Филогенетическая номенклатура
Филогенетическая номенклатура — это метод номенклатуры таксонов . в биологии, в котором используются филогенетические определения названий таксонов, как описано ниже Это контрастирует с традиционным методом , при котором названия таксонов определяются типом , которым может быть экземпляр или таксон более низкого ранга , и описанием в словах. [ 1 ] Филогенетическая номенклатура в настоящее время регулируется Международным кодексом филогенетической номенклатуры ( PhyloCode ).
Определения
[ редактировать ]Филогенетическая номенклатура связывает имена с кладами , группами, состоящими из предка и всех его потомков. Такие группы называются монофилетическими . Существуют несколько разные методы указания предка, которые обсуждаются ниже. После указания предка значение названия фиксируется: предок и все организмы, являющиеся его потомками, включаются в названный таксон. Для перечисления всех этих организмов (т.е. предоставления полного описания ) необходимо знать полное филогенетическое древо. На практике почти всегда существует одна или несколько гипотез относительно правильного соотношения. Различные гипотезы приводят к тому, что разные организмы считаются включенными в названный таксон, но не влияют на то, к каким организмам это название действительно относится; таким образом, название не зависит от пересмотра теории. [ нужна ссылка ]
Филогенетические определения названий клад
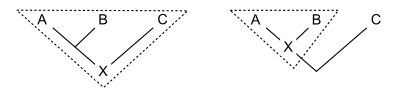
[ редактировать ]Филогенетическая номенклатура присваивает названия кладам , группам, состоящим исключительно из предка и всех его потомков. Таким образом, все, что необходимо для определения клады, — это указать предка. Есть несколько способов сделать это. Обычно предок указывается по его отношению к двум или более спецификаторам (видам, экземплярам или признакам), которые упоминаются явно. На схеме показаны три распространенных способа сделать это. Для ранее определенных клад A, B и C клада X может быть определена как:
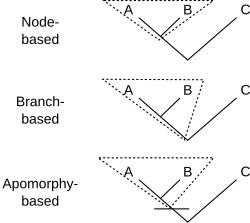
- может Определение на основе узла гласить: « последний общий предок A и B и все потомки этого предка». Таким образом, вся линия ниже места соединения А и В не принадлежит той кладе, к которой относится название с этим определением. Кронная группа — это тип группы, основанной на узлах, где A и B — существующие (живые) таксоны.
- Пример: динозавры- зауроподы состоят из последнего общего предка вулканодона (А) и апатозавра (Б). [ 2 ] и все потомки этого предка. Этот предок был первым зауроподом. C может включать других динозавров, таких как Стегозавр .
- Определение на основе ветвей , часто называемое определением на основе основы , может гласить: « первый предок A, который не является также предком C, и все потомки этого предка». Таким образом, вся линия ниже соединения A и B (кроме самой нижней точки) действительно принадлежит кладе, к которой относится название с этим определением. Пан-группа или общая группа - это тип ветвящейся группы, где A и C являются современными (живыми) таксонами.
- Пример (также общая группа): Грызуны состоят из первого предка домовой мыши (А), который не является также предком восточного кролика (С), вместе со всеми потомками этого предка. Здесь предком А (но не С) является самый первый грызун. Б — какой-то другой потомок того первого грызуна, возможно, рыжая белка .
- Определение , основанное на апоморфии, может гласить: «первый предок А, обладающий признаком М, унаследованным А, и все потомки этого предка». На диаграмме M развивается в месте пересечения горизонтальной линии с деревом. Таким образом, клада, к которой относится имя с этим определением, содержит ту часть линии ниже последнего общего предка А и В, которая соответствует предкам, обладающим апоморфией М. Нижняя часть линии исключается. Не обязательно, чтобы B имел признак M; возможно, он исчез в линии, ведущей к B.
- Пример: четвероногие состоят из первого предка человека (А), от которого люди унаследовали конечности с пальцами рук и ног (М), и всех потомков этого предка. К этим потомкам относятся змеи (Б), не имеющие конечностей.
Несколько других альтернатив представлены в PhyloCode . [ 3 ] (см. ниже ), хотя мы не претендуем на исчерпывающий характер.
Филогенетическая номенклатура допускает использование не только родственных связей , но и свойства дошедшего до нас . один из многих методов определения неорнитов Например, (современных птиц):
- Неорниты состоят из последнего общего предка ныне живущих членов наиболее инклюзивной клады, включающей какаду Cacatua galerita , но не динозавра Stegosaurus Armatus , а также всех потомков этого предка.
Neornithes — это коронная клада , клада, для которой последний общий предок ее ныне живущих членов также является последним общим предком всех ее членов.
Имена узлов
[ редактировать ]- Коронный узел : Самый недавний общий предок выбранных видов интересующей клады.
- Стеблевой узел : самый недавний общий предок интересующей клады и ее родственной клады.
Определения названий парафилетических и полифилетических таксонов на основе происхождения
[ редактировать ]Для Филокода «филогенетическое определение» может получить только клада, и это ограничение соблюдается в настоящей статье. Однако также возможно создать определения названий других групп, которые являются филогенетическими в том смысле, что они используют только наследственные отношения, основанные на видах или образцах. [ 4 ] Например, если предположить, что Mammalia и Aves (птицы) определены таким образом, Amniotes можно определить как «самого недавнего общего предка Mammalia и Aves и всех его потомков, кроме Mammalia и Aves». Это пример парафилетической группы, клады минус одна или несколько подчиненных клад. Названия полифилетических групп, характеризующихся признаком, который развивался конвергентно в двух или более подгруппах, могут быть определены аналогичным образом как сумма нескольких клад. [ 4 ]
Звания
[ редактировать ]Используя традиционные номенклатурные коды , такие как Международный кодекс зоологической номенклатуры и Международный кодекс номенклатуры водорослей, грибов и растений , таксоны, которые не связаны явно с рангом , не могут быть названы формально, поскольку применение названия к таксон основан как на типе, так и на ранге. Так, например, «семейство» Hominidae использует в качестве своего типа род Homo; его ранг (семейство) обозначается суффиксом -idae (см. обсуждение ниже). Требование ранга является основным различием между традиционной и филогенетической номенклатурой. Это имеет несколько последствий: оно ограничивает количество вложенных уровней, на которых можно применять имена; он вызывает изменение окончаний имен, если у группы изменился ее ранг, даже если в ней точно такие же члены (т. е. тот же самый объем ); и это логически несовместимо с тем, что все таксоны являются монофилетическими. [ нужна ссылка ]
В действующих кодексах есть правила, гласящие, что имена должны иметь определенные окончания в зависимости от ранга таксонов, к которым они применяются. Если группа имеет разный ранг в разных классификациях, ее название должно иметь другой суффикс. Эрешефский (1997:512) [ 5 ] привел пример. Он отметил, что Симпсон в 1963 году и Уайли в 1981 году согласились, что одну и ту же группу родов, включающую род Homo , следует объединять в таксон. Симпсон рассматривал этот таксон как семейство и поэтому дал ему название «Hominidae»: «Homin-» от «Homo» и «-idae» как суффикс семейства, используя зоологический код. Уайли считал, что оно относится к рангу «племени», и поэтому дал ему имя «Гоминини», где «-ini» было суффиксом племени. Племя Уайли Hominini составляло лишь часть семейства, которое он назвал Hominidae. Таким образом, используя зоологический код, две группы с совершенно одинаковым объёмом получили разные названия (Hominidae Симпсона и Hominini Уайли), а две группы с одинаковым названием имели разные объёмы (Hominidae Симпсона и Hominidae Уайли).
Особенно в последние десятилетия (благодаря достижениям в области филогенетики ) систематики назвали множество «вложенных» таксонов (т. е. таксонов, которые содержатся внутри других таксонов). Ни одна система номенклатуры не пытается назвать каждую кладу; это было бы особенно сложно с традиционной номенклатурой, поскольку каждому названному таксону должен быть присвоен более низкий ранг, чем любому названному таксону, в который он вложен, поэтому количество названий, которые могут быть присвоены во вложенном наборе таксонов, не может быть больше, чем число общепризнанных званий. Готье и др. (1988) [ 6 ] предположил, что если Reptilia присвоен традиционный ранг «класса», то филогенетическая классификация должна присвоить Aves ранг рода. [ 5 ] В такой классификации все ~ 12 000 известных видов современных и вымерших птиц должны были бы быть включены в этот род.
Были предложены различные решения при сохранении кодов ранговой номенклатуры. Паттерсон и Розен (1977) [ 7 ] предложили девять новых рангов между семейством и надсемейством, чтобы иметь возможность классифицировать кладу сельди, а Маккенна и Белл (1997) [ 8 ] введено большое количество новых рангов, чтобы справиться с разнообразием млекопитающих; они не получили широкого распространения. Для ботаники группа покрытосеменных филогении , ответственная за наиболее широко используемую в настоящее время классификацию цветковых растений , выбрала другой метод. Они сохранили традиционные ранги семейства и отряда, считая их ценными для преподавания и изучения взаимоотношений между таксонами, но также ввели именованные клады без формальных рангов. [ 9 ]
В филогенетической номенклатуре ранги не влияют на написание названий таксонов (см., например, Готье (1994). [ 10 ] и Филокод ). Однако ранги не полностью запрещены для филогенетической номенклатуры. Они просто отделены от номенклатуры: они не влияют на то, какие названия можно использовать, какие таксоны с какими названиями связаны и какие имена могут относиться к вложенным таксонам. [ 11 ] [ 12 ] [ 13 ]
Принципы традиционной ранговой номенклатуры логически несовместимы с тем, что все таксоны строго монофилетичны. [ 11 ] [ 14 ] Например, каждый организм должен принадлежать к какому-то роду , поэтому для каждого общего предка млекопитающих и птиц должен существовать род. Чтобы такой род был монофилетическим, он должен включать как класс Mammalia, так и класс Aves. Однако в ранговой номенклатуре классы должны включать роды, а не наоборот. [ нужна ссылка ]
Философия
[ редактировать ]Конфликт между филогенетической и традиционной номенклатурой представляет собой разные взгляды на метафизику и эпистемологию таксонов. Для сторонников филогенетической номенклатуры таксон — это отдельная сущность, сущность, которая может приобретать или терять атрибуты с течением времени. [ 15 ] Точно так же, как человек не становится кем-то другим, когда его или ее свойства меняются в результате взросления, старения или более радикальных изменений, таких как амнезия, потеря конечности или смена пола, так и таксон остается тем же самым, какие бы характеристики ни приобретались. или потерян. [ 16 ] Учитывая метафизические утверждения сторонников филогенетической номенклатуры относительно ненаблюдаемых сущностей, критики назвали их метод эссенциализмом происхождения. [ 17 ] [ 18 ]
Для любого индивида должно быть что-то, что связывает его временные стадии друг с другом, благодаря чему он остается одной и той же сущностью. Для человека пространственно-временная непрерывность тела обеспечивает соответствующую понятийную непрерывность; от младенчества до старости тело прокладывает непрерывный путь по миру и именно эта непрерывность, а не какие-либо особенности личности, связывает малыша с восьмидесятилетним человеком. [ 19 ] Это похоже на известную философскую проблему «Корабль Тесея» . Для таксона ЕСЛИ характеристики не имеют значения, ТО только предковые отношения могут связывать девонскую Rhyniognatha hirsti с современной бабочкой-монархом как представителей, разделенных 400 миллионами лет, таксона Insecta. [ 16 ] Противоположное мнение ставит под сомнение предпосылку этого силлогизма и утверждает, с эпистемологической точки зрения, что члены таксонов распознаются только эмпирически на основе их наблюдаемых характеристик, а гипотезы об общем происхождении являются результатами теоретической систематики, а не априорными предпосылками. Если нет характеристик, позволяющих ученым признать принадлежность ископаемого к таксономической группе, то это просто не поддающийся классификации кусок породы. [ 20 ]
Если происхождение достаточно для непрерывности таксона, то все потомки члена таксона также будут включены в таксон, поэтому все добросовестные таксоны являются монофилетическими; названия парафилетических групп не заслуживают формального признания. Поскольку « Pelycosauria » относится к парафилетической группе, включающей некоторых пермских четвероногих, но не их нынешних потомков, оно не может быть признано действительным названием таксона. Опять же, не соглашаясь с мнением о том, что следует называть только монофилетические группы, систематики-эмпирики противостоят этому эссенциализму происхождения, указывая, что пеликозавры признаны парафилетическими именно потому, что они демонстрируют комбинацию синапоморфий и симплезиоморфий, указывающих на то, что некоторые из них более тесно связаны между собой. для млекопитающих, чем для других пеликозавров. Материальное существование комплекса окаменелостей и его статус клады — это не одно и то же. Монофилетические группы заслуживают внимания и названий, потому что они обладают общими интересными свойствами — синапоморфиями — которые являются свидетельством, позволяющим сделать вывод об общем происхождении. [ 21 ]
История
[ редактировать ]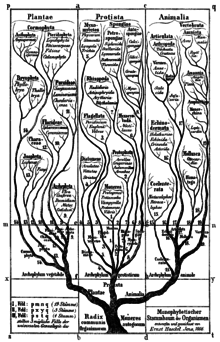
Филогенетическая номенклатура — это семантическое расширение общепринятой идеи ветвления в ходе эволюции, представленной в диаграммах Жана-Батиста Ламарка и более поздних авторов, таких как Чарльз Дарвин и Эрнст Геккель . [ 23 ] [ 24 ] В 1866 году Геккель впервые построил единую реляционную диаграмму всего живого на основе существующей принятой в то время классификации жизни. Эта классификация была ранговой, но не содержала таксонов, которые Геккель считал полифилетическими . В нем Геккель ввел ранг типа , который в своем названии несет в себе оттенок монофилии (буквально означающий «ствол»). [ нужна ссылка ]
С тех пор ведутся споры о том, каким образом и в какой степени понимание филогении жизни должно использоваться в качестве основы для ее классификации, причем мнения включают «числовую таксономию» ( фенетику ), « эволюционную таксономию » (градистику), и «филогенетическая систематика». Начиная с 1960-х годов время от времени предлагались безранговые классификации, но в целом принципы и общий язык традиционной номенклатуры использовались всеми тремя школами мысли. [ нужна ссылка ]
Однако большинство основных принципов филогенетической номенклатуры (отсутствие обязательных рангов и нечто близкое к филогенетическим определениям) можно проследить до 1916 года, когда Эдвин Гудрич [ 25 ] интерпретировал название Sauropsida , определенное 40 годами ранее Томасом Генри Хаксли , как включающее птиц ( Aves ), а также часть Reptilia , и изобрел новое название Theropsida, чтобы включить млекопитающих, а также другую часть Reptilia. Поскольку эти таксоны были отделены от традиционной зоологической номенклатуры, Гудрич не подчеркивал ранги, но ясно обсуждал диагностические признаки, необходимые для распознавания и классификации окаменелостей, принадлежащих к различным группам. Например, что касается пятой плюсневой кости задней ноги, он сказал: «Факты подтверждают нашу точку зрения, поскольку у этих ранних рептилий были нормальные плюсневые кости, как и у их предков-амфибий. Помогите нам решить, принадлежит ли данный вид к линии эволюции Теропсидов или Зауропсидов». Гудрич заключил свою статью: «Обладание этими признаками показывает, что все ныне живущие рептилии принадлежат к группе Sauropsidan, в то время как строение стопы позволяет нам определить родство многих не полностью известных ископаемых родов и заключить, что только некоторые вымершие отряды могут принадлежат к ветви Теропсидана». Гудрич полагал, что от названия Reptilia следует отказаться, как только филогения рептилий станет более известной. [ нужна ссылка ]
Принцип, согласно которому формально следует называть только клады , стал популярен среди некоторых исследователей во второй половине ХХ века. Она распространилась вместе с методами открытия клад ( кладистикой ) и является составной частью филогенетической систематики (см. выше). В то же время стало очевидно, что обязательные звания, являющиеся частью традиционных систем номенклатуры, создают проблемы. Некоторые авторы предлагали вообще отказаться от них, начиная с Вилли Хеннига. отказа [ 26 ] из его более раннего предложения определить ранги как классы геологического возраста. [ 27 ] [ 28 ]
Первое использование филогенетической номенклатуры в публикации можно датировать 1986 годом. [ 29 ] Вскоре последовали теоретические статьи, излагающие принципы филогенетической номенклатуры, а также дальнейшие публикации, содержащие приложения филогенетической номенклатуры (в основном к позвоночным животным) (см. Раздел «Литература»).
Пытаясь избежать раскола в сообществе систематиков, « Готье предложил двум членам МКЗН применять формальные таксономические названия, регулируемые зоологическим кодексом, только к кладам (по крайней мере, для надвидовых таксонов) и отказаться от линнеевских рангов, но эти два члены сразу отвергли эти идеи». [ 30 ] Посылка названий в традиционной номенклатуре базируется, в конечном счете, на типовых экземплярах, а определение групп рассматривается как таксономический выбор, сделанный систематиками, работающими над отдельными группами, а не номенклатурное решение, принятое на основе априорных правил Кодексов о Номенклатура . [ 31 ] Желание включить таксономические ограничения в номенклатурные определения побудило Кевина де Кейруша и ботаника Филиппа Кантино начать разработку собственного кодекса номенклатуры, PhyloCode , для регулирования филогенетической номенклатуры. [ нужна ссылка ]
Споры
[ редактировать ]Новаторская работа Вилли Хеннига вызвала споры. [ 32 ] об относительных преимуществах филогенетической номенклатуры по сравнению с таксономией Линнея или родственным методом эволюционной таксономии , который существует и по сей день. [ 33 ] Некоторые споры, которыми занимались кладисты, происходили с XIX века. [ 34 ] Хотя Хенниг настаивал на том, что разные схемы классификации полезны для разных целей, [ 35 ] он отдавал предпочтение своим собственным, утверждая, что категории его системы обладают «индивидуальностью и реальностью» в отличие от «вневременных абстракций» классификаций, основанных на морфологии. [ 36 ]
Говорят, что формальные классификации, основанные на кладистических рассуждениях, подчеркивают происхождение за счет описательных характеристик. Тем не менее, большинство систематиков в настоящее время избегают парафилетических групп всякий раз, когда считают, что это возможно в рамках таксономии Линнея; полифилетические таксоны уже давно вышли из моды. Многие кладисты утверждают, что традиционные Кодексы зоологической и ботанической номенклатуры полностью совместимы с кладистическими методами и что нет необходимости заново изобретать систему названий, которая хорошо функционировала на протяжении 250 лет. [ 37 ] [ 38 ] [ 39 ] но другие утверждают, что эта система не так эффективна, как должна быть, и что пришло время принять номенклатурные принципы, которые представляют дивергентную эволюцию как механизм, объясняющий большую часть известного биоразнообразия. [ 40 ] [ 41 ]
Международный кодекс филогенетической номенклатуры
[ редактировать ]ICPN . или PhyloCode — это свод правил и рекомендаций по филогенетической номенклатуре
- ICPN названия регулирует только клад . Названия видов основаны на правилах традиционных кодов номенклатуры .
- Принцип приоритета (или «первоочередности») применяется к именам и определениям в рамках ICPN. Отправной точкой приоритета стало 30 апреля 2020 года.
- Определения существующих названий, а также новые имена вместе с их определениями должны быть опубликованы в рецензируемых работах (не позднее даты начала) и должны быть зарегистрированы в онлайн-базе данных, чтобы быть действительными.
Число сторонников широкого внедрения PhyloCode по -прежнему невелико, и неизвестно, насколько широко ему будут следовать.
Ссылки
[ редактировать ]- ^ Международная комиссия по зоологической номенклатуре (1999). «Глоссарий». Международный кодекс зоологической номенклатуры (4-е изд.). Международный фонд зоологической номенклатуры, Музея естественной истории. ISBN 978-0-85301-006-7 .
- ^ Бентон, Майкл Дж. (2005). Палеонтология позвоночных . Блэквелл. п. 214. ИСБН 978-0-632-05637-8 .
- ^ Кантино, Филип Д. и де Кейрос, Кевин (2010). «Статья 9. Общие требования к установлению кладовых названий» . Международный кодекс филогенетической номенклатуры . 4в. примечание 9.3.1. .
- ^ Перейти обратно: а б де Кейрос, К.; Готье, Дж. (1990). «Филогения как центральный принцип систематики: филогенетические определения названий таксонов». Систематическая зоология . 39 (4): 307–322. дои : 10.2307/2992353 . JSTOR 2992353 .
- ^ Перейти обратно: а б Эрешефский, М. (1997). «Эволюция линнеевской иерархии». Биология и философия . 12 (4): 493–519. дои : 10.1023/А:1006556627052 . S2CID 83251018 .
- ^ Готье Дж., Эстес Р. и де Кейроз К. 1988. Филогенетический анализ лепидозавроморф . Стр. 15–98 в Р. Эстесе и Г. Прегилле (ред.): Филогенетические взаимоотношения семейств ящериц: очерки, посвященные Чарльзу Л. Кэмпу . Издательство Стэнфордского университета. ISBN 978-0-8047-1435-8
- ^ Паттерсон, К. и Розен, Д. 1977 Обзор ихтиодектиформных и других мезозойских костистых рыб, а также теория и практика классификации окаменелостей. Бюллетень Американского музея естественной истории 158: 81–172.
- ^ Маккенна, MC и Белл, SK 1997. Классификация млекопитающих выше уровня вида . Издательство Колумбийского университета. ISBN 0-231-11012-X
- ^ Группа филогенеза покрытосеменных (1998). «Порядковая классификация семейств цветковых растений». Анналы ботанического сада Миссури . 85 (4): 531–553. дои : 10.2307/2992015 . JSTOR 2992015 . S2CID 82134384 .
- ^ Готье, Ж.А. (1994). «Диверсификация амниот». В ДР Протеро; Райнер Р. Шох (ред.). Основные особенности эволюции позвоночных . Палеонтологическое общество. стр. 129–159.
- ^ Перейти обратно: а б де Кейрос, К.; Готье, Дж. (1992). «Филогенетическая систематика». Анну. Преподобный Экол. Сист . 23 : 449–480. doi : 10.1146/annurev.es.23.110192.002313 .
- ^ Кантино, П.Д. (2000). «Филогенетическая номенклатура: решение некоторых проблем». Таксон . 49 (1): 85–93. дои : 10.2307/1223935 . JSTOR 1223935 .
- ^ Брайант, Х.Н.; Кантино, П.Д. (2002). «Обзор критики филогенетической номенклатуры: является ли таксономическая свобода фундаментальным вопросом?». Биол. Преподобный . 77 (1): 39–55. дои : 10.1017/S1464793101005802 . ПМИД 11911373 . S2CID 20518066 .
- ^ Казлев М.А. "Кладистическая и линнеевская системы — несовместимы или дополняют друг друга?" . palaeos.com . Архивировано из оригинала 10 июля 2017 года . Проверено 30 сентября 2012 г.
- ^ Ассис, LCS; Бригандт, И. (2009). «Гомология: виды кластеров гомеостатических свойств в систематике и эволюции» (PDF) . Эволюционная биология . 36 (2): 248–255. дои : 10.1007/s11692-009-9054-y . S2CID 363300 . [ постоянная мертвая ссылка ]
- ^ Перейти обратно: а б Роу, Тимоти (1988). «Определение, диагностика и происхождение млекопитающих» (PDF) . Журнал палеонтологии позвоночных . 8 (3): 241–264. дои : 10.1080/02724634.1988.10011708 .
- ^ Уинзор, Мэри П. (2009). «Таксономия была основой эволюции Дарвина». Таксон . 58 : 43–49. дои : 10.1002/tax.581007 .
- ^ Риппель, Оливье (2010). «Новый эссенциализм в биологии». Философия науки . 36 (5): 662–673. дои : 10.1086/656539 . S2CID 86958171 .
- ^ Виггинс, Дэвид (1967). Тождество и пространственно-временная непрерывность . Издательство Оксфордского университета. ISBN 978-0631103707 .
- ^ Брауэр, Эндрю В.З. (2016). «Древовидное мышление» . Вывод . 2 .
- ^ Хенниг 1966 , с. 93.
- ^ Геккель, EH Ph. А. 1866. Общая морфология организмов . Джордж Реймер.
- ^ Рэган, Марк А. (2009). «Деревья и сети до и после Дарвина» . Биология Директ . 4 (43): 43. дои : 10.1186/1745-6150-4-43 . ПМЦ 2793248 . ПМИД 19917100 .
- ^ Тасси, Паскаль (май 2011 г.). «Деревья до и после Дарвина: Деревья до и после Дарвина» . Журнал зоологической систематики и эволюционных исследований . 49 (2): 89–101. дои : 10.1111/j.1439-0469.2010.00585.x .
- ^ Гудрич, ES (1916). «О классификации рептилий». Труды Королевского общества Б. 89 (615): 261–276. Бибкод : 1916РСПСБ..89..261Г . дои : 10.1098/rspb.1916.0012 .
- ^ Хенниг, В. 1969. Филогения насекомых . Вальдемар Крамер.
- ^ Хенниг, В. 1950. Основы теории филогенетической систематики . Центральное издательство Германии.
- ^ Хенниг, В. (1965). «Филогенетическая систематика». Ежегодный обзор энтомологии . 10 : 97–116. doi : 10.1146/annurev.en.10.010165.000525 .
- ^ Готье, Дж. (1986). «Ящеричья монофилия и происхождение птиц». В К. Падиане (ред.). Происхождение птиц и эволюция полета . Мемуары 8 Калифорнийской академии наук. стр. 1–55.
- ^ Лаурин, М. (2008). «Великолепная изоляция биологической номенклатуры». Зоологика Скрипта . 37 (2): 223–233. дои : 10.1111/j.1463-6409.2007.00318.x . S2CID 85020798 .
- ^ Брауэр, Эндрю В.З. (2020). «Мертв по прибытии: посмертная оценка «филогенетической номенклатуры», более 20 лет спустя» . Кладистика . 37 (6): 627–637. дои : 10.1111/cla.12432 . S2CID 224927279 .
- ^ Уиллер, Квентин (2000). Концепции видов и филогенетическая теория: дискуссия . Издательство Колумбийского университета. ISBN 978-0-231-10143-1 .
- ^ Бентон, MJ (2000). «Стебли, узлы, коронные клады и списки без рангов: Линней мертв?» (PDF) . Биологические обзоры . 75 (4): 633–648. CiteSeerX 10.1.1.573.4518 . дои : 10.1111/j.1469-185X.2000.tb00055.x . ПМИД 11117201 . S2CID 17851383 . Архивировано из оригинала (PDF) 9 августа 2017 г. Проверено 26 августа 2011 г.
- ^ Халл, Дэвид (1988). Наука как процесс . Издательство Чикагского университета. стр. 232–276. ISBN 978-0-226-36051-5 .
- ^ Хенниг 1966 , с. 9.
- ^ Хенниг 1966 , стр. 81.
- ^ Никсон, Кевин С. и Джеймс М. Карпентер. «О другой «филогенетической систематике».» Кладистика 16, вып. 3 (2000): 298-318.
- ^ Шу, Рэндалл Т. «Система Линнея и ее 250-летнее существование». Ботаническое обозрение 69, вып. 1 (2003): 59.
- ^ Брауэр, Эндрю В.З. «Мертв по прибытии: посмертная оценка «филогенетической номенклатуры», более 20 лет спустя». (2020) Кладистика 36(6):627-637.
- ^ Лорен, Мишель (3 августа 2023 г.). Появление PhyloCode: продолжающаяся эволюция биологической номенклатуры . ЦРК Пресс. дои : 10.1201/9781003092827 . ISBN 978-1-003-09282-7 .
- ^ Лорен, Мишель (23 июля 2023 г.). «Филокод: логический результат тысячелетней эволюции биологической номенклатуры?» . Зоологика Скрипта . 52 (6): 543–555. дои : 10.1111/zsc.12625 . ISSN 0300-3256 .
Источники
[ редактировать ]- Хенниг, Вилли (1966). Филогенетическая систематика . Перевод Д. Дуайта Дэвиса; Райнер Цангерль. Урбана, Иллинойс: Univ. из Иллинойс Пресс. п. 9. ISBN 978-0-252-06814-0 . (переиздано в 1979 и 1999 гг.)
Дальнейшее чтение
[ редактировать ]Здесь цитируются несколько публикаций, не упомянутых в списке литературы. Исчерпывающий список публикаций по филогенетической номенклатуре можно найти на сайте Международного общества филогенетической номенклатуры .
- Брайант, Гарольд Н. (1994). «Комментарии к филогенетическому определению названий таксонов и соглашениям о наименовании коронных клад». Сист. Биол . 43 : 124–129. дои : 10.1093/sysbio/43.1.124 .
- Кантино, Филип Д.; Олмстед, Ричард Г. (2008). «Применение филогенетически определенных имен не требует, чтобы каждый спецификатор присутствовал на дереве». Сист. Биол . 57 (1): 157–160. дои : 10.1080/10635150701883873 . ПМИД 18300028 .
- де Кейрос, Кевин (1992). Филогенетические определения и таксономическая философия. Биол. Филос. 7 : 295–313.
- Готье, Жак А., Арнольд Г. Клюге и Тимоти Роу (1988). Ранняя эволюция амниоты. Страницы 103–155 в книге Майкла Дж. Бентона (ред.): Филогения и классификация четвероногих, Том 1: Земноводные, рептилии, птицы. Сист. Жопа. Спец. Том. 35А . Кларендон Пресс, Оксфорд.
- Готье, Жак, Дэвид Каннателла, Кевин де Кейроз, Арнольд Г. Клюге и Тимоти Роу (1989). Филогения четвероногих. Страницы 337–353 в книге Б. Фернхольма, К. Бремера и Х. Йорнвалля (ред.): Иерархия жизни . Elsevier Science BV (Биомедицинский отдел), Нью-Йорк.
- Гизелин, МТ (1984). «Определение», «характер» и другие двусмысленные термины». Syst. Zool . 33 (1): 104–110. doi : 10.2307/2413135 . JSTOR 2413135 .
- Кизи, Т. Майкл (2007). «Математический подход к определению названий клад с потенциальным применением для компьютерного хранения и обработки». Зоол. Скр. 36 (6): 607–621. дои : 10.1111/j.1463-6409.2007.00302.x . S2CID 83862527 .
- Лорен, Мишель (2005). Преимущества филогенетической номенклатуры перед номенклатурой Линнея. Страницы 67–97 в книге А. Минелли, Г. Орталли и Г. Санги (ред.): Названия животных . Венетоский институт наук, литературы и искусства; Венеция.
- Ли, Майкл С.Ю. (2005). «Выбор эталонных таксонов в филогенетической номенклатуре». Зоол. Скр. 34 (3): 329–331. дои : 10.1111/j.1463-6409.2005.00196.x . S2CID 86329828 .
- Роу, Тимоти (1987). «Определение и диагностика в филогенетической системе». Сист. Зоол . 36 (2): 208–211. дои : 10.2307/2413270 . JSTOR 2413270 .
- Роу, Тимоти; Готье, Жак (1992). «Происхождение, палеонтология и определение названия млекопитающих». Сист. Биол . 41 (3): 372–378. дои : 10.1093/sysbio/41.3.372 . S2CID 86132781 .
- Серено, Пол К. (1998). «Обоснование филогенетических определений с применением к таксономии динозавров более высокого уровня». Новогодний ежегодник геологии и палеонтологии - Трактаты . 210 : 41–83. дои : 10.1127/njgpa/210/1998/41 .
- Серено, Пол К. (1999). «Определения в филогенетической таксономии: критика и обоснование». Сист. Биол . 48 (2): 329–351. дои : 10.1080/106351599260328 . ПМИД 12066711 .
- Серено, Пол К. (2005). «Логические основы филогенетической систематики» . Сист. Биол . 54 (4): 595–619. дои : 10.1080/106351591007453 . ПМИД 16109704 .
- Тейлор, Майкл П. (2007). «Филогенетические определения в эпоху до появления Филокода; последствия для наименования клад в соответствии с Филокодом». ПалеоБиос . 27 : 1–6.
- Уилкинсон, Марк (2006). «Определение стабильных эталонных таксонов для филогенетической номенклатуры». Зоол. Скр. 35 : 109–112. дои : 10.1111/j.1463-6409.2005.00213.x . S2CID 85702468 .
- Висс, Арканзас; Мэн, Дж. (1996). «Применение филогенетической таксономии к плохо разрешенным кладам короны: определение Rodentia на основе модифицированных стеблей узлов». Сист. Биол . 45 (4): 559–568. дои : 10.1093/sysbio/45.4.559 .
| Соответствующие поля | ||
|---|---|---|
| Основные понятия | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||