Глаз
| Глаз | |
|---|---|
 Сложный глаз антарктического криля | |
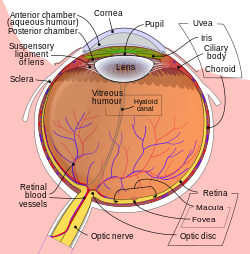 Схема человеческого глаза | |
| Подробности | |
| Система | Нервный |
| Идентификаторы | |
| латинский | окулус |
| ТА98 | А15.2.00.001 А01.1.00.007 |
| ТА2 | 113 , 6734 |
| Анатомическая терминология | |
Глаз , – орган чувств позволяющий организму воспринимать зрительную информацию. Он улавливает свет и преобразует его в электрохимические импульсы в нейронах (нейронах). организма Это часть зрительной системы .
У высших организмов глаз представляет собой сложную оптическую систему, которая собирает свет из окружающей среды, регулирует его интенсивность посредством диафрагмы , фокусирует его с помощью регулируемого набора линз для формирования изображения , преобразует это изображение в набор электрических сигналов и передает эти сигналы в мозг через нервные пути, которые соединяют глаз через зрительный нерв со зрительной корой и другими областями мозга.
Глаза с разрешающей способностью бывают десяти принципиально разных форм, которые подразделяются на сложные глаза и несложные глаза. Сложные глаза состоят из множества небольших зрительных единиц и часто встречаются у насекомых и ракообразных . Несложные глаза имеют одну линзу и фокусируют свет на сетчатке, образуя единое изображение. Этот тип глаз распространен у млекопитающих. Человеческий глаз – несложный глаз.
Простейшие глаза – ямочные. Это глазные пятна, которые можно поместить в ямку, чтобы уменьшить угол света, который попадает и воздействует на глазное пятно, чтобы позволить организму определить угол падающего света. [1]
Глаза обеспечивают несколько функций фотореакции, независимых от зрения. В организме с более сложными глазами светочувствительные ганглиозные клетки сетчатки посылают сигналы по ретиногипоталамическому тракту к супрахиазматическим ядрам для осуществления циркадной регулировки и в претектальную область для контроля зрачкового светового рефлекса .
Обзор

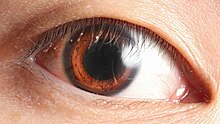
Сложные глаза различают формы и цвета . Зрительные поля многих организмов, особенно хищников, задействуют большие области бинокулярного зрения для восприятия глубины . У других организмов, особенно животных-жертв, глаза расположены так, чтобы максимально увеличить поле зрения, например, у кроликов и лошадей , которые обладают монокулярным зрением .
Первые протоглаза появились у животных 600 миллионов лет назад, примерно во время кембрийского взрыва . [2] Последний общий предок животных обладал биохимическим набором инструментов, необходимым для зрения, а более развитые глаза развились у 96% видов животных в шести из примерно 35 видов. [а] основной тип . [1] У большинства позвоночных и некоторых моллюсков глаз позволяет свету проникать и проецироваться на светочувствительный слой клеток, известный как сетчатка . Колбочки (для контрастов при слабом освещении) в сетчатке обнаруживают и преобразуют свет в нервные сигналы , (для цвета) и палочки которые передаются в мозг через зрительный нерв для формирования зрения. Такие глаза обычно имеют сферическую форму, заполнены прозрачным гелеобразным стекловидным телом , имеют фокусирующую линзу и часто радужную оболочку . Мышцы вокруг радужной оболочки изменяют размер зрачка , регулируя количество света, попадающего в глаз. [3] и уменьшение аберраций при достаточном освещении. [4] Глаза большинства головоногих моллюсков , рыб , амфибий и змей имеют фиксированную форму линз, а фокусировка достигается путем телескопирования линзы аналогично тому, как это делается в фотоаппарате . [5]
Сложные глаза членистоногих состоят из множества простых фасеток, которые, в зависимости от анатомических деталей, могут давать либо одно пиксельное изображение, либо несколько изображений на глаз. Каждый датчик имеет собственную линзу и светочувствительную ячейку(и). В некоторых глазах имеется до 28 000 таких датчиков, расположенных шестиугольно, что обеспечивает поле зрения на 360°. Сложные глаза очень чувствительны к движению. Некоторые членистоногие, в том числе многие Strepsiptera , имеют сложные глаза, состоящие всего из нескольких фасеток, каждый из которых имеет сетчатку, способную создавать изображение. Когда каждый глаз создает разное изображение, в мозгу создается слитое изображение с высоким разрешением.

Креветка -богомол обладает самой сложной в мире системой цветового зрения. Он обладает детальным гиперспектральным цветовым зрением. [6]
Трилобиты , ныне вымершие, имели уникальные сложные глаза. Прозрачные кристаллы кальцита образовали линзы их глаз. Этим они отличаются от большинства других членистоногих, у которых мягкие глаза. Количество линз в таком глазу сильно различалось; у некоторых трилобитов была только одна линза, а у других - тысячи линз в глазу.
В отличие от сложных глаз, простые глаза имеют одну линзу. Пауки-скакуны имеют одну пару больших простых глаз с узким полем зрения , дополненную множеством глаз меньшего размера для периферического зрения . насекомых Некоторые личинки , например гусеницы , имеют тип простого глаза ( стемматы ), который обычно дает лишь грубое изображение, но (как и у личинок пилильщика ) может обладать разрешающей способностью в 4 градуса дуги, быть поляризационно-чувствительным и способным увеличивать его абсолютная чувствительность ночью в 1000 и более раз. [7] Глазки , одни из самых простых глаз, встречаются у животных, например, у улиток . У них есть светочувствительные клетки, но нет линз или других средств проецирования изображения на эти клетки. Они могут различать свет и тьму, но не более того, что позволяет им избегать прямых солнечных лучей .У организмов, обитающих вблизи глубоководных жерл , сложные глаза приспособлены видеть инфракрасный свет, излучаемый горячими жерлами, что позволяет существам избежать сварения заживо. [8]
Типы
Существует десять различных раскладок глаз. Типы глаз можно разделить на «простые глаза» с одной вогнутой фоторецепторной поверхностью и «сложные глаза», которые состоят из ряда отдельных линз, расположенных на выпуклой поверхности. [1] «Простой» не подразумевает пониженный уровень сложности или остроты. Действительно, любой тип глаз можно адаптировать практически к любому поведению и окружающей среде. Единственными ограничениями, специфичными для типов глаз, являются ограничения разрешения: физика сложных глаз не позволяет им достичь разрешения лучше 1°. Кроме того, глаза суперпозиции могут достигать большей чувствительности, чем глаза аппозиции , поэтому они лучше подходят для существ, обитающих в темноте. [1] Глаза также делятся на две группы в зависимости от клеточного строения фоторецепторов: фоторецепторные клетки либо реснитчатые (как у позвоночных), либо рабдомерные . Эти две группы не монофилетичны; также у Cnidaria есть реснитчатые клетки, [9] и некоторые брюхоногие моллюски [10] и кольчатые черви обладают обоими. [11]
У некоторых организмов есть светочувствительные клетки, которые только и делают, что определяют, светло ли окружающее пространство или темно , чего достаточно для вовлечения циркадных ритмов . Они не считаются глазами, поскольку у них недостаточно структуры, чтобы считаться органом, и они не создают изображения. [12]
Все технологические методы получения оптического изображения, которые обычно используют люди, встречаются в природе, за исключением зум-объективов и линз Френеля . [1]
Несложные глаза
Простые глаза довольно распространены повсеместно, а глаза с линзами развивались по крайней мере семь раз у позвоночных , головоногих моллюсков , кольчатых червей , ракообразных и кубозоа . [13] [ не удалось пройти проверку ]
Ямы глаза
Ямочные глаза, также известные как стеммы , представляют собой глазные пятна, которые могут быть помещены в ямку, чтобы уменьшить углы света, который попадает в глазное пятно и воздействует на него, чтобы позволить организму определить угол падающего света. [1] Эти основные формы, обнаруженные примерно у 85% типов, вероятно, были предшественниками более продвинутых типов «простых глаз». Они маленькие, содержат до примерно 100 клеток размером около 100 мкм. [1] Направленность можно улучшить, уменьшив размер апертуры, включив отражающий слой позади рецепторных клеток или заполнив яму преломляющим материалом. [1]
У гадюк появились ямки, которые функционируют как глаза, воспринимая тепловое инфракрасное излучение, в дополнение к своим глазам с оптической длиной волны, как и у других позвоночных (см. Чувствительность инфракрасного излучения у змей ). Однако ямочные органы снабжены рецепторами, весьма отличными от фоторецепторов, а именно специфическим каналом временного рецепторного потенциала (каналы TRP), называемым TRPV1 . Основное отличие состоит в том, что фоторецепторы представляют собой рецепторы, связанные с G-белком , а TRP — это ионные каналы .
Глаз сферической линзы
Разрешение ямочных глаз можно значительно улучшить, включив материал с более высоким показателем преломления , что может значительно уменьшить возникающий радиус размытия, тем самым увеличивая достижимое разрешение. в линзу [1] Самая основная форма, наблюдаемая у некоторых брюхоногих моллюсков и кольчатых червей, состоит из линзы с одним показателем преломления. Гораздо более четкое изображение можно получить, используя материалы с высоким показателем преломления, уменьшающимся к краям; это уменьшает фокусное расстояние и, таким образом, позволяет формировать четкое изображение на сетчатке. [1] Это также позволяет использовать большую диафрагму для заданной резкости изображения, позволяя большему количеству света проникать в объектив; и более плоская линза, уменьшающая сферическую аберрацию . [1] Такая неоднородная линза необходима для того, чтобы фокусное расстояние уменьшилось примерно с 4-х радиусов линзы до 2,5 радиусов. [1]
Так называемые недофокусированные линзовые глаза, обнаруженные у брюхоногих моллюсков и многощетинковых червей, имеют глаза, которые занимают промежуточное положение между чашечными глазами без линз и настоящими камерами. Также у кубомедуз есть глаза со сферической линзой, роговицей и сетчаткой, но зрение размытое. [14] [15]
Гетерогенные глаза развивались по меньшей мере девять раз: четыре и более раз у брюхоногих моллюсков , один раз у копепод , один раз у кольчатых червей , один раз у головоногих моллюсков . [1] и однажды в хитонах , имеющих линзы арагонита . [16] Ни один из современных водных организмов не обладает однородными линзами; предположительно, эволюционное давление гетерогенной линзы достаточно велико, чтобы эта стадия быстро «переросла». [1]
Этот глаз создает изображение достаточно четкое, поэтому движение глаза может вызвать значительное размытие. Чтобы свести к минимуму эффект движения глаз во время движения животного, большинство таких глаз имеют стабилизирующие глазные мышцы. [1]
Глазки насекомых несут простую линзу, но их фокус обычно лежит за сетчаткой; следовательно, они не могут сформировать четкое изображение. Глазки (глаза членистоногих ямчатого типа) размывают изображение по всей сетчатке и, следовательно, превосходно реагируют на быстрые изменения интенсивности света во всем поле зрения; эта быстрая реакция дополнительно ускоряется большими нервными пучками, которые доставляют информацию в мозг. [17] Фокусировка изображения также приведет к тому, что изображение солнца будет сфокусировано на нескольких рецепторах, что может привести к повреждению под интенсивным светом; экранирование рецепторов блокирует часть света и, таким образом, снижает их чувствительность. [17] Такая быстрая реакция привела к предположению, что глазки насекомых используются главным образом в полете, поскольку их можно использовать для обнаружения внезапных изменений направления движения (поскольку свет, особенно ультрафиолетовый свет, который поглощается растительностью, обычно исходит сверху). . [17]
Несколько линз
Некоторые морские организмы несут более одной линзы; например, у веслоногих раков Понтеллы их три. Внешняя поверхность имеет параболическую поверхность, которая противодействует эффектам сферической аберрации и позволяет формировать четкое изображение. У другого копепода, Copilia , в каждом глазу есть по две линзы, расположенные как в телескопе. [1] Такие механизмы редки и плохо изучены, но представляют собой альтернативную конструкцию.
У некоторых охотников, таких как орлы и пауки-прыгуны, наблюдаются множественные линзы, у которых есть преломляющая роговица: у них есть отрицательная линза, увеличивающая наблюдаемое изображение до 50% по сравнению с рецепторными клетками, тем самым увеличивая их оптическое разрешение. [1]
Рефракционная роговица
В глазах большинства млекопитающих , птиц , рептилий и большинства других наземных позвоночных (а также пауков и некоторых личинок насекомых) стекловидное тело имеет более высокий показатель преломления, чем воздух. [1] В целом линза не сферическая. Сферические линзы создают сферическую аберрацию. При рефракционной роговице ткань хрусталика корректируется неоднородным материалом линзы (см. Линза Люнебурга ), либо асферической формой. [1] У сплющивания линзы есть недостаток; качество зрения ухудшается вдали от основной линии фокусировки. Таким образом, животные, у которых в ходе эволюции появилось широкое поле зрения, часто имеют глаза, в которых используется неоднородная линза. [1]
Как упоминалось выше, преломляющая роговица полезна только вне воды. В воде разница в показателе преломления между стекловидным телом и окружающей водой невелика. Следовательно, существа, вернувшиеся в воду — например, пингвины и тюлени — теряют сильно изогнутую роговицу и возвращаются к зрению, основанному на хрусталике. Альтернативное решение, которого придерживаются некоторые дайверы, — это иметь очень сильно фокусирующую роговицу. [1]

Уникальной особенностью глаз большинства млекопитающих является наличие век , которые вытирают глаз и распределяют слезы по роговице, чтобы предотвратить обезвоживание. Эти веки также дополняются наличием ресниц — нескольких рядов сильно иннервируемых и чувствительных волосков, которые растут по краям век и защищают глаза от мелких частиц и мелких раздражителей, таких как насекомые.
Глаза-отражатели
Альтернативой линзам является размещение внутри глаза «зеркал» и отражение изображения для фокусировки в центральной точке. [1] Природа этих глаз означает, что если бы кто-то заглянул в зрачок глаза, он увидел бы то же самое изображение, которое видел бы организм, отраженное обратно. [1]
Многие мелкие организмы, такие как коловратки , копеподы и плоские черви , используют такие органы, но они слишком малы, чтобы создавать пригодные изображения. [1] Некоторые более крупные организмы, такие как гребешки , также используют глаза-рефлекторы. У гребешка Pecten есть глаза-рефлекторы размером до 100 миллиметров, окаймляющие край его панциря. Он обнаруживает движущиеся объекты, когда они проходят мимо последовательных линз. [1]
Существует по крайней мере одно позвоночное животное, рыба-призрак , чьи глаза оснащены отражающей оптикой для фокусировки света. Каждый из двух глаз рыбы-призрака собирает свет как сверху, так и снизу; Свет, идущий сверху, фокусируется линзой, а свет, идущий снизу, — изогнутым зеркалом, состоящим из множества слоев маленьких отражающих пластинок из гуанина кристаллов . [18]
Сложные глаза



Сложный глаз может состоять из тысяч отдельных фоторецепторных единиц или омматидий ( омматидий , единственное число). Воспринимаемое изображение представляет собой комбинацию сигналов от многочисленных омматидий (отдельных «глазных единиц»), которые расположены на выпуклой поверхности и, таким образом, направлены в несколько разных направлениях. По сравнению с простыми глазами сложные глаза обладают очень большим углом обзора и могут обнаруживать быстрое движение и, в некоторых случаях, поляризацию света . [19] Поскольку отдельные линзы настолько малы, эффекты дифракции налагают ограничение на возможное разрешение, которое можно получить (при условии, что они не функционируют как фазированные решетки ). Противостоять этому можно только увеличением размера и количества линз. Чтобы видеть с разрешением, сравнимым с разрешением наших простых глаз, людям потребуются очень большие сложные глаза, радиусом около 11 метров (36 футов). [20]
Сложные глаза делятся на две группы: аппозиционные глаза, формирующие несколько перевернутых изображений, и суперпозиционные глаза, формирующие одно вертикальное изображение. [21] Сложные глаза распространены у членистоногих, кольчатых червей и некоторых двустворчатых моллюсков. [22] Сложные глаза членистоногих растут по краям за счет добавления новых омматидий. [23]
Аппозиционные глаза
Аппозиционные глаза — наиболее распространенная форма глаз и, предположительно, предковая форма сложных глаз. Они встречаются во всех группах членистоногих , хотя, возможно, они эволюционировали более одного раза в пределах этого типа. [1] У некоторых кольчатых червей и двустворчатых моллюсков также есть аппозиционные глаза. Они также имеются у Limulus , мечехвоста, и есть предположения, что другие хелицераты развили свои простые глаза путем редукции из сложной исходной точки. [1] (По-видимому, у некоторых гусениц сложные глаза развились из простых глаз противоположным образом.)
Аппозиционные глаза работают, собирая несколько изображений, по одному от каждого глаза, и объединяя их в мозгу, при этом каждый глаз обычно вносит одну точку информации. Типичный аппозиционный глаз имеет хрусталик, фокусирующий свет с одного направления на рабдоме, тогда как свет с других направлений поглощается темной стенкой омматидия .
Суперпозиция глаз
Второй тип называется глазом суперпозиции. Глаз суперпозиции делится на три типа:
- преломляющийся,
- отражающий и
- параболическая суперпозиция
Преломляющий суперпозиционный глаз имеет зазор между хрусталиком и рабдомом и не имеет боковой стенки. Каждая линза воспринимает свет под углом к своей оси и отражает его под таким же углом на другой стороне. В результате получается изображение на половине радиуса глаза, где находятся кончики рабдомов. Этот тип сложного глаза, для которого существует минимальный размер, ниже которого не может произойти эффективная суперпозиция, [24] обычно встречается у ночных насекомых, поскольку он может создавать изображения в 1000 раз ярче, чем эквивалентные аппозиционные глаза, хотя и за счет снижения разрешения. [25] В сложном типе глаз с параболической суперпозицией, наблюдаемом у членистоногих, таких как подёнки , параболические поверхности внутренней части каждой фасетки фокусируют свет от отражателя к матрице датчиков. Длиннотелые десятиногие ракообразные, такие как креветки , креветки , раки и омары , единственные, у кого есть отражающие суперпозиционные глаза, которые также имеют прозрачную щель, но используют угловые зеркала вместо линз .
Параболическая суперпозиция
Глаза этого типа преломляют свет, а затем используют параболическое зеркало для фокусировки изображения; он сочетает в себе черты суперпозиции и аппозиции глаз. [8]
Другой
Другой вид сложных глаз, обнаруженный у самцов отряда Strepsiptera , состоит из ряда простых глаз — глаз, имеющих одно отверстие, обеспечивающее освещение всей сетчатки, формирующей изображение. Несколько таких глазков вместе образуют сложный глаз стрептокрылых, который похож на «шизохроальные» сложные глаза некоторых трилобитов . [26] Поскольку каждое ушко представляет собой простой глаз, оно создает перевернутое изображение; эти изображения объединяются в мозгу, образуя одно единое изображение. Поскольку апертура глазка больше фасеток сложного глаза, такое расположение позволяет видеть при слабом уровне освещенности. [1]
Хорошие летчики, такие как мухи или медоносные пчелы, или насекомые, ловящие добычу, такие как богомолы или стрекозы , имеют специализированные зоны омматидиев, организованные в область ямки , которая обеспечивает острое зрение. В острой зоне глаза уплощены, фасетки крупнее. Уплощение позволяет большему количеству омматидиев получать свет от пятна и, следовательно, более высокому разрешению. Черное пятно, которое можно увидеть на сложных глазах таких насекомых, которое, кажется, всегда смотрит прямо на наблюдателя, называется псевдозрачком . Это происходит потому, что омматидии , которые наблюдают «в лоб» (вдоль оптических осей ), поглощают падающий свет , а те, что по бокам, отражают его. [27]
Из перечисленных выше типов есть некоторые исключения. У некоторых насекомых есть так называемый однохрусталиковый сложный глаз, переходный тип, который представляет собой что-то среднее между типом суперпозиции многолинзового сложного глаза и однохрусталичным глазом, встречающимся у животных с простыми глазами. Еще есть -мизида креветка Dioptromysis paucispinosa . У креветки глаз преломляющего суперпозиционного типа, сзади за ним в каждом глазу имеется одна большая фасетка, в три раза превышающая в диаметре остальные в глазу, а за ней - увеличенный кристаллический конус. Это проецирует вертикальное изображение на специализированную сетчатку. Полученный глаз представляет собой смесь простого глаза и сложного глаза.
Другая версия — сложный глаз, часто называемый «псевдофасетным», как у Scutigera . [28] Глаза этого типа состоят из скопления многочисленных омматидий на каждой стороне головы, организованных таким образом, что напоминают настоящий сложный глаз.
Тело Ophiocoma wendtii , разновидности хрупкой звезды , покрыто омматидиями, превращающими всю его кожу в сложный глаз. То же самое относится и ко многим хитонам . Ножки-трубки морских ежей содержат белки-фоторецепторы, которые вместе действуют как сложный глаз; у них отсутствуют экранирующие пигменты, но они могут определять направленность света по тени, отбрасываемой их непрозрачным телом. [29]
Питательные вещества
Ресничное тело треугольной формы в горизонтальном сечении и покрыто двойным слоем мерцательного эпителия. Внутренний слой прозрачен, покрывает стекловидное тело и является продолжением нервной ткани сетчатки. Наружный слой сильно пигментирован, непрерывен с пигментным эпителием сетчатки и представляет собой клетки мышцы-расширителя.
— Стекловидное тело это прозрачная, бесцветная, студенистая масса, заполняющая пространство между хрусталиком глаза и сетчаткой, выстилающей заднюю часть глаза. [30] Его производят определенные клетки сетчатки. По составу она очень похожа на роговицу, но содержит очень мало клеток (в основном фагоциты, удаляющие нежелательный клеточный мусор в поле зрения, а также гиалоциты Балаша поверхности стекловидного тела, перерабатывающие гиалуроновую кислоту ). кровеносные сосуды, а 98–99% ее объема составляет вода (в отличие от 75% в роговице) с солями, сахарами, витрозином (разновидность коллагена), сетью волокон коллагена II типа с мукополисахаридом гиалуроновой кислотой и также широкий спектр белков в микроколичествах. Удивительно, но при таком небольшом количестве твердого вещества он крепко приковывает взгляд.
Эволюция
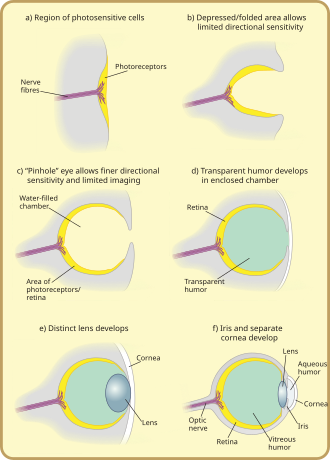
Фоторецепция филогенетически очень стара, существуют различные теории филогенеза. [31] Общее происхождение ( монофилия ) глаз всех животных в настоящее время широко признано фактом. Это основано на общих генетических особенностях всех глаз; то есть все современные глаза, какими бы разными они ни были, произошли от протоглаза, который, как полагают, развился около 650-600 миллионов лет назад, [32] [33] [34] и ген PAX6 считается ключевым фактором в этом. Считается, что на развитие большинства достижений в области ранних глаз потребовалось всего несколько миллионов лет, поскольку первый хищник, получивший истинное изображение, спровоцировал бы «гонку вооружений». [35] среди всех видов, не покинувших фотопическую среду. Без таких возможностей животные-жертвы и конкурирующие хищники окажутся в явно невыгодном положении и у них будет меньше шансов выжить и размножиться. Следовательно, параллельно развивались многочисленные типы и подтипы глаз (за исключением групп, таких как позвоночные, которые были вынуждены войти в фотопическую среду только на поздней стадии).
Глаза у различных животных демонстрируют адаптацию к их потребностям. Например, глаз хищной птицы имеет гораздо большую остроту зрения, чем глаз человека , и в некоторых случаях может улавливать ультрафиолетовое излучение. Различные формы глаз, например, у позвоночных и моллюсков являются примерами параллельной эволюции , несмотря на их отдаленное общее происхождение. Фенотипическая конвергенция геометрии глаз головоногих моллюсков и большинства позвоночных создает впечатление, что глаз позвоночных произошел от визуализирующего глаза головоногих моллюсков , но это не так, поскольку их соответствующие классы цилиарного и рабдомерного опсина поменялись местами. [36] и различные кристаллины линз показывают. [37]
Самые ранние «глаза», называемые глазными пятнами, представляли собой простые участки фоторецепторного белка у одноклеточных животных. У многоклеточных существ появились многоклеточные глазные пятна, физически похожие на участки рецепторов вкуса и запаха. Эти глазные пятна могли чувствовать только окружающую яркость: они могли различать свет и тьму, но не направление источника света. [1]
В результате постепенных изменений глазные пятна у видов, живущих в хорошо освещенной среде, приобрели неглубокую форму «чаши». Возможность слегка различать направленную яркость была достигнута за счет использования угла, под которым свет попадает на определенные клетки, для идентификации источника. Ямка со временем углублялась, отверстие уменьшалось в размерах, а количество фоторецепторных клеток увеличивалось, образуя эффективную камеру-обскуру , способную смутно различать формы. [38] Однако предки современных миксин , считающихся протопозвоночными, [36] очевидно, были вытеснены в очень глубокие темные воды, где они были менее уязвимы для зрячих хищников и где выгодно иметь выпуклое глазное пятно, собирающее больше света, чем плоское или вогнутое. Это привело бы к несколько иной траектории эволюции глаз позвоночных, чем глаз других животных.
Тонкий разрастание прозрачных клеток над апертурой глаза, изначально сформированное для предотвращения повреждения глазного пятна, позволило сегрегированному содержимому глазной камеры специализироваться на прозрачной жидкости, которая оптимизировала цветовую фильтрацию, блокировала вредное излучение, улучшала показатель преломления глаза и разрешенная функциональность вне воды. Прозрачные защитные клетки в конечном итоге разделились на два слоя, между которыми находилась циркулирующая жидкость, что позволило расширить углы обзора и повысить разрешение изображений, а толщина прозрачного слоя постепенно увеличивалась, у большинства видов с прозрачным белком -кристаллином . [39]
Зазор между слоями ткани естественным образом образует двояковыпуклую форму, оптимально идеальную структуру для нормального показателя преломления. Независимо друг от друга вперед от хрусталика отделяются прозрачный и непрозрачный слои: роговица и радужная оболочка . При отделении переднего слоя снова образовалась влага, водянистая влага . Это увеличило преломляющую силу и снова облегчило проблемы с кровообращением. Формирование непрозрачного кольца позволило увеличить количество кровеносных сосудов, улучшить кровообращение и увеличить размер глаз. [39]
Отношение к жизненным требованиям
Глаза обычно приспособлены к окружающей среде и жизненным потребностям организма, который их носит. Например, распределение фоторецепторов имеет тенденцию соответствовать области, в которой требуется наибольшая острота зрения: организмы, сканирующие горизонт, такие как те, которые живут на африканских равнинах, имеют горизонтальную линию ганглиев с высокой плотностью, а обитают на деревьях. существа, которым требуется хорошее всестороннее зрение, как правило, имеют симметричное расположение ганглиев, при этом острота зрения снижается кнаружи от центра.
Конечно, для большинства типов глаз невозможно отклониться от сферической формы, поэтому изменять можно только плотность оптических рецепторов. У организмов со сложными глазами именно количество омматидий, а не ганглиев, отражает область наибольшего сбора данных. [1] : 23–24 Глаза оптической суперпозиции ограничены сферической формой, но другие формы сложных глаз могут деформироваться до формы, в которой большее количество омматидий ориентировано, скажем, на горизонт, без изменения размера или плотности отдельных омматидий. [40] Глаза организмов, сканирующих горизонт, имеют стебли, поэтому их можно легко выровнять по горизонту, когда он наклонен, например, если животное находится на склоне. [27]
Расширением этой концепции является то, что глаза хищников обычно имеют зону очень острого зрения в центре, помогающую идентифицировать добычу. [40] У глубоководных организмов увеличен не центр глаза. Гиперидные . амфиподы — глубоководные животные, питающиеся организмами, находящимися над ними Их глаза почти разделены на две части, причем верхняя часть, как полагают, участвует в обнаружении силуэтов потенциальной добычи (или хищников) на фоне слабого света неба над головой. Соответственно, более глубокие водные гиперииды, у которых свет, с которым необходимо сравнивать силуэты, более тусклый, имеют более крупные «верхние глаза» и могут вообще потерять нижнюю часть глаз. [40] У гигантского антарктического изопода Glyptonotus небольшой вентральный сложный глаз физически полностью отделен от гораздо большего дорсального сложного глаза. [41] Восприятие глубины можно улучшить, увеличив глаза в одном направлении; небольшое искажение глаза позволяет оценить расстояние до объекта с высокой степенью точности. [8]
Острота зрения выше у самцов, которые спариваются в воздухе, поскольку им необходимо иметь возможность обнаружить и оценить потенциальных партнеров на очень большом фоне. [40] С другой стороны, глаза организмов, которые действуют при слабом уровне освещенности, например, на рассвете и в сумерках или на глубокой воде, имеют тенденцию быть больше, чтобы увеличить количество света, которое можно уловить. [40]
Образ жизни может влиять не только на форму глаз. Глаза могут быть наиболее видимыми частями организмов, и это может оказывать на организмы давление, заставляя их иметь более прозрачные глаза в ущерб их функции. [40]
Глаза можно установить на стебельках, чтобы обеспечить лучший круговой обзор, подняв их над панцирем организма; это также позволяет им отслеживать хищников или добычу, не двигая головой. [8]
Физиология
Острота зрения

Острота зрения , или разрешающая способность, — это «способность различать мелкие детали» и является свойством колбочек . [42] Его часто измеряют в циклах на градус (CPD), которые измеряют угловое разрешение или то, насколько глаз может отличить один объект от другого с точки зрения углов обзора. Разрешение в CPD можно измерить с помощью гистограмм различного количества циклов белой/черной полосы. Например, если каждый узор имеет ширину 1,75 см и расположен на расстоянии 1 м от глаза, он образует угол в 1 градус, поэтому количество пар белых/черных полос на узоре будет мерой количества циклов на один градус. степень этого паттерна. Наибольшее такое число, которое глаз может различить как полосы или отличить от серого блока, является тогда измерением остроты зрения глаза.
Для человеческого глаза с превосходной остротой максимальное теоретическое разрешение составляет 50 CPD. [43] (1,2 угловых минуты на пару линий или пару линий 0,35 мм на расстоянии 1 м). Крыса может разрешить только около 1-2 CPD. [44] У лошади более высокая острота большей части поля зрения, чем у человека, но она не соответствует высокой остроте центральной ямки человеческого глаза . [45]
Сферическая аберрация ограничивает разрешение зрачка диаметром 7 мм примерно до 3 угловых минут на пару линий. При диаметре зрачка 3 мм сферическая аберрация значительно уменьшается, что приводит к улучшению разрешения примерно до 1,7 угловых минут на пару линий. [46] Разрешение в 2 угловые минуты на пару линий, что эквивалентно промежутку в 1 угловую минуту в оптотипе , соответствует 20/20 ( нормальному зрению ) у людей.
Однако в сложном глазу разрешение зависит от размера отдельных омматидий и расстояния между соседними омматидиями. Физически их невозможно уменьшить в размерах, чтобы достичь остроты зрения, наблюдаемой в однохрусталиковых глазах, как у млекопитающих. Сложные глаза имеют гораздо меньшую остроту, чем глаза позвоночных. [47]
Восприятие цвета
«Цветовое зрение — это способность организма различать свет разных спектральных качеств». [48] Все организмы ограничены небольшим диапазоном электромагнитного спектра; это варьируется от существа к существу, но в основном находится в диапазоне длин волн от 400 до 700 нм. [49] Это довольно небольшой участок электромагнитного спектра, вероятно, отражающий подводную эволюцию органа: вода блокирует все, кроме двух небольших окон ЭМ-спектра, и среди наземных животных не было эволюционного давления, направленного на расширение этого диапазона. [50]
Самый чувствительный пигмент родопсин имеет пиковый отклик при длине волны 500 нм. [51] Небольшие изменения в генах, кодирующих этот белок, могут изменить пиковую реакцию на несколько нм; [52] Пигменты в линзах также могут фильтровать входящий свет, изменяя пиковый отклик. [52] Многие организмы не способны различать цвета, вместо этого видят оттенки серого; Для цветового зрения необходим ряд пигментных клеток, которые в первую очередь чувствительны к меньшим диапазонам спектра. У приматов, гекконов и других организмов они принимают форму колбочек , из которых произошли более чувствительные палочки . [51] Даже если организмы физически способны различать разные цвета, это не обязательно означает, что они могут воспринимать разные цвета; Это можно выяснить только с помощью поведенческих тестов. [52]
Большинство организмов, обладающих цветовым зрением, способны улавливать ультрафиолетовый свет. Этот свет высокой энергии может повредить рецепторные клетки. За некоторыми исключениями (змеи, плацентарные млекопитающие), большинство организмов избегают этих эффектов, располагая абсорбирующими каплями масла вокруг своих колбочек. Альтернатива, разработанная организмами, которые потеряли эти капли масла в ходе эволюции, состоит в том, чтобы сделать хрусталик непроницаемым для ультрафиолетового света — это исключает возможность обнаружения любого ультрафиолетового света, поскольку он даже не достигает сетчатки. [51]
Палочки и колбочки
Сетчатка содержит два основных типа светочувствительных фоторецепторных клеток, используемых для зрения: палочки и колбочки .
Палочки не различают цвета, но отвечают за при слабом освещении ( скотопическое монохромное ( черно-белое ) зрение ); они хорошо работают при тусклом свете, поскольку содержат пигмент родопсин (зрительный фиолетовый), который чувствителен при низкой интенсивности света, но насыщает при более высокой ( фотопической ) интенсивности. Палочки распределены по всей сетчатке, но их нет ни в ямке , ни в слепом пятне . Плотность палочек больше в периферической сетчатке, чем в центральной.
Колбочки отвечают за цветовое зрение . Для работы им требуется более яркий свет, чем требуется стержням. У человека есть три типа колбочек, максимально чувствительных к длинноволновому, средневолновому и коротковолновому свету (часто называемому красным, зеленым и синим соответственно, хотя пики чувствительности на самом деле не приходятся на эти цвета). ). Видимый цвет представляет собой совокупный эффект стимулов и реакций этих трех типов колбочек. Колбочки в основном сосредоточены в ямке и рядом с ней. Лишь немногие из них присутствуют по бокам сетчатки. Объекты наиболее четко видны в фокусе, когда их изображения попадают в ямку, например, когда человек смотрит на объект прямо. Колбочки и палочки через промежуточные клетки сетчатки соединяются с нервными волокнами зрительного нерва . Когда палочки и колбочки стимулируются светом, они соединяются через соседние клетки сетчатки и посылают электрический сигнал волокнам зрительного нерва. Зрительные нервы посылают импульсы через эти волокна в мозг. [51]
Пигментация
Молекулы пигментов, используемые в глазах, разнообразны, но их можно использовать для определения эволюционной дистанции между различными группами, а также помочь определить, какие из них тесно связаны, хотя проблемы конвергенции действительно существуют. [51]
Опсины – это пигменты, участвующие в фоторецепции. Другие пигменты, такие как меланин , используются для защиты фоторецепторных клеток от света, проникающего с боков.Группа белков опсинов возникла задолго до появления последнего общего предка животных и с тех пор продолжает диверсифицироваться. [52]
В зрении участвуют два типа опсина; c-опсины, которые связаны с фоторецепторными клетками цилиарного типа, и r-опсины, связанные с рабдомерными фоторецепторными клетками. [53] Глаза позвоночных обычно содержат ресничные клетки с c-опсинами, а у (двусторонних) беспозвоночных в глазу имеются рабдомерные клетки с r-опсинами. Однако некоторые ганглиозные клетки позвоночных животных экспрессируют r-опсины, что позволяет предположить, что их предки использовали этот пигмент для зрения, и что его остатки выживают в глазах. [53] Аналогичным образом было обнаружено, что к-опсины экспрессируются в мозге некоторых беспозвоночных. Возможно, они экспрессировались в реснитчатых клетках личиночных глаз, которые впоследствии резорбировались в мозг при метаморфозе во взрослую форму. [53] C-опсины также обнаружены в некоторых производных глазах двусторонних беспозвоночных, таких как мантийные глаза двустворчатых моллюсков; однако боковые глаза (которые предположительно были предковым типом для этой группы, если глаза когда-то там развились) всегда используют r-опсины. [53] Cnidaria , которые являются внешней группой упомянутых выше таксонов, экспрессируют к-опсины, но в этой группе р-опсины еще не обнаружены. [53] Между прочим, меланин, вырабатываемый книдариями, вырабатывается таким же образом, как и у позвоночных, что позволяет предположить общее происхождение этого пигмента. [53]
Дополнительные изображения
 Структуры глаза, обозначенные
Структуры глаза, обозначенные Другой вид глаза и его структур, обозначенных как
Другой вид глаза и его структур, обозначенных как


См. также
- Аккомодация (глаз позвоночного) (фокусировка)
- Адаптация (глаз) (ночное видение)
- Капсула линзы
- Роговица
- Теория выбросов (видение)
- Цвет глаз
- Развитие глаз
- Болезни глаз
- Травма глаза
- Движение глаз
- веко
- Линза (анатомия позвоночных)
- Мигательная перепонка
- Офтальмология
- Орбита (анатомия)
- Простой глаз у беспозвоночных
- Яркий ковер
- Слезы
Примечания
- ^ Не существует универсального консенсуса относительно точного общего количества типов Animalia; заявленная цифра незначительно варьируется от автора к автору.
Ссылки
Цитаты
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление Земля, МФ; Фернальд, Р.Д. (1992). «Эволюция глаз». Ежегодный обзор неврологии . 15 : 1–29. дои : 10.1146/annurev.ne.15.030192.000245 . ПМИД 1575438 .
- ^ Брейтмейер, Бруно (2010). Слепые пятна: множество способов, которыми мы не можем видеть . Нью-Йорк: Издательство Оксфордского университета. п. 4 . ISBN 978-0-19-539426-9 .
- ^ Нэрн, Джеймс (2005). Психология . Бельмонт: Издательство Wadsworth. ISBN 978-0-495-03150-5 . OCLC 61361417 . Архивировано из оригинала 17 января 2023 г. Проверено 19 октября 2020 г.
- ^ Брюс, Вики; Грин, Патрик Р.; Джорджсон, Марк А. (1996). Зрительное восприятие: физиология, психология и экология . Психология Пресс. п. 20. ISBN 978-0-86377-450-8 . Архивировано из оригинала 17 января 2023 г. Проверено 19 октября 2020 г.
- ^ Кирк, Молли; Деннинг, Дэвид (2001). «У какого животного более развитый глаз, у осьминога или насекомого?» . БиоМедиа Ассошиэйтс. Архивировано из оригинала 26 февраля 2017 года.
- ^ "Кого ты называешь "креветкой"?" . Национальный журнал дикой природы . Национальная федерация дикой природы . 1 октября 2010 года. Архивировано из оригинала 9 августа 2010 года . Проверено 3 апреля 2014 г.
- ^ Мейер-Рохов, В.Б. (1974). «Строение и функции личиночного глаза пилильщика Перга ». Журнал физиологии насекомых . 20 (8): 1565–1591. дои : 10.1016/0022-1910(74)90087-0 . ПМИД 4854430 .
- ^ Jump up to: а б с д Кронин, Т.В.; Портер, ML (2008). «Исключительная вариация на общую тему: эволюция сложных глаз ракообразных» . Эволюция: образование и информационно-пропагандистская деятельность . 1 (4): 463–475. дои : 10.1007/s12052-008-0085-0 .
- ^ Козьмик З.; Ружичкова Дж.; Йонасова, К.; Мацумото, Ю.; Вопаленский, П.; Козьмикова И.; Стрнад, Х.; Кавамура, С.; Пятигорский Ю.; и др. (2008). «Сборка книдарийного камерного глаза из компонентов, подобных позвоночным» (Полный текст бесплатно) . Труды Национальной академии наук Соединенных Штатов Америки . 105 (26): 8989–8993. Бибкод : 2008PNAS..105.8989K . дои : 10.1073/pnas.0800388105 . ПМЦ 2449352 . ПМИД 18577593 .
- ^ Жуков З.Х.; Бориссеко, С.Л.; Зигер, М.В.; Ваколюк И.А.; Мейер-Рохов, В.Б. (2006). «Глаз пресноводного переднежаберного брюхоногого моллюска Viviparus viviparus: ультраструктура, электрофизиология и поведение». Акта Зоология . 87 : 13–24. дои : 10.1111/j.1463-6395.2006.00216.x .
- ^ Фернальд, Рассел Д. (2006). «Проливая генетический свет на эволюцию глаз». Наука . 313 (5795): 1914–1918. Бибкод : 2006Sci...313.1914F . дои : 10.1126/science.1127889 . ПМИД 17008522 . S2CID 84439732 .
- ^ «Информационный бюллетень о циркадных ритмах» . Национальный институт общих медицинских наук. Архивировано из оригинала 13 марта 2020 года . Проверено 3 июня 2015 г.
- ^ Нильссон, Дэн-Э. (1989). «Оптика зрения и эволюция». Бионаука . 39 (5): 298–307. дои : 10.2307/1311112 . JSTOR 1311112 .
- ^ Глаза животных
- ^ Глубокое восприятие, Том 1: Основные механизмы
- ^ Спейзер, Д.И.; Эрниссе, диджей; Джонсен, С.Н. (2011). «Хитон использует линзы из арагонита для формирования изображений» . Современная биология . 21 (8): 665–670. Бибкод : 2011CBio...21..665S . дои : 10.1016/j.cub.2011.03.033 . ПМИД 21497091 . S2CID 10261602 .
- ^ Jump up to: а б с Уилсон, М. (1978). «Функциональная организация глазков саранчи». Журнал сравнительной физиологии . 124 (4): 297–316. дои : 10.1007/BF00661380 . S2CID 572458 .
- ^ Вагнер, HJ; Дуглас, Р.Х.; Фрэнк, ТМ; Робертс, Н.В. и Партридж, Дж.К. (27 января 2009 г.). «Новый глаз позвоночных, использующий как преломляющую, так и отражающую оптику» . Современная биология . 19 (2): 108–114. Бибкод : 2009CBio...19..108W . дои : 10.1016/j.cub.2008.11.061 . ПМИД 19110427 . S2CID 18680315 .
- ^ Фёлькель, Р; Эйснер, М; Вейбл, К.Дж. (июнь 2003 г.). «Миниатюрные системы визуализации» (PDF) . Микроэлектронная инженерия . 67–68 (1): 461–472. дои : 10.1016/S0167-9317(03)00102-3 . Архивировано из оригинала (PDF) 1 октября 2008 г.
- ^ Лэнд, Майкл (1997). «Острота зрения насекомых» (PDF) . Ежегодный обзор энтомологии . 42 : 147–177. дои : 10.1146/annurev.ento.42.1.147 . ПМИД 15012311 . Архивировано из оригинала (PDF) 23 ноября 2004 года . Проверено 27 мая 2013 г.
- ^ Гейтен, Эдвард (1998). «Оптика и филогения: есть ли понимание? Эволюция суперпозиционных глаз у десятиногих (ракообразных)» . Вклад в зоологию . 67 (4): 223–236. дои : 10.1163/18759866-06704001 .
- ^ Ричи, Александр (1985). « Ainiktozoon loganense Scourfield, протохордовый из силура Шотландии». Алчеринга . 9 (2): 137. Бибкод : 1985Alch....9..117R . дои : 10.1080/03115518508618961 .
- ^ Майер, Г. (2006). «Строение и развитие онихофорных глаз: какой предковый орган зрения у членистоногих?». Строение и развитие членистоногих . 35 (4): 231–245. Бибкод : 2006АртСД..35..231М . дои : 10.1016/j.asd.2006.06.003 . ПМИД 18089073 .
- ^ Мейер-Рохов, В.Б.; Гал, Дж (2004). «Предельные размеры глаз членистоногих с суперпозиционной оптикой» . Исследование зрения . 44 (19): 2213–2223. дои : 10.1016/j.visres.2004.04.009 . ПМИД 15208008 .
- ^ Грейнер, Биргит (16 декабря 2005 г.). Адаптации ночного зрения в аппозиционных глазах насекомых (PDF) (доктор философии). Лундский университет. Архивировано из оригинала (PDF) 9 февраля 2013 года . Проверено 13 ноября 2014 г.
- ^ Хорват, Габор; Кларксон, Юан НК (1997). «Обзор современных аналогов глаз шизохроального трилобита: структурные и функциональные сходства и различия». Историческая биология . 12 (3–4): 229–263. Бибкод : 1997HBio...12..229H . дои : 10.1080/08912969709386565 .
- ^ Jump up to: а б Йохен Цайль; Маха М. Аль-Мутаири (1996). «Вариации оптических свойств сложных глаз Uca Lactea annulipes » (PDF) . Журнал экспериментальной биологии . 199 (7): 1569–1577. дои : 10.1242/jeb.199.7.1569 . ПМИД 9319471 . Архивировано (PDF) из оригинала 25 февраля 2009 г. Проверено 15 сентября 2008 г.
- ^ Мюллер, CHG; Розенберг, Дж; Рихтер, С; Мейер-Рохов, В.Б. (2003). «Сложный глаз Scutigera coleoptrata (Linnaeus, 1758) (Chilopoda; Notostigmophora): ультраструктурное повторное исследование, которое дополняет концепцию Mandibulata». Зооморфология . 122 (4): 191–209. дои : 10.1007/s00435-003-0085-0 . S2CID 6466405 .
- ^ Ульрих-Лютер, EM; Дюпон, С.; Арболеда, Э.; Хаузен, Х.; Арноне, Мичиган (2011). «Уникальная система фоторецепторов в трубчатых ножках морского ежа» . Труды Национальной академии наук . 108 (20): 8367–8372. Бибкод : 2011PNAS..108.8367U . дои : 10.1073/pnas.1018495108 . ПМК 3100952 . ПМИД 21536888 .
- ^ Али и Клайн 1985 , с. 8
- ^ Отрум, Х (1979). "Введение". В Х. Отруме (ред.). Сравнительная физиология и эволюция зрения у беспозвоночных-А: Фоторецепторы беспозвоночных . Справочник по сенсорной физиологии. Том. VII/6А. Нью-Йорк: Springer-Verlag. стр. 4, 8–9. ISBN 978-3-540-08837-0 .
- ^ Гальдер, Г.; Каллаертс, П.; Геринг, WJ (1995). «Новые взгляды на эволюцию глаз». Курс. Мнение. Жене. Дев . 5 (5): 602–609. дои : 10.1016/0959-437X(95)80029-8 . ПМИД 8664548 .
- ^ Гальдер, Г.; Каллаертс, П.; Геринг, WJ (1995). «Индукция эктопических глаз путем целевой экспрессии безглазого гена у дрозофилы ». Наука . 267 (5205): 1788–1792. Бибкод : 1995Sci...267.1788H . дои : 10.1126/science.7892602 . ПМИД 7892602 .
- ^ Томарев С.И.; Каллаертс, П.; Кос, Л.; Зиновьева Р.; Гальдер, Г.; Геринг, В.; Пятигорский, Дж. (1997). «Кальмар Пакс-6 и развитие глаз» . Учеб. Натл. акад. наук. США . 94 (6): 2421–2426. Бибкод : 1997PNAS...94.2421T . дои : 10.1073/pnas.94.6.2421 . ЧВК 20103 . ПМИД 9122210 .
- ^ Конвей-Моррис, С. (1998). Горнило Творения . Оксфорд: Издательство Оксфордского университета
- ^ Jump up to: а б Тревор Д. Лэмб; Шон П. Коллин; Эдвард Н. Пью младший (2007). «Эволюция глаза позвоночных: опсины, фоторецепторы, сетчатка и глазная чашечка» . Обзоры природы Неврология . 8 (12): 960–976. дои : 10.1038/nrn2283 . ПМК 3143066 . ПМИД 18026166 .
- ^ Стаяслав Иванович Томарев; Рина Дмитриевна Зиновьева (1988). «Полипептиды основных хрусталиков кальмара гомологичны субъединицам глутатион S-трансфераз». Природа . 336 (6194): 86–88. Бибкод : 1988Natur.336...86T . дои : 10.1038/336086a0 . ПМИД 3185725 . S2CID 4319229 .
- ^ «Эволюция глаз?» . Library.thinkquest.org. Архивировано из оригинала 15 сентября 2012 г. Проверено 1 сентября 2012 г.
- ^ Jump up to: а б Фернальд, Рассел Д. (2001). Эволюция глаз: откуда появились линзы? Архивировано 19 марта 2006 г. в Wayback Machine Karger Gazette 64: «Глаз в фокусе».
- ^ Jump up to: а б с д и ж Земля, МФ (1989). «Глаза гипериидных амфипод: связь оптической структуры с глубиной». Журнал сравнительной физиологии А. 164 (6): 751–762. дои : 10.1007/BF00616747 . S2CID 23819801 .
- ^ Мейер-Рохов, Виктор Бенно (1982). «Разделенный глаз изоподы Glyptonotus antarcticus: эффекты односторонней темновой адаптации и повышения температуры». Труды Лондонского королевского общества . Б 215 (1201): 433–450. Бибкод : 1982РСПСБ.215..433М . дои : 10.1098/rspb.1982.0052 . S2CID 85297324 .
- ^ Али и Клайн 1985 , с. 28
- ^ Расс, Джон К. (2006). Руководство по обработке изображений . ЦРК Пресс. ISBN 978-0-8493-7254-4 . OCLC 156223054 . Архивировано из оригинала 17 января 2023 г. Проверено 19 октября 2020 г.
Верхний предел (мельчайшие детали), видимый человеческим глазом, составляет около 50 циклов на градус... (Пятое издание, 2007 г., стр. 94)
- ^ Клаассен, Кертис Д. (2001). Токсикология Казаретта и Дулла: фундаментальная наука о ядах . МакГроу-Хилл Профессионал. ISBN 978-0-07-134721-1 . OCLC 47965382 . Архивировано из оригинала 17 января 2023 г. Проверено 19 октября 2020 г.
- ^ «Сетчатка человеческого глаза» . гиперфизика.phy-astr.gsu.edu . Архивировано из оригинала 4 мая 2015 г. Проверено 3 июня 2015 г.
- ^ Фишер, Роберт Э.; Тадич-Галеб, Биляна; Плимптон, Рик (2000). Стив Чепмен (ред.). Проектирование оптических систем . МакГроу-Хилл Профессионал. ISBN 978-0-07-134916-1 . OCLC 247851267 . Архивировано из оригинала 17 января 2023 г. Проверено 19 октября 2020 г.
- ^ Барлоу, HB (1952). «Размер омматидий в аппозиционных глазах» . J Exp Biol . 29 (4): 667–674. дои : 10.1242/jeb.29.4.667 . Архивировано из оригинала 31 августа 2016 г. Проверено 1 января 2012 г.
- ^ Али и Клайн 1985 , с. 161
- ^ Барлоу, Гораций Бэзил; Моллон, JD (1982). Чувства . Кембридж: Издательство Кембриджского университета. п. 98 . ISBN 978-0-521-24474-9 .
- ^ Фернальд, Рассел Д. (1997). «Эволюция глаз» (PDF) . Мозг, поведение и эволюция . 50 (4): 253–259. дои : 10.1159/000113339 . ПМИД 9310200 . Архивировано из оригинала 20 ноября 2012 г. Проверено 16 сентября 2008 г.
- ^ Jump up to: а б с д и Голдсмит, TH (1990). «Оптимизация, ограничения и история эволюции глаз». Ежеквартальный обзор биологии . 65 (3): 281–322. дои : 10.1086/416840 . JSTOR 2832368 . ПМИД 2146698 . S2CID 24535762 .
- ^ Jump up to: а б с д Френтиу, Франческа Д.; Адриана Д. Бриско (2008). «Взгляд на птиц глазами бабочки». Биоэссе . 30 (11–12): 1151–1162. doi : 10.1002/bies.20828 . ПМИД 18937365 . S2CID 34409725 .
- ^ Jump up to: а б с д и ж Нильссон, Э.; Арендт, Д. (декабрь 2008 г.). «Эволюция глаз: размытое начало» . Современная биология . 18 (23): 1096–1098 р. Бибкод : 2008CBio...18R1096N . дои : 10.1016/j.cub.2008.10.025 . ПМИД 19081043 . S2CID 11554469 .
Библиография
- Али, Мохамед Атер; Клайн, Массачусетс (1985). Зрение у позвоночных . Нью-Йорк: Пленум Пресс . ISBN 978-0-306-42065-8 .
Дальнейшее чтение
- Йонг, Эд (14 января 2016 г.). «Внутри глаза: самое изысканное творение природы» . Нэшнл Географик . Архивировано из оригинала 14 января 2016 года.
Внешние ссылки
- Эволюция глаза
- Анатомия глаза – флеш-анимационный интерактив. ( Адобе Флеш )
- Вебвидение. Организация сетчатки и зрительной системы. Углубленное лечение функции сетчатки, доступное всем, но больше всего ориентированное на аспирантов.
- Исследование Калифорнийского университета в Беркли показывает, что глаза удаляют все изображения, кроме самого необходимого, прежде чем отправлять визуальную информацию в мозг.
