Тилоцефале
| Тилоцефале Временной диапазон: поздний кампан ,
| |
|---|---|

| |
| Схема голотипа черепа | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Неорнитишия |
| Клэйд : | † Пахицефалозаврия |
| Семья: | † Пахицефалозавриды |
| Род: | † Тилоцефале Марьянская и Осмольская, 1974 г. |
| Разновидность: | † Т. Гилморей
|
| Биномиальное имя | |
| † Tylocephale gilmorei Марьянская и Осмольская, 1974 г.
| |
Tylocephale (что означает «опухшая голова») — род пахицефалозавридных динозавров позднего , группы куполообразных травоядных птицетазовых животных , которые жили во время позднекампанского этапа (75–73 миллиона лет назад) мела на территории современной Монголии . Он известен по частичному черепу и связанной с ним нижней челюсти , которые были обнаружены в 1971 году польско-монгольской экспедицией в формацию Барун-Гойот в пустыне Гоби . Образец был описан в 1974 году польскими палеонтологами Терезой Марьянской и Гальшкой Осмолской как новый род и вид.
Он был среднего размера для пахицефалозавра: достигал 2 м (6,6 футов) в длину и 40 кг (88 фунтов) массы тела. Череп имеет треугольную форму при виде сзади, самая широкая точка находится в области скуловых костей, с вершиной наверху купола. Купол Тилоцефала — самый высокий из известных среди пахицефалозавров. Этот купол также необычайно толстый и морщинистый снаружи. За куполом сзади над шеей выступает множество шипов, узлов и бугорков. Надбровная дуга была украшена мелкими костными узелками и была толще, чем у других родов. Tylocephale тесно связан с другими азиатскими пахицефалозаврами, такими как Homalocephale и Goyocephale , будучи частью ветви Старого Света группы.
Будучи пахицефалозавром, он был травоядным и имел маленькие ребристые зубы, приспособленные для расщепления волокнистых растений. Его зубные ряды также имеют зазубрины , что предполагает потенциально более гетерогенную диету из листьев, орехов, семян и насекомых. Окаменелости были найдены в местности Хулсан, где сохранились окаменелости многих других групп динозавров, таких как цератопсы , анкилозавры , овирапторозавры , дромеозавры и титанозавры . Формация Барун-Гойот представляла собой аллювиальную равнину с крупными реками в засушливом или полузасушливом климате.
Открытие и присвоение имени
[ редактировать ]
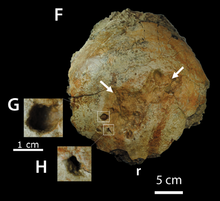
Во время совместной польско-монгольской экспедиции к обнажению Хулсан формации Барун-Гойот в пустыне Гоби в 1971 году были обнаружены большой череп и нижняя челюсть пахицефалозавра ( каталожный номер ZPAL MgD-I/105 ). [ 1 ] [ 2 ] Слои горных пород формации Барун-Гойот происходят из позднекампанского этапа позднего мела и имеют возраст от 75 до 73 миллионов лет. [ 1 ] Это была одна из серии экспедиций, проведенных в период с 1963 по 1971 год под руководством польского палеонтолога Зофии Килан-Яворовской , в ходе которых было собрано множество хорошо сохранившихся скелетов динозавров . [ 3 ] К предприятию присоединились еще несколько польских ученых, в том числе Тереза Марьянска и Хальшка Осмолска , которым помогали местные монгольские помощники. Череп был неполным, в нем отсутствовала большая часть передних частей и остальная часть костного купола. Нижняя челюсть и череп сохранились в сочлененном виде и лежали в выветренных блоках песчаника из бывшего русла реки . материалы анкилозавров Tarchia и Saichania , цератопсид Breviceratops и теропод Hulsanpes . Этот экземпляр был одним из нескольких особей динозавров, обнаруженных в Кхулсане в 1970-х годах. В этом месте были обнаружены [ 4 ] [ 5 ] Все окаменелости, обнаруженные во время этой экспедиции, были затем перевезены в Польскую академию наук в Варшаве , где они были описаны в последующие годы. [ 2 ]
Череп пахицефалозавра, найденный в Хулсане, был одним из нескольких, собранных во время польско-монгольских экспедиций, а и другие экземпляры из близлежащей формации Немегт в дополнение к Баруну Гойоту были собраны . Материал пахицефалозавра из обеих формаций был описан в научной литературе в журнале Palaeontologica Polonica в 1974 году Терезой Марьянской и Гальской Осмольской. Череп, найденный в Хулсане, был обозначен как типовой экземпляр нового рода и вида Tylocephale gilmorei . Название рода Tylocephale происходит от греческих слов tyle («опухший») и cephale («голова») и относится к выступающему куполу черепа. Видовое название дано в честь американского палеонтолога Чарльза Гилмора , написавшего первое подробное описание пахицефалозавра. В статье 1974 года также были названы два новых пахицефалозавра на основе окаменелостей из Nemegt: Homalocephale и Prenocephale . Все эти таксоны были сгруппированы в новый порядок Maryańska и Osmólska под названием Pachycephalosauria, который включал североамериканские роды, такие как стегоцерас и пахицефалозавр . А также [ 6 ]
Описание
[ редактировать ]
Тилоцефале был пахицефалозавром среднего размера, его длина составляла 2 м (6,6 фута), а масса тела - 40 кг (88 фунтов). [ 7 ] Никаких посткраниальных окаменелостей, принадлежащих Tylocephale, обнаружено не было, хотя имеются хорошо сохранившиеся скелеты родственных Stegoceras, Homalocephale и Prenocephale . Судя по этим таксонам, у Tylocephale была короткая шея , крошечные передние конечности , длинные задние конечности и толстый стержнеобразный хвост для равновесия. Шея была тонкой, U-образной формы, имела изогнутую позу и прикреплялась к затылочному мыщелку на задней части черепа. В его позвоночнике были прочные связи между позвонками, укрепленные окостеневшими сухожилиями . Его руки были легкими и тонкими, заканчивающимися кистью с пятью пальцами. Конечности заканчивались стопой с тремя пальцами, средний из которых был самым длинным, и все они имели когти . [ 8 ] [ 7 ]
Череп и украшения
[ редактировать ]Единственный известный экземпляр представляет собой частичный череп без черепной коробки , неба и передних частей черепа. Сохранилась и ее задняя половина нижней челюсти. есть три фенестры У большинства динозавров в черепе (полые пространства), но у Tylocephale только два. Подвисочное окно расположено под вертикальным углом и имеет одинаковую ширину на большей части своей длины. Череп очень высокий и узкий сзади, с короткой заглазничной частью. Примечательно, что крыша черепа высокая, с вершиной, очень близкой к заднему краю черепа. Эта крыша также толще и имеет вершину дальше назад, чем у других членов группы, что является отличительной особенностью таксона. Дорсальная часть чешуйчатой кости является наиболее плотным элементом черепа, при этом заострена, а не сглажена и не вздута. Однако вентральная поверхность чешуйчатой кости тоньше и контактирует с экзозатылочной . [ 6 ]

Все затылочные кости тонкие по сравнению с остальной частью черепа. Квадраты . удлинены и почти идеально совпадают с нижними челюстями Это позволило обеспечить прочное сочленение черепа и нижней челюсти. Ее квадратная кость расположена вертикально и перпендикулярно краю верхней челюсти. Напротив, скуловая кость (щечная кость) крепко построена и ориентирована латерально. Скуловые кости — самая широкая точка черепа, имеют треугольную форму в поперечном сечении. Боковая стенка спланхнокраниума квадратноскуловая (задняя часть черепа), скуловая кость и кость (скула) образуют поперечно широкую структуру. Орбита . (глазница) очень широкая, с надбровным валиком над отверстием, как и у других пахицефалозавров Верхний край ее уплощенный, с узкой заглазничной перемычкой, параллельной квадратной кости. Обе надглазничные кости сохранились, но неполные. Они высокие и толстые в поперечном сечении и составляют большую часть купола. [ 6 ]
Черепная орнаментация характерна для пахицефалозавров, которые часто имеют три структуры; узлы, клубни и колосья. Эти украшения становились наиболее крупными в чешуйчатой кости и меньшими ближе к передней части черепа. Надглазничные и заглазничные кости имеют некоторый орнамент, но он не является исключительно грубым. Tylocephale Югалы имеют гигантские, выступающие и неравномерно расположенные клубни. Купол, в отличие от некоторых других пахицефалозавров, имел грубую текстуру. Чешуйчатые кости заднего края черепа имели ряд гигантских шипов и бугорков. Из них самый большой располагался под самым крайним узлом ряда. Эти шипастые узлы продолжаются по всей длине чешуйчатой и заглазничной кости. [ 6 ]
Зубы и нижняя челюсть
[ редактировать ]Зубной ряд неполный, но сохранились девять зубов задней части челюстей. Все зубы, кроме последнего, расположены в прямую линию, что является уникальной чертой этого рода. Зубы были повреждены из-за внешних факторов, таких как эрозия и тафономия . Марьянска и Осмольска отметили, что пропорционально зубной ряд Tylocephale намного крупнее, чем у других пахицефалозавров, таких как Homalocephale. Зубы, числом семь, имеют высокие коронки и дугообразные режущие поверхности. Как и зубы, нижняя челюсть сохранилась очень плохо и состоит только из задних частей. У него слабо приподнятый отросток ветви венечный , который сочленяется со скуловой костью. Однако приводящая ямка очень глубока и хорошо развита в поперечном разрезе. [ 6 ] Эта ямка, расположенная между зубной и суставной поверхностью , служила для соединения мышц , нервов и вен со скуловой костью. [ 9 ]
Классификация
[ редактировать ]Tylocephale был членом группы Pachycephalosauria, семейства толсточерепных травоядных двуногих динозавров, которые жили в меловой период в Азии и Северной Америке. [ 6 ] Последние пахицефалозавры вымерли во время мел-палеогенового вымирания , причем последним выжившим родом был сам Pachycephalosaurus . [ 10 ] [ 11 ] Tylocephale Однако кладистический анализ 2020 года выявил гетеродонтозаврид как раннюю ветвь группы, что увеличивает возраст пахицефалозавров еще до ранней юры . [ 12 ] В настоящее время пахицефалозавры признаны частью более крупной группы Marginocephalia , которая включает в себя его и гигантских рогатых цератопсов. [ 13 ] [ 14 ]
В пределах Pachycephalosauria филогенетическое положение Tylocephale и других родов меняется из-за отсутствия многих хорошо сохранившихся экземпляров. [ 15 ] Несмотря на это, пахицефалозавры Азии, такие как Tylocephale, Homalocephale и Goyocephale, обнаруживаются в одном классе, тогда как североамериканские пахицефалозавры, Stygimoloch и часто Alaskacephale находятся в отдельной группе. [ 16 ] [ 17 ] Азиатские члены также являются более базальными и имеют меньше продвинутых характеристик по сравнению со своими североамериканскими коллегами. [ 18 ] Это связано с тем, что пахицефалозавры возникли в Азии, а затем распространились в Северную Америку во время ее краткого воссоединения с Азией в позднем меловом периоде. [ 19 ] [ 20 ] Tylocephale наиболее тесно связан с куполообразными Foraminacephale и плоскоголовыми Homalocephale . Согласно последним филогенетическим анализам, [ 21 ] Было высказано предположение, что Tylocephale gilmorei является не отдельным видом, а синонимом Prenocephale prenes . [ 22 ] Аналогичное предположение было сделано и в отношении Homalocephale . [ 23 ] Более поздние исследования гистологии более молодых экземпляров Prenocephale также доказывают его отличие от Homalocephale и Tylocephale . [ 23 ]
Ниже слева показано положение Tylocephale внутри Pachycephalosauridae согласно публикации Schott & Evans 2016 года о классификации Foraminacephale , которая считает его более базальным по отношению к более крупной североамериканской кладе. [ 24 ] Внизу справа показано филогенетическое расположение Pachycephalosauria в целом по данным Dieudonné et al (2020): [ 12 ]
Палеобиология
[ редактировать ]Диета
[ редактировать ]Неизвестно, чем питались пахицефалозавры; Имея очень маленькие ребристые зубы, они не могли жевать жесткие волокнистые растения так же эффективно, как другие динозавры того же периода. Предполагается, что их острые, зазубренные зубы идеально подходили для смешанной диеты, состоящей из листьев, семян, фруктов и насекомых. [ 25 ] Tylocephale , возможно, питался полностью травоядными животными, поскольку их зубные коронки были похожи на коронки зубов игуанид . На предчелюстных зубах имеются фасетки износа из-за контакта с предзубной костью, а на верхнечелюстных зубах имеются двойные фасетки износа, подобные тем, которые наблюдаются у других птицетазовых динозавров. [ 6 ] Каждый третий верхнечелюстной зуб у UALVP 2 является прорезывающимся замещающим зубом , а замена зубов происходила в обратном порядке в последовательных трех зубах. Затылочная область Stegoceras была хорошо разграничена для прикрепления мышц, и считается, что движение челюсти Stegoceras и других пахицефалозавров в основном ограничивалось движениями вверх и вниз с лишь небольшой способностью к вращению челюсти. Это основано на строении челюсти, микроизносе зубов и фасетках износа зубов, что указывает на то, что сила укуса использовалась больше для сдвига, чем для дробления. [ 26 ] [ 27 ] Однако было высказано предположение, что Tylocephale отличается от Stegoceras тем, что челюсть движется вперед и назад, а не вверх и вниз. Это пропалинальное движение перемещало пищу во рту вперед и назад. [ 28 ]
Функция купола
[ редактировать ]Tylocephale известен своим выступающим куполом, характерным для других пахицефалозаврид, который был покрыт кератином . Функция купола самой Tylocephale подробно не анализировалась, но аналогичный купол Prenocephale был протестирован биологами Эриком Снивли и Адамом Коксом в 2008 году. В ходе исследования был проведен анализ методом конечных элементов 2D и 3D черепов пахицефалозавров, который обнаружил, что высокая сводчатые купола, подобные куполу Тилоцефала, могли выдерживать более высокие силы удара, чем у других пахицефалозавров. Купол Tylocephale наиболее похож на купол Pachycephalosaurus с наличием сросшихся швов, бугорков на нижней челюсти и носовой кости , а также расширенных полочек на чешуйчатой кости. Эти черты отсутствуют у примитивных таксонов, таких как Stegoceras, Homalocephale и Goyocephale . Дополнительные украшения Tylocephale, Prenocephale и Pachycephalosaurus позволяют предположить, что купол предназначался не только для демонстрации или распознавания видов , но и для агонистического поведения, такого как бодание головой. [ 29 ] Другое исследование показало, что корреляция между ударением головой и морфологией черепа, обнаруженная у живых животных, также существовала у изученных пахицефалозавров. Stegoceras и Prenocephale имели форму черепа, похожую на снежного барана , с губчатой костью, защищающей мозг. Они также имели сходство в распределении компактных и губчатых областей с снежным бараном, белобрюхим дукером и жирафом . Белобрюхий дукер оказался ближайшим морфологическим аналогом Stegoceras ; у этого вида, бьющегося головой, купол меньшего размера, но такой же округлый. Stegoceras был лучше способен рассеивать силу, чем парнокопытные, которые бодались головами с большой силой, но менее васкуляризированные купола старых пахицефалозавров и, возможно, пониженная способность к заживлению после травм, выступали против такого боя у пожилых людей. В ходе исследования также проверили влияние кератинового покрытия купола и обнаружили, что оно улучшает производительность. [ 30 ]
Палеосреда
[ редактировать ]
Формация Барун-Гойот, основанная на отложениях , считается позднемеловой по возрасту (средний-верхний кампан). [ 31 ] [ 32 ] Эта формация в основном характеризуется серией красных пластов , преимущественно светлых песков (желтоватых, серо-коричневых и реже красноватых), местами сцементированных. На всей территории также распространены песчанистые аргиллиты (часто красного цвета), алевролиты , конгломераты и крупномасштабная троговая поперечная слоистость песков. бесструктурные, среднезернистые, мелкозернистые и очень мелкозернистые песчаники Кроме того, в отложениях барун-гойотской свиты преобладают . Отложения этой формации отлагались на аллювиальных равнинах (равнинах, состоящих из отложений горных рек ), озерных и эоловых палеосредах в условиях относительно засушливого и полузасушливого климата . [ 33 ] [ 34 ] [ 31 ]
Tylocephale является эндемиком формации Барун-Гойот, которая также была домом для многих других позвоночных животных , включая анкилозаврид Saichania , Tarchia и Zaraapelta ; [ 35 ] [ 36 ] альваресзавриды Khulsanurus и Parvicursor ; [ 37 ] птицы Гобипипус , Гобиптерикс и Голландия ; [ 38 ] протоцератопсиды Bagaceratops и Breviceratops ; [ 32 ] дромеозавриды Куру и Шри ; [ 39 ] [ 40 ] халшкарапторин Hulsanpes ; [ 41 ] и овирапториды Conchoraptor , Heyuannia и Nemegtomaia . [ 42 ] [ 43 ] Остальные таксоны представлены крупным титанозавром Quaesitosaurus , [ 44 ] и большое разнообразие млекопитающих и чешуйчатых . [ 45 ] [ 46 ] [ 47 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Фанти, Ф.; Кантелли, Л.; Анжеликола, Л. (01 апреля 2018 г.). «Карты высокого разрешения местонахождений Хулсан и Немегт (бассейн Немегта, южная Монголия): стратиграфические значения» . Палеогеография, Палеоклиматология, Палеоэкология . ПОЗДНЕМЕЛОВАЯ ЭКОСИСТЕМА НЕМЕГТА: РАЗНООБРАЗИЕ, ЭКОЛОГИЯ И ГЕОЛОГИЧЕСКАЯ ОСОБЕННОСТЬ. 494 : 14–28. Бибкод : 2018PPP...494...14F . дои : 10.1016/j.palaeo.2017.10.015 . ISSN 0031-0182 .
- ^ Jump up to: а б Карри, Пи Джей (2016). Динозавры Гоби: По следам польско-монгольских экспедиций . Палеонтология Полоника , 67 , 83-100.
- ^ Лавас, младший (2016). Зофия Килан-Яворовская и палеонтологические экспедиции Гоби. Палеонтология Полоника , 67 , 13-24.
- ^ Марьянская, Т. (1977). Ankylosauridae (Dinosauria) из Монголии. Палеонтология Полоника , 37 , 85-151.
- ^ Осмольская, Гальшка (5 августа 1982 г.). «Hulsanpes perlei ngnsp. (Deinonychosauria, Saurischia, Dinosauria) из верхнемеловой барун-гойотской свиты Монголии» . Новогодний ежегодник по геологии и палеонтологии - ежемесячные выпуски . 1982 (7): 440–448. дои : 10.1127/njgpm/1982/1982/440 .
- ^ Jump up to: а б с д и ж г Марьянская, Т.; Осмольска, Х. (1974). «Пахицефалозаврия, новый подотряд птицетазовых динозавров» (PDF) . Palaeontologica Polonica (30): 45–102.
- ^ Jump up to: а б Пол, GS (2016). Принстонский полевой справочник по динозаврам (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. п. 269. ИСБН 9780691167664 .
- ^ Гальтон, Питер М.; Сьюс, Ханс-Дитер (1 марта 1983 г.). «Новые данные о динозаврах-пахицефалозавридах (Reptilia: Ornithischia) из Северной Америки» . Канадский журнал наук о Земле . 20 (3): 462–472. Бибкод : 1983CaJES..20..462G . дои : 10.1139/e83-043 . ISSN 0008-4077 .
- ^ Ромер, Альфред Шервуд (1976). «Человек». Остеология рептилий . Издательство Чикагского университета. ISBN 978-0-226-72491-1 .
- ^ Гальтон, Питер М.; Сьюс, Ханс-Дитер (1 марта 1983 г.). «Новые данные о динозаврах-пахицефалозавридах (Reptilia: Ornithischia) из Северной Америки» . Канадский журнал наук о Земле . 20 (3): 462–472. Бибкод : 1983CaJES..20..462G . дои : 10.1139/e83-043 . ISSN 0008-4077 .
- ^ Бойд, Клинт А. (22 декабря 2015 г.). «Систематические связи и биогеографическая история птицетазовых динозавров» . ПерДж . 3 : е1523. дои : 10.7717/peerj.1523 . ISSN 2167-8359 . ПМК 4690359 . ПМИД 26713260 .
- ^ Jump up to: а б Дьедонне, П.-Э.; Крузадо-Кабальеро, П.; Годфруа, П.; Тортоса, Т. (3 октября 2021 г.). «Новая филогения цераподских динозавров» . Историческая биология . 33 (10): 2335–2355. Бибкод : 2021HBio...33.2335D . дои : 10.1080/08912963.2020.1793979 . ISSN 0891-2963 . S2CID 221854017 .
- ^ Батлер, Р.Дж., Апчерч, П., и Норман, Д.Б. (2008). Филогения птицетазовых динозавров. Журнал систематической палеонтологии , 6 (1), 1-40.
- ^ Хайлу, Ты; Син, Сюй; Сяолинь, Ван (07 сентября 2010 г.). «Новый род Psittacosauridae (Dinosauria: Ornithopoda) и происхождение и ранняя эволюция маргиноцефалических динозавров» . Acta Geologica Sinica — английское издание . 77 (1): 15–20. Бибкод : 2003AcGlS..77...15Y . дои : 10.1111/j.1755-6724.2003.tb00105.x . S2CID 89051352 .
- ^ Салливан, РМ (2006). «Таксономический обзор Pachycephalosauridae (Dinosauria: Ornithischia)» . Бюллетень Музея естественной истории и науки Нью-Мексико (35): 347–365.
- ^ Эванс, Дэвид С.; Ваврек, Мэтью Дж.; Ларссон, Ханс CE (01 декабря 2015 г.). «Черепные останки пахицефалозаврид (Dinosauria: Ornithischia) из поздней меловой (маастрихтской) формации Сколлард в Альберте, Канада» . Палеобиоразнообразие и палеосреда . 95 (4): 579–585. Бибкод : 2015PdPe...95..579E . дои : 10.1007/s12549-015-0188-x . ISSN 1867-1608 . S2CID 129253322 .
- ^ Вудрафф, округ Колумбия, Гудвин, МБ, Лайсон, Т. Р., и Эванс, округ Колумбия (2021). Онтогенез и изменчивость пахицефалозавринового динозавра Sphaerotholus buchholtzae и его систематика внутри рода. Зоологический журнал Линнеевского общества , 193 (2), 563-601.
- ^ Лонгрич, Н.Р., Санки, Дж., и Танке, Д. (2010). Texacephale langstoni, новый род пахицефалозаврид (Dinosauria: Ornithischia) из верхней кампанской формации Агуджа, южный Техас, США. Меловые исследования , 31 (2), 274–284.
- ^ Ганглофф, Р.А., Фиорилло, А.Р., и Нортон, Д.В. (2005). Первый пахицефалозавр (Dinosauria) из палеоарктики Аляски и его палеогеографическое значение. Журнал палеонтологии , 79 (5), 997-1001.
- ^ Салливан, Р.М., и Лукас, С.Г. (2006). Динозавр-пахицефалозаврид Stegoceras validum из верхнемеловой формации Fruitland, бассейн Сан-Хуан, Нью-Мексико . Бюллетень Музея естественной истории и науки Нью-Мексико , 35 , 329–330.
- ^ Эванс, Дэвид С.; Браун, Калеб М.; Ты, Хайлу; Кампионе, Николас Э. (2021). «Описание и пересмотренный диагноз первого зарегистрированного пахицефалозавриды в Азии, Sinocephale bexelli gen. nov., из верхнего мела Внутренней Монголии, Китай» . Канадский журнал наук о Земле . 58 (10): 981–992. Бибкод : 2021CaJES..58..981E . doi : 10.1139/cjes-2020-0190 . ISSN 0008-4077 . S2CID 244227050 .
- ^ Салливан, РМ (2006). Таксономический обзор Pachycephalosauridae (Dinosauria: Ornithischia). Бюллетень Музея естественной истории и науки Нью-Мексико , 35 (47), 347–365.
- ^ Jump up to: а б Эванс, Дэвид С.; Хаяси, Сёдзи; Тиба, Кентаро; Ватабе, Махито; Райан, Майкл Дж.; Ли, Юнг-Нам; Карри, Филип Дж.; Цогтбаатар, Хишигджав; Барсболд, Ринчен (01 апреля 2018 г.). «Морфология и гистология новых образцов черепов Pachycephalosauridae (Dinosauria: Ornithischia) из нэмэгтинской свиты, Монголия» . Палеогеография, Палеоклиматология, Палеоэкология . ПОЗДНЕМЕЛОВАЯ ЭКОСИСТЕМА НЕМЕГТА: РАЗНООБРАЗИЕ, ЭКОЛОГИЯ И ГЕОЛОГИЧЕСКАЯ ОСОБЕННОСТЬ. 494 : 121–134. Бибкод : 2018PPP...494..121E . дои : 10.1016/j.palaeo.2017.11.029 . ISSN 0031-0182 .
- ^ Шотт, Райан К.; Эванс, Дэвид К. (ноябрь 2016 г.). «Краниальные вариации и систематика Foraminacephale brevis gen. nov. и разнообразие пахицефалозавридных динозавров (Ornithischia: Cerapoda) в группе реки Белли в Альберте, Канада» . Зоологический журнал Линнеевского общества . дои : 10.1111/zoj.12465 .
- ^ Марьянская, Т.; Чепмен, RE; Вейшампель, Д.Б. (2004). «Пахицефалозаврия». В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 464–477 . ISBN 978-0-520-24209-8 .
- ^ Сьюс, HD и Гальтон, PM (1987). «Анатомия и классификация североамериканских пахицефалозавров (Dinosauria: Ornithischia)». Палеонтографика Абтейлунг А. 198 : 1–40.
- ^ Набавизаде, А. (2016). «Эволюционные тенденции в механике приводящих челюстей птицетазовых динозавров» . Анатомическая запись . 299 (3): 271–294. дои : 10.1002/ar.23306 . ПМИД 26692539 .
- ^ Барретт, премьер-министр (1998). Травоядность нептичьих динозавров (докторская диссертация, Кембриджский университет).
- ^ Снайвли, Э.; Кокс, А. (2008). «Структурная механика черепа пахицефалозавра допускает бодание головой» . Электронная палеонтология . 11 : 1–17.
- ^ Снайвли, Эрик; Теодор, Джессика М. (28 июня 2011 г.). «Общие функциональные корреляты поведения при ударе головой у пахицефалозавра Stegoceras validum (Ornithischia, Dinosauria) и боевых парнокопытных» . ПЛОС ОДИН . 6 (6): e21422. Бибкод : 2011PLoSO...621422S . дои : 10.1371/journal.pone.0021422 . ISSN 1932-6203 . ПМК 3125168 . ПМИД 21738658 .
- ^ Jump up to: а б Эберт, Д.А. (2018). «Стратиграфия и палеоэкологическая эволюция богатой динозаврами последовательности Баруунгойот-Нэмегт (верхний мел), бассейн Нэмэгт, южная Монголия». Палеогеография, Палеоклиматология, Палеоэкология . 494 : 29–50. Бибкод : 2018PPP...494...29E . дои : 10.1016/j.palaeo.2017.11.018 .
- ^ Jump up to: а б Чепинский, Л. (2019). «Онтогенез и изменчивость протоцератопсидного динозавра Bagaceratops rozhdestvenskyi из позднего мела пустыни Гоби». Историческая биология . 32 (10): 1394–1421. дои : 10.1080/08912963.2019.1593404 . S2CID 132780322 .
- ^ Градзинский, Р.; Ежикевич, Т. (1974). «Осадконакопление формации Барун-Гойот» (PDF) . Палеонтология Полоника . 30 : 111–146.
- ^ Градзинский, Р.; Яворовска, ЗК; Марьянская, Т. (1977). «Верхнемеловые джадохтинская, барун-гоётская и немегтинская свиты Монголии, включая замечания по предыдущим подразделениям» . Акта Геологика Полоника . 27 (3): 281–326.
- ^ Арбур, ВМ; Карри, Пи Джей; Бадамгарав, Д. (2014). "Анкилозавридные динозавры верхнемеловых баруунгойотских и немегтских формаций Монголии " Зоологический журнал Линнеевского общества . 172 (3): 631–652. дои : 10.1111/zoj.12185 .
- ^ Парк, Ж.-Ю.; Ли, Ю.Н.; Карри, Пи Джей; Райан, MJ; Белл, П.; Сиссонс, Р.; Коппельхус, Е.Б.; Барсболд, Р.; Ли, С.; Ким, С.-Х. (2021). «Новый скелет анкилозаврид из верхнемеловой формации Баруунгойот в Монголии: его значение для посткраниальной эволюции анкилозаврид» . Научные отчеты . 11 (4101): 4101. doi : 10.1038/s41598-021-83568-4 . ПМЦ 7973727 . ПМИД 33737515 .
- ^ Аверьянов, АО; Лопатин, А.В. (2022). «Переоценка Parvicursor remotus из позднего мела Монголии: значение для филогении и таксономии динозавров-теропод-альваресзаврид». Журнал систематической палеонтологии . 19 (16): 1097–1128. дои : 10.1080/14772019.2021.2013965 . S2CID 247222017 .
- ^ Белл, Алисса К.; Кьяппе, Луис М.; Эриксон, Грегори М.; Сузуки, Сигэру; Ватабе, Махито; Барсболд, Ринчен; Цогтбаатар, К. (2010). «Описание и экологический анализ Hollanda luceria, позднемеловой птицы из пустыни Гоби (Монголия)» . Меловые исследования . 31 (1): 16–26. Бибкод : 2010CrRes..31...16B . дои : 10.1016/j.cretres.2009.09.001 . ISSN 0195-6671 .
- ^ Тернер, А.Х.; Монтанари, С.; Норелл, Массачусетс (2021). «Новый дромеозаврид из позднемелового местонахождения Хулсан в Монголии» (PDF) . Американский музей Novitates (3965): 1–48. дои : 10.1206/3965.1 . hdl : 2246/7251 . ISSN 0003-0082 . S2CID 231597229 .
- ^ Наполи, JG; Рубеншталь, А.А.; Бхуллар, Б.-А.С.; Тернер, А.Х.; Норелл, Массачусетс (2021). «Новый дромеозаврид (Dinosauria: Coelurosauria) из Хулсана, Центральная Монголия» (PDF ) Американский музей Novitates (3982): 1–47. дои : 10.1206/3982.1 . hdl : 2246/7286 . ISSN 0003-0082 . S2CID 243849373 .
- ^ Кау, А.; Мадзия, Д. (2018). «Переописание и родство Hulsanpes perlei (Dinosauria, Theropoda) из верхнего мела Монголии» . ПерДж . 6 : е4868. дои : 10.7717/peerj.4868 . ПМЦ 5978397 . ПМИД 29868277 .
- ^ Фанти, Ф.; Карри, Пи Джей; Бадамгарав, Д.; Лалуэса-Фокс, К. (2012). «Новые экземпляры немегтомайи из баруунгойотской и нэмегтинской свит (поздний мел) Монголии » ПЛОС ОДИН 7 (2): e31330. Бибкод : 2012PLoSO... 731330F дои : 10.1371/journal.pone.0031330 . ПМЦ 3275628 . ПМИД 22347465 .
- ^ Фанстон, Г.Ф.; Мендонка, ЮВ; Карри, Пи Джей; Барсболд, Р.; Барсболд, Р. (2018). «Анатомия, разнообразие и экология овирапторозавра в бассейне Немегта». Палеогеография, Палеоклиматология, Палеоэкология . 494 : 101–120. Бибкод : 2018PPP...494..101F . дои : 10.1016/j.palaeo.2017.10.023 .
- ^ Курзанов С.М.; Банников, А.Ф. (1983). «Новый зауропод из верхнего мела Монголии» . Палеонтологический журнал . 2 : 90−96.
- ^ Килан-Яворовска, З. (1974). «Мультитуберкулезная сукцессия в позднем мелу пустыни Гоби (Монголия)» (PDF) . Палеонтология Полоника . 30 : 23–44.
- ^ Градзинский, Р.; Ежикевич, Т. (1974). «Эоловые и связанные с ними отложения верхнего мела пустыни Гоби (Монголия)», носящие динозавров и млекопитающих. Осадочная геология . 12 (4): 249–278. Бибкод : 1974SedG...12..249G . дои : 10.1016/0037-0738(74)90021-9 .
- ^ Кецин, Г.; Норелл, Массачусетс (2000). «Таксономический состав и систематика позднемеловых комплексов ящериц из Ухаа Толгода и прилегающих местонахождений, монгольская пустыня Гоби» . Новитаты Американского музея (249): 1–118. doi : 10.1206/0003-0090(2000)249<0001:TCASOL>2.0.CO;2 . hdl : 2246/1596 . S2CID 129367764 .
