Bovidae
| Bovidae Временной диапазон: Ранний миоцен настоящее время
| |
|---|---|

| |
| Пример Bovidae (по часовой стрелке сверху слева) — аддакс ( Addax nasomaculatus ), домашний крупный рогатый скот ( Bos taurus ), горная газель ( Gazella gasella ), импала ( Aepyceros melampus ), голубая антилопа гну ( Connochaetes taurinus ) и муфлон ( Ovis gmelini ). | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Парнокопытные |
| Инфрапорядок: | Овца |
| Суперсемейство: | Бовоидеа |
| Семья: | Bovidae Грей , 1821 г. |
| Типовой род | |
| Бос | |
| Подсемейства | |
Альтернативная таксономия:
| |
Bovidae рогатый составляют биологическое семейство парнокопытных жвачных , крупный млекопитающих в которое входят скот , яки , бизоны , буйволы , антилопы (включая козлиных антилоп ), овцы и козы . Члена этого семейства называют быковым . насчитывающее 143 существующих вида и 300 известных вымерших видов Семейство Bovidae, , состоит из 11 (или двух) основных подсемейств и тринадцати основных триб. Семейство возникло 20 миллионов лет назад, в раннем миоцене .
Крупные рогатые животные сильно различаются по размеру и окраске шерсти . За исключением некоторых одомашненных форм , все самцы крупного рогатого скота имеют два или более рогов , а у многих видов рога есть и у самок. Размер и форма рогов сильно различаются, но основная структура всегда представляет собой одну или несколько пар простых костных выступов без ветвей, часто имеющих спиральную, скрученную или рифленую форму, каждый из которых покрыт постоянной оболочкой из кератина . У большинства быков от 30 до 32 зубов.
Большинство быков ведут дневной образ жизни . Социальная активность и кормление обычно достигают пика на рассвете и в сумерках. Бовиды обычно отдыхают перед рассветом, в полдень и после наступления темноты. У них имеются различные способы социальной организации и социального поведения , которые подразделяются на одиночное и стадное поведение. Крупные рогатые животные используют различные формы голосового, обонятельного и материального общения. Большинство видов поочередно питаются и жуют пищу в течение дня. В то время как мелкие быки кормятся в плотных и закрытых местах обитания, более крупные виды питаются растительностью с высоким содержанием клетчатки на открытых лугах. Большинство быков полигинны . Взрослые быки спариваются не реже одного раза в год, а более мелкие виды могут спариваться даже дважды. У некоторых видов новорожденные крупного рогатого скота остаются скрытыми от недели до двух месяцев, и их матери регулярно выкармливают их; у других видов новорожденные являются последователями, сопровождающими своих самок, а не склонными оставаться скрытыми.
Наибольшее разнообразие быков встречается в Африке . Максимальная концентрация вида — в саваннах Восточной Африки . Другие виды крупного рогатого скота также встречаются в Европе, Азии и Северной Америке. Bovidae включает трех из пяти одомашненных млекопитающих, использование которых распространилось за пределы их первоначального ареала, а именно крупный рогатый скот, овцы и козы. Молочные продукты , такие как молоко , масло и сыр , производятся в основном из домашнего скота. Бовидов также выращивают ради кожи , мяса и шерсти .
Именование и этимология
[ редактировать ]Название Bovidae дал британский зоолог Джон Эдвард Грей в 1821 году. [ 1 ] Слово «Bovidae» представляет собой комбинацию префикса bov- (происходящего от латинского bos , «бык», через позднелатинский bovinus ) и суффикса -idae . [ 2 ]
Таксономия
[ редактировать ]Семейство Bovidae относят к отряду Парнокопытные который входят парнокопытные ( в ). Он включает 143 современных вида, составляющих почти 55% копытных , и 300 известных вымерших видов. [ 3 ]
До начала XXI века считалось, что семейство Moschidae (кабарга) сестрой является Cervidae . Однако филогенетическое исследование 2003 года, проведенное Александром Хассанином ( Национальный музей естественной истории, Франция ) и его коллегами, основанное на митохондриальном и ядерном анализе, показало, что Moschidae и Bovidae образуют родственную кладу Cervidae . Согласно исследованию, Cervidae отделились от клады Bovidae-Moschidae 27–28 миллионов лет назад. [ 4 ] Следующая кладограмма основана на исследовании 2003 года. [ 4 ]



Молекулярные исследования подтвердили монофилию семейства Bovidae (группа организмов, состоящая из предкового вида и всех их потомков). [ 5 ] [ 6 ] Количество подсемейств Bovidae оспаривается: предполагается, что их может быть от десяти до двух подсемейств. [ 6 ] Однако молекулярные, морфологические и ископаемые данные указывают на существование восьми различных подсемейств: Aepycerotinae (состоящие только из импалы), Alcelaphinae (бонтебок, антилопа гну и их родственники), Antilopinae (несколько антилоп, газелей и их родственников), Bovinae (крупный рогатый скот , буйволы, бизоны и другие антилопы), Caprinae (козы, овцы, козерог, сероу и родственники), Cephalophinae (дукеры), Hippotraginae (аддакс, орикс и родственники) и Reduncinae (антилопы ридбак и коб). Кроме того, известны три вымерших подсемейства: Hypsodontinae (средний миоцен ), Oiocerinae ( туролийский ) и подсемейство Tethytraginae , к которому относится Tethytragus (средний миоцен ). [ 7 ] [ 8 ]
В 1992 году Алан В. Джентри из Музея естественной истории в Лондоне разделил восемь основных подсемейств Bovidae на две основные клады на основе их эволюционной истории: Boodontia, в которую входили только Bovinae, и Aegodontia, состоявшая из остальные подсемейства. У боодонтов несколько примитивные зубы, напоминающие зубы быков , тогда как у эгодонтов зубы более развиты , как у коз. [ 9 ]
Существуют разногласия по поводу признания Peleinae и Pantholopinae , включающих роды Pelea и Pantholops соответственно, как подсемейства . В 2000 году американский биолог Джордж Шаллер и палеонтолог Элизабет Врба предложили включить Пелею в состав Reduncinae , [ 10 ] хотя серый ребок, единственный вид Pelea , по морфологии сильно отличается от кобов и редунцинов. [ 11 ] Пантолопсы , ранее отнесенные к Antilopinae , позже были помещены в отдельное подсемейство Pantholopinae . Однако молекулярный и морфологический анализ подтверждает включение Pantholops в Caprinae . [ 12 ]
Ниже представлена кладограмма, основанная на Yang et al . , 2013 г. и Каламари, 2021 г.: [ 13 ] [ 14 ] [ 15 ]
| Bovidae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||







Альтернативно, все представители Aegodontia могут быть отнесены к подсемейству Antilopinae, при этом отдельные подсемейства в этом случае считаются племенами. [ 14 ] [ 15 ]
Эволюционная история
[ редактировать ]Ранний миоцен и ранее
[ редактировать ]
В раннем миоцене полорогие начали отделяться от оленьих (оленей) и жирафов . Самые ранние полорогие, присутствие которых в Африке и Евразии во второй половине раннего миоцена (20 млн лет назад ) было установлено, были мелкими животными, чем-то похожими на современных газелей и, вероятно, обитавшими в лесной среде. [ 16 ] Эотрагус , самый ранний из известных быков, весил 18 кг (40 фунтов) и был почти такого же размера, как газель Томсона . [ 17 ] В начале своей эволюционной истории быки разделились на две основные клады: Boodontia (евразийского происхождения) и Aegodontia (африканского происхождения). Этот ранний раскол между Boodontia и Aegodontia объясняется континентальным водоразделом между этими массивами суши. Когда эти континенты позже воссоединились, этот барьер был устранен, и обе группы расширились на территорию другой. [ 18 ] Племена бовини и трагелафины разошлись в раннем миоцене. [ 19 ] Известно, что полорогие достигли Америки в плейстоцене, перейдя Берингов мост . [ 17 ]
Современные роды Alcelaphinae появились в плиоцене . Вымерший род Alcelaphine Paramularius , который был такого же размера, как и хартебест, как полагают, возник в плиоцене, но вымер в среднем плейстоцене . [ 6 ] Несколько родов Hippotraginae известны с плиоцена и плейстоцена. Это подсемейство, по-видимому, отделилось от Alcelaphinae во второй половине раннего миоцена. [ 19 ] Считается, что Bovinae отделились от остальных Bovidae в раннем миоцене . [ 20 ] Боселафины вымерли в Африке в раннем плиоцене; их последние окаменелости были раскопаны в Лангебанвеге (Южная Африка) и Лотагаме (Кения). [ 21 ]
Средний миоцен
[ редактировать ]Средний миоцен ознаменовал распространение быков в Китай и на Индийский субконтинент. [ 17 ] По мнению Врбы, распространение подсемейства Alcelaphinae началось во второй половине среднего миоцена. [ 6 ] Племена Caprinae, вероятно, разошлись в начале среднего миоцена. Caprini появились в среднем миоцене и, по-видимому, были заменены другими быками и оленьими в Евразии. [ 22 ] Самые ранние окаменелости антилопинов относятся к среднему миоцену, хотя исследования показывают существование подсемейства из раннего миоцена. Видообразование произошло трибы Antilopini в среднем или верхнем миоцене, преимущественно в Евразии. Триба Neotragini , по-видимому, появилась в Африке к концу миоцена и получила широкое распространение к плиоцену. [ 19 ]
Поздний миоцен
[ редактировать ]К концу миоцена, около 10 млн лет назад, полорогие быстро диверсифицировались , что привело к созданию 70 новых родов. [ 17 ] Эта радиация позднего миоцена отчасти объяснялась тем, что многие коровы адаптировались к более открытым местам обитания на лугах. [ 16 ] Aepycerotinae впервые появились в позднем миоцене, при этом существенной разницы в размерах примитивной и современной импалы не отмечено. [ 23 ] Окаменелости яйцебыков, трибы Caprinae, в Африке датируются поздним миоценом. [ 19 ] Самые ранние окаменелости Hippotragine относятся к позднему миоцену и были раскопаны в таких местах, как Лотагам и долина Аваш . [ 19 ] Первые африканские окаменелости Reduncinae датируются 6-7 млн лет назад. [ 24 ] Reduncinae и Peleinae, вероятно, разошлись в середине миоцена. [ 6 ]
Характеристики
[ редактировать ]
Все бычьи имеют одинаковую основную форму - морду с тупым концом, одну или несколько пар рогов (обычно присутствующих у самцов) сразу за овальными или заостренными ушами, отчетливую шею и конечности, а также хвост, различающийся по длине и густоте. вид. [ 25 ] У большинства быков наблюдается половой диморфизм : самцы обычно крупнее и тяжелее самок. Половой диморфизм более выражен у коров среднего и крупного размера. У всех полорогих на каждой ноге по четыре пальца — они ходят на двух центральных (копытцах ) , а два внешних ( прибылые пальцы ) гораздо меньше и редко касаются земли. [ 3 ]
Размер крупного рогатого скота сильно различается: гаур может весить более 1500 кг (3300 фунтов) и иметь высоту в холке 2,2 м (87 дюймов). [ 26 ] Королевская антилопа , напротив, имеет рост всего 25 см (9,8 дюйма) и весит не более 3 кг (6,6 фунта). [ 27 ] Клипспрингер , еще одна маленькая антилопа, имеет высоту 45–60 см ( 18–24 дюйма) в холке и весит всего 10–20 кг (22–44 фунта). [ 28 ]
Различия наблюдаются в окраске шерсти : от бледно-белой (как у арабского орикса ). [ 29 ] до черного цвета (как у черного антилопы гну ). [ 30 ] только промежуточные оттенки, такие как коричневый и красновато-коричневый (как у ридбака ). Однако обычно наблюдаются [ 31 ] У некоторых видов шерсть самок и молодых особей светлая, а у самцов с возрастом темнеет. Как и у антилопы гну , на шерсти могут быть заметные или слабые полосы. У некоторых видов, таких как аддакс , цвет шерсти может меняться в зависимости от сезона. [ 32 ] ароматические и сальные железы . Часто присутствуют [ 25 ]

Некоторые виды, такие как гемсбок , черная антилопа и газель Гранта , замаскированы сильно разрушительными отметинами на лице, скрывающими хорошо узнаваемый глаз. [ 34 ] Многие виды, такие как газели, можно сделать плоскими и, следовательно, слиться с фоном с помощью контрзатенения . [ 35 ] Очертания многих быков разделены яркой разрушительной окраской, а сильно контрастирующие узоры помогают задержать распознавание хищниками. [ 36 ] Однако все Hippotraginae (включая гемсбока) имеют бледные тела и лица с заметными отметинами. Зоолог Тим Каро считает, что это трудно объяснить, но, учитывая, что эти виды ведут дневной образ жизни, он предполагает, что эти отметины могут использоваться для общения. Сильно контрастная окраска ног обычна только у Bovidae, у которых, например, Bos , Ovis , бонтебок и гемсбок имеют белые чулки. Опять же, общение является вероятной функцией. [ 33 ]
За исключением некоторых одомашненных форм, все самцы крупного рогатого скота имеют рога, а у многих видов рогами обладают и самки. Размер и форма рогов сильно различаются, но основная структура представляет собой пару простых костных выступов без ветвей, часто имеющих спиральную, скрученную или рифленую форму, каждый из которых покрыт постоянной оболочкой из кератина. Хотя рога встречаются в одной паре почти у всех видов быков, есть исключения, такие как четырехрогая антилопа. [ 37 ] и овца Иакова . [ 38 ] [ 39 ] Уникальное строение рогов — единственная однозначная морфологическая особенность быков, отличающая их от других пекоран . [ 40 ] [ 41 ] Существует высокая корреляция между морфологией рогов и боевым поведением особи. Например, длинные рога предназначены для борьбы и фехтования, а изогнутые – для тарана. [ 42 ] Самцы с рогами, направленными внутрь, моногамны и одиночки, а с рогами, направленными наружу, как правило, полигинны . Эти результаты не зависели от размера тела. [ 43 ]
Развитие мужских рогов связано с половым отбором . [ 44 ] [ 45 ] Рога представляют собой небольшие шипы у моногамных дукеров и других мелких антилоп, тогда как у полигинных они крупные и сложной формы (например, имеют спиральную структуру, как у гигантской канны ). Таким образом, рога в некоторой степени отражают степень конкуренции между самцами одного вида. [ 31 ] Однако наличие рогов у самок, скорее всего, обусловлено естественным отбором . [ 44 ] [ 46 ] Рога самок обычно меньше, чем у самцов, а иногда и другой формы. Считается, что рога самок быков развились для защиты от хищников или для выражения территориальности, поскольку нетерриториальные самки, которые могут использовать крипсис для защиты от хищников, часто не имеют рогов. [ 46 ] Рога у самок есть только у половины родов быков, причем самки этих родов тяжелее, чем самки остальных. Самки используют рога преимущественно для колющих ударов. [ 47 ]
Анатомия
[ редактировать ]
У крупного рогатого скота третье и четвертое метаподии объединены в пушечную кость . Локтевая редуцируются и малоберцовая кости и срастаются с лучевой и большеберцовой костями соответственно. длинные лопатки Имеются , а ключицы отсутствуют. У жвачных животных желудок состоит из четырех камер: рубца (80%), книжки , сетки и сычуга . Инфузории жирные и бактерии рубца ферментируют сложную целлюлозу в более простые кислоты , которые затем всасываются через стенку рубца. У крупного рогатого скота длинный тонкий кишечник ; длина тонкой кишки у крупного рогатого скота составляет 29–49 м (95–161 фут). Температура тела колеблется в течение дня; например, у коз температура может незначительно меняться от почти 37 ° C (99 ° F) ранним утром до 40 ° C (104 ° F) днем. У крупного рогатого скота температура регулируется за счет потоотделения, тогда как у коз для этого используется дыхание. Правое легкое , состоящее из четырех-пяти долей , примерно в 1,5 раза больше левого, у которого три доли. [ 3 ] [ 25 ]
Зубной ряд
[ редактировать ]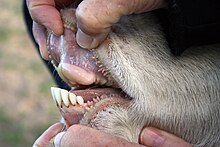
У большинства быков от 30 до 32 зубов. [ 31 ] В то время как верхние резцы отсутствуют, верхние клыки либо редуцированы, либо отсутствуют. Вместо верхних резцов у коров есть толстый и жесткий слой ткани, называемый зубной подушечкой , который обеспечивает поверхность для захвата травы и листвы. Они бывают гипсодонтными и селенодонтными , так как коренные зубы и премоляры имеют низкую коронку и серповидные бугорки . Нижние резцы и клыки выступают вперед. За резцами следует длинная беззубая щель, известная как диастема . [ 48 ] Общая зубная формула крупного рогатого скота составляет 0,0,2-3,3 3,1.3,3 . Большинство представителей семейства травоядны , но большинство дукеров всеядны . Как и у других жвачных животных, у коров четырехкамерный желудок, что позволяет им переваривать растительный материал, например траву , которую многие другие животные не могут использовать. Жвачные животные (и некоторые другие, такие как кенгуру , кролики и термиты ) способны использовать микроорганизмы, живущие в их кишечнике, для расщепления целлюлозы путем ферментации . [ 3 ]
Экология и поведение
[ редактировать ]

У полорогих имеются различные способы социальной организации и социального поведения, которые подразделяются на одиночное и стадное поведение. Кроме того, каждый из этих типов можно разделить на территориальное и нетерриториальное поведение. [ 31 ] Мелкие быки, такие как клипспрингер, ориби и стенбок, обычно ведут одиночный образ жизни и территориальны. Они владеют небольшими территориями, на которые другим представителям вида не разрешен вход. Эти антилопы образуют моногамные пары. Многие виды, такие как дик-дик, используют феромонов выделения из предглазничных желез , а иногда и навоз, чтобы пометить свою территорию. [ 49 ] Потомство рассеивается в подростковом возрасте, и самцы должны приобрести территории до спаривания. [ 3 ] Бушбак . — единственный крупный рогатый скот, который ведет одиночный и нетерриториальный образ жизни Эта антилопа практически не проявляет агрессии, склонна к самоизоляции или формированию разрозненных стад, хотя в благоприятной среде обитания несколько бушбаков можно встретить довольно близко друг к другу. [ 50 ]
За исключением цефалофинов (дукеров), трагелафинов (спиральнорогих антилоп) и неотрагинов, большинство африканских быков стадны и территориальны. Самцы вынуждены расселяться по достижении половой зрелости и должны образовывать свои собственные территории, в то время как самки не обязаны это делать. Самцы, не владеющие территориями, образуют холостяцкие стада. драки, как правило, становятся более ожесточенными Среди самцов происходит конкуренция за доминирование, и в ограниченные сезоны гона . За исключением мигрирующих самцов, самцы обычно владеют одной и той же территорией на протяжении всей своей жизни. [ 31 ] У водяного козла некоторым особям мужского пола, известным как «самцы-спутники», может быть разрешено проникать на территории других самцов, и им придется ждать, пока владелец состарится, чтобы они могли завладеть его территорией. [ 51 ] Известно, что спаривание лек , при котором самцы собираются вместе и соревнуются с потенциальными партнерами, существует среди топи , кобов и лехве . [ 52 ] Трагелафины, крупный рогатый скот, овцы и козы стадны и не территориальны. У этих видов самцы должны получить абсолютное доминирование над всеми остальными самцами, и драки не ограничиваются территориями. Таким образом, самцы тратят годы на рост тела. [ 31 ]
Активность
[ редактировать ]
Большинство полорогих ведут дневной образ жизни, хотя некоторые из них, такие как буйвол, бушбак, ридбак и грисбок, являются исключением. Социальная активность и кормление обычно достигают пика на рассвете и в сумерках. Полорогие обычно отдыхают перед рассветом, в полдень и после наступления темноты. Уход обычно осуществляется путем облизывания языком. Антилопы редко валяются в грязи или пыли. Антилопы гну и буйволы обычно валяются в грязи, тогда как антилопы гну и топи натирают голову и рога грязью, а затем размазывают ею свое тело. Крупные рогатые животные используют различные формы голосового, обонятельного и материального общения. Они включают в себя различные позы шеи, головы, рогов, волос, ног и ушей, чтобы передать сексуальное возбуждение, эмоциональное состояние или тревогу. Одним из таких выражений является ответ Флемена . Почувствовав опасность, полорогие обычно стоят неподвижно, с высоко поднятой головой и пристальным взглядом. Некоторые из них, такие как импала, куду и канна, могут даже прыгать на высоту нескольких футов. [ 31 ] Крупные рогатые скоты могут реветь или хрюкать, чтобы предостеречь других и отпугнуть хищников. [ 3 ] Крупные рогатые животные, такие как газели , в ответ на хищников ходят или натыкаются на ноги , совершая высокие прыжки на жестких ногах, что честно указывает как на то, что хищник был замечен, так и на то, что стоящая особь сильна и ее не стоит преследовать. [ 53 ]

В брачный период самцы во время гона ревут, чтобы сообщить самкам о своем присутствии. Овцебыки ревут во время драк между самцами, а самцы сайгаков нагнетают воздух через нос, издавая рев, чтобы отпугнуть соперничающих самцов и привлечь самок. Матери также используют голосовое общение, чтобы определить местонахождение своих телят, если их разлучают. Во время борьбы за доминирование самцы обычно занимают прямую позу с ровной мордой. [ 54 ] [ 55 ]
Техники боя различаются среди семейств коровьих, а также зависят от их телосложения. В то время как хартбесты сражаются на коленях, другие обычно сражаются на четвереньках. Газели разных размеров используют разные методы боя. Газели обычно боксируют, а в серьезных драках могут сталкиваться и фехтовать, состоя из сильных ударов с близкого расстояния. Самцы козла, козла и овцы стоят вертикально и сталкиваются друг с другом вниз. Антилопы гну используют мощные удары головой в агрессивных столкновениях. Если рога запутываются, противники двигаются по кругу, чтобы их разблокировать. Овцебыки будут таранить друг друга на высоких скоростях. Как правило, в драку вступают только два быка одинакового телосложения и уровня защиты, целью которой является определение превосходства из двоих. Индивиды, которые явно уступают другим, скорее предпочтут бежать, чем сражаться; например, неполовозрелые самцы не дерутся со взрослыми быками. Обычно быки направляют свои атаки на голову противника, а не на его тело. Рога S-образной формы, например, на Импала имеет различные секции, которые помогают таранить, удерживать и наносить удары. Серьезные драки, приводящие к травмам, случаются редко. [ 31 ] [ 54 ] [ 56 ]
Диета
[ редактировать ]
Большинство крупного рогатого скота в течение дня попеременно питаются и жуют пищу. В то время как те, кто питается концентратами, кормят и переваривают через короткие промежутки времени, кормушкам, питающимся грубыми кормами, нужны более длительные интервалы. Только мелкие виды, такие как дукер, пасутся в течение нескольких часов днем или ночью. [ 31 ] Привычки питания зависят от размера тела; в то время как мелкие быки кормятся в плотных и закрытых местах обитания, более крупные виды питаются растительностью с высоким содержанием клетчатки на открытых лугах. Подсемейства демонстрируют разные стратегии питания. В то время как виды Bovinae активно питаются свежей травой и разбросанным кормом, виды Cephalophinae (за исключением Sylvicapra ) в основном потребляют фрукты. [ 3 ] Виды Reduncinae и Hippotraginae зависят от нестабильных источников пищи, но последние специально адаптированы к засушливым районам. Представители Caprinae, будучи гибкими в питании, добывают корм даже на участках с низкой продуктивностью. Племена Alcelaphini, Hippotragini и Reduncini имеют высокую долю однодольных растений в своем рационе . Напротив, Tragelaphini и Neotragini (за исключением Ourebia ) обильно питаются двудольными растениями . [ 57 ] Никакой заметной связи между размером тела и потреблением однодольных растений не существует. [ 58 ]
Сексуальность и воспроизводство
[ редактировать ]
Большинство быков полигинны. У некоторых видов особи моногамны, что приводит к минимальной агрессии между самцами и уменьшению отбора самцов в пользу крупных размеров тела. Таким образом, половой диморфизм практически отсутствует. Самки могут быть немного крупнее самцов, возможно, из-за конкуренции между самками за приобретение территорий. Так обстоит дело с дукерами и другими мелкими быками. [ 59 ] [ 60 ] Время, необходимое для достижения половой зрелости обоими полами, широко варьируется среди быков. Половая зрелость может даже предшествовать спариванию или следовать за ним. Например, самцы импал, хотя и достигают половой зрелости к году, могут спариваться только после четырехлетнего возраста. [ 61 ] Напротив, самки варварских овец могут рожать потомство еще до того, как они достигнут половой зрелости. [ 62 ] Задержка полового созревания самцов более заметна у видов с половым диморфизмом, особенно у редунцинов, вероятно, из-за конкуренции между самцами. [ 3 ] Например, самки голубого антилопы гну становятся способными к размножению в течение года или двух после рождения, а самцы становятся половозрелыми только в возрасте четырех лет. [ 30 ]
Все быки спариваются не реже одного раза в год, а более мелкие виды могут спариваться даже дважды. Сезон спаривания у большинства коров обычно приходится на дождливые месяцы. Таким образом, пик размножения может оказаться дважды в экваториальных регионах. У овец и коз наблюдается замечательная сезонность размножения, в определении которой годовой цикл суточного фотопериода решающую роль играет . Другие факторы, которые оказывают существенное влияние на этот цикл, включают температуру окружающей среды, состояние питания, социальные взаимодействия, дату родов и период лактации. Исследование этого явления показало, что козы и овцы являются производителями короткого дня . Спаривание у большинства пород овец начинается летом или в начале осени. [ 63 ] На спаривание овец также влияет мелатонин, который опережает наступление сезона размножения; [ 64 ] и тироксин , завершающий период размножения. [ 65 ] У крупного рогатого скота течка длится не более суток, за исключением крупного рогатого скота и трагелафинов. За исключением хартебеста и топи, все быки могут обнаружить течку у самок путем анализа мочи с использованием вомероназального органа . [ 31 ] Как только самец убеждается, что у самки течка, он начинает ухаживать; эти проявления сильно различаются: от тщательно продуманных маршей у стадных видов до пылкого облизывания женских гениталий у одиночных видов. Самки, изначально невосприимчивые, в конечном итоге спариваются с самцом, добившимся доминирования над другими. Восприимчивость выражается в разрешении самцу садиться на животное и отложении хвоста самке. Копуляция обычно занимает несколько секунд. [ 31 ] [ 59 ]
Срок беременности у коров варьируется: у дукеров беременность длится от 120 до 150 дней, у африканских буйволов - от 300 до 330 дней. Обычно рождается один потомок (реже близнецы), способный самостоятельно стоять и бегать уже через час после рождения. У моногамных видов самцы помогают защищать своих детенышей, но у полигинных видов это не так. Большинство новорожденных телят остаются скрытыми от недели до двух месяцев, и их матери регулярно кормят их грудью. У некоторых видов крупного рогатого скота новорожденные начинают следовать за матерью сразу или через несколько дней, как у импалы. [ 61 ] У разных коров разные стратегии защиты молоди. Например, в то время как матери антилоп гну защищают исключительно своих детенышей, буйволы демонстрируют коллективную защиту. Отлучение от груди может произойти уже через два месяца (как у королевской антилопы) или через год (как у овцебыка ). [ 59 ] [ 60 ]
Продолжительность жизни
[ редактировать ]Большинство диких быков живут от 10 до 15 лет. Более крупные виды, как правило, живут дольше; [ 3 ] например, американский бизон может жить до 25 лет, а гаур - до 30 лет. Средняя продолжительность жизни одомашненных особей составляет почти десять лет. Например, средняя продолжительность жизни домашних коз составляет 12 лет. Обычно самцы, преимущественно у полигинных видов, имеют более короткую продолжительность жизни, чем самки. Это можно объяснить несколькими причинами: ранним расселением молодых самцов, агрессивными драками самцов с самцами, уязвимостью перед хищниками (особенно, когда самцы менее подвижны, как у куду), а также недоеданием (из-за больших размеров тело самца имеет высокий уровень питательных веществ). требования, которые могут быть не удовлетворены). [ 66 ] [ 67 ] Ричард Деспард Эстес предположил, что самки имитируют мужские вторичные половые признаки, такие как рога, чтобы защитить свое потомство мужского пола от доминирующих самцов. Эта особенность, по-видимому, была тщательно выбрана для предотвращения мужской смертности и несбалансированного соотношения полов из-за нападений агрессивных самцов и принудительного расселения молодых самцов в подростковом возрасте . [ 68 ]

Распределение
[ редактировать ]Большинство разнообразных видов крупного рогатого скота встречается в Африке. Максимальная концентрация – в саваннах восточной Африки. В зависимости от своих пищевых привычек несколько видов распространились на большие территории, и, следовательно, наблюдаются некоторые различия в морфологии зубов и конечностей. Дукеры населяют тропические экваториальные леса, ситатунга и лечве встречаются вблизи болот, канна обитает на лугах, спрингбок и сернобык встречаются в пустынях, бонго и аноа обитают в густых лесах, а горные козлы и такины обитают на больших высотах. [ 31 ] Несколько видов быков также встречаются в Европе, Азии и Северной Америке. Овцы и козы встречаются в основном в Евразии, хотя берберийские овцы и козероги составляют часть африканской фауны. Овцебык приурочен к арктической тундре . Несколько видов быков были одомашнены человеком. Одомашнивание коз и овец началось 10 тысяч лет назад, а крупный рогатый скот — около 7,5 тысяч лет назад. [ 3 ] [ 59 ]
Взаимодействие с людьми
[ редактировать ]Домашние животные
[ редактировать ]
Одомашнивание быков способствовало смещению зависимости человека от охоты и собирательства к сельскому хозяйству. К Bovidae относятся три из шести крупных домашних травоядных животных, использование которых распространилось за пределы их первоначального ареала, а именно крупный рогатый скот, овцы и козы; все они из Евразии и сейчас встречаются по всему миру. Остальные три вида — это лошадь, осел и свинья. Другими крупными быками, одомашненными, но остающимися в пределах ареала своих диких предков, являются домашний буйвол (от дикого водяного буйвола ), домашний як ), зебу (от (от дикого яка индийского зубра ), гаял (от дикого яка), зебу (от индийского зубра ), гаял (от дикого яка ). гаур ) и балийский скот (от бантенга ). [ 59 ] Некоторые антилопы были одомашнены, в том числе ориксы , аддаксы , канны и вымерший бубал-хартебест . В Древнем Египте на резных стенах изображались ориксы, аддаксы и бубалы-хартебы. [ нужна ссылка ]
Самые ранние свидетельства одомашнивания крупного рогатого скота относятся к 8000 году до нашей эры, что позволяет предположить, что этот процесс начался на Кипре и в бассейне Евфрата . [ 69 ]
Продукты животного происхождения
[ редактировать ]
Молочные продукты, такие как молоко , масло , топленое масло , йогурт , пахта и сыр, производятся в основном из домашнего скота, хотя молоко овец, коз, яков и буйволов также используется в некоторых частях мира и для производства деликатесов. Например, из буйволиного молока делают моцареллу в Италии и гулаб джамун в Индии. десерт [ 70 ] а овечье молоко используется для изготовления голубого сыра Рокфор во Франции. [ 71 ] Говядина является источником пищи с высоким содержанием цинка , селена , фосфора , железа и витаминов группы В. [ 72 ] В мясе бизона меньше жира и холестерина, чем в говядине, но больше белка. [ 73 ]
крупного рогатого скота Кожа прочная и долговечная, а дополнительным преимуществом является то, что из нее можно изготавливать кожу различной толщины — от мягкой одежной кожи до жесткой обувной кожи. В то время как кожа коз и крупного рогатого скота имеет широкое применение, овчина пригодна только для изготовления одежды. [ 74 ] свиней мериносовых . Шерсть – самая тонкая и ценная Мериносовая шерсть имеет длину 3–5 дюймов (7,6–12,7 см) и очень мягкая. Грубая шерсть, прочная и устойчивая к катышкам , используется для изготовления прочной одежды. [ 75 ] и ковры.

Костная мука — важное удобрение, богатое кальцием, фосфором и азотом , эффективно снижающее кислотность почвы . [ 76 ] Рога крупного рогатого скота использовались в качестве сосудов для питья с древности. [ 77 ]
В человеческой культуре
[ редактировать ]Bovidae фигурируют в историях, по крайней мере, со времен басен Эзопа из Древней Греции около 600 г. до н.э. Басни Эзопа включают «Ворона и овца» , «Лягушка и бык» , «Волк и ягненок» . [ 78 ] Мифологическое существо Химера, изображавшееся в виде льва, с головой козла, выступающей из спины, и хвостом, который мог заканчиваться головой змеи, было одним из потомков Тифона и Ехидны и родным братом таких чудовищ, как Цербер и Лернейская гидра . [ 79 ] Овца, синоним козы в китайской мифологии, является восьмым животным китайского зодиака и символом сыновней почтительности. [ 80 ]
Ссылки
[ редактировать ]- ^ Грабб, П. (2005). «Семейство Bovidae» . В Уилсоне, Делавэр ; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джонса Хопкинса. стр. 637–722. ISBN 978-0-8018-8221-0 . OCLC 62265494 .
- ^ «Бовиды» . Онлайн-словарь Мерриам-Вебстера . Архивировано из оригинала 11 октября 2014 года . Проверено 7 октября 2014 г.
- ^ Перейти обратно: а б с д и ж г час я дж Гомес, В.; Паттерсон, штат Калифорния; Суинтон, Дж.; Берини, Дж. «Bovidae: антилопы, крупный рогатый скот, газели, козы, овцы и их родственники» . Сеть разнообразия животных . Зоологический музей Мичиганского университета. Архивировано из оригинала 7 октября 2014 года . Проверено 7 октября 2014 г.
- ^ Перейти обратно: а б Хасанин А.; Дузери, EJP (2003). «Молекулярная и морфологическая филогения Ruminantia и альтернативное положение Moschidae» . Систематическая биология . 52 (2): 206–28. дои : 10.1080/10635150390192726 . ПМИД 12746147 .
- ^ Гейтси, Дж.; Амато, Г.; Врба, Э.; Шаллер, Г. (1997). «Кладистический анализ митохондриальной рибосомальной ДНК Bovidae» . Молекулярная филогенетика и эволюция . 7 (3): 303–19. дои : 10.1006/mpev.1997.0402 . ПМИД 9187090 .
- ^ Перейти обратно: а б с д и Фернандес, Миннесота; Врба, ЕС (2005). «Полная оценка филогенетических взаимоотношений жвачных: датированное супердерево современных жвачных животных на видовом уровне». Биологические обзоры . 80 (2): 269–302. дои : 10.1017/S1464793104006670 . ПМИД 15921052 . S2CID 29939520 .
- ^ Харрисон, Т. (2011). Палеонтология и геология эволюции человека Лаэтоли в контексте . Дордрехт: Спрингер. стр. 363–465. ISBN 978-9048-199-624 .
- ^ Демигель, Д.; Санчес, ИМ; Альба, DM; Галиндо, Дж.; Роблес, Дж. М.; Моя-Сола, С. (2012). «Первые свидетельства существования Азанзы и Моралеса, 1994 г. (Ruminantia, Bovidae) в миоцене бассейна Валлес-Пенедес (Испания)». Журнал палеонтологии позвоночных . 32 (6): 1457–62. дои : 10.1080/02724634.2012.696082 . S2CID 86664298 .
- ^ Харрисон, Т. (1997). Неогеновая палеонтология долины Манонга, Танзания: окно в эволюционную историю Восточной Африки . Нью-Йорк: Пленум Пресс. п. 113. ИСБН 978-0-306-45471-4 .
- ^ Врба, ЕС; Шаллер, Г. (2000). Антилопы, олени и их родственники: летопись окаменелостей, поведенческая экология, систематика и охрана . Нью-Хейвен: Издательство Йельского университета. ISBN 978-0300-081-428 .
- ^ Грабб, П. (2005). «Отряд Парнокопытные» . В Уилсоне, Делавэр ; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джонса Хопкинса. п. 719. ИСБН 978-0-8018-8221-0 . OCLC 62265494 .
- ^ Грабб, П. (2005). «Отряд Парнокопытные» . В Уилсоне, Делавэр ; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джонса Хопкинса. п. 699. ИСБН 978-0-8018-8221-0 . OCLC 62265494 .
- ^ Ян, Чэнчжун; Сян, Чанкуй; Ци, Вэньхуа; Ся, Шань; Ту, Фейюнь; Чжан, Сююэ; Моермонд, Тимоти; Юэ, Бисонг (01 июня 2013 г.). «Филогенетический анализ и улучшенное разрешение семейства Bovidae на основе полных митохондриальных геномов» . Биохимическая систематика и экология . 48 : 136–143. дои : 10.1016/j.bse.2012.12.005 . ISSN 0305-1978 . Архивировано из оригинала 31 января 2022 г. Проверено 31 января 2022 г.
- ^ Перейти обратно: а б Мэти, Конрад А.; Робинсон, Теренс Дж. (1 июня 1999 г.). «ЦитохромебФилогения семейства Bovidae: разрешение внутри Alcelaphini, Antilopini, Neotragini и Tragelaphini» . Молекулярная филогенетика и эволюция . 12 (1): 31–46. дои : 10.1006/mpev.1998.0573 . ISSN 1055-7903 . ПМИД 10222159 .
- ^ Перейти обратно: а б Каламари, Закари Т. (июнь 2021 г.). «Филогенетический анализ полных доказательств подтверждает новые морфологические синапоморфии Bovidae (Mammalia, Artiodactyla)» . Американский музей Novitates (3970): 1–38. дои : 10.1206/3970.1 . hdl : 2246/7267 . ISSN 0003-0082 . S2CID 235441087 . Архивировано из оригинала 14 марта 2023 г. Проверено 08 февраля 2022 г.
- ^ Перейти обратно: а б Сэвидж, RJG; Лонг, MR (1986). Эволюция млекопитающих: иллюстрированное руководство . Нью-Йорк: факты в архиве. стр. 232–5 . ISBN 978-0-8160-1194-0 .
- ^ Перейти обратно: а б с д Протеро, ДР; Шох, Р.М. (2002). Рога, бивни и ласты: эволюция копытных млекопитающих . Балтимор: Издательство Университета Джонса Хопкинса. стр. 87–90. ISBN 978-0-8018-7135-1 .
- ^ Хасанин, Д.; Дузери, Э.Дж. (1999). «Племенная радиация семейства Bovidae (Artiodactyla) и эволюция гена митохондриального цитохрома b » (PDF) . Молекулярная филогенетика и эволюция . 13 (2): 227–43. дои : 10.1006/mpev.1999.0619 . ПМИД 10603253 . Архивировано из оригинала (PDF) 20 июля 2011 г.
- ^ Перейти обратно: а б с д и Гилберт, штат Вашингтон; Асфау, Б. (2008). Homo Erectus: свидетельства плейстоцена из Среднего Аваша, Эфиопия . Беркли: Издательство Калифорнийского университета. стр. 100-1 45–84. ISBN 978-0-520-25120-5 .
- ^ Биби, Ф. (август 2013 г.). «Мультикалиброванная митохондриальная филогения современных Bovidae (Artiodactyla, Ruminantia) и важность летописи окаменелостей для систематики» . Эволюционная биология BMC . 13 : 166. дои : 10.1186/1471-2148-13-16 . ПМК 3607928 . ПМИД 23339550 .
- ^ Гераадс, Д.; Эль Бугаби, С.; Зухри, С. (2012). «Новый козий бык (Mammalia) из позднего миоцена Марокко». Palaeontologica Africana (47): 19–24. ISSN 0078-8554 .
- ^ Кингдон, Дж. (1989). Млекопитающие Восточной Африки: Атлас эволюции Африки (Том III, Часть C) . Чикаго: Издательство Чикагского университета. стр. 1–33. ISBN 978-0-226-43724-8 .
- ^ Стэнли, С.М.; Элдридж, Н. (1984). «Эволюционная модель и процесс в сестринской группе Alcelaphini-Aepycerotini (Mammalia: Bovidae)». Живые ископаемые . Спрингер. стр. 62–79. ISBN 978-1461-382-737 .
- ^ Врба, ЕС; Беркл, Л.Х.; Партридж, Калифорния; Дентон, GH (1995). Палеоклимат и эволюция с акцентом на происхождение человека . Нью-Хейвен: Издательство Йельского университета. стр. 24–5. ISBN 978-0300-063-486 .
- ^ Перейти обратно: а б с Уолтон, Д.В. (1989). Фауна Австралии (Том 1Б) . Канберра: Публикационная служба правительства Австралии. стр. 1–14. ISBN 978-0644-060-561 .
- ^ Хасанин, А. (2015). «Систематика и филогения крупного рогатого скота» . В Гаррике, Д.; Рувинский А. (ред.). Генетика крупного рогатого скота (второе изд.). Оксфордшир, Бостон: Каби. стр. 1–18. ISBN 978-1-78064-221-5 . Архивировано из оригинала 14 марта 2023 г. Проверено 13 января 2021 г.
- ^ Хаффман, Б. «Королевская антилопа» . Окончательное копытное . Архивировано из оригинала 16 декабря 2014 года . Проверено 8 октября 2014 г.
- ^ Хилдьярд, А. (2001). Находящаяся под угрозой исчезновения дикая природа и растения мира . Нью-Йорк: Маршалл Кавендиш. стр. 769–70. ISBN 978-0-7614-7200-1 .
- ^ "Oryx leucoryx" в Энциклопедии жизни.
- ^ Перейти обратно: а б Лундриган, Б.; Бидлингмейер, Дж. (2000). « Connochaetes gnou : черный гну» . Сеть разнообразия животных . Мичиганский университет. Архивировано из оригинала 5 октября 2013 г. Проверено 21 августа 2013 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л Эстес, Р.Д. (2004). Руководство по поведению африканских млекопитающих: включая копытных, хищников и приматов (4-е изд.). Беркли: Издательство Калифорнийского университета. стр. 7–25 . ISBN 978-0-520-08085-0 .
- ^ Краусман, PR; Кейси, AL (2012). « Адакс назомакулатус ». Виды млекопитающих (807): 1–4. дои : 10.1644/807.1 . S2CID 198123785 .
- ^ Перейти обратно: а б Каро, Тим (2009). «Контрастная окраска наземных млекопитающих» . Философские труды Королевского общества Б. 364 (1516): 537–548. дои : 10.1098/rstb.2008.0221 . ПМК 2674080 . ПМИД 18990666 .
- ^ Котт, HB (1940). Адаптивная окраска животных . Лондон: Метуэн. стр. 88 и фото 25.
- ^ Килти, РА (январь 1998 г.). «Контршейдинг: универсально обманчиво или обманчиво универсально?». Тенденции в экологии и эволюции . 3 (1): 21–23. дои : 10.1016/0169-5347(88)90079-1 . ПМИД 21227055 .
- ^ Котт, HB (1940). Адаптивная окраска животных . Лондон: Метуэн. п. 53 .
- ^ Лесли, DM; Шарма, К. (25 сентября 2009 г.). «Tetracerus Quadricornis (Парнокопытные: Bovidae)» . Виды млекопитающих (843): 1–11. дои : 10.1644/843.1 .
- ^ ЕС, Мунгалл (2007). Полевой справочник по экзотическим животным: Неместные копытные млекопитающие в США (1-е изд.). Колледж-Стейшн: Издательство Техасского университета A&M. п. 197. ИСБН 978-1-58544-555-4 .
- ^ Американское общество по охране пород домашнего скота (2009 г.). «Овечка Джейкоб» . Питтсборо, Северная Каролина: Организация по охране американских пород домашнего скота. Архивировано из оригинала 10 августа 2011 г. Проверено 5 мая 2011 г.
- ^ Биби, Ф.; Бухсианидзе, М.; Джентри, А.; Гераадс, Д.; Костопулос, Д.; Врба, Э. (2009). «Летопись окаменелостей и эволюция Bovidae: состояние поля» . Электронная палеонтология . 12 (3):10А. Архивировано из оригинала 16 июня 2011 г. Проверено 14 ноября 2010 г.
- ^ Гейтси, Дж.; Йелон, Д.; ДеСалле, Р.; Врба, Э. (1992). «Филогения Bovidae (Artiodactyla, Mammalia) на основе последовательностей митохондриальной рибосомальной ДНК» . Молекулярная биология и эволюция . 9 (3): 433–446. doi : 10.1093/oxfordjournals.molbev.a040734 . ПМИД 1584013 .
- ^ Лундриган, Б. (1996). «Морфология рогов и боевое поведение семейства Bovidae» . Журнал маммологии . 77 (2): 462–75. дои : 10.2307/1382822 . JSTOR 1382822 .
- ^ Каро, ТМ; Грэм, CM; Стоунер, CJ; Флорес, ММ (2003). «Корреляты формы рогов и рогов у быков и оленьих». Поведенческая экология и социобиология . 55 (1): 32–41. дои : 10.1007/s00265-003-0672-6 . S2CID 12030618 .
- ^ Перейти обратно: а б Бро-Йоргенсен, Дж. (2007). «Интенсивность полового отбора предсказывает размер оружия самцов крупного рогатого скота». Эволюция . 61 (6): 1316–1326. дои : 10.1111/j.1558-5646.2007.00111.x . ПМИД 17542842 . S2CID 24278541 .
- ^ Эзенва, В. ; Джоллес, А. (июнь 2008 г.). «Рога честно сигнализируют о заражении паразитами самцов и самок африканского буйвола». Поведение животных . 75 (6): 2013–2021. дои : 10.1016/J.ANBEHAV.2007.12.013 . ISSN 0003-3472 . S2CID 49240459 . Викиданные Q55899859 .
- ^ Перейти обратно: а б Станкович, Т.; Каро, Т. (2009). «Эволюция вооружения самок крупного рогатого скота» . Труды Королевского общества B: Биологические науки . 276 (1677): 4329–34. дои : 10.1098/rspb.2009.1256 . ПМК 2817105 . ПМИД 19759035 .
- ^ Пакер, К. (1983). «Половой диморфизм: рога африканских антилоп». Наука . 221 (4616): 1191–3. Бибкод : 1983Sci...221.1191P . дои : 10.1126/science.221.4616.1191 . ПМИД 17811523 . S2CID 37000507 .
- ^ Дженис, К.; Джарман, П. (1984). Макдональд, Д. (ред.). Энциклопедия млекопитающих . Нью-Йорк: факты в архиве. стр. 498–9 . ISBN 978-0-87196-871-5 .
- ^ Вятт, Т.Д. (2003). Феромоны и поведение животных: общение посредством запаха и вкуса . Кембридж: Издательство Кембриджского университета. п. 97 . ISBN 978-0-521-48526-5 .
- ^ Чишек, Д. «Бушбак» . Сеть разнообразия животных . Зоологический музей Мичиганского университета. Архивировано из оригинала 28 июля 2013 года . Проверено 28 октября 2014 г.
- ^ ТЛ, Ньюэлл. «Вотербак» . Сеть разнообразия животных . Зоологический музей Мичиганского университета. Архивировано из оригинала 28 сентября 2012 года . Проверено 28 октября 2014 г.
- ^ Лотт, Дейл Ф. (1991). Внутривидовая изменчивость социальных систем диких позвоночных . Издательство Кембриджского университета. стр. 37 . ISBN 978-0-521-37024-0 .
- ^ Бигалке, Р.К. (1972). «Наблюдения за поведением и питанием спрингбока Antidorcas marsupialis » . Зоология Африканская . 7 (1): 333–359. дои : 10.1080/00445096.1972.11447448 . hdl : 10499/AJ19390 .
- ^ Перейти обратно: а б Паркер, СП (1990). Энциклопедия млекопитающих Гржимека (том 5) (1-е изд.). Нью-Йорк: Издательство McGraw-Hill. стр. 288–324, 338–9, 354–5, 432–3, 444–5, 460–1, 482–3 . ISBN 978-0-07-909508-4 .
- ^ Чаплевски, Нью-Джерси; Райан, Дж. М.; Воган, Т.А. (2011). Маммология (5-е изд.). Садбери: Издательство Jones and Bartlett. ISBN 978-0-7637-6299-5 .
- ^ Пост, Э.; Форчхаммер, MC (июль 2008 г.). «Изменение климата снижает репродуктивный успех арктических травоядных из-за трофического несоответствия» . Философские труды Королевского общества B: Биологические науки . 363 (1501): 2367–2373. дои : 10.1098/rstb.2007.2207 . ПМК 2606787 . ПМИД 18006410 .
- ^ Ганьон, М.; Чу, А.Е. (май 2000 г.). «Диетические предпочтения современных африканских Bovidae» . Журнал маммологии . 81 (2): 490–511. doi : 10.1644/1545-1542(2000)081<0490:DPIEAB>2.0.CO;2 .
- ^ Спонхаймер, М.; Ли-Торп, Дж.А.; ДеРюитер, диджей; Смит, Дж. М.; ван дер Мерве, Нью-Джерси; Рид, К.; Грант, CC; Эйлифф, ЛК; Робинсон, Т.Ф. (2003). «Рацион южноафриканских коров: данные о стабильных изотопах» . Журнал маммологии . 84 (2): 471–9. doi : 10.1644/1545-1542(2003)084<0471:DOSABS>2.0.CO;2 .
- ^ Перейти обратно: а б с д и Фельдхамер, Джордж А.; Дрикамер, Ли С.; Весси, Стивен Х.; Мерритт, Джозеф Ф.; Краевски, Кэри (2007). Маммология: адаптация, разнообразие, экология . Издательство Университета Джонса Хопкинса. стр. 519–522. ISBN 978-0-8018-8695-9 . Архивировано из оригинала 14 марта 2023 г. Проверено 7 ноября 2016 г.
- ^ Перейти обратно: а б Кребс, младший; Дэвис, НБ (1997). Поведенческая экология: эволюционный подход (4-е изд.). Уайли-Блэквелл. ISBN 978-0-86542-731-0 .
- ^ Перейти обратно: а б Эстес, Р.Д. (2004). Руководство по поведению африканских млекопитающих: включая копытных, хищников и приматов (4-е изд.). Беркли: Издательство Калифорнийского университета. стр. 158–66 . ISBN 978-0-520-08085-0 .
- ^ Грей, Гэри Г.; Симпсон, К. Дэвид (1980). «Аммотрагус лервия». Виды млекопитающих (144): 1–7. дои : 10.2307/3504009 . JSTOR 3504009 .
- ^ Роза, HJD; Брайант, MJ (2003). «Сезонность размножения овец». Исследования мелких жвачных животных . 48 (3): 155–71. дои : 10.1016/S0921-4488(03)00038-5 .
- ^ Шемино, П.; Пеллетье, Дж.; Герен, Ю.; Колас, Г.; Раво, Япония; Туре, Г.; Алмейда, Г.; Тимонье, Ж.; Ортавант, Р. (1988). «Фотопериодическое лечение и лечение мелатонином для контроля сезонного размножения овец и коз» . Размножение, питание, развитие . 28 (2Б): 409–22. дои : 10.1051/rnd:19880307 . ПМИД 3413339 .
- ^ Прендергаст, Би Джей; Мосингер, Б.; Колаттукуди, ЧП; Нельсон, Р.Дж. (2002). «Экспрессия гипоталамических генов у репродуктивно фотореактивных и фоторефрактерных сибирских хомяков» . Труды Национальной академии наук . 99 (25): 16291–6. Бибкод : 2002PNAS...9916291P . дои : 10.1073/pnas.232490799 . ПМК 138604 . ПМИД 12456888 .
- ^ Оуэн-Смит, Н. (1993). «Сравнительный уровень смертности самцов и самок куду: цена полового диморфизма размеров». Журнал экологии животных . 62 (3): 428–40. дои : 10.2307/5192 . JSTOR 5192 . S2CID 87280870 .
- ^ Тойго, К.; Гайяр, JM (2003). «Причины выживания взрослых особей копытных с учетом пола: диморфизм полового размера, тактика спаривания или суровость окружающей среды?». Ойкос . 101 (2): 376–84. дои : 10.1034/j.1600-0706.2003.12073.x .
- ^ Эстес, Р.Д. (1991). «Значение рогов и других мужских вторичных половых признаков у самок крупного рогатого скота». Прикладная наука о поведении животных . 29 (1–4): 403–51. дои : 10.1016/0168-1591(91)90264-X .
- ^ Зедер, Массачусетс (2006). Документирование одомашнивания: новые генетические и археологические парадигмы . Беркли, Калифорния: Издательство Калифорнийского университета. п. 317. ИСБН 978-0-520-24638-6 .
- ^ Фелан, Бенджамин; Фелан, Бенджамин (24 июля 2013 г.). «Чужое молоко» . Слейт.com. Архивировано из оригинала 6 октября 2014 года . Проверено 10 октября 2014 г.
- ^ Хьюз, Том; Хьюз, Мередит Сэйлс (2005). Гастрономия!: Музеи еды и объекты культурного наследия Франции . Издательство Банкер Хилл. п. 19. ISBN 978-1-59373-029-1 .
- ^ «Говядина нежирная органическая» . WHFoods. 18 октября 2004 г. Архивировано из оригинала 2 апреля 2015 г. Проверено 1 апреля 2015 г.
- ^ «| Национальная ассоциация зубров» . Bisoncentral.com. Архивировано из оригинала 20 января 2011 года . Проверено 1 апреля 2015 г.
- ^ Вельдмейер, А.Дж.; Харрис, С. (2014). Почему кожа?: Материальные и культурные аспекты кожи . Сайдстоун Пресс. стр. 31–6. ISBN 978-90-8890-261-1 .
- ^ «Овцы-мериносы в Австралии» . Архивировано из оригинала 5 ноября 2006 г. Проверено 1 апреля 2015 г.
- ^ Колай, АК (2007). Навоз и удобрения . Нью-Дели: Atlantic Publications. п. 98. ИСБН 978-81-269-0810-3 .
- ^ Эвисон, Вера И. (1975). «Германские стеклянные рога для питья». Журнал исследований стекла . 17 : 74–87. ISSN 0075-4250 . JSTOR 24188063 .
- ^ «Басни Эзопа» . Басни Эзопа . Архивировано из оригинала 16 октября 2014 года . Проверено 10 октября 2014 г.
- ^ Пек. «Вступление: Химера» . Архивировано из оригинала 11 октября 2022 года . Проверено 31 марта 2015 г.
- ^ Эберхард, В. (1986). Словарь китайских символов: скрытые символы в китайской жизни и мысли . Лондон: Рутледж. ISBN 978-0-415-00228-8 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Bovidae, на Викискладе?
СМИ, связанные с Bovidae, на Викискладе?  Данные, относящиеся к Bovidae, в Wikispecies
Данные, относящиеся к Bovidae, в Wikispecies - . Британская энциклопедия (11-е изд.). 1911.
- . Новая энциклопедия Кольера . 1921.