Головоногий глаз
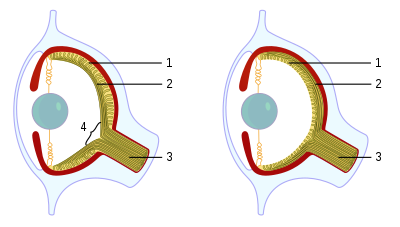
Головоногие моллюски , как активные морские хищники, обладают органами чувств, специализированными для использования в водных условиях. [1] У них есть глаз типа камеры , который состоит из радужной оболочки, круглой линзы, полости стекловидного тела (глазного геля), пигментных клеток и фоторецепторных клеток, которые преобразуют свет от светочувствительной сетчатки в нервные сигналы, которые проходят по зрительному нерву к глазу. мозг. [2] В течение последних 140 лет глаз головоногих моллюсков сравнивали с глазом позвоночных как пример конвергентной эволюции , когда оба типа организмов независимо развили признак глаз-камеры и оба имеют схожие функциональные возможности. Существуют споры о том, действительно ли это конвергентная эволюция или параллельная эволюция . [3] В отличие от глаза-камеры позвоночных , головоногие моллюски имеют форму впячиваний на поверхность тела (а не выростов головного мозга), и, следовательно, роговица лежит над верхней частью глаза, а не является его структурной частью. [4] В отличие от глаз позвоночных, глаз головоногих фокусируется за счет движения, подобно линзе камеры или телескопа, а не за счет изменения формы, как это делает хрусталик в человеческом глазу. Глаз хрусталик примерно сферический, как и , который полностью внутренний. [5]
Глаза головоногих моллюсков развиваются таким образом, что у них есть ретинальные аксоны, которые проходят через заднюю часть сетчатки, поэтому зрительный нерв не должен проходить через фоторецепторный слой, чтобы выйти из глаза, и не имеет естественной центральной физиологической слепоты. пятно позвоночных. [2]
Кристаллины , используемые в хрусталике, по-видимому, развились независимо от кристаллинов позвоночных, что указывает на гомоплазное происхождение хрусталика. [6]
Большинство головоногих моллюсков обладают сложной системой экстраокулярных мышц, которая позволяет очень точно контролировать положение глаз. Осьминоги обладают вегетативной реакцией, которая поддерживает ориентацию их зрачков так, что они всегда находятся в горизонтальном положении. [1]
Поляризованный свет
[ редактировать ]Некоторые виды головоногих моллюсков, особенно кальмары и осьминоги, а также, возможно, каракатицы, имеют глаза, способные различать ориентацию поляризованного света . Эта чувствительность обусловлена ортогональной организацией соседних фоторецепторов . (У головоногих есть рецепторные клетки, называемые рабдомами, подобные таковым у других моллюсков.) Напротив, глаз позвоночных обычно нечувствителен к различиям в поляризации, поскольку опсины в палочках и колбочках расположены полуслучайно. Таким образом, глаз одинаково чувствителен к любой ориентации электронной оси света. Из-за своей ортогональной организации опсины в глазах головоногих имеют наибольшее светопоглощение при правильном выравнивании с осью электронного вектора света, что обеспечивает чувствительность к различиям в поляризации. [7] Точная функция этой способности не доказана, но предполагается, что она предназначена для обнаружения добычи, навигации и, возможно, общения между меняющими цвет головоногими моллюсками. [7] [8]
 Глаз Bathyteuthis sp.
Глаз Bathyteuthis sp. Глаз осьминога ( Octopus vulgaris )
Глаз осьминога ( Octopus vulgaris ) кальмара Глаз
кальмара Глаз каракатицы Глаз
каракатицы Глаз Наутилус ( Nautilus pompilius ) глаз
Наутилус ( Nautilus pompilius ) глаз





Эволюционные дебаты
[ редактировать ]Разногласия относительно того, является ли эволюция камерного глаза у головоногих и позвоночных параллельными или конвергентными , все еще существуют, хотя в основном разрешены. В настоящее время наблюдается конвергентная эволюция их аналогичного глаза камерного типа.
Параллельная эволюция
[ редактировать ]Те, кто утверждает, что это параллельная эволюция, утверждают, что существуют доказательства существования общего предка, содержащего генетическую информацию для развития этого глаза. Об этом свидетельствуют все двулатеральные организмы , содержащие ген Pax6 , отвечающий за развитие глаз. [9]
Конвергентная эволюция
[ редактировать ]Сторонники конвергентной эволюции утверждают, что этот общий предок значительно предшествовал как головоногим, так и позвоночным. Общий предок с выражением глаза камерного типа должен был существовать примерно за 270 миллионов лет до эволюции глаза камерного типа у головоногих моллюсков и примерно за 110–260 миллионов лет до эволюции глаза камерного типа у позвоночных. [10] Другим источником доказательств этого являются различия в экспрессии, обусловленные независимыми вариантами Pax6 , возникающими как у головоногих, так и у позвоночных. Головоногие моллюски содержат в своих геномах пять вариантов Pax6 , которые возникли независимо и не являются общими для позвоночных, хотя они допускают аналогичную экспрессию генов по сравнению с Pax6 позвоночных. [11]
Исследования и медицинское использование
[ редактировать ]Основное медицинское применение в этой области – исследования развития глаз и глазных заболеваний . Новые исследования экспрессии глазных генов проводятся с использованием глаз головоногих моллюсков в связи с доказательствами их конвергентной эволюции с аналогичным человеческим глазом. Эти исследования заменяют предыдущие исследования экспрессии генов дрозофилы во время развития глаз как наиболее точные, хотя исследования дрозофилы остаются наиболее распространенными. Вывод о том, что они аналогичны, придает достоверность их сравнению, прежде всего, для медицинского использования, поскольку признак в обоих случаях сформировался в результате естественного отбора под одинаковым давлением в одинаковых условиях; это означает, что в глазах обоих организмов будет одинаковое проявление глазного заболевания. [2]
Преимущество экспериментов на глазах с головоногими моллюсками состоит в том, что головоногие могут регенерировать свои глаза благодаря своей способности возобновлять процессы развития, что позволяет продолжить исследования одних и тех же головоногих после одного пробного образца при изучении последствий болезни. Это также позволяет провести более комплексное исследование того, как регенерация может сохраняться в геномах головоногих моллюсков и может ли она в некоторой степени сохраняться в геноме человека наряду с генами, экспрессирующими глаз-камеру. [2]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Будельманн БУ. «Органы чувств, нервы и мозг головоногих моллюсков: адаптация к высокой производительности и образу жизни». Морское и пресноводное поведение и физиология. Том 25, выпуск 1–3, страницы 13–33.
- ^ Перейти обратно: а б с д Сербка Жанна М. (2008). «На пути к разработке моделей для изучения заболеваний, экологии и эволюции глаза моллюсков *» (PDF) . Американский малакологический бюллетень . 26 (1–2): 3–18. дои : 10.4003/006.026.0202 . S2CID 1557944 . Архивировано из оригинала (PDF) 18 декабря 2014 г. Проверено 18 ноября 2014 г.
- ^ Серб, Дж.; Эрнисс, Д. (2008). «Изложение траектории эволюции: использование разнообразия глаз моллюсков для понимания параллельной и конвергентной эволюции» . Эволюция: образование и информационно-пропагандистская деятельность . 1 (4): 439–447. дои : 10.1007/s12052-008-0084-1 .
- ^ Ханке, Фредерика Д.; Кельбер, Альмут (14 января 2020 г.). «Глаз осьминога обыкновенного (Octopus vulgaris)» . Границы в физиологии . 10 : 1637. doi : 10.3389/fphys.2019.01637 . ISSN 1664-042X . ПМК 6971404 . ПМИД 32009987 .
- ^ Ямамото, М. (февраль 1985 г.). «Онтогенез зрительной системы каракатицы Sepiella japonica . I. Морфологическая дифференциация зрительной клетки». Журнал сравнительной неврологии . 232 (3): 347–361. дои : 10.1002/cne.902320307 . ISSN 0021-9967 . ПМИД 2857734 . S2CID 24458056 .
- ^ САМИР К. БРАХМА1 (1978). «Онтогенез кристаллинов хрусталика морских головоногих моллюсков» (PDF) . Эмбриол. Эксп. Морф . 46 (1): 111–118. ПМИД 359745 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Перейти обратно: а б Матгер, LM; Шашар, Н.; Хэнлон, RT (2009). «Общаются ли головоногие моллюски, используя отражение поляризованного света от их кожи?». Журнал экспериментальной биологии . 212 (Часть 14): 2133–2140. дои : 10.1242/jeb.020800 . ПМИД 19561202 .
- ^ Шашар, Н; Ратледж, П; Кронин, Т. (1996). «Поляризационное зрение у каракатиц в скрытом канале связи?». Журнал экспериментальной биологии . 199 (9): 2077–2084. дои : 10.1242/jeb.199.9.2077 . ПМИД 9319987 .
- ^ Геринг, WJ (2004). «Исторический взгляд на развитие и эволюцию глаз и фоторецепторов» . Международный журнал биологии развития . 48 (8–9): 707–717. doi : 10.1387/ijdb.041900wg . ПМИД 15558463 .
- ^ Фернальд, Рассел Д. (29 сентября 2006 г.). «Проливая генетический свет на эволюцию глаз». Наука . 313 (5795): 1914–1918. Бибкод : 2006Sci...313.1914F . дои : 10.1126/science.1127889 . ПМИД 17008522 . S2CID 84439732 .
- ^ Ёсида, Масааки; Юра, Кей; Огура, Ацуши (2014). «Эволюция глаз головоногих моллюсков была модулирована приобретением вариантов сплайсинга Pax-6» . Научные отчеты . 4 (4256): 4256. Бибкод : 2014NatSR...4E4256Y . дои : 10.1038/srep04256 . ПМК 3942700 . ПМИД 24594543 .
головоногих моллюсков Анатомия |
|---|



Зрение у животных |
|---|
