать ]
Первые остатки тираннозавридов были обнаружены во время экспедиций, проводимых геологической службой Канады , которая располагала многочисленными разбросанными зубами. Этим отличительным зубам динозавров было дано название Deinodon («ужасный зуб») Джозефа Лейди Первые хорошие образцы тиранозаврида были обнаружены в формировании каньона подкова Альберты в 1856 году . и состояли из почти полных черепа с частичными скелетами. Эти останки были впервые изучены Эдвардом Дим -Коупом в 1876 году, который считал их видом восточного тиранозавра . В 1905 году Генри Фэйрфилд Осборн признал, что Альберта остается значительно отличается от сухожира и придумал для них новое название: саркофаг Альбертозавра («Ящерица Альберты, питающаяся, питающаяся»). [ 5 ] Коп описал больше материала тиранозавра в 1892 году в форме изолированных позвонков и дал этому животному имя Manospondylus Gigas . Это открытие в основном было упущено более века и вызвало противоречие в начале 2000 -х годов, когда было обнаружено, что этот материал фактически принадлежал и имел приоритет над названием Tyrannosaurus rex . [ 6 ]

В своем документе 1905 года Albertosaurus Осборн описал два дополнительных образца тиранозавра, которые были собраны в Монтане и Вайоминге во время экспедиции Американского музея естественной истории 1902 года во главе с Барнумом Брауном . Первоначально Осборн считал, что это различные виды. Первый, он назвал империоз Dynamosaurus («Императорская ящерица»), а второй - Tyrannosaurus rex («Ящерица King Tyrant»). Год спустя Осборн признал, что эти два образца действительно поступили от одного и того же вида. Несмотря на то, что Dynamosaurus был найден в первую очередь, имя Tyrannosaurus появилось на одной странице ранее в своей оригинальной статье, описывающей оба образца. Поэтому, согласно Международному кодексу зоологической номенклатуры имя Tyrannosaurus . (ICZN), было использовано [ 7 ]
Барнум Браун продолжил собирать еще несколько образцов тиранозавридов из Альберты, в том числе первые, кто сохранил укороченные двухколотые передние конечности для группы (которую Лоуренс Ламбе назвал Горгозавр Libratus , «Сбалансированная ожесточенная ячейка», в 1914 году). Вторая значительная находка, приписываемая Gorgosaurus , была сделана в 1942 году в виде хорошо сохранившегося, хотя и необычно маленького, полного черепа. Образец ждал, пока после окончания Второй мировой войны будет изучен Чарльз У. Гилмор , который назвал его Gorgosaurus Lancenis . [ 5 ] Этот череп был переосмыслен Робертом Т. Баккером , Филом Керри и Майклом Уильямсом в 1988 году и назначен на новый род Nanotyrannus . [ 8 ] Также в 1946 году палеонтологи из Советского Союза начали экспедиции в Монголию и обнаружили первые останки тиранозавра из Азии. Евгений Малеев описал новые монгольские виды тиранозавра и Gorgosaurus в 1955 году и один новый род: Tarbosaurus («ужасающий ящерица»). Последующие исследования, однако, показали, что все виды тиранозавров Малеева на самом деле были одним из видов тарбозавра на разных стадиях роста. Позже был обнаружен второй вид монгольского тиранозаврида, описанный Сергеем Курзанов в 1976 году, и учитывая название Alioramus remotus («отдаленная различная ветвь»), хотя его статус настоящего тиранозаврида, а не более примитивный тираннозавр все еще остается спорным. [ 9 ] [ 5 ]
Описание
[ редактировать ]
Все тиранозавриды были крупными животными, со всеми видами, способными весом не менее 1 метрической тонны. [ 10 ] Был обнаружен один образец алиорамуса индивидуального, оцененный в длину от 5 до 6 метров (16 и 20 футов), [ 9 ] Хотя некоторые эксперты считаются несовершеннолетними. [ 10 ] [ 11 ] Albertosaurus , Gorgosaurus и Daspletosaurus измеряются от 8 до 9 метров (26 и 30 футов) [ 12 ] в то время как Tarbosaurus достиг длины 11 метров (36 футов) от морды до хвоста. [ 13 ] Массивный тиранозавр достиг 13 метров (43 фута) в самом большом образце, RSM P2523.8 . [ 14 ]
Череп и зубной зуб
[ редактировать ]



Анатомия черепа тиранозаврида хорошо понята, так как полные черепа известны всеми родами, но алиорамусом , который известен только по остаткам частичного черепа. [ 15 ] Tyrannosaurus , Tarbosaurus и Daspletosaurus имели черепа, которые превышали 1 м (3,3 фута) в длину. [ 12 ] У взрослых тиранозавридов были высокие, массивные черепа, с множеством костей, слитых и усиленных для силы. В то же время полые камеры во многих костях черепа и большие отверстия ( Fenestrae ) между этими костями помогли снизить вес черепа. Многие особенности черепов тиранозавридов также были обнаружены у их непосредственных предков, в том числе высокие предчелюрки и слитые носовые кости . [ 10 ]
Черепа Tyrannososaurid имели много уникальных характеристик, в том числе слитые теменные кости с выдающимся сагиттальным гребнем , который продольно проходил вдоль сагиттального шва и разделял два супратропоральных фенестра на крыше черепа. За этими фенестрами у тиранозавридов был характерно высокий нучальный гребень, который также возник из теменных, но бегал по поперечной плоскости, а не в продольном направлении. Нюшальный гребень был особенно хорошо развит в Тираннозавре , Тарбозавре и Алиорамусе . Альбертозавр , Дасплетозавр и Горгозавр имели высокие гребни перед глазами на слезных костях , в то время как у тарбозавра и тиранозавра были чрезвычайно утолщенные посторбитальные кости, образующие гребни в форме полумесяца за глазами. У Алиорамуса было ряд шести костных гребней поверх своей морды, возникающих из костей носа; Сообщалось о более низких гребнях в некоторых образцах дасплетозавра и тарбозавра , а также более базальных тиранозавра Аппалачиозавра . [ 11 ] [ 16 ] Торца и другие части черепа также носят многочисленные форамины . Согласно исследованию 2017 года, в котором описано D. horneri , чешуйчатый покров, а также тактильную чувствительность коррелировали с множественными рядами нервно -сосудистых отверстий, наблюдаемых у крокодилов и тиранозавридов. [ 17 ]


Тиранозавриды, как и их предки тиранозавра, были гетеродонтами , с предчелюстными зубами D, взысканными в поперечном сечении и меньше, чем остальные. В отличие от более ранних тиранозавра и большинства других теропод, верхнечелюстные и нижнечелюстные зубы зрелых тиранозавридов не являются лезвия, а чрезвычайно утолщенные и часто круговые по поперечному сечению, причем у некоторых видов есть пониженные зубчатые разрезы. [ 10 ] Число зубов, как правило, согласовано внутри видов, а более крупные виды имеют тенденцию иметь более низкий уровень зубов, чем более мелкие. Например, у Алиорамуса было от 76 до 78 зубов в челюстях, в то время как у Тиранозавра было от 54 до 60. [ 18 ]
В 2001 году Уильям Эблер заметил, что Albertosaurus зубные зубцы напоминают трещину в зубе, заканчивающемся круглой пустотой, называемой ампуллой. [ 19 ] Зубы тиранозавридов использовались в качестве удержания для стягивания мяса с тела, поэтому, когда тиранозавр потянул на кусок мяса, натяжение могло вызвать чисто, похожее на трещины, распространяться через зуб. [ 19 ] Однако наличие ампуллы распределяло бы эти силы по более крупной площади поверхности и снизило риск повреждения зуба при штамме. [ 19 ] Наличие разрезов, заканчивающихся пустотами, имеет параллели в человеческой инженерии. Производители гитары используют разрезы, заканчивающиеся пустотами, как описывает Аблер, «придают чередующиеся области гибкости и жесткости» древесину, с которой они работают. [ 19 ] Использование тренировки для создания своего рода «ампуллы» и предотвращения распространения трещин через материал также используется для защиты поверхностей самолета . [ 19 ] Эблер продемонстрировал, что плекс из плексигласа с разрезами, называемыми «керфами» и просверленными отверстиями, была более чем на 25% сильнее, чем один, с регулярно размещенными разрезами. [ 19 ] В отличие от тиранозавров и других теропод, древние хищники, такие как фитозавры и диметродон, не имели адаптации, чтобы предотвратить распространение зубьев их зубов, похожих на трещины. [ 19 ]
Посткраниальный скелет
[ редактировать ]
Череп был расположен в конце толстой, S -образной шеи, а длинный тяжелый хвост действовал как противовес, чтобы уравновесить голову и туловище, с центром масс над бедрами. Тиранозавриды известны своими пропорционально очень маленькими передниками с двумя пальцами, хотя рудимента . иногда встречаются остатки третьей цифры [ 10 ] [ 20 ] У Tarbosaurus были самые короткие передние конечности по сравнению с размером тела, в то время как Daspletosaurus имел самый длинный.

Тиранозавриды ходили исключительно по задним конечностям, поэтому их кости ноги были массивными. В отличие от передних конечностей, задние конечности были длиннее по сравнению с размером тела, чем почти любые другие тероподы. У несовершеннолетних и даже некоторых меньших взрослых, таких как более базальные тиранозавра, имели более длинные голени , чем бедра , характерная для качественных (быстро бегающих) динозавров, таких как орнитомимиды . У более крупных взрослых были пропорции ноги, характерные для медленных животных, но не в той степени, в которой можно увидеть другие крупные тероподы, такие как абелизавриды или карнозавры . Третьи плюсневые кости тиранозавридов были ущипнуты между вторым и четвертым плюсновиками, образуя структуру, известную как Arctometatarsus . [ 10 ] У тиранозавридов также были большие прикрепления мышц ног и низкую вращательную инерцию по сравнению с массой их тела, что указывает на то, что они могут повернуться быстрее, чем другие крупные тероподы. [ 21 ] [ 22 ]
Неясно, когда Arctometatarsus впервые развивался; Это не присутствовало в самых ранних тиранозавроидах, таких как Dilong , [ 23 ] но был найден в более позднем Аппалачиозавра . [ 16 ] Эта структура также охарактеризовала Troodontids , Ornithomimids и Caenagnathids , [ 24 ] Но его отсутствие в самых ранних тиранозавроидах указывает на то, что оно было получено путем конвергентной эволюции . [ 23 ]
Реконструкция мягких тканей
[ редактировать ]Сравнительный анализ зубов предполагает, что тиранозавриды, а также другие большие тероподы имели губы, которые защищали их зубы от внешнего повреждения. Эта анатомическая особенность дала этим животным больше визуального сходства с лепидозаврами, чем с тесно связанными крокодильцами. [ 25 ]
Классификация
[ редактировать ]Название Deinodontidae было придумано Эдвардом Дрибцом Коуп в 1866 году для этой семьи, [ 26 ] и продолжал использоваться вместо нового названия Tyrannososauridae до 1960 -х годов. [ 27 ] Типовым родом Deinodontidae является Deinodon , который был назван в честь изолированных зубов из Монтаны . [ 28 ] Однако в обзоре североамериканских тиранозавров в 1970 году Дейл Рассел пришел к выводу, что Деинодон не был действительным таксоном, и использовал имя Tyrannosauridae вместо Deinodontidae, заявив, что это соответствовало правилам ICZN . [ 12 ] Следовательно, Tyrannososauridae предпочитают современные эксперты. [ 5 ]
Tyrannosaurus был назван Генри Фэрфилдом Осборном в 1905 году вместе с семейством Tyrannosauridae. [ 29 ] Имя происходит от древнегреческих слов τυραννος ( тиран ) ('тиран') и σαυρος ( sauros ) ('ящерица'). Очень распространенный суффикс -idae обычно добавляется к зоологическим фамилиям и получен из греческого суффикса -ιΔαι -Idai , который указывает на существительное во множественном числе. [ 30 ]
Таксономия
[ редактировать ]
Tyrannosauridae-это семейство в рангевой таксономии Linnaean , в пределах суперсемейства Tyrannosauroidea и подрядной Theropoda .
Tyrannososauridae непревзойденно разделяется на две подсемейства. Albertosaurinae включает в себя североамериканские роды Albertosaurus и Gorgosaurus , в то время как Tyrannosaurinae включает Daspletosaurus , Teratophoneus , Bistabersor , Tarbosaurus , Nanuqsaurus , Zhuchengtyrannus и самих тиранозавра . [ 31 ] Некоторые авторы включают вида Gorgosaurus Libratus в роде Albertosaurus и Tarbosaurus Bataar в роду Tyrannosaurus , [ 16 ] [ 5 ] [ 32 ] в то время как другие предпочитают сохранять Gorgosaurus и Tarbosaurus в качестве отдельных родов. [ 10 ] [ 11 ] Альбертозавра характеризуются более тонкими сборками, более низкими черепами и пропорционально более длинными голенями , чем тиранозавра. [ 10 ] В тиранозаврах сагиттальный гребень на теменных велах продолжается вперед на лобовые. [ 11 ] В 2014 году Lü Junchang et al. Описал алиормин как племя в тиранозавре, содержащих алиорамуса и Qanzhoususaurus роды Их филогенетический анализ показал, что племя находилось у основания тиранозавра. [ 33 ] [ 34 ] Некоторые авторы, такие как Джордж Ольшевский и Трейси Форд, создали другие подразделения или племена для различных комбинаций тираннозавридов в подсемействах. [ 35 ] [ 36 ] Тем не менее, они не были филогенетически определены и обычно состояли из родов, которые в настоящее время считаются синонимом других родов или видов. [ 18 ]
Дополнительные подсемейства были названы в честь более фрагментарных родов, включая Aublysodontinae и Deinodontinae . Тем не менее, роды aublysodon и deinodon обычно считаются номинами Дубии , поэтому они и их одноименные подсемейства обычно исключаются из таксономий тираннозавридов. Дополнительный тиранозаврид, Raptorex , был первоначально описан как более примитивный тиранозавроид, но, вероятно, представляет собой ювенильный тираннозаврин, аналогичный Tarbosaurus . Однако, как это известно только из юношеского образца, в настоящее время он также считается номенем -дубимом . [ 37 ] Тем не менее, Томас Карр поддерживает свою достоверность и считает, что она похожа на тиранозавра. [ 38 ]
Филогения
[ редактировать ]С появлением филогенетической таксономии в палеонтологии позвоночных, Tyrannosauridae получили несколько явных определений. Оригинал был произведен Полом Серено в 1998 году и включал все тиранозавра ближе к Тираннозавру, чем к Alectrosaurus , Aublysodon или Nanotyrannus . [ 39 ] Тем не менее, нанотиранус часто считается юношеским тиранозавром Rex , в то время как Aublysodon обычно считается номенем -дубием, не подходящим для использования в определении клады . [ 10 ] Определения с тех пор основывались на более устоявшихся родах.
В 2001 году Томас Р. Хольц -младший опубликовал кладистический анализ Tyrannosauridae. [ 40 ] Он пришел к выводу, что было два подсемейства : более примитивные aublysodontinae , характеризующиеся несердинированными доносиловыми зубами ; и Tyrannosaurinae . [ 40 ] Aublysodontinae включал Aublysodon , « Kirtland Aublysodon » и Alectrosaurus . [ 40 ] Хольц также обнаружил, что Siamotyrannus демонстрировал некоторые синапоморфии Tyrannososauridae , но лежал «за пределами [ семьи ]». [ 40 ]
Позже в той же статье он предложил определить Tyrannosauridae как «все потомки самого последнего общего предка Tyrannosaurus и Aublysodon ». [ 40 ] Он также раскритиковал определения, ранее предложенные другими работниками, как, как предложенное Полом Серено , что Tyrannososauridae были «все таксоны ближе к« тиранозавру », чем к Alectrosaurus , Aublysodon и Nanotyrannus ». [ 40 ] Хольц заметил, что, поскольку нанотиран был, вероятно, ошибочно идентифицированным несовершеннолетним Т. Рексом , предлагаемое Серено было бы иметь семейный тиранозаурид в качестве субтаксона рода Tyrannosaurus . [ 40 ] Кроме того, его предлагаемое определение подсемейства Tyrannosaurinae также будет ограничено Tyrannosaurus . [ 40 ]
В 2003 году Кристофер Брочуу включает в себя Albertosaurus , Alectrosaurus , Alioramus , Daspletosaurus , Gorgosaurus , Tarbosaurus и Tyrannosaurus в определении. [ 41 ] Хольц пересмотрел кладу в 2004 году, чтобы использовать все вышеперечисленные в качестве спецификаторов, за исключением Алиорамуса и Альтрозавра , которые его анализ не мог с уверенностью достать. Однако в одной и той же статье Holtz также предоставил совершенно другое определение, включая все тероподы, более тесно связанные с тиранозавром, чем с эотираном . [ 10 ] Самым последним определением является определение Серено в 2005 году, которое определило Tyrannosauridae как наименее инклюзивная клада, содержащая Albertosaurus , Gorgosaurus и Tyrannosaurus . [ 42 ]
Кладистический тиранозавридов анализ филогения часто считает, что Tarbosaurus и Tyrannosaurus являются родственными таксонами , причем Daspletosaurus более базально, чем. Тесная связь между тарбозавром и тиранозавром подтверждается многочисленными особенностями черепа, включая схему швов между определенными костями, наличие гребня в форме полумесяца на посторбитальной кости за каждым глазом и очень глубокую верхнюю верхнюю верхнюю часть с заметной кривой вниз на Нижний край, среди прочих. [ 10 ] [ 16 ] Альтернативная гипотеза была представлена в исследовании Фила Керри и его коллег в 2003 году, в котором обнаружила слабую поддержку Daspletosaurus в качестве базального члена клады , включая Tarbosaurus и Alioramus , оба из Азии, на основе отсутствия костинного оборота, соединяющего носа и и Лесковые кости. [ 18 ] Было обнаружено, что Alioramus является наиболее близким родственником Tarbosaurus в этом исследовании, основанной на сходной схеме распределения напряжений в черепе.
Связанное исследование также отмечало механизм блокировки в нижней челюсти, разделенный между двумя родами. [ 43 ] В отдельной статье Керри отметила возможность того, что Алиорамус может представлять юношеский тарбозавр , но заявил, что гораздо более высокое количество зубов и более заметные носовые гребня у Алиорамуса предполагают, что это отличный род. Точно так же Керри использует высокое количество зубов Nanotyrannus, чтобы предположить, что это может быть отдельный род, [ 11 ] Вместо юношеского тиранозавра , как считают большинство других экспертов. [ 10 ] [ 44 ] Тем не менее, открытие и описание Qianzhousaurus показывают, что алиорамус не является тесным отношением к Tarbosaurus , вместо этого принадлежащий к новорожденному племени тираннозавридов; Алиоримини. Qianzhousaurus далее показывает, что аналогичные давно выпущенные тиранозавриды широко распространены по всей Азии и поделились бы той же средой, избегая при этом конкуренции с более крупными и более надежными тиранозаврами, охотясь за различной добычей. [ 45 ]
|
|
|
Палеобиология
[ редактировать ]Рост
[ редактировать ]
Палеонтолог Грегори Эриксон и его коллеги изучили рост и историю жизни тиранозавридов. Анализ гистологии кости может определить возраст образца, когда он умер. Темпы роста могут быть изучены, когда возраст различных людей наносится на график их размера на графике. Эриксон показал, что после долгого времени, как несовершеннолетние, тиранозавры подвергались огромным всплескам роста в течение примерно четырех лет в середине своей жизни. После того, как фаза быстрого роста закончился половой зрелостью , рост значительно замедлился у взрослых животных. Кривая роста тиранозавридов является S-образной формой, с максимальными темпами роста людей около 14 лет. [ 49 ]

По оценкам, самый маленький известный Tyrannosaurus rex индивидуум ( LACM 28471, «Иордан Теропод») весил всего 29,9 килограмма (66 фунтов) в возрасте всего 2 года, в то время как самый большой, такой как FMNH PR2081 (« Сью »), скорее всего, наибольшее весил около 5654 кг (12 465 фунтов), по оценкам, было 28 лет, возраст, который мог быть близок к максимуму для вида. [ 49 ] Несовершеннолетние Т. Рекса оставались менее чем в 1800 кг (4000 фунтов) до приблизительно 14 лет, когда размер тела начал резко увеличиваться. На этом быстром этапе роста молодой Т. Рекс получит в среднем 600 кг (1300 фунтов) в год в течение следующих четырех лет. Это замедлилось через 16 лет, и в 18 лет снова критические плато, что указывает на то, что рост резко замедлился. [ 50 ] Например, только 600 кг (1300 фунтов) отделили 28-летнюю «Сью» от 22-летнего канадского образца ( RTMP 81.12.1). [ 49 ] Это внезапное изменение скорости роста может указывать на физическую зрелость, гипотеза, которая подтверждается открытием медуллярной ткани в бедре 18-летнего Т. Рекса из Монтаны ( MOR 1125, также известный как «B-Rex») Полем [ 51 ] Медуллярная ткань обнаруживается только у женщин-птиц во время овуляции, что указывает на то, что «B-Rex» был репродуктивного возраста. [ 52 ]
Другие тиранозавриды демонстрируют чрезвычайно сходные кривые роста, хотя с более низкими темпами роста, соответствующими их более низким размерам взрослых. [ 53 ] По сравнению с альбертозаврами Daspletosaurus показал более быстрые темпы роста в течение быстрого периода роста из -за его более высокого веса взрослого. Максимальная темпа роста в Daspletosaurus составляла 180 килограммов (400 фунтов) в год, основываясь на массовой оценке 1800 кг (4000 фунтов) у взрослых. Другие авторы предложили более высокие веса взрослых для Daspletosaurus ; Это изменит величину скорости роста, но не общую модель. [ 49 ] Самым молодым известным Albertosaurus является двухлетний ребенок, обнаруженный в кости сухого острова, который весил бы около 50 кг (110 фунтов) и измерял немного более 2 метров (6,6 фута) в длину. 10-метровый (33-футовый) образец из того же карьера является самым старым и крупнейшим известным в возрасте 28 лет. Самый быстрый темп роста, по оценкам, возникает около 12–16 лет, достигая 122 кг (269 фунтов) в год, на основе взрослых 1300 кг (2900 фунтов), что составляет около пятого из скорости для T.-rex Полем Для Gorgosaurus рассчитанная максимальная скорость роста составляет около 110 килограммов (240 фунтов) во время фазы быстрого роста, что сопоставимо с тем, что у Albertosaurus . [ 49 ]
Обнаружение эмбрионального тиранозавра того, пока не упреждающего рода предполагает, что тиранозавриды разработали свои отличительные скелетные особенности при развитии в яйце. Кроме того, размер образца, 1,1 дюйма (2,8 см) из нижней челюсти, обнаруженной в двух лекарственном образовании Монтаны в 1983 году, и когтя ног, обнаруженный в формировании каньона Подкова в 2018 году и описанный в 2020 году, предполагает, что новорожденный Тиранозавриды родились с черепами размером с мыши или грызунов аналогичного размера и, возможно, были примерно размером с маленькую собаку при рождении. Считается, что образец челюсти был получен от животного примерно 2,5 фута (0,76 м), в то время как когтя, как полагают, принадлежит образцу, размещенному около 3 футов (0,91 м). В то время как яичные скорлупы не были обнаружены в связи с одним образцом, место, где были обнаружены эти новорожденные тиранозавриды, предполагает, что эти животные использовали те же места гнезда, что и другие виды, с которыми они жили, и охотились. [ 54 ] Отсутствие яичных скорлупы, связанных с этими образцами, также открыло предположения о возможности того, что тиранозавриды откладывают яйца с мягкими оболочкой в качестве яиц в родах муссура и протокератопов . [ 55 ]
Отопаемые следы из формирования вапити предполагают, что по мере роста тиранозавридов ноги стали шире с более толстыми пальцами, чтобы поддерживать их вес. Более широкие ноги предполагают, что взрослые тиранозавриды были медленнее, чем их потомство. [ 56 ] [ 57 ]
История жизни
[ редактировать ]
Конец фазы быстрого роста предполагает возникновение половой зрелости у Альбертозавра , хотя рост продолжался с медленной скоростью на протяжении всей жизни животных. [ 49 ] [ 53 ] Сексуальное созревание, в то время как все еще активно расти, по -видимому, является общей чертой среди маленьких [ 58 ] и большой [ 59 ] динозавры, а также у крупных млекопитающих, таких как люди и слоны . [ 59 ] Эта схема относительно раннего полового созревания поразительно отличается от схемы птиц, что задерживает их сексуальную зрелость до тех пор, пока они не закончат расти. [ 59 ] [ 60 ]
Таблица с числом образцов каждой возрастной группы, Эриксон и его коллеги смогли сделать выводы о истории жизни в популяциях Tyranosauridae. Их анализ показал, что в то время как несовершеннолетние были редки в ископаемом отчете, субдумы на фазе быстрого роста и взрослые были гораздо чаще. Более половины известных образцов T. rex , по-видимому, умерли в течение шести лет после достижения сексуальной зрелости, схемы, которая также наблюдается у других тираннозавров и сегодня у некоторых крупных, долгоживущих птиц и млекопитающих. Эти виды характеризуются высокими показателями младенческой смертности, а затем относительно низкая смертность среди несовершеннолетних. Смертность снова увеличивается после половой зрелости, отчасти из -за стрессов размножения. Хотя это может быть связано с сохранением или смещениями сбора , Эриксон предположил, что разница была связана с низкой смертностью среди несовершеннолетних в определенном размере, что также наблюдается у некоторых современных крупных млекопитающих, таких как слоны . Эта низкая смертность, возможно, была вызвана отсутствием хищничества, поскольку тиранозавры превзошли всех современных хищников в возрасте до двух лет. Палеонтологи не нашли достаточно Daspletosaurus остается для аналогичного анализа, но Эриксон отмечает, что, по -видимому, применяется та же общая тенденция. [ 53 ]
Тиранозавриды провели целую половину своей жизни в юношеской фазе, прежде чем вылететь до почти максимума всего за несколько лет. [ 49 ] Это, наряду с полным отсутствием хищников, промежуточных по размеру между огромными взрослыми тиранозавридами и другими небольшими тероподами, предполагает, что эти ниши могли быть заполнены ювенильными тиранозавридами. Это видно у современных драконов Комодо , где мысли начинаются как насекомоядные, жилые деревья , и медленно разразились в массивные хищники вершины, способные снимать больших позвоночных. [ 10 ] Например, Albertosaurus были обнаружены в агрегациях, которые, как предполагают, представляют упаковки смешанного возраста . [ 61 ] [ 62 ]
Локомоция
[ редактировать ]Возможности локомоции лучше всего изучаются для Tyrannosaurus , и есть две основные проблемы, касающиеся этого: насколько хорошо это может стать; и какова была максимальная прямая скорость, вероятно, была. Тиранозавр, возможно, медленно повернулся, возможно, потребовалось от одной-двух секунд, чтобы повернуть только 45 °-количество людей, которые были вертикально ориентированы и без хвоста, могут вращаться через часть секунды. [ 63 ] Причиной сложности является ротационная инерция , поскольку большая часть массы Тиранозавра . была на некотором расстоянии от центра тяжести, как человек, несущий тяжелую древесину [ 64 ]
Ученые произвели широкий диапазон максимальных скоростей, в основном около 11 метров в секунду (25 миль в час), но на 5–11 метров в секунду (11–25 миль в час), и несколько до 20 метров на один Второй (45 миль в час). Исследователи должны полагаться на различные методы оценки, потому что, хотя существует множество треков очень больших ходьбы теропод, до сих пор ни один из них не был обнаружен из очень больших теропод, и это отсутствие может указывать на то, что они не работали. [ 65 ]

Джек Хорнер и Дон Лесс в 1993 году утверждали, что Тираннозавр был медленным и, вероятно, не мог бежать (без воздушного фазы в середине). [ 66 ] Тем не менее, Холц (1998) пришел к выводу, что тиранозавриды и их близкие родственники были самыми быстрыми большими тероподами. [ 67 ] Кристиансен (1998) подсчитал, что кости ног тиранозавра не были значительно сильнее, чем колы Было около 11 метров в секунду (25 миль в час), что касается скорости человеческого спринтера. [ 68 ] Фарлоу и его коллеги (1995) утверждают, что тиранозавр от 6 до 8 тонн был бы критически или даже смертельно ранен, если бы он упал быстро, поскольку его торс врезался бы в землю при замедлении 6 г (шесть Время ускорения из -за гравитации или около 60 метров/с 2 ) и его крошечные руки не могли уменьшить воздействие. [ 69 ] [ 70 ] Однако, , жирафы скакают со скоростью 50 км/ч (31 миль в час), несмотря на риск того, что они могут сломать ногу или хуже, что может быть фатальным даже в «безопасной» среде, такой как зоопарк. как известно [ 71 ] [ 72 ] Таким образом, вполне возможно, что Тираннозавр также двигался быстро, когда это необходимо, и должен был принять такие риски; Этот сценарий также был изучен для аллозавра . [ 73 ] [ 74 ] Самое последнее исследование по локомоции Tyrannosaurus не сужает скорости дальше, чем диапазон от 17 до 40 км/ч (от 11 до 25 миль в час), т.е. от ходьбы или медленного бега до бега с умеренной скоростью. [ 65 ] [ 75 ] [ 76 ] В исследование компьютерной модели в 2007 году оценивалась скорость бега, основываясь на данных, взятых непосредственно из окаменелостей, и утверждалось, что T. rex обладает максимальной скоростью бега 8 метров в секунду (18 миль в час). [ 77 ] [ 78 ] (Вероятно, несовершеннолетний человек. [ 79 ] )
Исследования, проведенные Eric Snively et al., Опубликованные в 2019 году, показывают, что тиранозавриды, такие как Tarbosaurus и сами тиранозавра, были более маневренными, чем аллозавра сопоставимого размера, из -за низкой инерции вращения по сравнению с массой их тела в сочетании с большими мышцами ног. В результате предполагается, что тираннозавриды были способны делать относительно быстрые повороты и, вероятно, могли бы быстрее разворачиваться их тела, когда близко к их добыче или что во время поворота они могли «пируэт» на одной посаженной ноге, пока чередующая нога была продерживался приостановленным качанием во время преследования. Результаты этого исследования потенциально могут пролить свет на то, как ловкость могла бы способствовать успеху эволюции тиранозавридов. [ 80 ]
Кроме того, исследование 2020 года показывает, что тиранозавриды были исключительно эффективными ходунками. Исследования Dececchi et al. сравнивали пропорции ноги, массу тела и походки более 70 видов динозавров теропод, включая тиранозавридов. Затем исследовательская группа применила множество методов для оценки максимальной скорости каждого динозавра при беге, а также того, сколько энергии каждый динозавр потратил при перемещении на более расслабленных скоростях, таких как при ходьбе. Среди видов меньшего до среднего размера, таких как Dromaeosaurids, более длинные ноги, по-видимому, являются адаптацией для более быстрого бега, в соответствии с предыдущими результатами других исследователей. Но для теропод весом более 1000 кг (2200 фунтов), верхняя скорость бега ограничена размером тела, поэтому было обнаружено, что более длинные ноги коррелировали с ходьбой с низкой энергией. Результаты исследования также показали, что меньшие тероподы эволюционировали длинные ноги для скорости в качестве средства для помощи как в охоте, так и в сбежек от больших хищников, в то время как более крупные хищные тероподы, которые развивали длинные ноги, сделали это, чтобы снизить затраты на энергию и повышение эффективности кормления, поскольку они, как они были освобождены от требований давления хищничества из -за их роли в качестве хищников вершины. По сравнению с более базальными группами теропод в исследовании, тиранозавриды показали заметное повышение эффективности кормления из -за снижения расходов на энергию во время охоты и похвалы. Это, вероятно, привело к тому, что у тиранозавров была снижена потребность в охотничьях и потребовало меньшего количества пищи, чтобы поддерживать себя в результате. Кроме того, исследование в сочетании с исследованиями, которые показывают, что тиранозавры были более проворными, чем другие тероподы с крупным телом, указывает на то, что они были вполне хорошо адаптированы к подходу к дальней дистанции, за которым следует быстрый взрыв скорости, чтобы пойти на убийство. В результате можно отметить аналогии между тиранозавридами и современными волками, подтверждаемые доказательствами того, что, по крайней мере, некоторые тиранозавриды, такие как Альбертозавр охотился в групповых условиях. [ 81 ] [ 82 ]
Покров
[ редактировать ]
Продолжающиеся дебаты в палеонтологическом сообществе окружают степень и природу интенсивного покрытия тиранозавридов. Давние нитевидные структуры сохранились вместе со скелетными останками многочисленных коелурозавров из ранней меловой формирования Йиксиан и других близлежащих геологических формирований из Ляонинг , Китай. [ 83 ] Эти нити обычно интерпретируются как «протофичики», гомологичные с разветвленными перьями, обнаруженными у птиц, и некоторых неавийских теропод , [ 84 ] [ 85 ] Хотя другие гипотезы были предложены. [ 86 ] Скелет Dilong был описан в 2004 году, который включал первый пример «протофичи» в тиранозавра. Как и вниз по перьям современных птиц, «протофичи», найденные в Dilong, были разветвлены, но не выпечками , и, возможно, использовались для изоляции . [ 23 ] Обнаружение и описание 9-метрового (30 футов) пернатых тиранозавра Ютирана в 2012 году указывают на то, что вероятность крупных тираннозавридов также пернасодится в качестве взрослых. [ 87 ]

Основываясь на принципе филогенетического кронштейна , было предсказано, что тиранозавриды также могут обладать такими перьями. Тем не менее, исследование в 2017 году, опубликованное командой исследователей в биологических письмах, описано в тиранозавридных впечатлениях, собранных в Альберте, Монтане и Монголии, которые происходили из пяти родов ( Tyrannosaurus , Albertosaurus , Gorgosaurus , Daspletosaurus и Tarbosaurus ). [ 88 ] Хотя впечатления от кожи небольшие, они широко рассеяются по посткраниуму, все вместе расположены на животе, торакальной области, подвздошной кости, таза, хвоста и шеи. Они показывают жесткую схему прекрасных, непересекающихся катлевых масштабов (которые соавтор Скотт по сравнению с теми, которые наблюдаются на флангах крокодила [ 89 ] ) и не сохранять никаких намеков на перья. Основная текстура состоит из крошечных «фундаментальных масштабов» диаметром приблизительно от 1 до 2 мм, с некоторыми впечатлениями, показывающими 7 мм «шкалы функций» между ними. Дополнительные масштабы можно увидеть на следах Тиранозаврида. [ 90 ] Исследования показывают, что покров личиков тиранозавридов имел масштабы на зубной и верхней челюсти, корнифицированный эпидермис и броня, похожая на кожу на подчиненных областях. [ 91 ] [ 92 ]
Bell et al. выполнил реконструкцию наследственного персонажа, основанную на том, что известно о распределении покровных по интуированию у тиранозавра. Несмотря на 89% вероятность того, что тиранозавра начались с перьев, они определили, что чешуйчатые тиранозавриды имеют 97% вероятность быть истинной. Данные «предоставляют убедительные доказательства совершенно плоскоклеточного покрытия в Тираннозавре», написала команда, хотя они признали, что оперение, возможно, все еще присутствовало в дорсальном регионе, где кожные впечатления еще не были найдены. Bell et al. Гипотезы определяют, что масштабные впечатления от тираннозавридов, возможно, являются ретикулями, которые вторично получены из перьев, хотя для этого необходимы доказательства. [ 88 ] Однако другие утверждают, что это связано с тафономическим уклоном у тираннозавридов. [ 93 ]
Еще предстоит определить, почему такое покровное изменение могло произойти. Прецедент для потери перья можно увидеть в других группах динозавров, таких как орнитишки , в которых нитевидные структуры были потеряны, и вновь появились весы. [ 94 ] Хотя гигантизм был предложен в качестве механизма, Фил Р. Белл, который соавторил исследование, отметил, что пернатый Ютиранус перекрыл по размеру с Gorgosaurus и Albertosaurus . «Проблема здесь в том, что у нас есть большие тиранозавры, некоторые с перьями, некоторые без этого живут в довольно похожих климатах. Так в чем причина этой разницы? Мы действительно не знаем». [ 95 ]
Зрение
[ редактировать ]
Глаза тиранозавра расположены так, что глаза будут указывать вперед, давая им бинокулярное зрение, немного лучше, чем у современных ястребов . В то время как хищные тероподы в целом имели бинокулярное зрение непосредственно перед черепом, у тиранозавров была значительно большая площадь перекрытия. Джек Хорнер также отметил, что у линии тиранозавра была история постоянно улучшающего бинокль. Трудно понять, как естественный отбор предположил бы эту долгосрочную тенденцию, если бы тиранозавры были чистыми мусорщиками, что не потребовало бы развитого восприятия глубины , которое обеспечивает стереоскопическое зрение . [ 96 ] [ 97 ] У современных животных бинокулярное зрение встречается в основном у хищников (основными исключениями являются приматы , которые нуждаются в том, чтобы перейти из ветви к ветвью). В отличие от Tyrannosaurus , у Tarbosaurus был более узкий череп, более типичный для других тиранозавридов, в которых глаза сталкиваются в основном вбок. Все это говорит о том, что Tarbosaurus больше полагался на чувства запаха и слуха, чем на зрение. [ 98 ] В Gorgosaurus образцах глазной розетка была круглой, а не овальной или в форме замочной скважины, как в других родах тиранозаврида. [ 11 ] В Daspletosaurus это был высокий овал, где -то между круговой формой, видимой в Gorgosaurus и форме «замочной скважины» тиранозавра . [ 10 ] [ 11 ] [ 44 ]
Чувствительность лица
[ редактировать ]Основываясь на сравнении кости дасплетозавра текстуры с существующими крокодильцами , подробное исследование в 2017 году Томаса Д. Карр и соавт. обнаружил, что у тиранозавров были большие плоские весы на своих мордах . [ 99 ] [ 100 ] В центре этих весов были небольшие кератинизированные пятна. У крокодилов такие пятна покрывают пучки сенсорных нейронов , которые могут обнаруживать механические, тепловые и химические стимулы . [ 101 ] [ 102 ] Они предположили, что у тиранозавров , вероятно, также были пучки сенсорных нейронов лица под их масштабами и, возможно, использовали их для идентификации объектов, измерения температуры их гнезд и осторожно подбирают яйца и мысли . [ 99 ]
Костные гребни
[ редактировать ]
Костровая гребня встречаются на черепах многих теропод, в том числе много тираннозавридов. Алиорамус , возможный тираннозаврид из Монголии, несет один ряд из пяти выдающихся костных ударов на носовых костях; Аналогичный ряд гораздо более низких ударов присутствует на черепе Аппалачиозавра , а также некоторые образцы Daspletosaurus , Albertosaurus и Tarbosaurus . [ 16 ] В Albertosaurus , Gorgosaurus и Daspletosaurus перед каждым глазом на слезной кости есть выдающийся рог. Лескимальный рог отсутствует в Tarbosaurus и Tyrannosaurus , которые вместо этого имеют герб в форме полумесяца за каждым глазом на посторбитальной кости . Эти гребни, возможно, использовались для демонстрации , возможно, для распознавания видов или поведения ухаживания . [ 10 ]
Терморегуляция
[ редактировать ]Тиранозавр , как и большинство динозавров, долгое время считалось, что он имел эктотермический («хладнокровный») метаболизм были брошены такие ученые, как Роберт Т. Баккер и Джон Остром рептилий, но в первые годы « Ренессанса динозавров » 1960 -е годы [ 103 ] [ 104 ] Сам Tyrannosaurus rex , как утверждалось, был эндотермическим («теплокровным»), что подразумевает очень активный образ жизни. [ 105 ] С тех пор несколько палеонтологов стремились определить способность регулировать температуру тиранозавра тела своего . Гистологические доказательства высоких темпов роста у молодых Т. Рекса , сравнимых с тем млекопитающими и птицами, могут подтвердить гипотезу высокого метаболизма. Кривые роста показывают, что, как и у млекопитающих и птиц, рост T. rex был ограничен в основном незрелыми животными, а не неопределенным ростом, наблюдаемым у большинства других позвоночных . [ 50 ] Было указано, что разница температуры могла быть не более 4-5 ° C (от 7 до 9 ° F) между позвонками торса и голени нижней ноги. Этот небольшой температурный диапазон между ядром тела и конечностями был заявлен палеонтологом Ризом Барриком и геохимиком Уильямом, чтобы указать, что Т. Рекс сохранил постоянную внутреннюю температуру тела ( гомеотермию ) и что он пользовался метаболизмом где -то между эктотермическими рептилиями и эндотермическими млекопитающими. [ 106 ] Позже они обнаружили аналогичные результаты в образцах Giganotosaurus , которые жили на другом континенте и десятках миллионов лет назад. [ 107 ] Даже если Tyrannosaurus rex действительно демонстрирует доказательства гомеотермии, это не обязательно означает, что это было эндотермическим. Такая терморегуляция также может быть объяснена гигантотермией , как у некоторых живых морских черепах . [ 108 ] [ 109 ] [ 110 ]
Палеоэкология
[ редактировать ]Сосуществование дамплетозавра и горгозавра
[ редактировать ]
В формировании парка Динозавров Горгозавр жил рядом с более редкими видами тиранозаврина Daspletosaurus . Это один из немногих примеров двух сосуществующих родов тиранозавра. Подобные размеры хищников в современных гильдиях хищников разделены на различные экологические ниши анатомическими, поведенческими или географическими различиями, которые ограничивают конкуренцию. Нишевая дифференциация между тиранозавридами парка динозавров не совсем понятна. [ 111 ] В 1970 году Дейл Рассел предположил , что более распространенный Gorgosaurus активно охотился на флота-ногах , в то время как реже и более неприятные цератопсианцы и анкилозавры (рогатые и сильно бронированные динозавры) остались для более сильно построенных Daspletosaurus . [ 12 ] Тем не менее, образец Daspletosaurus ( OTM 200) из современного двух лекарств в Монтане сохраняет перевариваемые останки ювенильного адрозавра в его области кишечника. [ 112 ] В отличие от некоторых других групп динозавров, ни один род не был более распространенным на более высоких или более низких высотах, чем другие. [ 111 ] Тем не менее, Gorgosaurus появляется чаще в северных формациях, таких как парк динозавров, с видами дамплетозавра, более распространенными на юге. Тот же шаблон виден в других группах динозавров. Chasmosaurine Ceratopsians и Hadrosaurine Hadrosaurs также чаще встречаются в двух лекарственных формах Монтаны и на юго -западе Северной Америки во время кампании, в то время как центрозарины и ламбеозавра доминируют в северных широтах. Хольц предположил, что этот паттерн указывает на общие экологические предпочтения между тиранозаврами, хасмосавринами и гадрозауринами. В конце более поздней маастрихтской сцены тиранозавра, такие как Tyrannosaurus rex , Hadrosaurines, такие как Edmontosaurus и Chasmosaurines, такие как трицератопс, были широко распространены по всей Западной Северной Америке, в то время как альбертозавра и центрозавра вымерли, а ламбейозавра были редкими. [ 10 ]
Социальное поведение
[ редактировать ]Существуют ограниченные доказательства социального поведения среди тиранозавридов. Исследователи сообщили, что в одном и том же карьере были обнаружены юношеский скелет и юношеский скелет, который был использован для поддержки гипотезы о том, что тираннозавры могли жить в каких -то социальных группах. [ 113 ] Хотя нет никаких доказательств общительного поведения в Gorgosaurus , [ 61 ] [ 62 ] Существуют свидетельства некоторого поведения упаковки для Albertosaurus и Daspletosaurus .
Молодой экземпляр видов Daspletosaurus Daspletosaurus ( TMP 94.143.1) показывает следы укусов на лице, которые были нанесены другим тиранозавром. Метки укуса исцеляются, что указывает на то, что животное пережило укус. Полбороченный парк динозавров Daspletosaurus (TMP 85.62.1) также демонстрирует следы укуса тиранозавра, показывая, что атаки на лицо не были ограничены молодыми животными. Хотя возможно, что укусы были связаны с другими видами, внутривидовая агрессия, включая укусы лица, очень распространена среди хищников. Укусы лица видны у других тиранозавров, таких как Gorgosaurus и Tyrannosaurus , а также в других родах Theropod, таких как Sinraptor и Saurornitholestes . Даррен Танк и Фил Керри предполагают, что укусы связаны с внутривидовой конкуренцией за территорию или ресурсы или доминирование в социальной группе. [ 61 ]

Доказательства того, что Daspletosaurus жил в социальных группах, исходит из кости, обнаруженной в двух лекарственном образовании Монтаны. Кости включает остатки трех дамплетозавра , в том числе большого взрослого, небольшого юношеского и другого индивидуума промежуточного размера. По крайней мере пять хадрозавров сохранились в том же месте. Геологические данные указывают на то, что останки были объединены не речными токами , а что все животные были похоронены одновременно в том же месте. Останки Хадразавра разбросаны и несут много отметок от зубов тиранозавра, что указывает на то, что дасплетозавр питался хадрозаврами во время смерти. Причина смерти неизвестна. Керри предполагает, что дамплетозавры образовали пакет , хотя это нельзя указать с уверенностью. [ 62 ] Другие ученые скептически относятся к доказательствам социальных групп в Daspletosaurus и других крупных тероподах; [ 114 ] Брайан Роуч и Даниэль Бринкман предположили, что социальное взаимодействие Daspletosaurus более близко напоминало бы современного дракона Комодо , где не-кооперативные люди туши толпы, часто атакующие и даже каннибалирующие друг друга в процессе. [ 115 ]
Сухой островной костяной клет, обнаруженная Барнумом Брауном, и его команда содержит останки 22 Альбертозавра , большинство людей, найденных в одном местном месте любого мела-теропода, и второй по величине любого крупного динозавра с динозавром Кливленда -Ллоид Карьер в Юте . Группа, кажется, состоит из одного очень старого взрослого; восемь взрослых от 17 до 23 лет; Семь подразделений проходят свои быстрые этапы роста в возрасте от 12 до 16 лет; и шесть несовершеннолетних в возрасте от 2 до 11 лет, которые еще не достигли фазы роста. [ 53 ] Почти поглощение осталось травоядного животного и аналогичное состояние сохранения между многими людьми в карьере Albertosaurus Bonebede заставило Фила Керри прийти к выводу, что местность не была хищной ловушкой, как La Brea Lat в Калифорнии , и что все сохранившиеся Животные умерли одновременно. Керри утверждает, что это доказательство поведения упаковки. [ 116 ] Другие ученые скептически относятся к тому, что животные могли быть объединены засухой, наводнением или по другим причинам. [ 53 ] [ 114 ] [ 117 ]
Хотя это обычно остается спорным, существуют доказательства, которые подтверждают теорию, что, по крайней мере, некоторые тиранозавриды были социальными. В Британской Колумбии в формировании Wapiti , трек , состоящий из следов трех отдельных тиранозавридов (названных как Ichnogenus bellatoripes Fredlundi ), был обнаружен местным экипировкой по имени Aaron Fredlund и описан в журнале Plos One Richard McCrea et al. Исследование Trackway не обнаружило никаких доказательств того, что один трек -эй был оставлен долго после того, как был сделан другой, что еще больше подтверждает гипотезу о том, что три отдельных тираннозавры путешествовали вместе как группа. Дальнейшие исследования показали, что животные путешествовали со скоростью от 3,9 до 5,2 мили в час (6,3 и 8,4 км/ч) и, вероятно, имели высоту бедра от 7 до 9 футов. В качестве трех разных родов тиранозавридов ( Gorgosaurus , Daspletosaurus и Albertosaurus , соответственно) известны из формирования, неизвестно, какой род был создатель Trackway. [ 118 ] [ 119 ] [ 120 ] Дополнительные доказательства в форме кости от карьеры радуг и единорогов в формировании кайпаровитса в южной части штата Юта, описанная в 2021 году, приписываемой Teratophoneus, предполагают, что другие тиранозавриды также были социальными животными. Окаменелости, состоящие из четырех или, возможно, пяти различных животных в возрасте от 4 до 22 лет, предполагают событие массовой смертности, возможно, вызванное наводнениями или менее вероятностью цианобактериальным токсикозом , пожаром или засухой. Тот факт, что все сохранившиеся животные, по -видимому, погибли в течение короткого срока, еще больше усиливает аргумент в пользу общительного поведения у тиранозавридов, при этом костные пласты таких родов, как тератофон , альбертозавр , тираннозавр и дампеттозавр, демонстрирующие, что социальное поведение могло быть распространенным среди распространения среди распространения среди распространенных среди распространенных поведений среди Tyrannosauridae в целом. [ 121 ] [ 122 ] [ 123 ]
Кормление
[ редактировать ]тиранозавра являются наиболее часто сохраняемыми следами кормления плотоядных динозавров. Зоны [ 124 ] Их сообщалось от цератопсийцев , хадрозавров и других тиранозавров. [ 124 ] Кости тиранозавридов с метками зубов представляют около 2% известных окаменелостей с сохраненными метками зубов. [ 124 ] Зубы тиранозавридов использовались в качестве удержания для вытягивания мяса с тела, а не для ножа, подобных функциям резки. [ 125 ] Схемы износа зубов намекают на то, что сложное поведение встряхивания головы, возможно, было вовлечено в кормление тиранозавра. [ 125 ]

Спекуляции о привычках охоты на упаковку Альбертозавра были сделаны несколькими исследователями, которые предполагают, что молодые члены пакета, возможно, были ответственны за то, чтобы двигать добычей к взрослым, которые были более крупными и мощными, но также медленными. [ 116 ] У несовершеннолетних также могли быть разные образ жизни, чем у взрослых, заполняя ниши хищников между огромными взрослыми и меньшими современными тероподами, самые большие из которых были на два порядка меньше, чем взрослый альбертозавр в массе. [ 10 ] Однако, поскольку сохранение поведения в ископаемом отчете чрезвычайно редко, эти идеи не могут быть легко проверены. Фил Керри предполагает, что Daspletosaurus образовал пакеты для охоты , хотя это нельзя с уверенностью указать. [ 62 ] Нет никаких доказательств такого общительного поведения в Gorgosaurus . [ 61 ] [ 62 ]
Дискуссия о том, ли Тираннозавр был хищником или чистым мусорщиком, так же старая, как и дебаты о его передвине. Ламбе (1917) описал хороший скелет Тираннозавра Gorgosaurus относительного близкого и пришел к выводу, что он и, следовательно, также тираннозавр был чистым мусорщиком, потому что Gorgosaurus зубы практически не проявляли износа. [ 126 ] Этот аргумент больше не воспринимается всерьез, потому что тероподы заменили их зубы довольно быстро. С тех пор, как первое открытие Tyrannosaurus Большинство ученых согласились с тем, что это был хищник, хотя, как и современные крупные хищники, было бы рад убить или украсть убийство другого хищника, если бы у него была возможность. [ 127 ] [ 128 ]
Отметил Hadrosaur эксперт Джек Хорнер в настоящее время главным сторонником идеи о том, что Tyrannosaurus был исключительно мусорщиком и вообще не участвовал в активной охоте. [ 66 ] [ 129 ] [ 130 ] Хорнер представил несколько аргументов, подтверждающих гипотезу чистого мусорщика. Присутствие больших обонятельных луковиц и обонятельных нервов предполагает высоко развитое обоняние для обнюхивания тушек на больших расстояниях. Зубы могут раздавить кость и, следовательно, могут извлечь как можно больше пищи ( костного мозга ) из остатков туши, обычно наименее питательных деталей. По крайней мере, часть его потенциальной добычи может двигаться быстро, в то время как данные свидетельствуют о том, что Тираннозавр гулял вместо бега. [ 129 ] [ 131 ]
Другие данные свидетельствуют о охотничьем поведении в Тираннозавре . Глаза тиранозавров расположены так, что глаза будут указывать вперед, давая им бинокулярное зрение, немного лучше, чем у современных ястребов . Ущерб, нанесенный тиранозавра, был обнаружен на скелетах хадрозавров и трицератоп, которые, казалось, пережили первоначальные атаки. [ 132 ] [ 133 ] [ 134 ] Некоторые исследователи утверждают, что если тиранозавр был мусорщиком, другой динозавр должен был быть лучшим хищником в верхнем мера америке. Лучшей добычей была более крупная края и орнитоподы . Другие тиранозавриды имеют так много характеристик с тиранозавром , что только мелкие дромаозавры остаются возможными лучшими хищниками. В этом свете приверженцы гипотезы мусорщика предположили, что размер и сила тираннозавров позволили им украсть убийства от меньших хищников. [ 131 ]

было сообщено о ювенильном Gorgosaurus с содержанием желудка in situ , содержащим два несовершеннолетних примерно года В 2023 году из парка динозавров . Этому несовершеннолетнему было бы 5-7 лет на момент смерти, имея длину около 4 метров (13 футов) и весом около 335 килограммов (739 фунтов). Он намного больше, чем у двух несовершеннолетних Citipes , которые весят около 9–12 килограммов (20–26 фунтов), в отличие от предположения, что тираннозавриды, получавшие добычу их размера, когда они достигли 16–32 килограммов (35–71 фунт), что указывает на что несовершеннолетние тиранозавриды по -прежнему потребляли гораздо меньшую добычу после превышения порога определенного размера. Обнаружение этого образца показало, что тиранозавриды, вероятно, не охотятся в многопользовательских пакетах, поскольку его размер добычи слишком мал, чтобы делиться с конспецификами. Это также прямые диетические доказательства, подтверждающие теорию «онтогенетического сдвига» для тиранозавридов, как ранее было выведено в результате экологического моделирования и анатомических особенностей среди разных возрастных групп. Только остатки задних конечностей и каудальных позвонков юношеских Ситипы присутствовали в полости желудка тиранозаврида, что позволяет предположить, что у юношеского Gorgosaurus было преимущественное потребление мышечных задних конечностей. [ 135 ]
Каннибализм
[ редактировать ]Доказательства также убедительно свидетельствуют о том, что тиранозавриды, по крайней мере, иногда каннибалистичны. Сам Тиранозавр имеет убедительные доказательства, указывающие на него, как каннибалистическую, по крайней мере, в способности по удалению, основанной на знаках зубов на костях стопы, плечевой кости и плюсневых атмосферах одного образца. [ 136 ] Окаменелости из формирования фруктов , формации Киртланда (как кампаниана по возрасту), и маастихтиново-возрастка Оджо Аламо Формирование предполагают, что каннибализм присутствовал в различных родах тиранозаврида бассейна Сан-Хуана. Доказательства, полученные из образцов, предполагают оппортунистическое поведение питания у тираннозавридов, которые каннибализировали членов их собственного вида. [ 137 ]
Распределение
[ редактировать ]
Самые ранние известные останки Тиранозавра произошли между 129,4 и 125 миллионов лет назад в Ишикаве, Япония, недалеко от LAT 36.166668 СПГ 136,633331. Найденным Маршем в 1881 году и идентифицированный HF Osborne в 1906 году, возраст останков был определен H. Matsuoka et al. в 2002 году.
В то время как более ранние тиранозавра обнаружены на всех трех северных континентах, окаменелости тиранозавридов известны только из Северной Америки и Азии. Иногда фрагментарные останки, обнаруженные в южном полушарии, сообщались как «тиранозавриды южного полушария», хотя, по -видимому, они были неверно идентифицированными окаменелостями абелисавридов . [ 138 ]
Останки Тиранозаврида никогда не были обнаружены из Восточной Северной Америки, в то время как более базальные тиранозавра, такие как сухой и Аппалачозавр , сохранялись до конца меховой Америки, что указывает на то, что тиранозавриды должны были развиваться или рассеяны на западную северную Америку, указывая на пополам тиранозавриды Западным внутренним морским путем посреди мелового. [ 16 ] Окаменелости тиранозавридов были обнаружены на Аляске , что, возможно, обеспечило путь для рассеивания между Северной Америкой и Азией. [ 139 ] Обнаружено, что алиорамус и тарбозавр связаны в одном кладистическом анализе, образуя уникальную азиатскую отрасль семьи. [ 18 ] Позже это было опровергнуто с открытием Qianzhousaurus и описанием семейства Тираннозавров Алиорамина. Зубы тиранозавридов от большого вида неизвестного сорта были обнаружены на полуострове Нагасаки исследователями из музея префектурных динозавров Фукуи, что еще больше расширило диапазон группы. Зубы, по оценкам, в возрасте 81 миллиона лет ( возраст Кампании ). [ 140 ]

Из двух подсемейств тиранозавра, по -видимому, были более распространенными. домом были тиранозавра, такие как Tarbosaurus и Zhuchengtyrannus , а также Qianzhousaurus и Alioramus alioramini Альбертозарины неизвестны в Азии, в которой . Как подсемейства Tyrannososaurinae, так и Albertosaurinae присутствовали на кампании и ранних маастрихтских этапах Северной Америки, при этом тираннозарины, такие как Daspletosaurus, на протяжении всей западной внутренней части, в то время как Albertosaurines Albertosaurus и Gorgosaurus в настоящее время известны только из северо -западного. [ 141 ]
Покойным маастрихтином альбертозавра, похоже, вымерли, в то время как тиранозавриновый тираннозавр бродил из Саскачевана в Техас . Эта модель отражается в других таксонах динозавров в Северной Америке. Во время кампанианского и раннего маастрихта ламбейозавра гадрозавров и центрозавриновых цератопсийцев распространены на северо -западе, в то время как гадрозарины и хасмосарины были более распространены для юга. К концу мелоговых центрозавра неизвестны, и ламбейозавра редки, в то время как гасрозавра и хасмосарины были обычными во всем западном внутреннем интерьере. [ 10 ] Исследование, опубликованное в журнале Scientific Reports 2 февраля 2016 года, Стивом Брусатте, Томасом Карром и др . Указывает, что во время более позднего маастрихта сам тираннозавр мог бы быть частично ответственной за вымирание других тираннозавридов в большинстве западной части Северной Америки. Исследование показывает, что Тираннозавр мог быть иммигрантом из Азии, в отличие от развития в Северной Америке (возможно, потомком тесно связанного тарбозавра ), который вытеснил и преодолел другие тиранозавриды. Эта теория дополнительно подтверждается тем фактом, что в известном диапазоне Тиранозавра не обнаружены лишь лишь лишь иные другие виды тиранозавридов . [ 142 ]
Временная шкала родов
[ редактировать ]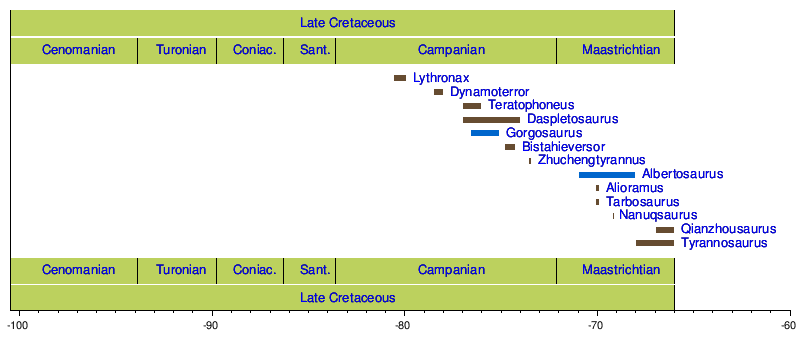
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Holtz, Thomas R. Jr. (2012) Динозавры: самая полная, современная энциклопедия для любителей динозавров всех возрастов, Приложение Winter 2011. Архивированный 2017-08-12 на The Wayback Machine
- ^ Статья о Скотти на osel.cz (в чешском)
- ^ Люди, SW; Керри, PJ; Эриксон, GM (2019). «Старый и исключительно большой взрослый образец Tyrannosaurus rex » . Анатомическая запись . 303 (4): 656–672. doi : 10.1002/ar.24118 . ISSN 1932-8486 . PMID 30897281 .
- ^ Лайл А. (22 марта 2019 г.). «Палеонтологи выявляют крупнейший тиранозавр , когда -либо обнаруженный» . Фолио, Университет Альберты . Получено 25 марта 2019 года .
- ^ Подпрыгнуть до: а беременный в дюймовый и Плотник, Кен. (1992). «Тираннозавриды (динозаврическая) Азии и Северная Америка». В Mateer, Niall J.; Чен Пейджи (ред.). Аспекты не -марной меловой геологии . Пекин: China Ocean Press. С. 250–268.
- ^ Breithaupt, BH; Саутвелл, эх; Мэтьюз, Н.А. (18 октября 2005 г.). «В рамках празднования 100 -летнего Tyrannosaurus rex : Manospondylus gigas , Ornithomimus grandis и Dynamosaurus Imperiosus , самые ранние открытия Tyrannosaurus rex на Западе» . Тезисы с программами . 2005 Ежегодное собрание Солт -Лейк -Сити . Тол. 37. Геологическое общество Америки . п. 406. Архивировано из оригинала 22 октября 2019 года . Получено 8 октября 2008 года .
- ^ Breithaup, BH; Southwell EH; Мэтьюз Н.А. (2006). « Империоз Dynamosaurus и самые ранние открытия Tyrannosaurus rex в Вайоминге и на западе». Музей естественной истории и науки Нью -Мексико . 35 : 257–258.
- ^ Bakker RT; Уильямс М.; Керри П. (1988). « Nanotyrannus , новый род Pygmy Tyrannosaur, из последнего мела Монтаны». Охотники . 1 : 1–30.
- ^ Подпрыгнуть до: а беременный Курзанов, Сергей М. «Новый карнозавр из покойного мела Ногон-цав, Монголия». Совместная советская монгольская палеонтологическая экспедиция (на русском языке). 3 : 93–104.
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий Хольц, Томас Р. (2004). "Tyrannosauroidea". В Weishampel, Дэвид Б .; Додсон, Питер ; Osmólska, Halszka (ред.). Динозаврия (второе изд.). Беркли: Университет Калифорнийской прессы. С. 111–136. ISBN 978-0-520-24209-8 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин Керри, Филипп Дж. (2003). «Черноальная анатомия тиранозавридов из покойного мела Альберты» (PDF) . Acta Palaeontologica Polonica . 48 (2): 191–226. Архивировано (PDF) из оригинала 2018-08-19 . Получено 2010-06-12 .
- ^ Подпрыгнуть до: а беременный в дюймовый Рассел, Дейл А. (1970). «Тираннозавры из покойного мела Западной Канады». Национальный музей естественных наук публикации в палеонтологии . 1 : 1–34.
- ^ Мортимер, Микки (21 июля 2003 г.). «И самый большой теропод -…» Список рассылки динозавров. Архивировано из оригинала 25 марта 2010 года . Получено 8 сентября 2007 года .
- ^ Статья о Скотти на osel.cz (в чешском)
- ^ Керри, Филипп Дж. (2000). «Тероподы из мелового из Монголии». Возраст динозавров в России и Монголии . Кембридж: издательство Кембриджского университета. С. 434–455. ISBN 978-0-521-54582-2 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон Карр, Томас Д.; Уильямсон, Томас Э.; Schwimmer, David R. (2005). «Новый род и виды тиранозавра из позднего мелового (среднего кампании) формирования демополиса Алабамы». Журнал палеонтологии позвоночных . 25 (1): 119–143. doi : 10.1671/0272-4634 (2005) 025 [0119: angaso] 2.0.co; 2 . ISSN 0272-4634 . S2CID 86243316 .
- ^ Карр, Томас Д.; Варриккио, Дэвид Дж.; Sedlmayr, Jayc C.; Робертс, Эрик М.; Мур, Джейсон Р. (2017-03-30). «Новый тираннозавр с доказательствами анагенеза и крокодилоподобной чувственной системы лица» . Научные отчеты . 7 (1): 44942. BIBCODE : 2017NATSR ... 744942C . doi : 10.1038/srep44942 . ISSN 2045-2322 . PMC 5372470 . PMID 28358353 .
- ^ Подпрыгнуть до: а беременный в дюймовый Керри, Филипп Дж .; Хурум, Джёрн Х; Сабат, Кароль. (2003). «Структура и эволюция черепа в филогении тиранозавридов» (PDF) . Acta Palaeontologica Polonica . 48 (2): 227–234. Архивировано (PDF) из оригинала 2008-10-31 . Получено 2010-10-02 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин Abler, WL 2001. Керф-и бурная модель зубных зубцов тиранозавра. п. 84–89. В: Жизнь мезозойских позвоночных . Ed.S Tanke, DH, Carpenter, K., Skrepnick, MW Indiana University Press.
- ^ Куинлан, Элизабет Д.; Дерстлер, Крайг; Миллер, Мерседес М. (2007). «Анатомия и функция цифр III тиранозавра Рекса Мануса» . Ежегодное собрание Геологического общества Америки-Тезисы с программами : 77. Архивировано из оригинала 2008-02-24 . Получено 2008-01-15 . [только аннотация]
- ^ Снаво, Эрик; О'Брайен, Хейли; Хендерсон, Дональд М.; Маллисон, Генрих; Сдача, Лара А.; Бернс, Майкл Э.; Хольц, Томас Р.; Рассел, Энтони П.; Витмер, Лоуренс М.; Керри, Филипп Дж.; Хартман, Скотт А.; Коттон, Джон Р. (2019-02-21). «Более низкая вращательная инерция и более крупные мышцы ног указывают на более быстрые повороты в тиранозавридах, чем в других крупных тероподах» . ПЕРЕЙ . 7 : E6432. doi : 10.7717/peerj.6432 . ISSN 2167-8359 . PMC 6387760 . PMID 30809441 .
- ^ Хартман, Скотт (2019-02-21). «Более низкая вращательная инерция и более крупные мышцы ног указывают на более быстрые повороты в тиранозавридах, чем в других крупных тероподах» . Скотт Хартман - доктор философии . Получено 2023-12-29 .
- ^ Подпрыгнуть до: а беременный в Сюй Син, х ; Norell, Mark A.; Куанг Сюэвен; Ван Ксаолин; Чжао Ци; Цзя Ченгкай. (2004). «Базальные тиранозаврауруроиды из Китая и доказательства протоферы у тиранозаврауроидов» (PDF ) Природа 431 (7009): 680–6 Bibcode : 2004natur.431..680x Doi : 10.1038/ nature0 PMID 15470426 S2CID 4381777
- ^ Хольц, Томас Р. (1994). «Филогенетическое положение Tyrannosauridae: последствия для системной теропод». Журнал палеонтологии . 68 (5): 1100–1117. Bibcode : 1994jpal ... 68.1100h . doi : 10.1017/s0022336000026706 . JSTOR 1306180 . S2CID 129684676 .
- ^ Томас М. Каллен, Дерек Уильям Ларсон, Марк П. Виттон, Дайан Скотт, Чай Махо, Кирстин С. Бринк, Дэвид С Эванс, Роберт Рейс (30 марта 2023 г.). «Реконструкция лицевого лицера Theropod и важность мягких тканей в палеобиологии» . Наука . 379 (6639): 1348–1352. Bibcode : 2023sci ... 379.1348c . doi : 10.1126/science.abo7877 . PMID 36996202 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Коуп Эд (1866). «Открытие гигантского динозавра в меловом периоде Нью -Джерси». Труды Академии естественных наук Филадельфии . 18 : 275–279.
- ^ Малев Э.А. (1955). «Новые плотоядные динозавры из верхней меловой Монголии». Doklady Akademii Nauk Sssr . 104 (5): 779–783.
- ^ Лейди, Джозеф (1856). «Уведомление о остатках вымерших рептилий и рыб, обнаруженных доктором Ф. Ф. Хейденом в бесплодных землях реки Джудит, территория штата Небраска». Труды Академии естественных наук Филадельфии . 8 : 72–73.
- ^ Осборн, Генри Ф. (1905). « Тиранозавр и другие меловые хищные динозавры». Бюллетень Американского музея естественной истории . 21 (3): 259–265. doi : 10.1111/j.1468-5965.2007.00735_17.x . HDL : 2246/1464 .
- ^ Лидделл, Генри Г .; Скотт, Роберт (1980). Грек-английский лексикон (сокращенное изд.). Оксфорд: издательство Оксфордского университета. ISBN 978-0-19-910207-5 .
- ^ Фиорильо, Ар ; Tykoski, RS (2014). Додсон, Питер (ред.). «Уменьшенный новый тираннозавр с вершины мира» . Plos один . 9 (3): E91287. Bibcode : 2014ploso ... 991287f . doi : 10.1371/journal.pone.0091287 . PMC 3951350 . PMID 24621577 .
- ^ Пол, Грегори С. (1988). Хищные динозавры мира . Нью -Йорк: Саймон и Шустер. С. 464pp . ISBN 978-0-671-61946-6 .
- ^ Лю, Джунчан; Yi, Laiping; Брусатт, Стивен Л.; Ян, Лин; Ли, Хуа; Чен, Лю (2014). «Новая клада азиатских поздних меловых длинношерских тираннозавридов» . Природная связь . 5 : 3788. Bibcode : 2014natco ... 5.3788L . doi : 10.1038/ncomms4788 . PMID 24807588 .
- ^ «Пиноккио Рекс давно выдвинутый тираннозавр обнаружен в Азии» . 7 мая 2014 года. Архивировано с оригинала 2 февраля 2019 года . Получено 8 мая 2014 года .
- ^ Ольшевский, Джордж (1995). Полем Кюраруко Сайзенсен [Dino Frontline ] 9–10 : 92–1
- ^ Ольшевский Г.; Ford TL (1995). «Происхождение и эволюция Tyrannosauridae, часть 2 [на японском языке]». Dino Frontline . 6 : 75–99.
- ^ Фаулер, DW; Вудворд, HN; Freedman, EA; Ларсон, PL; Хорнер, младший (2011). «Реанализ" Raptorex Kriegsteini ": юношеский динозавр тиранозаврида из Монголии" . Plos один . 6 (6): E21376. Bibcode : 2011ploso ... 621376f . doi : 10.1371/journal.pone.0021376 . PMC 3126816 . PMID 21738646 .
- ^ Карр, Томас Д. (2022-11-25). «Переоценка окаменелостей тиранозавра из формирования Ирен Дабасу (конец -кампаниан), внутренней Монголии, Китайская Народная Республика» . Журнал палеонтологии позвоночных . 42 (5). Bibcode : 2022jvpal..42e9817c . doi : 10.1080/02724634.2023.2199817 . ISSN 0272-4634 .
- ^ Серено, Пол С. (1998). «Рациональный для филогенетических определений, с применением к таксономии динозаврии более высокого уровня». Новый ежегодный книга по геологии и палеонтологии, трактаты (на немецком языке). 210 : 41–83. Doi : 10.1127/njgpa/210/998/41 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час Holtz, TR, Jr. (2001), Филогения и таксономия Tyrannosauridae в K Carpenter & D Tanke [Eds.], Жизнь мезозойских позвоночных. Индиана Univ. Пресс, стр. 64-83.
- ^ Брочу, Кристофер Р. (2003). «Остеология Tyrannosaurus rex : понимание почти полного компьютерного томографического анализа скелета и высокого разрешения». Общество палеонтологических мемуаров позвоночных . 7 : 1–138. doi : 10.2307/3889334 . JSTOR 3889334 .
- ^ Серено, Пол С. (7 ноября 2005 г.). «Архосаврия стебля - Taxonsearch» . Архивировано из оригинала 19 февраля 2007 года . Получено 14 января 2008 года .
- ^ Hurum, Jørn H.; Сабат, Кароль. (2003). «Гигантские динозавры Theropod из Азии и Северной Америки: Черепа Tarbosaurus Bataar и Tyrannosaurus rex сравнивали» (Аннотация) . Acta Palaeontologica Polonica . 48 (2): 161–190. Архивировано из оригинала 2012-03-08 . Получено 2008-09-10 .
- ^ Подпрыгнуть до: а беременный Карр, Томас Д. (1999). «Чернофациальная онтогенция у Tyrannosauridae (Dinosauria, Coelurosauria)» . Журнал палеонтологии позвоночных . 19 (3): 497–520. Bibcode : 1999jvpal..19..497c . doi : 10.1080/02724634.1999.10011161 . S2CID 83744433 . Архивировано из оригинала 2020-02-20 . Получено 2019-12-02 .
- ^ «Недавно найденный динозавр-давно носовой двоюродный брат тиранозавра Рекса» . Scienceday . Архивировано с оригинала 2018-03-17 . Получено 2018-03-09 .
- ^ Citreerefreted _ & _ carr2016
- ^ Citirefiriririllo _ & _ tykoski2014
- ^ Лоуэн, Массачусетс ; Ирмис, РБ ; Сертич, JJW ; Керри, PJ ; Сэмпсон, SD (2013). Эванс, Дэвид С (ред.). «Эволюция динозавров тирана отслеживает рост и падение поздне -меловых океанов» . Plos один . 8 (11): E79420. BIBCODE : 2013PLOSO ... 879420L . doi : 10.1371/journal.pone.0079420 . PMC 3819173 . PMID 24223179 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час Эриксон, Грегори М., GM; Маковицки, Питер Дж.; Керри, Филипп Дж .; Norell, Mark A.; Йерби, Скотт А.; Брочу, Кристофер А. (2004). «Гигантизм и сравнительные параметры жизни динозавров тиранозавридов» (PDF) . Природа . 430 (7001): 772–775. Bibcode : 2004natur.430..772e . doi : 10.1038/nature02699 . PMID 15306807 . S2CID 4404887 .
Отбрасываемые долговечность для Т. Рекса варьировались от 2 до 28 лет
(ошибка: doi : 10.1038/nature16487 , PMID 26675726 , реверкция Watch ) и соответствующие оценки массы тела варьировались от 29,9 до 5654 кг - ^ Подпрыгнуть до: а беременный Хорнер, JR & Padian, K. (2004). «Динамика возраста и роста тиранозавра Рекса» . Труды Королевского общества Лондона б . 271 (1551): 1875–1880. doi : 10.1098/rspb.2004.2829 . PMC 1691809 . PMID 15347508 .
- ^ Ли, Эндрю Х.; Вернинг, Сара (2008). «Сексуальная зрелость в растущих динозаврах не соответствует моделям роста рептилии» . Труды Национальной академии наук . 105 (2): 582–587. Bibcode : 2008pnas..105..582L . doi : 10.1073/pnas.0708903105 . PMC 2206579 . PMID 18195356 .
- ^ Schweitzer, MH, Wittmeyer, JL, & Horner, Jr (2005). «Гендерная специфическая репродуктивная ткань в RATITES и Tyrannosaurus rex ». Наука . 308 (5727): 1456–1460. Bibcode : 2005sci ... 308.1456s . doi : 10.1126/science.1112158 . PMID 15933198 . S2CID 30264554 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Подпрыгнуть до: а беременный в дюймовый и Erickson, GM, Currie, PJ, Inouye, BD, & Winn, AA (2006). «Жизненные таблицы тиранозавров: пример биологии неавийской биологии населения динозавров». Наука . 313 (5784): 213–217. Bibcode : 2006sci ... 313..213e . doi : 10.1126/science.1125721 . PMID 16840697 . S2CID 34191607 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ «Первые выявлены эмбрионные окаменелости эмбриона тиранозавра» . Наука . 19 октября 2020 года. Архивировано с оригинала 5 декабря 2020 года . Получено 23 ноября 2020 года .
- ^ Геггель, Лора (15 октября 2020 года). «1 -й из их добрых малышек окаменелостей тиранозавра обнаружено» . LivesCience.com . Архивировано из оригинала 17 октября 2020 года . Получено 17 октября 2020 года .
- ^ Энрикес, Натан Джеймс; Кампионе, Николас; Белл, Фил (21 апреля 2021 года). «Родители тиранозавров с жирной ножкой не могли не отставать от своего худого потомства, показывают ископаемые следы» . Разговор . Архивировано из оригинала 2021-04-26 . Получено 2021-04-28 .
- ^ Энрикес, Натан Дж.; Campione, Nicolás E.; Броум, Том; Фанти, Федерико; Белый, Мэтт А.; Sissons, Robin L.; Салливан, Корвин; Ваврек, Мэтью Дж.; Белл, Фил Р. (21 апреля 2021 г.). «Изучение возможных онтогенетических траекторий у тираннозавридов с использованием треков из формации Wapiti (верхнего кампании) Альберты, Канада». Журнал палеонтологии позвоночных . 40 (6): E1878201. doi : 10.1080/02724634.2021.1878201 . S2CID 234814620 .
- ^ Эриксон, Грегори М.; Карри Роджерс, Кристи; Варриккио, Дэвид Дж.; Норэлл, Марк; Сюй, Син (2007). «Модели роста в задумчивых динозаврах выявляют время сексуальной зрелости у неавийских динозавров и генезиса состояния птиц» . Биологические письма . 3 (5): 558–561. doi : 10.1098/rsbl.2007.0254 . PMC 2396186 . PMID 17638674 .
- ^ Подпрыгнуть до: а беременный в Ли, Эндрю Х.; Вернинг, Сара (2008). «Сексуальная зрелость в растущих динозаврах не соответствует моделям роста рептилии» . ПНА . 105 (2): 582–587. Bibcode : 2008pnas..105..582L . doi : 10.1073/pnas.0708903105 . PMC 2206579 . PMID 18195356 .
- ^ Риклефс, Роберт Э. (2007). «Старение тиранозавра» . Биологические письма . 3 (2): 214–217. doi : 10.1098/rsbl.2006.0597 . PMC 2375931 . PMID 17284406 .
- ^ Подпрыгнуть до: а беременный в дюймовый Танк, Даррен Х.; Керри, Филипп Дж. (1998). «Поведение с кусочкой в тероподных динозаврах: палеопатологические данные» (PDF) . Гайя . 15 : 167–184. Архивировано (PDF) из оригинала на 2008-02-27 . Получено 2008-09-13 . [Не напечатано до 2000 года]
- ^ Подпрыгнуть до: а беременный в дюймовый и Керри, Филипп Дж .; Трекслер, Дэвид; Коппелхус, Ева Б .; Уикс, Келли; Мерфи, Нейт (2005). «Необычный мультидивидуальный костный лепесток тиранозавридов в двух медицине (позднем меловом, кампании) Монтаны (США)». В Карпентере, Кеннет (ред.). Плотноядные динозавры . Блумингтон: издательство Университета Индианы. С. 313–324. ISBN 978-0-253-34539-4 .
- ^ «У Тиранозавра был плохой поворотный круг» . Архивировано из оригинала 7 мая 2012 года . Получено 2007-09-25 .
{{cite web}}: CS1 Maint: Bot: исходный статус URL неизвестен ( ссылка ) - ^ Carrier, DR, Walter, RM и Lee, DV (15 ноября 2001 г.). «Влияние ротационной инерции на поворотные показатели динозавров теропод: подсказки от людей с повышенной инерцией вращения» . Журнал экспериментальной биологии . 204 (Pt 22): 3917–3926. doi : 10.1242/jeb.204.22.3917 . PMID 11807109 . Архивировано из оригинала 6 января 2008 года . Получено 12 октября 2008 года .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Подпрыгнуть до: а беременный Хатчинсон, младший (2004). «Биомеханическое моделирование и анализ чувствительности двуногих способностей к бегу. II. Вымершие таксоны» (PDF) . Журнал морфологии . 262 (1): 441–461. doi : 10.1002/jmor.10240 . PMID 15352202 . S2CID 15677774 . Архивировано из оригинала (PDF) на 2008-10-31.
- ^ Подпрыгнуть до: а беременный Хорнер, Джон Р .; Дон Лесем (1993). Полный Т. Рекс . Нью -Йорк : Саймон и Шустер . с. 120 . ISBN 978-0-671-74185-3 .
- ^ Хольц, Томас Р. (1 мая 1996 г.). «Филогенетическая таксономия коэлурозаврии (динозаврия; Theropoda)» . Журнал палеонтологии . 70 (3): 536–538. Bibcode : 1996jpal ... 70..536H . doi : 10.1017/s0022336000038506 . S2CID 87599102 . Архивировано с оригинала 26 октября 2008 года . Получено 3 октября 2008 года .
- ^ Кристиансен П. (1998). «Значения индикатора прочности костей Theropod, с комментариями о пропорциях конечности и потенциале курсора» (PDF) . Гайя . 15 : 241–255. ISSN 0871-5424 . Архивировано (PDF) из оригинала 2008-10-31 . Получено 2008-10-12 .
- ^ Фарлоу, Джеймс О . ; Смит, Мэтью Б.; Робинсон, Джон М. (1995). «Массовая масса, кость« индикатор силы »и заклинание потенциала Tyrannosaurus rex » . Журнал палеонтологии позвоночных . 15 (4): 713–725. Bibcode : 1995jvpal..15..713f . doi : 10.1080/02724634.1995.10011257 . Архивировано из оригинала 2008-10-23.
- ^ «Чем больше они приходят, тем тяжелее они падают», архивировал 2015-05-31 в The Wayback Matche New Scientist, 7 октября 1995 г., с. 18
- ^ "Жираф" . Wildlifesafari.info. Архивировано из оригинала 12 июня 2010 года . Получено 29 апреля 2006 года .
- ^ "История зоопарка Вудленд -парка - глава 4" . Архивировано из оригинала 2 июня 2007 года . Получено 29 апреля 2006 года .
- ^ Александр, RM (7 августа 2006 г.). «Биомеханика динозавра» . Proc Biol Sci . 273 (1596): 1849–1855. doi : 10.1098/rspb.2006.3532 . PMC 1634776 . PMID 16822743 .
- ^ Ханна, Ребекка Р. (2002). «Многочисленные травмы и инфекция в динозавре с суб-адольтом ( Allosaurus fragilis ) с сравнением с патологией аллозавра в коллекции карьеры динозавров Кливленд-Ллойд». Журнал палеонтологии позвоночных . 22 (1): 76–90. doi : 10.1671/0272-4634 (2002) 022 [0076: miaiia] 2.0.co; 2 . ISSN 0272-4634 . S2CID 85654858 . Каталог травмы аллозавра , известного как « Большой Ал » - по крайней мере один был приписан падению.
- ^ Хатчинсон, Джон Р.; Гарсия, М (28 февраля 2002 г.). « Тираннозавр не был быстрым бегуном» . Природа . 415 (6875): 1018–1021. Bibcode : 2002natur.415.1018h . doi : 10.1038/4151018a . PMID 11875567 . S2CID 4389633 .
- ^ Хайдул Р. (1997). Сухожилия. Карты динозавров . Orbis Publishing Ltd. D36044311.
- ^ Продавцы, WI & Manning, PL (июль 2007 г.). «Оценка максимальной скорости бега динозавра с использованием эволюционной робототехники» . Прокурор R. Soc. Беременный 274 (1626): 2711–6. doi : 10.1098/rspb.2007.0846 . PMC 2279215 . PMID 17711833 . Архивировано из оригинала 2011-02-07 . Получено 2008-10-12 . Это может быть предварительная версия продавцов, WI, Manning, PL, Crompton, RH и Codd, Jr,. (2007), «Изучение эластичных эффектов хранения энергии в двуночном локоме с использованием эволюционной робототехники», Журнал биомеханики , In-Review
- ^ Лиз Сьюард: «Т. Рекс» опередил бы футболиста »« Архивировал 2012-01-07 на машине Wayback . Веб-сайт BBC News , вторник, 21 августа 2007 . года Получено 22 августа 2007 года.
- ^ Callison, G.; HM Quimby (1984). «Крошечные динозавры: они полностью выросли?». Журнал палеонтологии позвоночных . 3 (4): 200–209. Bibcode : 1984JVPAL ... 3...200C . doi : 10.1080/02724634.1984.10011975 .
- ^ Снаво, Эрик; О'Брайен, Хейли; Хендерсон, Дональд М.; Маллисон, Генрих; Сдача, Лара А.; Бернс, Майкл Э.; JR, Томас Р. Хольц; Рассел, Энтони П.; Витмер, Лоуренс М.; Керри, Филипп Дж.; Хартман, Скотт А.; Коттон, Джон Р. (21 февраля 2019 г.). «Более низкая вращательная инерция и более крупные мышцы ног указывают на более быстрые повороты в тиранозавридах, чем в других крупных тероподах» . ПЕРЕЙ . 7 : E6432. doi : 10.7717/peerj.6432 . PMC 6387760 . PMID 30809441 .
- ^ Dececchi, T. Alexander; Mloszewska, Aleksandra M.; Хольц, Томас Р.; Хабиб, Майкл Б.; Ларссон, Ханс Се (2020). «Быстрые и скромные: дивергентные локомоторивые стратегии способствуют удлинению конечностей у динозавров с теропод» . Plos один . 15 (5): E0223698. Bibcode : 2020ploso..1523698d . doi : 10.1371/journal.pone.0223698 . PMC 7220109 . PMID 32401793 .
- ^ «Т. Рекс был чемпионом Уокера, сверхэффективным на более низких скоростях» . Эврикалерт! Полем Архивировано из оригинала 2020-06-27 . Получено 2020-06-27 .
- ^ Zhou Zhonghe, z; Барретт, Пол М.; Хилтон, Джейсон. (2003). «Исключительно сохранившаяся нижняя меловая экосистема». Природа . 421 (6925): 807–814. Bibcode : 2003natur.421..807z . doi : 10.1038/nature01420 . PMID 12594504 . S2CID 4412725 .
- ^ Чен Пейджи, Пей-дзи; Донг Жиминг ; Чжэнь Шуонан. (1998). «Исключительно хорошо сохранившийся динозавр Theropod из формирования Йикса в Китае» (PDF) . Природа . 391 (6663): 147–152. Bibcode : 1998natur.391..147c . doi : 10.1038/34356 . S2CID 4430927 .
- ^ Сюй Син, х ; Чжоу Чжонге; Прум, Ричард А. (2003). «Разветвленные покровные структуры в Sinornithosaurus и происхождение перьев». Природа . 410 (6825): 200–204. Bibcode : 2001natur.410..200x . doi : 10.1038/35065589 . PMID 11242078 . S2CID 4426803 .
- ^ Lingham-Soliar, Theagarten; Feduccia, Alan ; Ван, Сяолин (2007). «Новый китайский образец показывает, что« протофичики »в раннем мела -тероподном динозавре Sinosauropteryx являются деградированными коллагеновыми волокнами» . Труды Королевского общества Лондона. Серия B, биологические науки . 274 (1620): 1823–1829. doi : 10.1098/rspb.2007.0352 . PMC 2270928 . PMID 17521978 .
- ^ «Архивная копия» (PDF) . Архивировано из оригинала (PDF) 2012-04-17 . Получено 2012-04-04 .
{{cite web}}: CS1 Maint: архивная копия как заголовок ( ссылка ) - ^ Подпрыгнуть до: а беременный Белл, PR; Кампионе, NE; Люди, WS; Керри, PJ; Ларсон, PL; Танк, DH; Баккер, RT (2017). «Интуирование тиранозавра выявляет противоречивые закономерности гигантизма и эволюции перьев» . Биологические письма . 13 (6): 20170092. DOI : 10.1098/rsbl.2017.0092 . PMC 5493735 . PMID 28592520 .
- ^ http://globalnews.ca/news/3506400/tyrannosaurus-rex-fossil-skin-cales-university-of-alberta/ архивировано 2017-06-17 на машине Wayback, что чувствует Т. Рекс? U of the Paleontologists узнают в новом обнаружении ископаемых
- ^ Currie, PJ, Badamgarav, D., Koppelhus, EB 2003. Первые поздние меловые следы от местности Немегта в Гоби Монголии. Ichnos. Vol.10: 1-12.currie, PJ, Badamgarav, D., Koppelhus, EB 2003.
- ^ Carr, TD, Varricchio, DJ, Sedlmayr, JC, Roberts, EM, & Moore, Jr (2017). Новый тираннозавр с доказательствами анагенеза и крокодилоподобной чувствительной системы лица. Научные отчеты , 7.
- ^ Виттон, Марк; Хон, Дэвид (сентябрь 2018 г.). "Tyrannososaurid Theropods: они когда -нибудь улыбались, как крокодилы?" (PDF) . Ежегодный симпозиум палеонтологии позвоночных и сравнительной анатомии .
- ^ Сатта, Эван Т.; Флетчер, Ян; Мартин, Питер; Питман, Майкл; Кэй, Томас Г.; Правда, Лоуренс Д.; Norell, Mark A.; Эббот, Джеффри Д.; Призыв, Роджер Э.; Пенкман, Кирсти; Винтер, Якоб (2018-11-01). «Сохранение перьевых волокон от позднего мелового динозавра Shuvuuia Deserti вызывает обеспокоенность по поводу иммуногистохимического анализа о окаменелостях» (PDF) . Органическая геохимия . 125 : 142–151. Bibcode : 2018Rogeo.125..142S . doi : 10.1016/j.orggeochem.2018.09.008 . ISSN 0146-6380 . S2CID 105753275 .
- ^ Блогер, гость. «Десять вещей, которые мы не знаем о тиранозаврах» . Архивировано из оригинала 2020-09-30 . Получено 2021-01-05 .
- ^ Гуарино, Бен (2021-10-27). «Tyrannosaurus rex имела чешуйчатую кожу и не был покрыт перьями, говорится в новом исследовании» . Вашингтон пост . ISSN 0190-8286 . Получено 2023-08-07 .
- ^ Стивенс, Кент А. (июнь 2006 г.). «Бинокулярное зрение у динозавров с теропод» (PDF) . Журнал палеонтологии позвоночных . 26 (2): 321–330. doi : 10.1671/0272-4634 (2006) 26 [321: bvitd] 2.0.co; 2 . ISSN 0272-4634 . S2CID 85694979 . Архивировано из оригинала (PDF) 2012-03-23.
- ^ Джаффе, Эрик (1 июля 2006 г.). «Зрелище для« Саур Глаза: Т. Рекс Видение было одним из лучших природы » . Science News . 170 (1): 3–4. doi : 10.2307/4017288 . JSTOR 4017288 . Архивировано с оригинала 29 сентября 2012 года . Получено 6 октября 2008 года .
- ^ Савелев, Сергей В.; Алифанов, Владимир Р. (2005). «Новое исследование мозга хищного динозавра Tarbosaurus Bataar (Theropoda, Tyrannosauridae)». Палеонтологический журнал . 41 (3): 281–289. Bibcode : 2007palj ... 41..281s . doi : 10.1134/s0031030107030070 . S2CID 53529148 .
- ^ Подпрыгнуть до: а беременный Карр, Томас Д.; Варриккио, Дэвид Дж.; Sedlmayr, Jayc C.; Робертс, Эрик М.; Мур, Джейсон Р. (2017-03-30). «Новый тираннозавр с доказательствами анагенеза и крокодилоподобной чувственной системы лица» . Научные отчеты . 7 : 44942. Bibcode : 2017natsr ... 744942c . doi : 10.1038/srep44942 . ISSN 2045-2322 . PMC 5372470 . PMID 28358353 .
- ^ Баркер, Крис; Найш, Даррен; Katsamenis, Orestis; Дайк, Гарет (16 июня 2017 г.). «Сложная нейроанатомия в ростроме острова Уайт Теропод Невейтор Салерии» . Научные отчеты . 7 (1): 3749. Bibcode : 2017Natsr ... 7.3749b . doi : 10.1038/s41598-017-03671-3 . PMC 5473926 . PMID 28623335 .
- ^ Лейтч, Дункан Б.; Катания, Кеннет С. (2012-12-01). «Структура, иннервация и свойства ответа покровных сенсорных органов у крокодилов» . Журнал экспериментальной биологии . 215 (23): 4217–4230. doi : 10.1242/jeb.076836 . ISSN 0022-0949 . PMC 4074209 . PMID 23136155 .
- ^ Ди-Пой, Николас; Milinkovitch, Michel C. (2013-07-02). «Крокодилианы эволюционировали рассеянные мультисенсорные микроорганы» . Эводево . 4 (1): 19. doi : 10.1186/2041-9139-4-19 . ISSN 2041-9139 . PMC 3711810 . PMID 23819918 .
- ^ Баккер, Роберт Т. (1968). «Превосходство динозавров». Открытие . 3 (2): 11–12.
- ^ Баккер, Роберт Т. (1972). «Анатомические и экологические доказательства эндотермии у динозавров». Природа . 238 (5359): 81–85. Bibcode : 1972natur.238 ... 81b . doi : 10.1038/238081a0 . S2CID 4176132 .
- ^ Баккер, Роберт Т. (1986). ДИНОСАВЕР ВЕРЕЗИ . Нью -Йорк: Kensington Publishing. ISBN 978-0-688-04287-5 Полем OCLC 13699558 . [ страница необходима ]
- ^ Баррик, Риз Э.; Уильям Дж. Лож (июль 1994 г.). «Термофизиология Tyrannosaurus rex: данные из изотопов кислорода». Наука . 265 (5169). Нью -Йорк : 222–224. Bibcode : 1994sci ... 265..222b . doi : 10.1126/science.265.5169.222 . PMID 17750663 . S2CID 39392327 .
- ^ Баррик, Риз Э.; Уильям Дж. Лож (октябрь 1999). «Термофизиология и биология гиганотозавра : сравнение с тиранозавром » . Palaeontologia Electronica . 2 (2). Архивировано из оригинала 2011-05-17 . Получено 2008-10-12 .
- ^ Паладино, Фрэнк В.; Spotila, Джеймс Р.; Додсон, Питер (1999). «План гиганта: моделирование физиологии крупных динозавров». В Джеймсе О. Фарлоу ; М.К. Бретт-Сурман (ред.). Полный динозавр . Блумингтон: издательство Университета Индианы. С. 491–504. ISBN 978-0-253-21313-6 .
- ^ Чинсами, Анусуя; Виллем Дж. Хиллуениус (2004). «Физиология неавийских динозавров». В Дэвиде Б. Вайшампел; Питер Додсон; Halszka Osmólska (ред.). Динозаврия . Беркли: Университет Калифорнийской прессы. С. 643–659. ISBN 978-0-520-24209-8 .
- ^ Сеймур, Роджер С. (2013-07-05). «Максимальная аэробная и анаэробная выработка электроэнергии в крупных крокодилах по сравнению с млекопитающими: последствия для гигантотермии динозавров» . Plos один . 8 (7): E69361. BIBCODE : 2013PLOSO ... 869361S . doi : 10.1371/journal.pone.0069361 . ISSN 1932-6203 . PMC 3702618 . PMID 23861968 .
- ^ Подпрыгнуть до: а беременный Фарлоу, Джеймс О . ; ПИАНКА, Эрик Р. (2002). «Размер тела перекрывается, распределение среды обитания и требования к жилой площади сухое позвоночные хищники: последствия для палеоэкологии крупных динозавров с теропод» (PDF) . Историческая биология . 16 (1): 21–40. Bibcode : 2002hbio ... 16 ... 21f . doi : 10.1080/0891296031000154687 . S2CID 18114585 .
- ^ Varricchio, David J. (2001). «Содержание кишечника из мелового тиранозаврида: последствия для пищеварительных трактов динозавров» (PDF) . Журнал палеонтологии . 75 (2): 401–406. doi : 10.1666/0022-3360 (2001) 075 <0401: gcfact> 2.0.co; 2 . ISSN 0022-3360 . S2CID 86113170 .
- ^ Guinness World Records Ltd. (2003). 2003 Guinness World Record . PG 90.
- ^ Подпрыгнуть до: а беременный Эберт, Дэвид А.; МакКри, Ричард Т. (2001). "Были ли крупные тероподстные?". Журнал палеонтологии позвоночных . 21 (дополнение к номеру 3): 46a. doi : 10.1080/02724634.2001.10010852 . S2CID 220414868 .
- ^ Роуч, Брайан Т.; Бринкман, Даниэль Л. (2007). «Переоценка кооперативной охоты и общительности в Deinonychus antirrhopus и других неавийских динозаврах теропод». Бюллетень музея естественной истории Пибоди . 48 (1): 103–138. doi : 10.3374/0079-032x (2007) 48 [103: Acocph] 2.0.co; 2 . S2CID 84175628 .
- ^ Подпрыгнуть до: а беременный Керри, Филипп Дж. (1998). «Возможные доказательства общительного поведения у тираннозавридов» (PDF) . Гайя . 15 : 271–277. Архивировано (PDF) из оригинала на 2009-03-26 . Получено 2008-09-13 . (не напечатано до 2000 года)
- ^ Роуч, Брайан Т.; Бринкман, Даниэль Т. (2007). «Переоценка кооперативной охоты и общительности в Deinonychus antirrhopus и других неавийских динозаврах теропод». Бюллетень музея естественной истории Пибоди . 48 (1): 103–138. doi : 10.3374/0079-032x (2007) 48 [103: Acocph] 2.0.co; 2 . S2CID 84175628 .
- ^ МакКри, Ричард Т.; Бакли, Лиза Г.; Фарлоу, Джеймс О.; Локли, Мартин Дж.; Керри, Филипп Дж.; Мэтьюз, Неффра А.; Пембертон, С. Джордж (2014). «Террор тиранозавров»: первые пути тиранозавридов и свидетельство общительности и патологии у тиранозауридов » . Plos один . 9 (7): E103613. Bibcode : 2014ploso ... 9J3613M . doi : 10.1371/journal.pone.0103613 . PMC 4108409 . PMID 25054328 .
- ^ «Tyrannosaurus Tracks показывает прогулку динозавра» . 2014-07-23. Архивировано с оригинала 2014-09-30 . Получено 2015-01-16 .
- ^ «Треки намекают на социальную жизнь динозавров тирана» . 2014-07-23. Архивировано из оригинала 2015-01-08 . Получено 2015-01-16 .
- ^ Титус, Алан Л.; Кнолл, Катя; Сертич, Джозеф Дж.В.; Ямамура, Дайго; Суарес, Селина А.; Glasspool, Ian J.; Джинувз, Джонатан Э.; Лукачич, Эбигейл К.; Робертс, Эрик М. (19 апреля 2021 г.). «Геология и тафономия уникального тиранозавридного костей из верхнего кампании кайпаровитса формирования южной части штата Юта: последствия для общительности тиранозавридов» . ПЕРЕЙ . 9 : E11013. doi : 10.7717/peerj.11013 . PMC 8061582 . PMID 33976955 .
- ^ Эйлперин, Джульетта (19 апреля 2021 года). «Тиранозавры, вероятно, охотились в пакетах, а не на соло, ученые находят» . The Washington Post . Архивировано из оригинала 20 апреля 2021 года . Получено 27 сентября 2021 года .
- ^ «Тираннозавр Рекс» охотился в пакетах » . 22 июня 2011. Архивировано с оригинала 2021-09-27 . Получено 2021-09-27 .
- ^ Подпрыгнуть до: а беременный в Jacobsen, AR 2001. Зубная, нанесенная на зубах маленькая тероподная кость: чрезвычайно редкая следов. п. 58-63. В: Жизнь мезозойских позвоночных. Ed.S Tanke, DH, Carpenter, K., Skrepnick, MW Indiana University Press.
- ^ Подпрыгнуть до: а беременный Abler, WL 2001. Керф-и бурная модель зубных зубцов тиранозавра. п. 84-89. В: Жизнь мезозойских позвоночных . Ed.S Tanke, DH, Carpenter, K., Skrepnick, MW Indiana University Press.
- ^ Lambe, LB (1917). «Цивяшний тероподный динозавр Gorgosaurus » . Мемуары геологической службы Канады . 100 : 1–84. doi : 10.4095/101672 . Архивировано из оригинала 2021-09-09 . Получено 2021-09-23 .
- ^ Фарлоу, Джо; Хольц, Т.Р. (2002). Kowalewski, M.; Келли, Ph (ред.). «Окаменечная запись хищничества» (PDF) . Документы палеонтологического общества. С. 251–266. Архивировано из оригинала (PDF) на 2008-10-31.
- ^ Дори, М. (1997). Тиранозавр. Карты динозавров . Orbis Publishing Ltd. D36045907.
- ^ Подпрыгнуть до: а беременный Хорнер, младший (1994). «Стейк -ножи, бусинки и крошечные руки (портрет тиранозавра в качестве мусорщика)». Специальное публикация палеонтологического общества . 7 : 157–164. doi : 10.1017/s2475262200009497 .
- ^ Amos, J. (31 июля 2003 г.). «Т. Рекс идет в суд» . Би -би -си. Архивировано из оригинала 13 марта 2012 года . Получено 12 октября 2008 года .
- ^ Подпрыгнуть до: а беременный Уолтерс М., Пейкер Дж. (1995). Словарь доисторической жизни. Клермонт книги. ISBN 1-85471-648-4 .
- ^ Эриксон GM; Олсон К.Х. (1996). «Знаки укуса, связанные с Tyrannosaurus Rex : предварительное описание и последствия» . Журнал палеонтологии позвоночных . 16 (1): 175–178. Bibcode : 1996jvpal..16..175e . doi : 10.1080/02724634.1996.10011297 . Архивировано из оригинала 2021-10-19 . Получено 2020-09-12 .
- ^ Карпентер К. (1998). «Свидетельство хищнического поведения тероподных динозавров» . Гайя . 15 : 135–144. Архивировано из оригинала 17 ноября 2007 года . Получено 5 декабря 2007 года .
- ^ Фаулер, Денвер W.; Роберт М. Салливан (2006). «Кератопсидный таз с зубцами из верхней меловой формирования Киртланда, Нью -Мексико: свидетельство поведения позднего кампании тиранозаврида». Музей естественной истории и науки Нью -Мексико . 35 : 127–130.
- ^ Терриен, Ф.; Зеленицкий, DK; Танака, К.; Ворис, JT; Эриксон, GM; Керри, PJ; Debuhr, cl; Kobayashi, Y. (2023). «Исключительно сохраненное содержание желудка молодых тиранозавридов выявляет онтогенетический сдвиг в пище в культовом вымершем хищнике» . Наука достижения . 9 (49): eadi0505. Bibcode : 2023scia .... 9i.505t . doi : 10.1126/sciadv.adi0505 . PMC 10846869 . PMID 38064561 .
- ^ Longrich, NR; Хорнер, младший; Эриксон, GM; Currie, PJ (2010). «Каннибализм в тиранозавре Рекс » . Plos один . 5 (10): E13419. BIBCODE : 2010PLOSO ... 513419L . doi : 10.1371/journal.pone.0013419 . PMC 2955550 . PMID 20976177 .
- ^ «Новые доказательства каннибализма у динозавров тиранозавридов из верхнего мела (Кампанский/Маастрихтский бассейн Сан -Хуан в Нью -Мексико» . Researchgate . Архивировано из оригинала 2021-11-10 . Получено 2021-01-06 .
- ^ Додсон, Питер; Бритт, Брукс; Карпентер, Кеннет; Форстер, Кэтрин А.; Джилетт, Дэвид Д.; Norell, Mark A.; Ольшевский, Джордж; Parrish, J. Michael & Weishampel, David B. (1993). «Абелисавр». Возраст динозавров . Publications International, Ltd. п. 105. ISBN 978-0-7853-0443-2 .
- ^ Фиорильо, Энтони Р.; Ганглофф, Роланд А. (2000). «Тероподные зубы из формации Принс -Крик (меловой) северной Аляски, с предположениями о палеоэкологии арктических динозавров». Журнал палеонтологии позвоночных . 20 (4): 675–682. doi : 10.1671/0272-4634 (2000) 020 [0675: ttftpc] 2.0.co; 2 . ISSN 0272-4634 . S2CID 130766946 .
- ^ «Эти 81-миллионные зубы являются первыми свидетельствами крупнейшего в Японии, когда-либо динозавре | Ирландский экзаменатор» . Архивировано с оригинала 2016-04-12 . Получено 2016-03-10 .
- ^ Томас Р. Хольц младший (1994). «Филогенетическое положение Tyrannosauridae: последствия для системной теропод». Журнал палеонтологии . 68 (5): 1100–1117. Bibcode : 1994jpal ... 68.1100h . doi : 10.1017/s0022336000026706 . JSTOR 1306180 . S2CID 129684676 .
- ^ Февраль 2016 года, Лора Геггель 29 (29 февраля 2016 г.). «Т. Рекс, вероятно, был инвазивным видом» . LivesCience.com . Архивировано из оригинала 14 июля 2019 года . Получено 5 января 2021 года .
{{cite web}}: CS1 Maint: числовые имена: список авторов ( ссылка )
Внешние ссылки
[ редактировать ]- Список образцов Tyrannosauridae и обсуждение в базе данных Theropod















- ^ Иеронима, Тобин; Витмер, Лоуренс; Танк, Даррен; Керри, Филипп (26 августа 2009 г.). «Лицевой покровные цератопсиды центрозаврина: морфологические и гистологические корреляты новых кожных структур» . Анатомическая запись . 292 (9): 1370–1396. doi : 10.1002/ar.20985 . PMID 19711467 .