Жирафа
| Жирафы Временной диапазон: Миоцен до недавнего времени | |
|---|---|
 | |
| Жираф Масаи ( Giraffa Tippelskirchi ) в национальном парке Микуми , Танзания | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Парнокопытные |
| Семья: | Жирафы |
| Род: | Жирафа Бриссон, 1762 г. |
| Разновидность | |
 | |
| Распространение жирафа | |

Жираф принадлежащее — крупное африканское копытное млекопитающее, к роду Giraffa . Это самое высокое из ныне живущих наземных животных и самое большое жвачное животное на Земле . Традиционно жирафы считались одним видом , Giraffa Camelopardalis , имеющим девять подвидов . Совсем недавно исследователи предложили разделить их на четыре существующих вида благодаря новым исследованиям их митохондриальной и ядерной ДНК , а отдельные виды можно отличить по рисунку их меха . Семь других вымерших видов жирафов летописи известны из окаменелостей .
Отличительными характеристиками жирафа являются чрезвычайно длинная шея и ноги, роговидные оссиконы и пятнистая шерсть. Он отнесен к семейству Giraffidae вместе со своим ближайшим родственником окапи . Его рассеянный ареал простирается от Чада на севере до Южной Африки на юге и от Нигера на западе до Сомали на востоке. Жирафы обычно обитают в саваннах и лесах . Их источником пищи являются листья, плоды и цветы древесных растений, в первую очередь видов акации , которые они поедают на высотах, которых не может достичь большинство других наземных травоядных животных .
Львы , леопарды , пятнистые гиены и африканские дикие собаки могут охотиться на жирафов. Жирафы живут стадами родственных самок и их потомков или холостяцкими стадами неродственных взрослых самцов, но они стадны и могут собираться в большие группы. Самцы устанавливают социальную иерархию посредством «обхватывания шеи», боевых схваток, в которых шея используется как оружие. Доминирующие самцы получают доступ к спариванию с самками, которые несут исключительную ответственность за воспитание детенышей.
Жираф заинтриговал различные древние и современные культуры своим своеобразным внешним видом и часто изображался на картинах, книгах и мультфильмах. классифицирует его Международный союз охраны природы (МСОП) как уязвимый к исчезновению. Он был истреблен во многих частях своего бывшего ареала. Жирафы по-прежнему встречаются во многих национальных парках и заповедниках насчитывается около 97 500 особей жирафов , но, по оценкам на 2016 год, в дикой природе . В 2010 году в зоопарках содержалось более 1600 особей.
Этимология
Название «жираф» имеет свое самое раннее известное происхождение от арабского слова «зарафах» ( زرافة ). [2] в конечном итоге от персидского زُرنَاپَا ( зурнапа ), соединения زُرنَا ( зурна , «флейта, зурна») и پَا ( па , «нога»). [3] [4] В раннем современном английском языке написания jarraf и ziraph , вероятно, непосредственно из арабского языка. использовались [5] а в среднеанглийском jarraf и ziraph — gerfauntz . Итальянская . форма жирафа возникла в 1590-х годах Современная английская форма возникла около 1600 года от французского жирафа . [2]
«Камелопард» / k ə ˈ m ɛ l ə ˌ p ɑːr d / — архаичное английское название жирафа; оно происходит от древнегреческого καμηλοπάρδαλις ( kamēlopárdalis ), от κάμηλος ( kámēlos ), « верблюд », и πάρδαλις ( párdalis ), « леопард », имея в виду его верблюжью форму и леопардовую окраску. [6] [7]
Таксономия
Эволюция
Жираф — один из двух ныне живущих родов семейства Giraffidae отряда парнокопытных , второй — окапи . [8] Это жвачные животные клады Pecora , наряду с Antilocapridae ( вилорогами ), Cervidae (олени), Bovidae (крупный рогатый скот, антилопы, козы и овцы) и Moschidae (кабарга). 2019 года Исследование генома (кладограмма ниже) показало, что Giraffidae являются родственным таксоном Antilocapridae, раскол которого, по оценкам, произошел более 20 миллионов лет назад. [9]



Семейство Giraffidae когда-то было гораздо более обширным: было описано более 10 ископаемых родов . [8] Удлинение шеи, по-видимому, началось на ранних этапах развития жирафов . Сравнение жирафов и их древних родственников позволяет предположить, что позвонки, расположенные вблизи черепа, удлинялись раньше, а затем удлинялись позвонки ниже. [10] Одним из ранних предков жирафов был Canthumeryx , который, по разным оценкам, жил от 25 до 20 миллионов лет назад , 17–15 млн лет назад или 18–14,3 млн лет назад, и чьи месторождения были обнаружены в Ливии. Это животное напоминало антилопу и имело легкое тело среднего размера. Жирафокерикс появился 15–12 млн лет назад на Индийском субконтиненте и напоминал окапи или маленького жирафа, имел более длинную шею и подобные оссиконы . [8] Жирафокерикс , возможно, принадлежал к одной кладе с более массивными жирафами, такими как Sivatherium и Bramatherium . [10]

Жирафы, такие как Palaeotragus , Shansitherium и Samotherium, появились 14 млн лет назад и жили по всей Африке и Евразии. Эти животные имели более широкие черепа с уменьшенными лобными полостями. [8] [10] Палеотрагус напоминал окапи и, возможно, был его предком. [8] Другие считают, что линия окапи разошлась раньше, до жираффокерикса . [10] Самотерий был особенно важным переходным ископаемым в линии жирафов, поскольку длина и структура его шейных позвонков находились между таковыми у современного жирафа и окапи, а положение его шеи, вероятно, было похоже на положение шеи первого. [11] Болиния , впервые появившаяся в юго-восточной Европе и жившая 9–7 млн лет назад, вероятно, была прямым предком жирафа. Болиния очень напоминала современных жирафов, имея длинную шею и ноги, а также схожие оссиконы и зубные ряды. [8]
Болиния колонизировала Китай и северную Индию и произвела на свет жирафов , которые около 7 миллионов лет назад достигли Африки. Изменения климата привели к исчезновению азиатских жирафов, в то время как африканские жирафы выжили и превратились в новые виды. Живые жирафы, по-видимому, возникли около 1 миллиона лет назад в Восточной Африке во время плейстоцена . [8] Некоторые биологи предполагают, что современные жирафы произошли от G. jumae ; [12] другие считают G. gracilis более вероятным кандидатом. G. jumae был крупнее и крепче, а G. gracilis — меньше и тоньше. [8]
Считается, что переход от обширных лесов к более открытым местам обитания , начавшийся 8 млн лет назад, стал основным фактором эволюции жирафов. [8] За это время тропические растения исчезли и были заменены засушливыми растениями C4 сухая саванна . , а на востоке и севере Африки и западной Индии возникла [13] [14] Некоторые исследователи выдвинули гипотезу, что эта новая среда обитания в сочетании с другой диетой, включающей виды акации, возможно, подвергла предков жирафов воздействию токсинов, которые вызвали более высокий уровень мутаций и более высокую скорость эволюции. [15] Рисунок шерсти современных жирафов также мог совпасть с этими изменениями среды обитания. Предполагается, что азиатские жирафы имели окраску, более похожую на окапи. [8]
Длина генома жирафа составляет около 2,9 миллиарда пар оснований по сравнению с 3,3 миллиарда пар оснований окапи. Из белков в генах жирафа и окапи 19,4% идентичны. Расхождение . линий жирафов и окапи датируется примерно 11,5 млн лет назад Небольшая группа регуляторных генов у жирафа, по-видимому, отвечает за рост животного и связанные с ним адаптации кровообращения. [16] [17]
Виды и подвиды
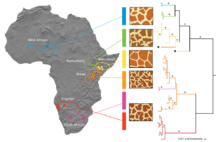
Международный союз охраны природы (МСОП) в настоящее время признает только один вид жирафов с девятью подвидами. [1]
Карл Линней первоначально классифицировал живых жирафов как один вид в 1758 году. Он дал ему биномиальное название Cervus Camelopardalis . Матюрен Жак Бриссон придумал родовое название «Жираф» в 1762 году. [19] В течение 1900-х годов были предложены различные таксономии с двумя или тремя видами. [20] Исследование генетики жирафов с использованием митохондриальной ДНК, проведенное в 2007 году, показало, что как минимум шесть линий можно признать видами. [18] Исследование 2011 года с использованием подробного анализа морфологии жирафов и применения концепции филогенетических видов описало восемь видов ныне живущих жирафов. [21] Исследование 2016 года также пришло к выводу, что современные жирафы состоят из нескольких видов. Исследователи предположили существование четырех видов, которые не обменивались генетической информацией между собой на протяжении от 1 до 2 миллионов лет. [22]
Исследование 2020 года показало, что в зависимости от выбранного метода для рода Giraffa можно рассматривать различные таксономические гипотезы, признающие от двух до шести видов . Это исследование также показало, что методы объединения нескольких видов могут привести к таксономическому чрезмерному разделению, поскольку эти методы ограничивают географические структуры, а не виды. Гипотеза трех видов, которая G. признает камелопардалис , Г. жираф и Г. Tippelskirchi , подтверждается филогенетическим анализом, а также подтверждается большинством популяционно-генетических и многовидовых объединенных анализов. [23] проведенное в 2021 году, Исследование полногеномного секвенирования, предполагает существование четырех отдельных видов и семи подвидов. [24] Исследование 2024 года выявило больший поток древних генов между популяциями, чем ожидалось. [25]
ниже На кладограмме показаны филогенетические связи между четырьмя предложенными видами и семью подвидами на основе анализа генома 2021 года. [24] Восемь линий соответствуют восьми традиционным подвидам в одновидовой гипотезе. Жираф Ротшильда отнесен к G. Camelopardalis Camelopardalis .
|
В следующей таблице сравниваются различные гипотезы относительно видов жирафов. В столбце описания показаны традиционные девять подвидов в гипотезе одного вида. [1] [26]
| Описание | Изображение | Таксономия восьми видов [21] | Таксономия четырех видов [22] [24] | Таксономия трех видов [23] |
|---|---|---|---|---|
| Кордофанский жираф ( G.c. antiquorum ) имеет ареал, включающий южный Чад , Центральноафриканскую Республику , северный Камерун и северо-восточную Демократическую Республику Конго . [1] Популяции Камеруна ранее включались в G. c. перальта , но это было неправильно. [27] По сравнению с нубийскими жирафами, этот подвид имеет более мелкие и неравномерные пятна. Пятна присутствуют на внутренней стороне ног, иногда под скакательными суставами . У мужчин имеется срединная шишка. [28] : 51–52 Считается, что около 2000 особей остаются в дикой природе. [1] Существовала значительная путаница по поводу статуса этого подвида и G. c. перальта в зоопарках. В 2007 году все предполагаемые G. c. peralta в европейских зоопарках на самом деле оказался G. c. антикворум . [27] С учетом этой поправки около 65 из них живут в зоопарках. [29] |  | Кордофанский жираф ( Г. древних ) [30] | Северный жираф ( Г. камелопардалис ) Три или четыре подвида:
| |
| Нубийский жираф ( G. c. Camelopardalis ) встречается в восточной части Южного Судана и юго-западной Эфиопии , а также в Кении и Уганде . [1] пятна с острыми краями, У него каштановые окруженные преимущественно белыми линиями, а на нижней стороне пятен нет. В середине головы самца выделяется шишка. [28] : 51 Считается, что около 2150 особей остаются в дикой природе, а еще 1500 особей принадлежат к Ротшильда экотипу . [1] С добавлением жирафа Ротшильда к нубийскому подвиду нубийский жираф стал очень распространён в неволе, хотя исходный фенотип встречается редко — группа содержится в зоопарке Аль-Айн в Объединенных Арабских Эмиратах . [31] В 2003 году эта группа насчитывала 14 человек. [32] |  | Нубийский жираф ( Г. камелопардалис ) [26] Также известен как Баринго жираф или угандийский жираф. Два подвида:
| ||
| Жираф Ротшильда ( G. c. rothschildi ) может быть экотипом G. Camelopardalis . Его ареал включает части Уганды и Кении . [1] Его присутствие в Южном Судане сомнительно. [33] У этого жирафа большие темные пятна с обычно четко выраженными краями, но иногда с разделенными краями. Внутри темных пятен также могут быть завитки бледного цвета. Пятнистость редко доходит до скакательных суставов и редко до копыт. У этого экотипа также может развиться пять «рогов». [28] : 53 Считается, что около 1500 особей остаются в дикой природе. [1] и более 450 живут в зоопарках. [29] Согласно генетическому анализу, проведенному примерно в сентябре 2016 года, он конспецифичен нубийскому жирафу ( G. c. Camelopardalis ). [22] |  | |||
| Западноафриканский жираф ( G.c.peralta ) является эндемиком юго-западного Нигера. [1] У этого животного более светлая шерсть (мех), чем у других подвидов. [34] : 322 с красными лопастными пятнами, доходящими до скакательных суставов. Оссиконы более прямостоячие, чем у других подвидов, у самцов хорошо развиты срединные бугорки. [28] : 52–53 Это самый исчезающий подвид среди жирафов : в дикой природе осталось 400 особей. [1] Раньше считалось, что жирафы в Камеруне принадлежат к этому виду, но на самом деле это G. c. антикворум . Эта ошибка привела к некоторой путанице в отношении его статуса в зоопарках, но в 2007 году было установлено, что все « G.c.peralta », хранящиеся в европейских зоопарках, на самом деле являются G.c.peralta. антикворум . То же исследование 2007 года показало, что западноафриканский жираф был более тесно связан с жирафом Ротшильда, чем кордофан, и его предок, возможно, мигрировал из восточной Африки в северную, а затем на запад по мере распространения пустыни Сахара . В самом крупном случае озеро Чад могло служить границей между западноафриканскими и кордофанскими жирафами во время голоцена (до 5000 г. до н. э.). [27] |  | Западноафриканский жираф (из Г. Пера ), [35] Также известен как нигерийский жираф или нигерийский жираф. | ||
| ( Сетчатый жираф G. c. reticulata ) обитает на северо-востоке Кении, на юге Эфиопии и в Сомали . [1] Его характерный рисунок шерсти состоит из красновато-коричневых многоугольных пятен с острыми краями, окруженных тонкими белыми линиями. Пятна могут распространяться или не распространяться под скакательными суставами, у кобелей имеется срединная шишка. [28] : 53 По оценкам, в дикой природе осталось 8660 особей. [1] и, согласно данным Международной системы информации о видах , более 450 особей живут в зоопарках. [29] Исследование 2024 года показало, что сетчатый жираф является результатом гибридизации северной и южной линий жирафов. [25] |  | Сетчатый жираф ( G. reticulata ), [36] Также известен как сомалийский жираф. | ||
| Ангольский жираф ( G.c. angolensis ) [37] встречается в северной Намибии , юго-западной Замбии , центральной Ботсване , западном Зимбабве , южном Зимбабве и, с середины 2023 года, снова в Анголе. [38] [1] [39] [40] Генетическое исследование этого подвида, проведенное в 2009 году, показало, что популяции северной пустыни Намиб и национального парка Этоша образуют отдельный подвид. [41] Этот подвид белый с крупными коричневыми пятнами с заостренными или срезанными краями. Пятнистый рисунок распространяется на все ноги, но не на верхнюю часть лица. Пятна на шее и крупе обычно довольно маленькие. У подвида также есть белая отметина на ухе. [28] : 51 По оценкам, около 13 000 животных остаются в дикой природе. [1] и около 20 живут в зоопарках. [29] |  | Ангольский жираф ( G. angolensis ) Также известен как намибийский жираф. | Южный жираф ( G. giraffa ) Два подвида:
| |
| ( Южноафриканский жираф G. c. giraffa ) встречается на севере Южной Африки , на юге Ботсваны, на севере Ботсваны и на юго-западе Мозамбика . [1] [39] [40] Он имеет желтовато-коричневый фон с темными, несколько округлыми пятнами «с небольшими выступами». Пятна распространяются вниз по ногам, становясь при этом меньше. Средний размер самцов относительно небольшой. [28] : 52 По оценкам, в дикой природе осталось максимум 31 500 особей. [1] и около 45 живут в зоопарках. [29] |  | Южноафриканский жираф ( Г. жираф ) [42] Также известен как Капский жираф. | ||
| Масаи -жирафа ( G.c. Tippelskirchi ) можно встретить в центральной и южной Кении, а также в Танзании . [1] Рисунок его шерсти очень разнообразен: пятна варьируются от в основном округлых и гладких до овальных с насечками или скошенными краями. [43] Срединная шишка обычно присутствует у мужчин. [28] : 54 [44] Предполагается, что в общей сложности в дикой природе осталось 32 550 особей. [1] и около 100 живут в зоопарках. [29] |  | Масаи жираф ( Г. Типпельскирчи ) [37] Также известен как Килиманджаро. жираф | Масаи жираф ( G. Tippelskirchi ) Два подвида:
| |
| Жираф Торникрофта ( G.c. thornicrofti ) обитает только в долине Луангва на востоке Замбии. [1] На нем есть зубчатые и звездообразные пятна, которые могут проходить по ногам, а могут и не проходить. Средний размер самцов скромных размеров. [28] : 54 В дикой природе осталось не более 550 особей. [1] ни одного в зоопарках. [29] Он был назван в честь Гарри Скотта Торникрофта . [37] |  | Жираф Торникрофта (« Г. Торникрофти ») Также известен как жираф Луангва или родезийский жираф. | ||
Первым вымершим видом, который был описан, был Giraffa sivalensis Falconer and Cautley 1843, переоценка позвонка, который первоначально был описан как окаменелость живого жирафа. [45] Хотя по некоторым названиям может отсутствовать таксономическое мнение, опубликованные вымершие виды включают: [46]
- Грацилис жираф
- Жираф Джумаэ
- Жираф помело
- Присцилла жираф
- Жираф пенджабиенсис
- Жираф пигмейный
- Жираф сиваленсис
- Стиллей жираф
Анатомия

Взрослые жирафы имеют рост 4,3–5,7 м (14–19 футов), причем самцы выше самок. [47] Средний вес составляет 1192 кг (2628 фунтов) для взрослого мужчины и 828 кг (1825 фунтов) для взрослой женщины. [48] Несмотря на длинную шею и ноги, тело относительно короткое. [49] : 66 Кожа преимущественно серая. [48] или загар, [50] и может достигать толщины 20 мм (0,79 дюйма). [51] : 87 Длина 80–100 см (31–39 дюймов). [37] хвост заканчивается длинным темным пучком волос и используется для защиты от насекомых. [51] : 94
есть На шерсти темные пятна или пятна, которые могут быть оранжевыми, каштановыми , коричневыми или почти черными, окруженными светлыми волосами, обычно белыми или кремовыми . [52] С возрастом самцы жирафов темнеют. [44] рисунок шерсти Утверждается, что служит камуфляжем в светотенях лесов саванны. [37] Стоя среди деревьев и кустов, их трудно увидеть даже с расстояния нескольких метров. Однако взрослые жирафы передвигаются, чтобы лучше видеть приближающегося хищника, полагаясь на свой размер и способность защитить себя, а не на маскировку, которая может быть более важна для детенышей. [8] У каждого жирафа уникальный рисунок шерсти. [53] [54] Телята наследуют некоторые черты рисунка шерсти от своих матерей, а изменение некоторых признаков пятен коррелирует с выживаемостью теленка. [43] Кожа под пятнами может регулировать температуру тела животного, поскольку является местом расположения сложной системы кровеносных сосудов и крупных потовых желез . [55] Беспятнистые или однотонные жирафы встречаются очень редко, но их наблюдали. [56] [57]
Мех может обеспечить химическую защиту животного, поскольку репелленты от паразитов придают ему характерный запах. по меньшей мере 11 основных ароматических В мехе содержится индол и 3-метилиндол химических веществ, хотя за большую часть запаха ответственны . Поскольку самцы имеют более сильный запах, чем самки, они также могут выполнять сексуальную функцию . [58]
Голова

У обоих полов есть заметные роговидные структуры, называемые оссиконами , которые могут достигать 13,5 см (5,3 дюйма). Они образованы из окостеневшего хряща , покрыты кожей и срослись с черепом у теменных костей . [44] [51] : 95–97 Будучи васкуляризированными , оссиконы могут играть роль в терморегуляции . [55] и используются в бою между самцами. [59] Внешний вид является надежным показателем пола или возраста жирафа: оссиконы самок и молодых особей тонкие, с пучками волос на макушке, тогда как у взрослых самцов, как правило, лысые и узловатые на макушке. [44] В середине черепа появляется шишка, более заметная у мужчин. [19] У мужчин с возрастом появляются отложения кальция , которые образуют шишки на черепе. [52] Множественные пазухи облегчают череп жирафа. [51] : 103 Однако с возрастом их черепа становятся тяжелее и похожими на булаву, что помогает им стать более доминирующими в бою. [44] Затылочные мыщелки в нижней части черепа позволяют животному наклонять голову более чем на 90 градусов и хватать языком пищу на ветках прямо над ними. [51] : 103, 110 [19]
Глаза расположены по бокам головы, поэтому жираф имеет широкое поле зрения . благодаря своему большому росту [51] : 85, 102 По сравнению с другими копытными , зрение жирафа более бинокулярное , а глаза крупнее и имеют большую сетчатки . площадь поверхности [60] Жирафы видят в цвете, [51] : 85 их слух и обоняние остры. [52] Уши подвижные. [51] : 95 Ноздри имеют щелевидную форму, возможно, чтобы противостоять песчаным ветрам. [61] жирафа Язык имеет длину около 45 см (18 дюймов). Он черный, возможно, для защиты от солнечных ожогов, может захватывать листву и деликатно срывать листья. [51] : 109–110 Верхняя губа гибкая и покрыта волосками, защищающими от острых колючек. [19] имеется твердое небо На верхней челюсти вместо передних зубов . Моляры коронками и премоляры широкие, с невысокими на поверхности. [51] : 106
Шея
У жирафа чрезвычайно удлиненная шея, длина которой может достигать 2,4 м (7 футов 10 дюймов). [62] Вдоль шеи расположена грива из коротких, стоячих волос. [19] Шея обычно лежит под углом 50–60 градусов, хотя у молодых особей угол ближе к 70 градусам. [51] : 72–73 Длинная шея является результатом непропорционального удлинения шейных позвонков , а не добавления большего количества позвонков. Каждый шейный позвонок имеет длину более 28 см (11 дюймов). [49] : 71 Они составляют 52–54 процента длины позвоночного столба жирафа по сравнению с 27–33 процентами, типичными для аналогичных крупных копытных, включая ближайшего из ныне живущих родственников жирафа, окапи . [15] Это удлинение в основном происходит после рождения, возможно, потому, что матерям-жирафам было бы трудно родить детенышей с такими же пропорциями шеи, как у взрослых. [63] Голова и шея жирафа поддерживаются крупными мышцами и выйной связкой , которые закреплены длинными грудными позвонками, образующими горб. [19] [64] [37]

Шейные позвонки жирафа имеют шаровидные суставы . [49] : 71 Точка сочленения шейных и грудных позвонков жирафов смещена между первым и вторым грудными позвонками (Т1 и Т2), в отличие от большинства других жвачных животных, у которых сочленение находится между седьмым шейным позвонком (С7) и Т1. [15] [63] Это позволяет C7 напрямую способствовать увеличению длины шеи и дает основание предположить, что T1 на самом деле является C8 и что у жирафов появился дополнительный шейный позвонок. [64] Однако это предположение не является общепринятым, поскольку T1 имеет другие морфологические особенности, такие как сочленяющееся ребро , которое считается диагностическим для грудных позвонков, а также потому, что исключения из семи шейных позвонков у млекопитающих обычно характеризуются повышенными неврологическими аномалиями и заболеваниями. [15]
Существует несколько гипотез относительно эволюционного происхождения и поддержания удлинения шеи жирафа. [59] Чарльз Дарвин изначально предложил «гипотезу конкурирующих браузеров », которая была оспорена лишь недавно. Это предполагает, что конкурентное давление со стороны более мелких браузеров, таких как куду , стинбок и импала , способствовало удлинению шеи, поскольку это позволяло жирафам добывать пищу, которую конкуренты не могли. Это преимущество вполне реально, поскольку жирафы могут и действительно кормятся на высоте до 4,5 м (15 футов), в то время как даже довольно крупные конкуренты, такие как куду, могут кормиться на высоте всего около 2 м (6 футов 7 дюймов). [65] Есть также исследования, предполагающие, что конкуренция за просмотр интенсивна на более низких уровнях, и жирафы питаются более эффективно (набирая больше биомассы листьев с каждым глотком) высоко в кроне. [66] [67] Однако ученые расходятся во мнениях относительно того, сколько времени жирафы тратят на кормление на уровнях, недоступных для других браузеров. [12] [59] [65] [68] а исследование 2010 года показало, что взрослые жирафы с более длинной шеей на самом деле страдают от более высокой смертности в условиях засухи, чем их собратья с более короткой шеей. Это исследование показывает, что для поддержания более длинной шеи требуется больше питательных веществ, что подвергает жирафов с более длинной шеей риску во время нехватки еды. [69]
Другая теория, гипотеза полового отбора , предполагает, что длинная шея развилась как вторичный половой признак , давая самцам преимущество в состязаниях по «шейям» (см. ниже), чтобы установить доминирование и получить доступ к сексуально восприимчивым самкам. [12] В подтверждение этой теории некоторые исследования показали, что у мужчин шея длиннее и тяжелее, чем у женщин того же возраста. [12] [59] и что мужчины не используют другие формы боя. [12] Однако исследование 2024 года показало, что, хотя у самцов шея толще, у самок она на самом деле пропорционально длиннее, что, вероятно, связано с их большей потребностью находить больше еды, чтобы поддерживать себя и своих зависимых детенышей. [70] Также было высказано предположение, что шея служит для повышения бдительности животного. [71] [72]
Ноги, движение и осанка

Передние ноги обычно длиннее задних, [51] : 109 а у самцов передние ноги пропорционально длиннее, чем у самок, что дает им лучшую поддержку при повороте шеи во время драки. [70] В костях ног отсутствуют первая, вторая и пятая метаподии . [51] : 109 Похоже, что поддерживающая связка позволяет долговязым ногам выдерживать большой вес животного. [73] Копыта крупных жирафов-самцов достигают 31 × 23 см ( 12,2 × 9,1 дюйма) в диаметре. [51] : 98 Пучок ноги расположен низко к земле, что позволяет копыту лучше выдерживать вес животного. У жирафов отсутствуют прибылые пальцы и межпальцевые железы. Хотя таз относительно короткий, гребни подвздошной кости вытянуты. [19]
У жирафа только две походки : ходьба и галоп. Ходьба осуществляется путем перемещения ног с одной стороны тела, а затем с другой стороны. [44] При галопе задние ноги обходят передние, прежде чем последние двинутся вперед. [52] и хвост свернется. [44] Движения головы и шеи обеспечивают баланс и контроль инерции во время галопа. [34] : 327–29 Жираф может развивать скорость до 60 км/ч (37 миль в час). [74] и может выдерживать скорость 50 км/ч (31 миль в час) на протяжении нескольких километров. [75] Жирафы, вероятно, не были бы умелыми пловцами, поскольку их длинные ноги были бы очень громоздкими в воде. [76] хотя они могут плавать. [77] При плавании передние ноги отягощают грудную клетку, что затрудняет гармоничное движение шеи и ног. [76] [77] или держать голову над поверхностью воды. [76]
Жираф отдыхает, лежа, положив тело на сложенные ноги. [34] : 329 Чтобы лечь, животное становится на колени на передние ноги, а затем опускает остальную часть тела. Чтобы снова подняться, он сначала встает на передние колени и кладет заднюю часть на задние ноги. Затем он тянет заднюю часть вверх, и передние ноги снова выпрямляются. На каждом этапе животное качает головой для равновесия. [51] : 67 Если жираф хочет напиться, он либо раздвигает передние ноги, либо сгибает колени. [44] Исследования в неволе показали, что жираф спит с перерывами около 4,6 часов в день, в основном ночью. Обычно он спит лежа; однако были зарегистрированы случаи сна стоя, особенно у пожилых людей. Прерывистые короткие фазы «глубокого сна» во время лежания характеризуются тем, что жираф сгибает шею назад и кладет голову на бедро или бедро - положение, которое, как полагают, указывает на парадоксальный сон . [78]
Внутренние системы

У млекопитающих левый возвратный гортанный нерв длиннее правого; у жирафа он длиннее более чем на 30 см (12 дюймов). Эти нервы у жирафа длиннее, чем у любого другого живого животного; [79] длина левого нерва превышает 2 м (6 футов 7 дюймов). [80] Каждая нервная клетка на этом пути начинается в стволе мозга и проходит вниз по шее вдоль блуждающего нерва , затем разветвляется в возвратный гортанный нерв, который проходит обратно вверх по шее в гортань. Таким образом, у самых крупных жирафов эти нервные клетки имеют длину почти 5 м (16 футов). [79] Несмотря на длинную шею и большой череп, мозг жирафа типичен для копытного. [81] Потеря тепла в результате испарения в носовых проходах сохраняет мозг жирафа прохладным. [55] Форма скелета придает жирафу небольшой объем легких по сравнению с его массой. Его длинная шея оставляет ему много мертвого пространства , несмотря на узкое дыхательное горло. У жирафа также высокий дыхательный объем , поэтому баланс мертвого пространства и дыхательного объема практически такой же, как и у других млекопитающих. Животное все еще может обеспечивать достаточное количество кислорода для своих тканей, а также может увеличивать частоту дыхания и диффузию кислорода во время бега. [82]

жирафа Кровеносная система имеет несколько приспособлений, компенсирующих его большой рост. [16] весом 11 кг (25 фунтов) и размером 60 см (2 фута) Его сердце должно создавать примерно вдвое большее кровяное давление, необходимое человеку для поддержания притока крови к мозгу. Таким образом, толщина стенки сердца может достигать 7,5 см (3,0 дюйма). [52] У жирафов относительно высокая частота пульса для их размера — 150 ударов в минуту. [49] : 76 Когда животное опускает голову, кровь почти беспрепятственно устремляется вниз, а rete mirabile в верхней части шеи с большой площадью поперечного сечения предотвращает избыточный приток крови к мозгу. Когда он снова поднимается, кровеносные сосуды сужаются и выталкивают кровь в мозг, чтобы животное не потеряло сознание. [83] Яремные вены содержат несколько (чаще всего семь) клапанов, которые предотвращают обратный ток крови в голову из нижней полой вены и правого предсердия , когда голова опущена. [84] И наоборот, кровеносные сосуды голеней находятся под большим давлением из-за давления на них жидкости. Чтобы решить эту проблему, кожу голеней делают толстой и плотной, что не позволяет слишком большому количеству крови вливаться в них. [37]
У жирафов мышцы пищевода достаточно сильны, чтобы обеспечить срыгивание пищи из желудка вверх по шее и в рот для пережевывания пищи . [49] : 78 У них четырехкамерный желудок , приспособленный к их специализированной диете. [19] Кишечник взрослого жирафа имеет длину более 70 м (230 футов) и имеет относительно небольшое соотношение тонкой и толстой кишки . [85] У жирафа маленькая компактная печень. [49] : 76 У плода может быть небольшой желчный пузырь , который исчезает еще до рождения. [19] [86] [87]
Поведение и экология
Среда обитания и питание

Жирафы обычно обитают в саваннах и редколесьях . Они предпочитают участки, где преобладают акации , коммифоры , комбретумы и терминалии деревья , а не брахистегии , расположенные более плотно. [34] : 322 Ангольского жирафа можно встретить в пустыне. [88] Жирафы грызут ветки деревьев, предпочитая ветки подсемейства Acacieae и родов Commiphora и Terminalia . [89] которые являются важными источниками кальция и белка для поддержания темпов роста жирафа. [8] Они также питаются кустарниками, травой и фруктами. [34] : 324 Жираф съедает около 34 кг (75 фунтов) растительных веществ ежедневно. [44] В состоянии стресса жирафы могут грызть большие ветки, сдирая с них кору. [34] : 325 Также зафиксировано, что жирафы жуют старые кости . [51] : 102
В сезон дождей еды много, и жирафы более рассредоточены, а в засушливый сезон они собираются вокруг оставшихся вечнозеленых деревьев и кустов. [89] Матери, как правило, кормятся на открытых местах, предположительно, чтобы было легче обнаружить хищников, хотя это может снизить эффективность их кормления. [68] Будучи жвачным животным , жираф сначала пережевывает пищу, затем заглатывает ее для переработки, а затем пропускает полупереваренную жвачку вверх по шее и обратно в рот, чтобы снова пережевать. [49] : 78–79 Жирафу требуется меньше пищи, чем многим другим травоядным животным, потому что листва, которую он поедает, содержит больше концентрированных питательных веществ и у него более эффективная пищеварительная система. [89] Фекалии животного имеют вид мелких гранул. [19] Когда у жирафа есть доступ к воде, он может обходиться без воды не более трех дней. [44]
Жирафы оказывают большое влияние на деревья, которыми питаются, задерживая рост молодых деревьев на несколько лет и придавая «талию» слишком высоким деревьям. Пик кормления приходится на первые и последние часы дня. В эти часы жирафы в основном стоят и размышляют. Размышление является доминирующим занятием в ночное время, когда оно в основном осуществляется лежа. [44]
Социальная жизнь

Жирафы обычно образуют группы, размер и состав которых различаются в зависимости от экологических, антропогенных, временных и социальных факторов. [90] Традиционно состав этих групп описывался как открытый и постоянно меняющийся. [91] В исследовательских целях «группа» определяется как «совокупность особей, находящихся на расстоянии менее километра друг от друга и движущихся в одном направлении». [92] Более поздние исследования показали, что жирафы имеют устойчивые социальные группы или клики, основанные на родстве, поле или других факторах, и эти группы регулярно общаются с другими группами в более крупных сообществах или подсообществах внутри общества разделения-слияния . [93] [94] [95] [96] Близость к людям может нарушить социальные механизмы. [93] Жирафы Масаи в Танзании разделяются на разные субпопуляции по 60–90 взрослых самок с перекрывающимися ареалами, каждая из которых различается уровнем воспроизводства и смертностью детенышей. [97] Рассредоточение имеет предвзятое отношение к мужчинам и может включать пространственное и/или социальное рассредоточение. [98] Субпопуляции взрослых самок связаны самцами в суперсообщества, насчитывающие около 300 животных. [99]

Число жирафов в группе может колебаться от одного до 66 особей. [90] [96] Группы жирафов, как правило, разделены по полу. [96] хотя встречаются и смешанные группы, состоящие из взрослых женщин и молодых мужчин. [92] Женские группы могут быть связаны по материнской линии . [96] Как правило, женщины более избирательны, чем мужчины, при выборе людей того же пола, с которыми они общаются. [95] Особенно стабильными являются группы жирафов, состоящие из самок и их детенышей. [92] который может длиться недели или месяцы. [100] Молодые самцы также образуют группы и участвуют в игровых боях. Однако по мере взросления самцы становятся более одинокими, но могут также объединяться в пары или группы самок. [96] [100] Жирафы не территориальны , [19] но у них есть ареалы обитания , которые различаются в зависимости от количества осадков и близости к населенным пунктам. [101] Жирафы-самцы иногда уходят далеко от мест, где они обычно часто бывают. [34] : 329
Ранние биологи предполагали, что жирафы немы и неспособны создать достаточный поток воздуха для вибрации своих голосовых связок . [102] Доказано обратное; Было зарегистрировано, что они общаются, используя фырканье, чихание, кашель, храп, шипение, взрывы, стоны, ворчание, рычание и звуки, похожие на звуки флейты. [44] [102] Во время ухаживания самцы издают громкий кашель. Самки призывают своих детенышей ревом. Телята издают блеяние, мычание и мяуканье. [44] Фырканье и шипение ассоциируются с бдительностью. [103] Ночью жирафы, кажется, напевают друг другу. [104] Есть некоторые свидетельства того, что жирафы используют резонанс Гельмгольца для создания инфразвука . [105] Они также общаются с помощью языка тела. Доминирующие самцы демонстрируют другим самцам прямую осанку; держа подбородок и голову вверх при скованной ходьбе и показывая бок. Менее доминирующие демонстрируют покорность, опуская голову и уши, опуская подбородок и убегая. [44]
Размножение и родительский уход

Размножение жирафов в целом полигамно : несколько самцов старшего возраста спариваются с плодовитыми самками. [92] Самки могут воспроизводить потомство в течение всего года, а эструс повторяется примерно каждые 15 дней. [106] [107] Самки жирафов в период течки рассредоточены в пространстве и времени, поэтому репродуктивные взрослые самцы принимают стратегию блуждания среди групп самок в поисках возможностей для спаривания с периодическим гормонально-индуцированным гоном примерно каждые две недели. [108] Самцы предпочитают молодых взрослых самок подросткам и пожилым людям. [92]
Самцы жирафов оценивают фертильность самок, пробуя на вкус мочу самки, чтобы обнаружить эструс, в рамках многоэтапного процесса, известного как реакция Флемена . [92] [100] Как только течная самка обнаружена, самец попытается ухаживать за ней. Во время ухаживания доминирующие самцы будут держать подчиненных на расстоянии. [100] Ухаживающий самец может лизнуть хвост самки, положить голову и шею на ее тело или подтолкнуть ее своими оссиконами. Во время совокупления самец стоит на задних лапах, подняв голову, а передние ноги опираются на бока самки. [44]
жирафа Беременность длится 400–460 дней, после чего обычно рождается один теленок, хотя в редких случаях случаются двойни. [106] Мать рожает стоя. Теленок выходит вперед головой и передними ногами, прорвав плодные оболочки , и падает на землю, перерезав пуповину . [19] Рост новорожденного жирафа составляет 1,7–2 м (5 футов 7 дюймов – 6 футов 7 дюймов). [47] Уже через несколько часов после рождения теленок может бегать и почти неотличим от недельного ребенка. Однако в течение первых одной-трех недель он проводит большую часть времени, прячась. [109] его рисунок шерсти обеспечивает маскировку. Оссиконы, пролежавшие в утробе матери, поднимаются через несколько дней. [44]

Матери с телятами собираются в стада на доращивании, вместе передвигаются или пасутся. Матери в такой группе могут иногда оставлять своих телят с одной самкой, пока они кормятся и пьют в другом месте. Это известно как « бассейн отела ». [109] Телята подвергаются риску нападения хищников, и мать-жираф стоит над ними и пинает приближающегося хищника. [44] Самки, наблюдающие за прудами для отела, предупредят своих детенышей только в том случае, если обнаружат беспокойство, хотя остальные заметят это и последуют за ними. [109] Алло-сосание, когда детеныш кормит грудью не свою мать, а самку, было зарегистрировано как у диких жирафов, так и у жирафов, содержащихся в неволе. [110] [111] Телята впервые жуют грудь в четыре-шесть месяцев и перестают есть в шесть-восемь месяцев. Молодые могут не достичь независимости, пока им не исполнится 14 месяцев. [51] : 49 Самки способны к размножению в возрасте четырех лет. [44] тогда как сперматогенез у самцов начинается в возрасте трех-четырех лет. [112] Самцам приходится ждать, пока им исполнится хотя бы семь лет, чтобы получить возможность спариваться. [44]
Выравнивание

Самцы жирафов используют свои шеи в качестве оружия в бою, такое поведение известно как «обнимание». Охватывание используется для установления доминирования, и самцы, побеждающие в схватках, добиваются большего репродуктивного успеха . [12] Такое поведение происходит при низкой или высокой интенсивности. При обхвате с низкой интенсивностью бойцы трутся и опираются друг на друга. Самец, который сможет держаться более вертикально, побеждает в схватке. При интенсивном схватке бойцы расставляют передние ноги и размахивают шеями друг друга, пытаясь нанести удары своими оссиконами. Участники будут стараться уклоняться от ударов друг друга, а затем готовиться к ответному удару. Сила удара зависит от веса черепа и дуги замаха. [44] Поединок на шее может длиться более получаса, в зависимости от того, насколько хорошо подобраны бойцы. [34] : 331 Хотя большинство драк не приводят к серьезным травмам, имеются записи о сломанных челюстях, сломанных шеях и даже смертельных случаях. [12]
После дуэли два самца жирафа обычно ласкают и ухаживают друг за другом. Было обнаружено, что такие взаимодействия между мужчинами встречаются чаще, чем гетеросексуальные связи. [113] По данным одного исследования, до 94 процентов наблюдаемых инцидентов с ростом числа случаев насилия имели место между мужчинами. Доля однополых связей колебалась от 30 до 75 процентов. Только один процент однополых инцидентов произошел между женщинами. [114]
Смертность и здоровье

Жирафы имеют высокую вероятность выживания взрослых особей. [115] и необычно долгая продолжительность жизни по сравнению с другими жвачными животными - до 38 лет. [116] Выживаемость взрослых самок существенно коррелирует с количеством социальных объединений. [117] Благодаря своим размерам, зрению и мощным ударам взрослые жирафы в основном защищены от хищников. [44] львы являются их единственной серьезной угрозой. [51] : 55 Телята гораздо более уязвимы, чем взрослые особи, и на них также охотятся леопарды , пятнистые гиены и дикие собаки . [52] От четверти до половины детенышей жирафов достигают взрослой жизни. [115] [118] Выживаемость телят варьируется в зависимости от сезона рождения: телята, рожденные в засушливый сезон, имеют более высокие показатели выживаемости. [119]
Местное сезонное присутствие больших стад мигрирующих антилоп гну и зебр снижает давление хищников на детенышей жирафов и увеличивает вероятность их выживания. [120] В свою очередь, было высказано предположение, что другим копытным может быть полезно общение с жирафами, поскольку их рост позволяет им замечать хищников издалека. Было обнаружено, что зебры оценивают риск нападения хищников, наблюдая за жирафами, и тратят меньше времени на осмотр вокруг, когда жирафы присутствуют. [121]

Некоторые паразиты питаются жирафами. Они часто являются хозяевами клещей , особенно в области гениталий, где кожа тоньше, чем на других участках. Виды клещей, которые обычно питаются жирафами, относятся к родам Hyalomma , Amblyomma и Rhipicephalus . Жирафы могут полагаться на красноклювых и желтоклювых быков, которые очищают их от клещей и предупреждают об опасности. Жирафы являются носителями множества видов внутренних паразитов и восприимчивы к различным заболеваниям. Они стали жертвами (ныне искорененной) вирусной болезни чумы крупного рогатого скота . [19] Жирафы также могут страдать от заболеваний кожи, которые проявляются в виде морщин, повреждений или трещин . симптомы болезни наблюдаются у 79% жирафов В национальном парке Руаха она не стала причиной смертности , однако в Тарангире и менее распространена в районах с плодородными почвами. [122] [123] [124]
Человеческие отношения
Культурное значение
Жираф, долговязый и пятнистый, вызывал восхищение на протяжении всей истории человечества, а его образ широко распространен в культуре. Он олицетворял гибкость, дальновидность, женственность, хрупкость, пассивность, грацию, красоту и сам африканский континент. [125] : 7, 116

Жирафы изображались в искусстве по всему африканскому континенту, в том числе у киффианцев , египтян и кушитов . [125] : 45–47 Киффианцы создали наскальную гравюру двух жирафов в натуральную величину, датированную 8000 лет назад, которую назвали «крупнейшим в мире петроглифом наскального искусства». [125] : 45 [126] То, как жираф добился такого роста, было предметом различных африканских сказок . [12] Народ тугенов в современной Кении использовал жирафа, чтобы изобразить своего бога Мда. [127] Египтяне дали жирафу собственный иероглиф ; «ср» в древнеегипетском языке и «мми» в более поздние периоды. [125] : 49
Жирафы присутствуют в современной западной культуре . Сальвадор Дали изображал их с горящими гривами на некоторых картинах сюрреалистов. Дали считал жирафа мужским символом. Пылающий жираф должен был быть «космическим апокалиптическим монстром мужского пола». [125] : 123 Жираф упоминается в нескольких детских книгах, в том числе в «Жирафе, который боялся высоты» Дэвида А. Уфера , » Джайлза Андреа « Жирафы не умеют танцевать и Роальда Даля «Жираф, Пелли и я» . Жирафы появлялись в анимационных фильмах в качестве второстепенных персонажей в диснеевских фильмах «Дамбо » и «Король Лев» , а также в более заметных ролях в фильмах «Дикая природа» и « Мадагаскар» . Жираф Софи была популярным прорезывателем с 1961 года. Еще один известный вымышленный жираф - талисман Toys «R» Us Жираф Джеффри . [125] : 127
Жирафа также использовали для некоторых научных экспериментов и открытий. Ученые использовали свойства кожи жирафа в качестве модели для костюмов космонавтов и летчиков-истребителей, поскольку людям этих профессий грозит потеря сознания, если кровь прильет к ногам. [49] : 76 Ученые-компьютерщики смоделировали рисунок шерсти нескольких подвидов, используя механизмы реакции-диффузии . [128] Созвездие Жирафа , введенное в 17 веке , изображает жирафа. [125] : 119–20 Народ тсвана в Ботсване традиционно рассматривает созвездие Креста как двух жирафов: Акрукс и Мимоза, образующие самца, а Гакрукс и Дельта Креста, образующие самку. [129]

плен
Египтяне были одними из первых людей, которые держали жирафов в неволе и отправляли их по Средиземному морю. [125] : 48–49 Жираф был среди многих животных, собранных и выставленных римлянами. Первую в Рим привез Юлий Цезарь в 46 г. до н.э. [125] : 52 С падением Западной Римской империи численность жирафов в Европе сократилась. [125] : 54 В средние века жирафы были известны европейцам благодаря контактам с арабами, которые почитали жирафов за их своеобразный внешний вид. [52]
На протяжении всей истории отдельные плененные жирафы получали статус знаменитостей. жирафа из Малинди привез в Китай В 1414 году путешественник Чжэн Хэ и поместил его в зоопарк династии Мин . Животное было источником восхищения для китайцев, которые связывали его с мифическим Цилинем . [125] : 56 Жираф Медичи был жирафом, подаренным Лоренцо Медичи в 1486 году. По прибытии во Флоренцию он вызвал большой резонанс . [130] Зарафа , еще один знаменитый жираф, был привезен из Египта в Париж в начале 19 века в качестве подарка французскому королю Карлу X. Сенсация: жираф стал предметом многочисленных памятных вещей или «жирафаналий». [125] : 81
Жирафы стали популярной достопримечательностью в современных зоопарках , хотя сохранить их здоровье сложно, поскольку им требуются огромные площади и они должны поедать большое количество травы. У содержащихся в неволе жирафов в Северной Америке и Европе уровень смертности выше, чем в дикой природе, причем наиболее распространенными причинами являются плохое содержание, питание и управление. [51] : 153 Жирафы в зоопарках демонстрируют стереотипное поведение , особенно облизывание неодушевленных предметов и хождение. [51] : 164 Владельцы зоопарков могут предлагать различные мероприятия, чтобы стимулировать жирафов, в том числе обучать их брать еду у посетителей. [51] : 167, 176 Конюшни для жирафов строят особенно высокими, чтобы соответствовать их росту. [51] : 183
Эксплуатация
Жирафы, вероятно, были обычной целью охотников по всей Африке. [34] : 337 Разные части их тел использовались для разных целей. [19] Их мясо употреблялось в пищу. Волосы на хвосте представляли собой мухобойки , браслеты, ожерелья и нити. Щиты, сандалии и барабаны делались из кожи, а струны музыкальных инструментов — из сухожилий. [19] [34] : 337 В Буганде дым горящей шкуры жирафа традиционно использовался для лечения носовых кровотечений. [34] : 337 Жители хумр Кордофана печени употребляют напиток Умм Нёлох, который готовят из и костного мозга жирафов. Ричард Радгли предположил, что Умм Нёлох может содержать ДМТ . [131] , являются призраками жирафов Говорят, что напиток вызывает галлюцинации жирафов, которые, по мнению Хумра . [132]
Статус сохранения
признал жирафов уязвимыми с точки зрения сохранения. В 2016 году МСОП [1] По оценкам, в 1985 году в дикой природе насчитывалось 155 000 жирафов. В 1999 году это число снизилось до более чем 140 000. [133] насчитывается около 97 500 особей жирафов . По оценкам на 2016 год, в дикой природе [134] [135] Масаи и сетчатые подвиды находятся под угрозой исчезновения . [136] [137] а подвид Ротшильдов находится под угрозой исчезновения . [33] Нубийский подвид находится под угрозой исчезновения . [138]

Основными причинами сокращения популяции жирафов являются утрата среды обитания и прямое уничтожение рынков мяса диких животных . Жирафы были истреблены на большей части своего исторического ареала, включая Эритрею , Гвинею , Мавританию и Сенегал . [1] Возможно, они также исчезли из Анголы , Мали и Нигерии , но были завезены в Руанду и Эсватини . [1] [138] По состоянию на 2010 год [update] , в неволе содержалось более 1600 особей В зоопарках, зарегистрированных Species360 . [29] Разрушение среды обитания нанесло вред жирафу. В Сахеле потребность в дровах и пастбищах для скота привела к вырубке лесов . Обычно жирафы могут сосуществовать с домашним скотом, поскольку они избегают прямой конкуренции, кормясь над ними. [37] В 2017 году сильная засуха на севере Кении привела к усилению напряженности из-за земель и истреблению диких животных пастухами, особенно пострадали популяции жирафов. [139]
Охраняемые территории, такие как национальные парки, обеспечивают важную среду обитания и от браконьерства . защиту популяций жирафов [1] Природоохранные усилия местных сообществ за пределами национальных парков также эффективны для защиты жирафов и их среды обитания. [140] [141] Частные заповедники внесли свой вклад в сохранение популяций жирафов в восточной и южной Африке. [37] Жираф является охраняемым видом на большей части своего ареала. Это национальное животное Танзании. [142] и охраняется законом, [143] а несанкционированное убийство может привести к тюремному заключению. [144] Поддержанная ООН Конвенция по мигрирующим видам отобрала жирафов для защиты в 2017 году. [145] В 2019 году жирафы были внесены в список Приложения II к Конвенции о международной торговле видами, находящимися под угрозой исчезновения ( СИТЕС ), что означает, что международная торговля, в том числе частями/производными, регулируется. [146]
Транслокации иногда используются для увеличения или восстановления уменьшенных или истребленных популяций, но эти действия рискованны и их трудно осуществить, используя лучшие практики обширных исследований до и после транслокации и обеспечения жизнеспособной основной популяции. [147] [148] Аэрофотосъемка является наиболее распространенным методом мониторинга тенденций численности жирафов на обширных бездорожных территориях африканских ландшафтов, но известно, что с помощью воздушных методов численность жирафов не учитывается. Методы наземных исследований более точны и могут использоваться в сочетании с воздушными съемками для точной оценки размеров и тенденций популяций. [149]
См. также
- Фауна Африки
- Центр Жирафов
- Giraffe Manor - отель в Найроби с жирафами
Ссылки
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и Мюллер, З.; Беркович, Ф.; Бранд, Р.; Браун, Д.; Браун, М.; Болджер, Д.; Картер, К.; Дьякон, Ф.; Доэрти, Дж.Б.; Феннесси, Дж.; Феннесси, С.; Хусейн, А.А.; Ли, Д.; Марэ, А.; Штраус, М.; Татчингс, А.; Вубе, Т. (2018) [дополненная версия оценки 2016 года]. « Жираф камелопардалис » . Красный список исчезающих видов МСОП . 2018 : e.T9194A136266699. doi : 10.2305/IUCN.UK.2016-3.RLTS.T9194A136266699.en . Проверено 12 ноября 2021 г.
- ^ Перейти обратно: а б «Жираф» . Интернет-словарь этимологии . Архивировано из оригинала 19 марта 2015 года . Проверено 1 ноября 2011 г.
- ^ Деххода, Али-Акбар. "Душа" . Институт лексики Деххода и Международный центр персидских исследований . Архивировано из оригинала 28 декабря 2022 года . Проверено 28 декабря 2022 г.
- ^ Ачарян, Грачеай (1973). «ւրափէ» [в Hayerēn amatakan baṙaran [Армянский этимологический словарь] (на армянском языке), том II, 2-е издание, переиздание оригинального семитомного издания 1926–1935 годов]. www.nayiri.com . Ереван: Университетское издательство. п. 110а. Архивировано из оригинала 28 декабря 2022 года . Проверено 28 декабря 2022 г.
- ^ Эрси, Тибор (2006). Французское лингвистическое влияние в хлопковой версии путешествий Мандевиля . Издательство «Инк». п. 113. ИСБН 9789637094545 . Архивировано из оригинала 22 сентября 2023 года . Проверено 20 апреля 2021 г.
- ^ «Определение ВЕРБЛЮДА» . mw.com . Британская энциклопедия: Мерриам-Вебстер. Архивировано из оригинала 25 апреля 2009 года . Проверено 3 сентября 2014 г.
- ^ «Определение камелопарда» . Словарь средневековых терминов и фраз . Архивировано из оригинала 4 сентября 2014 года . Проверено 3 сентября 2014 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л Митчелл, Г.; Скиннер, доктор медицинских наук (2003). «О происхождении, эволюции и филогении жирафов Giraffa Camelopardalis ». Труды Королевского общества Южной Африки . 58 (1): 51–73. Бибкод : 2003TRSSA..58...51M . дои : 10.1080/00359190309519935 . S2CID 6522531 .
- ^ Чен, Л.; Цю, К.; Цзян, Ю.; Ван, К. (2019). «Крупномасштабное секвенирование генома жвачных животных дает представление об их эволюции и отличительных чертах» . Наука . 364 (6446): eaav6202. Бибкод : 2019Sci...364.6202C . дои : 10.1126/science.aav6202 . ПМИД 31221828 .
- ^ Перейти обратно: а б с д Дановиц, М.; Васильев А.; Кортландт, В.; Солуниас, В. (2015). «Ископаемые свидетельства и стадии удлинения шеи Giraffa Camelopardalis » . Королевское общество открытой науки . 2 (10): 150393. Бибкод : 2015RSOS....250393D . дои : 10.1098/rsos.150393 . ПМЦ 4632521 . ПМИД 26587249 .
- ^ Перейти обратно: а б Дановиц, М.; Домальский, Р.; Солуниас, Н. (2015). «Анатомия шейки матки Samotherium , жирафа с промежуточной шеей» . Королевское общество открытой науки . 2 (11): 150521. Бибкод : 2015RSOS....250521D . дои : 10.1098/rsos.150521 . ПМЦ 4680625 . ПМИД 26716010 .
- ^ Перейти обратно: а б с д и ж г час Симмонс, Р.Э.; Шиперс, Л. (1996). «Победа за шею: половой отбор в эволюции жирафа» (PDF) . Американский натуралист . 148 (5): 771–786. дои : 10.1086/285955 . S2CID 84406669 . Архивировано из оригинала (PDF) 23 августа 2004 года.
- ^ Янис, CM (1993). «Третичная эволюция млекопитающих в контексте изменения климата, растительности и тектонических событий». Ежегодный обзор экологии и систематики . 24 : 467–500. дои : 10.1146/annurev.ecolsys.24.1.467 . JSTOR 2097187 .
- ^ Рамштайн, Г.; Флюто, Ф.; Бесс, Дж.; Жуссом, С. (1997). «Влияние складчатости, движения плит и распределения суши и моря на изменение климата в Евразии за последние 30 миллионов лет». Природа . 386 (6627): 788–795. Бибкод : 1997Natur.386..788R . дои : 10.1038/386788a0 . S2CID 4335003 .
- ^ Перейти обратно: а б с д Бадлангана, Луизиана; Адамс, Дж.В.; Менеджер, PR (2009). жирафа (Giraffa Camelopardalis) « Шейный позвоночник : эвристический пример в понимании эволюционных процессов?» . Зоологический журнал Линнеевского общества . 155 (3): 736–757. дои : 10.1111/j.1096-3642.2008.00458.x .
- ^ Перейти обратно: а б Холмс, Б. (2021). «Внимание! Секреты сердечно-сосудистой системы жирафов» . Знающий журнал . doi : 10.1146/knowable-051821-2 . S2CID 236354545 . Архивировано из оригинала 6 июля 2022 года . Проверено 1 августа 2022 г.
- ^ Агаба, М.; Ишенгома, Э.; Миллер, туалет; МакГрат, Британская Колумбия; Хадсон, Китай; Бедоя, Китайская республика; Ратан, А.; Бурханс, Р.; Чихи, Р.; Медведев П.; Праул Калифорния; Ву-Кавенер, Л.; Вуд, Б.; Робертсон, Х.; Пенфолд, Л.; Кавенер, ДР (2016). «Последовательность генома жирафа раскрывает ключ к его уникальной морфологии и физиологии» . Природные коммуникации . 7 : 11519. Бибкод : 2016NatCo...711519A . дои : 10.1038/ncomms11519 . ПМЦ 4873664 . ПМИД 27187213 .
- ^ Перейти обратно: а б Браун, DM; Бреннеман Р.А.; Кёпфли, К.-П.; Поллинджер, JP; Мила, Б.; Георгиадис, Нью-Джерси; Луис-младший, Э.Э.; Гретер, ГФ; Джейкобс, ДК; Уэйн РК (2007). «Обширная популяционная генетическая структура жирафа» . БМК Биология . 5 (1): 57. дои : 10.1186/1741-7007-5-57 . ПМК 2254591 . ПМИД 18154651 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Дагг, А.И. (1971). « Жираф камелопардалис » (PDF) . Виды млекопитающих (5): 1–8. дои : 10.2307/3503830 . JSTOR 3503830 . Архивировано из оригинала (PDF) 21 апреля 2017 года . Проверено 25 октября 2011 г.
- ^ Гипполити, С. (2018). «Влияние таксономической инерции на сохранение разнообразия африканских копытных: обзор» . Биологические обзоры . 93 (1): 115–130. дои : 10.1111/brv.12335 . ПМИД 28429851 . S2CID 5189968 . Архивировано из оригинала 3 апреля 2021 года . Проверено 21 апреля 2021 г.
- ^ Перейти обратно: а б Гроувс, К.; Грабб, П. (2011). Систематика копытных . JHU Пресс . стр. 68–70. ISBN 9781421400938 . Архивировано из оригинала 22 сентября 2023 года . Проверено 17 октября 2020 г.
- ^ Перейти обратно: а б с Феннесси, Дж.; Бидон, Т.; Ройсс, Ф.; Кумар, В.; Элкан, П.; Нильссон, Массачусетс; Вамбергер, М.; Фриц, У.; Янке, А. (2016). «Мультилокусный анализ выявил четыре вида жирафов вместо одного» . Современная биология . 26 (18): 2543–2549. Бибкод : 2016CBio...26.2543F . дои : 10.1016/j.cub.2016.07.036 . ПМИД 27618261 . S2CID 3991170 .
- ^ Перейти обратно: а б Петцольд, А.; Хасанин, А. (2020). «Сравнительный подход к разграничению видов, основанный на нескольких методах анализа последовательностей мультилокусной ДНК: тематическое исследование рода Giraffa (Mammalia, Cetartiodactyla)» . ПЛОС Один . 15 (2): e0217956. Бибкод : 2020PLoSO..1517956P . дои : 10.1371/journal.pone.0217956 . ПМК 7018015 . ПМИД 32053589 .
- ^ Перейти обратно: а б с Коимбра, РТФ; Зима, С.; Кумар, В.; Кёпфли, К.-П.; Гули, Р.М.; Добрынин П.; Феннесси, Дж.; Янке, А. (2021). «Полногеномный анализ жирафа позволяет выявить четыре различных вида» . Современная биология . 31 (13): 2929–2938.e5. Бибкод : 2021CBio...31E2929C . дои : 10.1016/j.cub.2021.04.033 . ПМИД 33957077 .
- ^ Перейти обратно: а б Бертола, LD; Куинн, Л; Ханхой, К; Гарсия-Эрилл, Э; Расмуссен, С.М.; Мейснер, Дж; Беггильд, Т; Ван, X; Лин, Л; Нурсифа, С; Лю, Х; Ли, З; Чеге, М; Мудли, Ю; Брюниче-Ольсен, А; Куджа, Дж; Шуберт, М; Агаба, М; Сантандер, Германия; Синдинг, М.Х. С.; Муваника, В; Масембе, К; Зигизмунд, HR; Мольтке, Я; Альбрехцен, А; Хеллер, Р. (2024). «Линия жирафов сформирована в результате крупных древних событий примеси» . Современная биология . 34 (7): 1576–1586.e5. Бибкод : 2024CBio...34E1576B . дои : 10.1016/j.cub.2024.02.051 . ПМИД 38479386 .
- ^ Перейти обратно: а б Линней, К. (1758). Система Природы .
- ^ Перейти обратно: а б с Хасанин А.; Ропике, А.; Гурман, Б.-Л.; Шардонне, Б.; Ригуле, Ж. (2007). «Изменчивость митохондриальной ДНК у Giraffa Camelopardalis : последствия для таксономии, филогеографии и сохранения жирафов в Западной и Центральной Африке». Comptes Rendus Biologies . 330 (3): 173–183. дои : 10.1016/j.crvi.2007.02.008 . ПМИД 17434121 .
- ^ Перейти обратно: а б с д и ж г час я Сеймур, Р. (2001). Закономерности подвидового разнообразия жирафа Giraffa Camelopardalis (L. 1758): сравнение систематических методов и их значение для природоохранной политики (докторская диссертация).
- ^ Перейти обратно: а б с д и ж г час «Жираф» . ИГИЛ . 2010. Архивировано из оригинала 6 июля 2010 года . Проверено 4 ноября 2010 г.
- ^ Swaison 1835. Camelopardalis antiquorum. Баггер-эль-Хомер, Кордофан, около 10° с.ш., 28° в.д. (фиксировано Харпером, 1940 г.)
- ^ «Экспонаты» . Зоопарк Аль-Айна . 25 февраля 2003 г. Архивировано из оригинала 29 ноября 2011 г. Проверено 21 ноября 2011 г.
- ^ «Нубийский жираф родился в зоопарке Аль-Айна» . ОАЭ Интеракт. Архивировано из оригинала 20 марта 2012 года . Проверено 21 декабря 2010 г.
- ^ Перейти обратно: а б Феннесси, С.; Феннесси, Дж.; Мюллер, З.; Браун, М.; Марэ, А. (2018). « Жираф камелопардалис вида Ротшильди » . Красный список исчезающих видов МСОП . 2018 : e.T174469A51140829. doi : 10.2305/IUCN.UK.2018-2.RLTS.T174469A51140829.en . Проверено 19 ноября 2021 г.
- ^ Перейти обратно: а б с д и ж г час я дж к Кингдон, Дж. (1988). Млекопитающие Восточной Африки: Атлас эволюции Африки . Том. 3, Часть Б: Крупные млекопитающие. Издательство Чикагского университета . стр. 313–337. ISBN 978-0-226-43722-4 .
- ^ Феннесси, Дж.; Марэ, А.; Татчингс, А. (2018). « Жираф камелопардалис подвид перальта » . Красный список исчезающих видов МСОП . 2018 : e.T136913A51140803.
- ^ Винтон, МЫ де (1899). «XXVIII.—О млекопитающих, собранных подполковником В. Гиффардом на северной территории Золотого Берега» . Анналы и журнал естественной истории . 4 (2 3): 353–359. дои : 10.1080/00222939908678212 . ISSN 0374-5481 . Архивировано из оригинала 1 августа 2020 года . Проверено 16 сентября 2019 г.
- ^ Перейти обратно: а б с д и ж г час я Пеллоу, Р.А. (2001). «Жираф и Окапи». В Макдональде, Д. (ред.). Энциклопедия млекопитающих (2-е изд.). Издательство Оксфордского университета . стр. 520–527. ISBN 978-0-7607-1969-5 .
- ^ «Впервые за десятилетия ангольские жирафы теперь населяют парк в Анголе» . ЭНЕРГЕТИЧЕСКИЙ ЯДЕРНЫЙ РЕАКТОР . 2023. Архивировано из оригинала 12 июля 2023 года . Проверено 12 июля 2023 г. , Меган Лим, NPR , 11 июля 2023 г.
- ^ Перейти обратно: а б Бок, Ф.; Феннесси, Дж.; Бидон, Т.; Татчингс, А.; Марэ, А.; Дьякон, Ф.; Янке, А. (2014). «Митохондриальные последовательности показывают четкое разделение между ангольскими и южноафриканскими жирафами вдоль загадочной рифтовой долины» . Эволюционная биология BMC . 14 (1): 219. Бибкод : 2014BMCEE..14..219B . дои : 10.1186/s12862-014-0219-7 . ISSN 1471-2148 . ПМК 4207324 . ПМИД 25927851 .
- ^ Перейти обратно: а б Зима, С.; Феннесси, Дж.; Феннесси, С.; Янке, А. (2018). «Матрилинейная структура популяции и распространение ангольских жирафов в пустыне Намиб и за ее пределами» . Экологическая генетика и геномика . 7–8 : 1–5. Бибкод : 2018EcoGG...7....1W . дои : 10.1016/j.egg.2018.03.003 . S2CID 90395544 .
- ^ Бреннеман, РА; Луи, Э.Э. младший; Феннесси, Дж. (2009). «Генетическая структура двух популяций намибийского жирафа Giraffa Camelopardalis angolensis ». Африканский журнал экологии . 47 (4): 720–728. Бибкод : 2009AfJEc..47..720B . дои : 10.1111/j.1365-2028.2009.01078.x .
- ^ Рукмакер, LC (1989). Зоологическое исследование Южной Африки 1650–1790 гг . ЦРК Пресс . ISBN 9789061918677 . Архивировано из оригинала 22 сентября 2023 года . Проверено 17 октября 2020 г.
- ^ Перейти обратно: а б Ли, Делавэр; Кавенер, ДР; Бонд, ML (2018). «Наблюдение пятен: количественная оценка сходства матери и потомства и оценка последствий приспособленности черт рисунка шерсти в дикой популяции жирафов ( Giraffa Camelopardalis )» . ПерДж . 6 : е5690. дои : 10.7717/peerj.5690 . ПМК 6173159 . ПМИД 30310743 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в Эстес, Р. (1992). Руководство по поведению африканских млекопитающих: включая копытных, хищников и приматов . Издательство Калифорнийского университета . стр. 202–207. ISBN 978-0-520-08085-0 .
- ^ ван Ситтерт, SJ; Митчелл, Г. (2015). «О реконструкции Giraffa sivalensis , вымершего жирафа с холмов Сивалик, Индия» . ПерДж . 3 : е1135. дои : 10.7717/peerj.1135 . ПМК 4540016 . ПМИД 26290791 .
- ^ " Жираф Бриссон 1762 (жираф)" . paleobiodb.org . Архивировано из оригинала 5 декабря 2021 года . Проверено 9 декабря 2021 г.
- ^ Перейти обратно: а б Новак, Р.М. (1999). Млекопитающие мира Уокера . Том. 1. Издательство Университета Джонса Хопкинса . стр. 1086–1089. ISBN 978-0801857898 . Архивировано из оригинала 22 сентября 2023 года . Проверено 30 августа 2021 г.
- ^ Перейти обратно: а б Скиннер, доктор медицинских наук; Смитерс, RHM (1990). Млекопитающие южноафриканского субрегиона . Университет Претории . стр. 616–20. ISBN 978-0-521-84418-5 .
- ^ Перейти обратно: а б с д и ж г час Сваби, С. (2010). "Жирафа". В Харрисе, Т. (ред.). Анатомия млекопитающих: Иллюстрированное руководство . Маршалл Кавендиш . стр. 64–84. ISBN 978-0-7614-7882-9 .
- ^ Лэнгли, Л. (2017). «У зебр есть полосы на коже?» . Нэшнл Географик . Архивировано из оригинала 1 апреля 2019 года . Проверено 2 июня 2020 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v Дагг, А.И. (2014). Жираф: биология, поведение и охрана . Издательство Кембриджского университета . ISBN 978-1107610170 .
- ^ Перейти обратно: а б с д и ж г Протеро, ДР; Шох, Р.М. (2003). Рога, бивни и ласты: эволюция копытных млекопитающих . Издательство Университета Джонса Хопкинса . стр. 67–72. ISBN 978-0-8018-7135-1 .
- ^ Фостер, Дж. Б. (1966). «Жираф национального парка Найроби: ареал обитания, соотношение полов, стадо и еда» . Африканский журнал экологии . 4 (1): 139–148. Бибкод : 1966AfJEc...4..139F . дои : 10.1111/j.1365-2028.1966.tb00889.x . Архивировано из оригинала 14 декабря 2022 года.
- ^ Ли, Делавэр; Лохай, Г.Г.; Кавенер, ДР; Бонд, ML (2022). «Использование точечного распознавания образов для изучения популяционной биологии, эволюционной экологии, социальности и движений жирафов: 70-летняя ретроспектива» . Биология млекопитающих . 102 (4): 1055–1071. дои : 10.1007/s42991-022-00261-3 . S2CID 252149865 . Архивировано из оригинала 22 сентября 2023 года . Проверено 14 декабря 2022 г.
- ^ Перейти обратно: а б с Митчелл, Г.; Скиннер, доктор медицинских наук (2004). «Терморегуляция жирафа: обзор» . Труды Королевского общества Южной Африки . 59 (2): 49–57. Бибкод : 2004TRSSA..59..109M . дои : 10.1080/00359190409519170 . S2CID 87321176 . Архивировано из оригинала 20 ноября 2018 года . Проверено 19 октября 2011 г.
- ^ Файн Марон, Дина (12 сентября 2023 г.). «Найден еще один редкий безупречный жираф — первый, когда-либо замеченный в дикой природе» . Нэшнл Географик . Проверено 6 марта 2024 г.
- ^ Ромо, Ванесса; Джонс, Дастин (6 сентября 2023 г.). «Редкий безупречный жираф получает подходящее имя» . ЭНЕРГЕТИЧЕСКИЙ ЯДЕРНЫЙ РЕАКТОР . Проверено 6 марта 2024 г.
- ^ Вуд, ВФ; Велдон, П.Дж. (2002). «Запах сетчатого жирафа ( Giraffa Camelopardalis reticulata )». Биохимическая систематика и экология . 30 (10): 913–17. Бибкод : 2002BioSE..30..913W . дои : 10.1016/S0305-1978(02)00037-6 .
- ^ Перейти обратно: а б с д Симмонс, Р.Э.; Альтвегг, Р. (2010). «Шеи для секса или конкурирующие браузеры? Критика идей эволюции жирафа». Журнал зоологии . 282 (1): 6–12. дои : 10.1111/j.1469-7998.2010.00711.x .
- ^ Митчелл, Г.; Робертс, генеральный директор; ван Ситтерт, SJ; Скиннер, доктор юридических наук (2013). «Ориентация орбиты и морфометрия глаз у жирафов ( Giraffa Camelopardalis )». Африканская зоология . 48 (2): 333–339. дои : 10.1080/15627020.2013.11407600 . hdl : 2263/37109 . S2CID 219292664 .
- ^ Петерсон, Д. (2013). Отражения жирафа . Издательство Калифорнийского университета . п. 30. ISBN 978-0520266858 .
- ^ Тейлор, член парламента; Ведель, MJ (2013). «Почему у зауроподов были длинные шеи и почему у жирафов шеи короткие» . ПерДж . 1 : е36. дои : 10.7717/peerj.36 . ПМЦ 3628838 . ПМИД 23638372 .
- ^ Перейти обратно: а б Ван Ситтерт, SJ; Скиннер, доктор медицинских наук; Митчелл, Г. (2010). «От плода к взрослому - аллометрический анализ позвоночного столба жирафа». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 314Б (6): 469–479. Бибкод : 2010JEZB..314..469V . дои : 10.1002/jez.b.21353 . ПМИД 20700891 .
- ^ Перейти обратно: а б Солуниас, Н. (1999). «Замечательная анатомия шеи жирафа» (PDF) . Журнал зоологии . 247 (2): 257–268. дои : 10.1111/j.1469-7998.1999.tb00989.x . Архивировано (PDF) из оригинала 25 марта 2009 г.
- ^ Перейти обратно: а б дю Туа, ЖТ (1990). «Стратификация по высоте кормления среди африканских жвачных животных» (PDF) . Африканский журнал экологии . 28 (1): 55–62. Бибкод : 1990AfJEc..28...55D . дои : 10.1111/j.1365-2028.1990.tb01136.x . Архивировано из оригинала (PDF) 10 ноября 2011 года . Проверено 21 ноября 2011 г.
- ^ Кэмерон, EZ ; дю Туа, ЖТ (2007). «Победа за шею: высокие жирафы избегают конкуренции с более короткими браузерами» . Американский натуралист . 169 (1): 130–135. дои : 10.1086/509940 . ПМИД 17206591 . S2CID 52838493 . Архивировано из оригинала 2 июня 2020 года . Проверено 5 декабря 2019 г.
- ^ Вулноу, AP; дю Туа, ЖТ (2001). «Вертикальная зональность качества побега в кронах деревьев, подвергающаяся воздействию гильдии африканских копытных, структурированных по размеру» (PDF) . Экология . 129 (1): 585–590. Бибкод : 2001Oecol.129..585W . дои : 10.1007/s004420100771 . ПМИД 24577699 . S2CID 18821024 . Архивировано из оригинала (PDF) 10 ноября 2011 года . Проверено 7 марта 2012 г.
- ^ Перейти обратно: а б Янг, Т.П.; Исбелл, Луизиана (1991). «Половые различия в экологии питания жирафов: энергетические и социальные ограничения» (PDF) . Этология . 87 (1–2): 79–89. Бибкод : 1991Этол..87...79Y . дои : 10.1111/j.1439-0310.1991.tb01190.x . Архивировано из оригинала (PDF) 16 мая 2013 года . Проверено 2 февраля 2012 г.
- ^ Митчелл, Г.; ван Ситтерт, С.; Скиннер, доктор медицинских наук (2010). «Демография смертности жирафов во время засухи». Труды Королевского общества Южной Африки . 65 (3): 165–168. Бибкод : 2010TRSSA..65..165M . дои : 10.1080/0035919X.2010.509153 . hdl : 2263/18957 . S2CID 83652889 .
- ^ Перейти обратно: а б Кавенер, ДР; Бонд, ML; Ву-Кавенер, Л; Лохай, Г.Г.; Кавенер, Миссури; Хоу, Х; Пирс, Д.Л.; Ли, Делавэр (2024). «Половой диморфизм пропорций тела жирафов Масаи и эволюция шеи жирафа» . Биология млекопитающих . дои : 10.1007/s42991-024-00424-4 .
- ^ Браунли, А. (1963). «Эволюция жирафа» . Природа . 200 (4910): 1022. Бибкод : 1963Natur.200.1022B . дои : 10.1038/2001022a0 . S2CID 4145785 .
- ^ Уильямс, EM (2016). «Рост жирафа и удлинение шеи: бдительность как эволюционный механизм» . Биология . 5 (3): 35. doi : 10.3390/biology5030035 . ПМК 5037354 . ПМИД 27626454 .
- ^ Вуд, К. (2014). «Заводные жирафы… особая структура костей удерживает этих животных в вертикальном положении» . Общество экспериментальной биологии . Архивировано из оригинала 25 ноября 2018 года . Проверено 7 мая 2014 г.
- ^ Гарланд, Т.; Дж., СМ (1993). «Предсказывает ли соотношение плюсны и бедра максимальной скорости бега у бегающих млекопитающих?» (PDF) . Журнал зоологии . 229 (1): 133–51. дои : 10.1111/j.1469-7998.1993.tb02626.x . Архивировано из оригинала (PDF) 20 ноября 2018 года . Проверено 25 апреля 2010 г.
- ^ Рафферти, JP (2011). Грейзерс (Британский справочник по хищникам и добыче) . Образовательное издательство Британника . п. 194. ИСБН 978-1-61530-336-6 .
- ^ Перейти обратно: а б с Хендерсон, DM; Нэйш, Д. (2010). «Прогнозирование плавучести, равновесия и потенциальной плавательной способности жирафов с помощью компьютерного анализа». Журнал теоретической биологии . 265 (2): 151–59. Бибкод : 2010JThBi.265..151H . дои : 10.1016/j.jtbi.2010.04.007 . ПМИД 20385144 .
- ^ Перейти обратно: а б Нэйш, Д. (2011). «Он будет плавать?» . Научный американец . 304 (1): 22. Бибкод : 2011SciAm.304a..22N . doi : 10.1038/scientificamerican0111-22 . Архивировано из оригинала 4 декабря 2013 года . Проверено 30 декабря 2010 г.
- ^ Тоблер, И.; Швиерин, Б. (1996). «Поведенческий сон жирафа ( Giraffa Camelopardalis ) в зоологическом саду» . Журнал исследований сна . 5 (1): 21–32. дои : 10.1046/j.1365-2869.1996.00010.x . ПМИД 8795798 . S2CID 34605791 .
- ^ Перейти обратно: а б Ведель, MJ (2012). «Памятник неэффективности: предполагаемый ход возвратного гортанного нерва у динозавров-зауроподов» (PDF) . Acta Palaeontologica Polonica . 57 (2): 251–256. дои : 10.4202/app.2011.0019 . S2CID 43447891 . Архивировано (PDF) из оригинала 29 октября 2013 г.
- ^ Харрисон, DFN (1995). Анатомия и физиология гортани млекопитающих . Издательство Кембриджского университета. п. 165. ИСБН 978-0-521-45321-9 .
- ^ Граик, Ж.-М.; Перуффо, А.; Балларин, К.; Коцци, Б. (2017). «Мозг жирафа ( Giraffa Camelopardalis ): конфигурация поверхности, коэффициент энцефализации и анализ существующей литературы» . Анатомическая запись . 300 (8): 1502–1511. дои : 10.1002/ar.23593 . ПМИД 28346748 . S2CID 3634656 .
- ^ Скиннер, доктор медицинских наук; Митчелл, Г. (2011). «Объемы легких жирафов, Giraffa Camelopardalis » (PDF) . Сравнительная биохимия и физиология А. 158 (1): 72–78. дои : 10.1016/j.cbpa.2010.09.003 . hdl : 2263/16472 . ПМИД 20837156 . Архивировано (PDF) из оригинала 20 ноября 2018 года . Проверено 27 ноября 2011 г.
- ^ Митчелл, Г.; Скиннер, доктор юридических наук (1993). «Как жирафы приспосабливаются к своей необычной форме». Труды Королевского общества Южной Африки . 48 (2): 207–218. Бибкод : 1993TRSSA..48..207M . дои : 10.1080/00359199309520271 .
- ^ Митчелл, Г.; ван Ситтерт, SJ; Скиннер, доктор юридических наук (2009). «Структура и функции клапанов яремной вены жирафа» (PDF) . Южноафриканский журнал исследований дикой природы . 39 (2): 175–180. дои : 10.3957/056.039.0210 . hdl : 2263/13994 . S2CID 55201969 . Архивировано (PDF) из оригинала 20 ноября 2018 года . Проверено 21 ноября 2011 г.
- ^ Перес, В.; Лима, М.; Клаусс, М. (2009). «Общая анатомия кишечника жирафа ( Giraffa Camelopardalis )» (PDF) . Анатомия, Гистология, Эмбриология . 38 (6): 432–435. дои : 10.1111/j.1439-0264.2009.00965.x . ПМИД 19681830 . S2CID 28390695 . Архивировано (PDF) из оригинала 22 июля 2018 года.
- ^ Кейв, AJE (1950). «О печени и желчном пузыре жирафа». Труды Лондонского зоологического общества . 120 (2): 381–93. дои : 10.1111/j.1096-3642.1950.tb00956.x .
- ^ Олдхэм-Отт, СК; Жиллото, Дж. (1997). «Сравнительная морфология желчного пузыря и желчевыводящих путей у позвоночных: различия в строении, гомология функций и желчные камни». Микроскопические исследования и техника . 38 (6): 571–579. doi : 10.1002/(SICI)1097-0029(19970915)38:6<571::AID-JEMT3>3.0.CO;2-I . ПМИД 9330347 . S2CID 20040338 .
- ^ Феннесси, Дж. (2004). Экология пустынного жирафа Giraffa Camelopardalis angolensis на северо-западе Намибии ( докторская диссертация). Университет Сиднея . Архивировано из оригинала 20 ноября 2018 года . Проверено 30 января 2012 г.
- ^ Перейти обратно: а б с Кингдон, Дж. (1997). «Жирафы-жирафиды» . Полевой путеводитель Кингдона по африканским млекопитающим . Академическая пресса . стр. 339–344. ISBN 978-0-12-408355-4 .
- ^ Перейти обратно: а б Бонд, ML; Ли, Дерек Э.; Озгуль А.; Кениг, Б. (2019). «Динамика деления-слияния мегатравоядных определяется экологическими, антропогенными, временными и социальными факторами» . Экология . 191 (2): 335–347. Бибкод : 2019Oecol.191..335B . дои : 10.1007/s00442-019-04485-y . ПМИД 31451928 . S2CID 201732871 . Архивировано из оригинала 22 сентября 2023 года . Проверено 22 августа 2023 г.
- ^ ван дер Югд, HP; Принс, HHT (2000). «Движения и групповая структура жирафа ( Giraffa Camelopardalis ) в национальном парке озера Маньяра, Танзания» (PDF) . Журнал зоологии . 251 (1): 15–21. дои : 10.1111/j.1469-7998.2000.tb00588.x . Архивировано из оригинала (PDF) 6 ноября 2013 года.
- ^ Перейти обратно: а б с д и ж Пратт, DM; Андерсон, В.Х. (1985). «Социальное поведение жирафа». Журнал естественной истории . 19 (4): 771–781. Бибкод : 1985JNatH..19..771P . дои : 10.1080/00222938500770471 .
- ^ Перейти обратно: а б Бонд, ML; Кениг, Б.; Ли, Делавэр; Озгуль А.; Фарин, ДР (2020). «Близость к людям влияет на местную социальную структуру метапопуляции жирафов» . Журнал экологии животных . 90 (1): 212–221. дои : 10.1111/1365-2656.13247 . ПМИД 32515083 .
- ^ Беркович, ФБ; Берри, ПСМ (2013). «Состав стада, родство и социальная динамика деления-слияния среди диких жирафов» . Африканский журнал экологии . 51 (2): 206–216. Бибкод : 2013AfJEc..51..206B . дои : 10.1111/aje.12024 .
- ^ Перейти обратно: а б Картер, К.Д.; Седдон, Дж. М.; Фререб, Швейцария; Картер, Дж. К. (2013). «Динамика деления-слияния у диких жирафов может быть обусловлена родством, пространственным перекрытием и индивидуальными социальными предпочтениями». Поведение животных . 85 (2): 385–394. дои : 10.1016/j.anbehav.2012.11.011 . S2CID 53176817 .
- ^ Перейти обратно: а б с д и ВандерВаал, КЛ; Ван, Х.; МакКоуэн, Б.; Фушинг, Х.; Исбелл, Луизиана (2014). «Многоуровневая социальная организация и использование пространства у сетчатого жирафа ( Giraffa Camelopardalis )». Поведенческая экология . 25 (1): 17–26. doi : 10.1093/beheco/art061 .
- ^ Бонд, ML; Кениг, Б.; Озгуль А.; Фарин, ДР; Ли, Делавэр (2021). «Социально определенные субпопуляции демонстрируют демографические различия в метапопуляции жирафов» . Журнал управления дикой природой . 85 (5): 920–931. Бибкод : 2021JWMan..85..920B . дои : 10.1002/jwmg.22044 . S2CID 233600744 . Архивировано из оригинала 8 марта 2022 года . Проверено 27 января 2024 г.
- ^ Бонд, ML; Ли, Делавэр; Озгуль А.; Фарин, ДР; Кениг, Б. (2021). «Уход, оставаясь: социальное рассредоточение жирафов» . Журнал экологии животных . 90 (12): 2755–2766. Бибкод : 2021JAnEc..90.2755B . дои : 10.1111/1365-2656.13582 . ПМК 9291750 .
- ^ Лависта Феррес, JM; Ли, Делавэр; Насир, Мэриленд; Чен, Ю.-К.; Биджрал, А.С.; Беркович, ФБ; Бонд, ML (2021). «Социальные связи и перемещения среди сообществ жирафов различаются в зависимости от пола и возраста» . Поведение животных . 180 : 315–328. дои : 10.1016/j.anbehav.2021.08.008 . S2CID 237949827 .
- ^ Перейти обратно: а б с д Лейтольд, Б.М. (1979). «Социальная организация и поведение жирафа в Восточном национальном парке Цаво». Африканский журнал экологии . 17 (1): 19–34. Бибкод : 1979AfJEc..17...19L . дои : 10.1111/j.1365-2028.1979.tb00453.x .
- ^ Кнюзель, М.; Ли, Д.; Кениг, Б.; Бонд, М. (2019). «Корреляты размеров домашнего ареала жирафов, Giraffa Camelopardalis » (PDF) . Поведение животных . 149 : 143–151. дои : 10.1016/j.anbehav.2019.01.017 . S2CID 72332291 . Архивировано (PDF) из оригинала 10 февраля 2020 г.
- ^ Перейти обратно: а б Касози, Х.; Монтгомери, РА (2018). «Как жирафы находят друг друга? Обзор визуального, слухового и обонятельного общения жирафов» . Журнал зоологии . 306 (3): 139–146. дои : 10.1111/jzo.12604 .
- ^ Володина Елена Владимировна; Володин Илья А.; Челышева Елена Владимировна; Фрей, Роланд (2018). «Шипение и фырканье называют типы диких жирафов Giraffa Camelopardalis : акустическая структура и контекст» . Исследовательские заметки BMC . 11 (12): 12. дои : 10.1186/s13104-017-3103-x . ПМК 5761111 . ПМИД 29316966 .
- ^ Баотич, А.; Сикс, Ф.; Штегер, А.С. (2015). «Ночные «гудящие» вокализации: добавление кусочка к загадке голосового общения жирафов» . Исследовательские заметки BMC . 8 : 425. doi : 10.1186/s13104-015-1394-3 . ПМК 4565008 . ПМИД 26353836 .
- ^ Фон Муггенталер, Э. (2013). «Жирафовый резонанс Гельмгольца» . Материалы совещаний по акустике . 19 (1): 010012. дои : 10.1121/1.4800658 .
- ^ Перейти обратно: а б дель Кастильо, СМ; Башоу, MJ; Паттон, ML; Ричес, Р.Р.; Беркович, ФБ (2005). «Анализ фекальных стероидов репродуктивного состояния самок жирафов ( Giraffa Camelopardalis ) и влияние эндокринного статуса на ежедневные бюджеты времени» . Общая и сравнительная эндокринология . 141 (3): 271–281. дои : 10.1016/j.ygcen.2005.01.011 . ПМИД 15804514 . Архивировано из оригинала 25 мая 2021 года . Проверено 10 февраля 2021 г.
- ^ Беркович, ФБ; Башоу, MJ; дель Кастильо, С.М. (2006). «Социосексуальное поведение, тактика спаривания самцов и репродуктивный цикл жирафа Giraffa Camelopardalis » . Гормоны и поведение . 50 (2): 314–321. дои : 10.1016/j.yhbeh.2006.04.004 . ПМИД 16765955 . S2CID 45843281 .
- ^ Сибер, Пенсильвания; Дункан, П.; Фриц, Х.; Гансвиндт, А. (2013). «Андрогенные изменения и гибкое поведение самцов жирафов во время гона» . Письма по биологии . 9 (5): 20130396. doi : 10.1098/rsbl.2013.0396 . ПМЦ 3971675 . ПМИД 23925833 .
- ^ Перейти обратно: а б с Лангман, В.А. (1977). «Отношения коровы и теленка у жирафа ( Giraffa Camelopardalis Giraffa )» . Журнал психологии животных . 43 (3): 264–286. дои : 10.1111/j.1439-0310.1977.tb00074.x . Архивировано из оригинала 12 января 2020 года . Проверено 12 января 2020 г.
- ^ Сайто, М.; Идани, Г. (2018). «Сосущее и сосущее поведение диких жирафов ( Giraffa Camelopardalis Tippelskirchi )». Биология млекопитающих . 93 : 1–4. Бибкод : 2018МамБи..93....1S . дои : 10.1016/j.mambio.2018.07.005 . S2CID 91472891 .
- ^ Глонекова, М.; Брандлова, К.; Плугачек, Ю. (2021). «Дальнейшие поведенческие параметры подтверждают взаимность и кражу молока как объяснение заботы жирафов» . Научные отчеты . 11 (1): 7024. Бибкод : 2021NatSR..11.7024G . дои : 10.1038/s41598-021-86499-2 . ПМК 8007720 . ПМИД 33782483 .
- ^ Холл-Мартин, AJ; Скиннер, доктор медицинских наук; Хопкинс, Би Джей (1978). «Развитие репродуктивных органов самца жирафа Giraffa Camelopardalis » (PDF) . Журнал репродукции и фертильности . 52 (1): 1–7. дои : 10.1530/jrf.0.0520001 . ПМИД 621681 . Архивировано (PDF) из оригинала 19 июля 2018 года . Проверено 12 июня 2017 г.
- ^ Коу, MJ (1967). « Поведение жирафа, тянущего шею». Журнал зоологии . 151 (2): 313–21. дои : 10.1111/j.1469-7998.1967.tb02117.x .
- ^ Багемил, Б. (1999). Биологическое изобилие: гомосексуализм животных и естественное разнообразие . Пресса Святого Мартина. стр. 391–393. ISBN 978-0-312-19239-6 .
- ^ Перейти обратно: а б Ли, Делавэр; Штраус, МКЛ (2016). Справочный модуль по системам Земли и наукам об окружающей среде . Эльзевир. дои : 10.1016/B978-0-12-409548-9.09721-9 . ISBN 9780124095489 .
- ^ Мюллер, Д.В.; Зербе, П.; Кодрон, Д.; Клаусс, М.; Хатт, Дж. М. (2011). «Долгая жизнь среди жвачных животных: жирафы и другие особые случаи». Швейцарский архив ветеринарной медицины . 153 (11): 515–519. дои : 10.1024/0036-7281/a000263 . ПМИД 22045457 . S2CID 10687135 .
- ^ Бонд, ML; Ли, Делавэр; Фарин, ДР; Озгуль А.; Кениг, Б. (2021). «Общительность увеличивает выживаемость взрослых самок жирафов» . Труды Королевского общества B: Биологические науки . 288 (1944): 20202770. doi : 10.1098/rspb.2020.2770 . ПМЦ 7893237 . ПМИД 33563118 .
- ^ Ли, Делавэр; Бонд, ML; Киссуи, Б.М.; Киванго, Ю.А.; Болджер, Т. (2016). «Пространственные вариации демографии жирафов: проверка двух парадигм» . Журнал маммологии . 97 (4): 1015–1025. дои : 10.1093/jmammal/gyw086 . S2CID 87117946 .
- ^ Ли, Делавэр; Бонд, ML; Болджер, Д.Т. (2017). «Сезон рождения влияет на выживаемость молоди жирафов» . Популяционная экология . 59 (1): 45–54. Бибкод : 2017PopEc..59...45L . дои : 10.1007/s10144-017-0571-8 . S2CID 7611046 . Архивировано из оригинала 22 сентября 2023 года . Проверено 22 августа 2023 г.
- ^ Ли, Делавэр; Киссуи, Б.М.; Киванго, Ю.А.; Бонд, ML (2016). «Мигрирующие стада антилоп гну и зебр косвенно влияют на выживаемость детенышей жирафов» . Экология и эволюция . 6 (23): 8402–8411. Бибкод : 2016EcoEv...6.8402L . дои : 10.1002/ece3.2561 . ПМК 5167056 . ПМИД 28031792 .
- ^ Шмитт, Миннесота; Стирс, К.; Шрейдер, AM (2016). «Зебры снижают риск нападения хищников в смешанных стадах, подслушивая сигналы жирафов» . Поведенческая экология . 27 (4): 1073–1077. дои : 10.1093/beheco/arw015 .
- ^ Ли, Делавэр; Бонд, ML (2016). «Возникновение и распространенность кожных заболеваний жирафов на охраняемых территориях северной Танзании». Журнал болезней дикой природы . 52 (3): 753–755. дои : 10.7589/2015-09-247 . ПМИД 27310168 . S2CID 10736316 .
- ^ Бонд, ML; Штраус, МКЛ; Ли, Делавэр (2016). «Почвенные корреляции и смертность от кожных заболеваний жирафов в Танзании». Журнал болезней дикой природы . 52 (4): 953–958. дои : 10.7589/2016-02-047 . ПМИД 27529292 . S2CID 46776142 .
- ^ Муньеза, AB; Монтгомери, РА; Феннесси, Джей Ти; Дикман, Эй Джей; Ролофф, Дж.Дж.; Макдональд, Д.В. (2016). «Региональные вариации проявления, распространенности и тяжести кожных заболеваний жирафов: обзор новых заболеваний в диких и содержащихся в неволе популяциях жирафов». Биологическая консервация . 198 : 145–156. Бибкод : 2016BCons.198..145M . дои : 10.1016/j.biocon.2016.04.014 .
- ^ Перейти обратно: а б с д и ж г час я дж к л Уильямс, Э. (2011). Жираф . Книги реакции . ISBN 978-1-86189-764-0 .
- ^ «Петрограф наскального рисунка «Жираф Дабус» . Фонд Брэдшоу . Архивировано из оригинала 7 ноября 2011 года . Проверено 6 ноября 2011 г.
- ^ Шоррокс, Б. (2016). Жираф: биология, экология, эволюция и поведение . Уайли . п. 3. ISBN 9781118587478 .
- ^ Уолтер, М.; Фурнье, А.; Менево, Д. (2001). «Интеграция формы и рисунка в моделях млекопитающих» (PDF) . Материалы 28-й ежегодной конференции «Компьютерная графика и интерактивные технологии» . стр. 317–26 . CiteSeerX 10.1.1.10.7622 . дои : 10.1145/383259.383294 . ISBN 978-1-58113-374-5 . S2CID 13488215 . Архивировано из оригинала 23 сентября 2015 года . Проверено 16 ноября 2011 г.
- ^ Клегг, А. (1986). «Некоторые аспекты космологии тсваны». Примечания и записи Ботсваны . 18 : 33–37. JSTOR 40979758 .
- ^ Рингмар, Э. (2006). «Аудитория жирафа: европейский экспансионизм и поиск экзотики» (PDF) . Журнал всемирной истории . 17 (4): 353–97. дои : 10.1353/jwh.2006.0060 . JSTOR 20079397 . S2CID 143808549 . Архивировано (PDF) из оригинала 10 сентября 2008 г.
- ^ Радгли, Ричард Энциклопедия психоактивных веществ , паб. Абакус 1998 г. ISBN 0 349 11127 8 стр. 20–21.
- ^ Ян Каннисон (1958). «Охота на жирафов среди племени Хумр». Суданские заметки и записи . 39 .
- ^ «Жираф – Факты: Текущий статус жирафа?» . Фонд охраны жирафов. Архивировано из оригинала 19 марта 2016 года . Проверено 21 декабря 2010 г.
- ^ Мэтт МакГрат (8 декабря 2016 г.). «Жирафам грозит «тихое вымирание» по мере сокращения их численности» . Новости Би-би-си . Архивировано из оригинала 21 мая 2019 года . Проверено 8 декабря 2016 г.
- ^ «Новые виды птиц и жирафы под угрозой – Красный список МСОП» . 8 декабря 2016 года. Архивировано из оригинала 5 марта 2017 года . Проверено 8 декабря 2016 г.
- ^ Болджер, Д.; Огуту, Дж.; Штраус, М.; Ли, Д.; Муньеза, А.; Феннесси, Дж.; Браун, Д. (2019). «Масайский жираф» . Красный список исчезающих видов МСОП . 2019 . doi : 10.2305/IUCN.UK.2019-1.RLTS.T88421036A88421121.en .
- ^ Муньеза, А.; Доэрти, Дж.Б.; Хусейн Али, А.; Феннесси, Дж.; Марэ, А.; О'Коннор, Д.; Вубе, Т. (2018). «Сетчатый жираф» . Красный список исчезающих видов МСОП . 2018 . doi : 10.2305/IUCN.UK.2018-2.RLTS.T88420717A88420720.en .
- ^ Перейти обратно: а б Вубе, Т.; Доэрти, Дж.Б.; Феннесси, Дж.; Марэ, А. (2018). « Жираф камелопардалис подвид камелопардалис » . Красный список исчезающих видов МСОП . 2018 . doi : 10.2305/IUCN.UK.2018-2.RLTS.T88420707A88420710.en .
- ^ Цю, Джейн (2017). «Всплеск убийств диких животных приводит к исчезновению жирафов» . Наука . дои : 10.1126/science.aan7000 . Архивировано из оригинала 9 февраля 2023 года . Проверено 30 июня 2022 г.
- ^ Ли, Дерек Э. (2018). «Оценка эффективности охраны природы в зоне управления дикой природой Танзании» . Журнал управления дикой природой . 82 (8): 1767–1774. Бибкод : 2018JWMan..82.1767L . дои : 10.1002/jwmg.21549 . ISSN 1937-2817 . S2CID 91251633 . Архивировано из оригинала 26 октября 2020 года . Проверено 16 ноября 2020 г.
- ^ Ли, Дерек Э; Бонд, Моника Л. (2018). «Количественная оценка экологического успеха общественного заповедника дикой природы в Танзании» . Журнал маммологии . 99 (2): 459–464. дои : 10.1093/jmammal/gyy014 . ISSN 0022-2372 . ПМК 5965405 . ПМИД 29867255 .
- ^ Баттонс, Дж (1987). Восточная Африка: Кения, Танзания и Уганда. Издательство Викас. п. 57. ИСБН 978-0-7069-2822-8 .
- ^ Чарльз Фоули; Лара Фоули; Алекс Лобора; Даниэла Де Лука; Маурус Мсуха; Тим Р.Б. Давенпорт; Сара М. Дюрант (2014). Полевой справочник по крупным млекопитающим Танзании . Издательство Принстонского университета . п. 179. ИСБН 978-1-4008-5280-2 . Архивировано из оригинала 22 сентября 2023 года . Проверено 13 июля 2016 г.
- ^ «Национальные символы: Национальное животное» . tanzania.go.tz . Правительственный портал Танзании. Архивировано из оригинала 18 января 2015 года . Проверено 14 января 2015 г.
- ^ «Шимпанзе среди 33 пород, отобранных под особую охрану» . Новости Би-би-си . 28 октября 2017 года. Архивировано из оригинала 29 октября 2017 года . Проверено 30 октября 2017 г.
- ^ «Хорошие новости для жирафов на 18-й конференции CITES > Отдел новостей» . newsroom.wcs.org . Архивировано из оригинала 27 сентября 2020 года . Проверено 16 ноября 2020 г.
- ^ Мюллер, Зоя; Ли, Дерек Э.; Шайен, Циска П.Дж.; Штраус, Меган К.Л.; Картер, Керрин Д.; Дьякон, Франсуа (2020). «Транслокации жирафов: обзор и обсуждение соображений» . Африканский журнал экологии . 58 (2): 159–171. Бибкод : 2020AfJEc..58..159M . дои : 10.1111/aje.12727 .
- ^ Ли, Де; Финиег, Э.; Ван Остерхаут, К.; Мюллер, З.; Штраус, М.; Картер, Кд.; Шайен, Cpj; Дикон, Ф. (27 февраля 2020 г.). «Анализ жизнеспособности транслокационной популяции жирафов» . Исследования исчезающих видов . 41 : 245–252. дои : 10.3354/esr01022 . Архивировано из оригинала 24 сентября 2020 года . Проверено 16 ноября 2020 г.
- ^ Ли, Дерек Э.; Бонд, Моника Л. (2016). «Точность, достоверность и стоимость методов исследования жирафов Giraffa Camelopardalis » . Журнал маммологии . 97 (3): 940–948. дои : 10.1093/jmammal/gyw025 . S2CID 87384776 .
Внешние ссылки
- Официальный сайт Фонда охраны жирафов.