Линза (анатомия позвоночных)
| Объектив | |
|---|---|
 Хрусталик глаза меняет форму, фокусируясь вблизи и вдалеке. | |
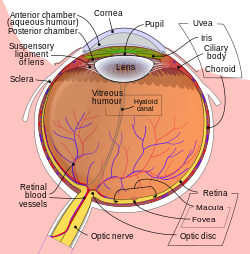 Принципиальная схема человеческого глаза . | |
| Подробности | |
| Часть | глазное яблоко |
| Система | Визуальная система |
| Функция | Преломлять свет |
| Идентификаторы | |
| латинский | хрусталик кристаллин |
| МеШ | D007908 |
| ТА98 | А15.2.05.001 |
| ТА2 | 6798 |
| ФМА | 58241 |
| Анатомическая терминология | |
Хрусталик прозрачную , или хрусталик , представляет собой двояковыпуклую структуру большинства наземных позвоночных в глазах . Относительно длинные и тонкие волокнистые клетки составляют большую часть хрусталика. Эти клетки различаются по архитектуре и расположены концентрическими слоями. Новые слои клеток рекрутируются из тонкого эпителия в передней части хрусталика, чуть ниже базальной мембраны, окружающей хрусталик. В результате хрусталик позвоночных растет на протяжении всей жизни. Окружающая хрусталик мембрана, называемая капсулой хрусталика, также систематически растет, обеспечивая сохранение оптически подходящей формы хрусталика вместе с нижележащими волокнистыми клетками. В капсулу наибольшего диаметра встроены тысячи поддерживающих связок, которые удерживают хрусталик внутри глаза. Большинство этих хрусталиковых структур происходят из эпителия эмбриона еще до рождения.
Вместе с роговицей , водянистой жидкостью и стекловидным телом он преломляет свет, фокусируя его на сетчатке . У многих наземных животных форма хрусталика может быть изменена, что позволяет эффективно изменять фокусное расстояние глаза, позволяя им фокусироваться на объектах на разных расстояниях. Такая регулировка хрусталика называется аккомодацией (см. также ниже ). У многих полностью водных позвоночных, таких как рыбы, используются другие методы аккомодации, такие как изменение положения хрусталика относительно сетчатки, а не изменение формы хрусталика. Аккомодация аналогична фокусировке фотокамеры путем смены линз . У наземных позвоночных хрусталик на передней стороне более плоский, чем на задней, а у рыб хрусталик часто близок к сферическому.
Аккомодация у людей хорошо изучена и позволяет использовать искусственные средства, дополняющие нашу концентрацию, такие как очки для коррекции зрения по мере старения. Преломляющая сила хрусталика молодого человека в естественной среде составляет примерно 18 диоптрий , что составляет примерно одну треть от общей силы глаза, составляющей около 60 диоптрий. К 25 годам способность хрусталика изменять путь света снижается до 10 диоптрий, а аккомодация с возрастом продолжает ухудшаться.
Структура
[ редактировать ]Позиция в глазу
[ редактировать ]Хрусталик расположен в передней части глаза позвоночного, называемой передним сегментом , который включает роговицу и радужную оболочку , расположенные перед хрусталиком. Хрусталик удерживается на месте поддерживающими связками (Zonule of Zinn) . [1] прикрепление линзы по экватору к остальной части глаза [2] [3] через цилиарное тело . За хрусталиком находится желеобразное стекловидное тело , которое удерживает хрусталик на месте. В передней части хрусталика находится жидкая водянистая влага , которая омывает хрусталик питательными веществами и другими веществами. Хрусталики наземных позвоночных обычно имеют эллипсоидную , двояковыпуклую форму. Передняя поверхность менее изогнута, чем задняя. У взрослого человека хрусталик обычно имеет диаметр около 10 мм и толщину 4 мм, но меняет форму в зависимости от аккомодации и размера из-за роста на протяжении всей жизни человека. [4]
Анатомия
[ редактировать ]



Хрусталик состоит из трех основных частей: капсулы хрусталика , эпителия хрусталика и волокон хрусталика. Капсула хрусталика представляет собой относительно толстую базальную мембрану, образующую внешний слой хрусталика. Внутри капсулы основную часть хрусталика составляют гораздо более тонкие волокна хрусталика. Клетки . эпителия хрусталика образуют тонкий слой между капсулой хрусталика и наружным слоем волокон хрусталика в передней части хрусталика, но не сзади В самом хрусталике отсутствуют нервы, кровеносные сосуды и соединительная ткань. [5] Анатомы часто ссылаются на положение структур в хрусталике, описывая его как глобус мира. Передняя и задняя части хрусталика называются передним и задним «полюсами», подобно северному и южному полюсам. «Экватор» — это внешний край хрусталика, часто скрытый радужной оболочкой, и область дифференцировки большинства клеток. Поскольку экватор обычно не находится на пути света глаза, структуры, участвующие в метаболической активности, избегают рассеивания света, который в противном случае мог бы повлиять на зрение.
Капсула линзы
[ редактировать ]

Капсула хрусталика представляет собой гладкую прозрачную базальную мембрану , полностью окружающую хрусталик. Капсула эластичная, ее основным структурным компонентом является коллаген . Предполагается, что он синтезируется эпителием хрусталика и его основными компонентами в порядке распространенности являются гепарансульфат-протеогликан (сульфатированные гликозаминогликаны (ГАГ)), энтактин, коллаген IV типа , ламинин. [6] Капсула очень эластична и позволяет хрусталику принимать более сферическую форму, когда напряжение поддерживающих связок снижается. Толщина капсулы человека варьируется от 2 до 28 микрометров, самая толстая вблизи экватора (приэкваториальная область) и обычно тоньше вблизи заднего полюса. [4]
На фотографиях электронного и светового микроскопов показан участок экватора хрусталика капсулы, где капсула растет и примыкает к местам прикрепления тысяч поддерживающих связок. [7] [8] Крепление должно быть достаточно прочным, чтобы предотвратить отсоединение связок от капсулы хрусталика. Силы возникают при удержании линзы на месте, а силы добавляются во время фокусировки. В то время как капсула самая тонкая на экваторе, где ее площадь увеличивается. [7] Передняя и задняя капсулы тоньше места прикрепления связок.
Эпителий хрусталика
[ редактировать ]хрусталика Эпителий представляет собой один слой клеток в передней части хрусталика между капсулой хрусталика и волокнами хрусталика. [4] Обеспечивая волокна хрусталика питательными веществами и удаляя отходы, клетки эпителия регулируют поддержание гомеостаза хрусталика . [9] Когда ионы, питательные вещества и жидкость попадают в хрусталик из водянистой влаги , Na + /К + -АТФазные насосы в эпителиальных клетках хрусталика откачивают ионы из хрусталика для поддержания соответствующей осмотической концентрации и объема хрусталика, при этом наибольший вклад в этот ток вносят экваториально расположенные эпителиальные клетки хрусталика. Активность Na + /К + -АТФазы удерживают воду и ток, протекающие через линзу от полюсов и выходящие через экваториальные области.
Клетки эпителия хрусталика также делятся на новые волокна хрусталика на экваторе хрусталика. [10] Хрусталик откладывает волокна с момента его первого формирования в эмбрионе до момента его смерти. [11]
Линзовые волокна
[ редактировать ]Волокна хрусталика составляют основную часть хрусталика. Это длинные, тонкие, прозрачные клетки, плотно упакованные, диаметром обычно 4–7 микрометров и длиной до 12 мм у человека. [4] Волокна хрусталика тянутся в длину от заднего полюса к переднему и, если их разрезать горизонтально, располагаются концентрическими слоями, напоминающими слои луковицы. Если разрезать по экватору, ячейки имеют шестиугольное поперечное сечение, напоминающее соты. [12] Приблизительная середина каждого волокна лежит вокруг экватора. [11] Эти плотно упакованные слои волокон хрусталика называются пластинками. Цитоплазмы волокон хрусталика связаны друг с другом посредством щелевых соединений , межклеточных мостиков и переплетений клеток, которые напоминают форму «шара и гнезда».
Хрусталик разделен на участки в зависимости от возраста волокон хрусталика конкретного слоя. Двигаясь наружу от центрального, самого старого слоя, хрусталик разделяется на эмбриональное ядро, эмбриональное ядро, взрослое ядро, внутреннюю и внешнюю кору. Новые волокна хрусталика, образующиеся из эпителия хрусталика, добавляются к внешней коре головного мозга. Зрелые волокна хрусталика не имеют органелл и ядер .
Слияние клеток, пустоты и вакуоли
[ редактировать ]

С появлением других способов рассмотрения клеточных структур хрусталика у живого животного стало очевидно, что участки волоконных клеток, по крайней мере в передней части хрусталика, содержат большие пустоты и вакуоли. Предполагается, что они участвуют в системах транспортировки хрусталика, связывающих поверхность хрусталика с более глубокими областями. [13] Очень похожие структуры также указывают на слияние клеток в хрусталике. Слияние клеток показано микроинъекцией с образованием стратифицированного синцития в культурах цельных хрусталиков. [10]
Разработка
[ редактировать ]
Развитие хрусталика позвоночных начинается, когда длина человеческого эмбриона составляет около 4 мм. На сопроводительном изображении показан процесс на более легко изучаемом курином эмбрионе. В отличие от остальной части глаза, которая образуется в основном из внутренних слоев эмбриона , хрусталик образуется из кожи вокруг эмбриона . Первый этап формирования хрусталика наступает, когда сфера клеток, образовавшаяся в результате отпочкования внутренних слоев зародыша, приближается к внешней коже эмбриона. Сфера клеток побуждает близлежащую внешнюю кожу начать превращаться в плакоду хрусталика . Плакод хрусталика — это первый этап трансформации участка кожи в хрусталик. На этой ранней стадии плакода хрусталика представляет собой один слой клеток . [14] [15]
По мере развития плакода хрусталика начинает углубляться и прогибаться внутрь. По мере того как плакода продолжает углубляться, отверстие в поверхностной эктодерме сужается. [16] и клетки хрусталика отпочковываются от кожи эмбриона, образуя сферу клеток, известную как «пузырек хрусталика». Когда эмбрион достигает длины около 10 мм, хрусталик полностью отделяется от кожи эмбриона.
Затем эмбрион посылает сигналы от развивающейся сетчатки, побуждая клетки, ближайшие к заднему концу пузырька хрусталика, удлиняться по направлению к переднему концу пузырька. [16] Эти сигналы также вызывают синтез белков, называемых кристаллинами . [17] Как следует из названия, кристаллины могут образовывать прозрачное желе с высокой преломляющей способностью. Эти удлиняющиеся клетки в конечном итоге заполняют центр пузырька клетками, длинными и тонкими, как прядь волос, называемыми волокнами. Эти первичные волокна становятся ядром зрелого хрусталика. Эпителиальные клетки, которые не образуют волокна, ближайшие к передней части хрусталика, дают начало эпителию хрусталика. [18]

Дополнительные волокна происходят из эпителиальных клеток хрусталика, расположенных на экваторе хрусталика. Эти клетки удлиняются вперед и назад, охватывая уже отложенные волокна. Новые волокна должны быть длиннее, чтобы покрывать предыдущие волокна, но по мере того, как хрусталик становится больше, концы новых волокон больше не доходят так далеко до передней и задней части хрусталика. Волокна хрусталика, не доходящие до полюсов, образуют с соседними волокнами плотные, переплетающиеся швы. Эти швы, менее кристаллические, чем основная часть хрусталика, более заметны и называются «швами». Рисунки швов становятся более сложными по мере того, как к внешней части хрусталика добавляется больше слоев волокон.
Хрусталик продолжает расти после рождения, при этом новые вторичные волокна добавляются в качестве внешних слоев. Новые волокна хрусталика образуются из экваториальных клеток эпителия хрусталика в области, называемой «герминативной зоной» и «луковой областью». Эпителиальные клетки хрусталика удлиняются, теряют контакт с капсулой и эпителием в задней и передней части хрусталика, синтезируют кристаллин , а затем, наконец, теряют свои ядра (энуклеируют), когда они становятся зрелыми волокнами хрусталика. У людей по мере роста хрусталика за счет отложения большего количества волокон вплоть до раннего взрослого возраста хрусталик приобретает более эллипсоидную форму. Примерно после 20 лет хрусталик снова становится круглым, и радужная оболочка очень важна для этого развития. [4]
Несколько белков контролируют эмбриональное развитие хрусталика, хотя PAX6 считается главным геном-регулятором этого органа. [19] Другие эффекторы правильного развития хрусталика включают передачи сигналов Wnt компоненты BCL9 и Pygo2 . [20] Весь процесс дифференцировки эпителиальных клеток в наполненные кристаллином волокнистые клетки без органелл происходит в пределах капсулы хрусталика. Старые клетки не могут оторваться и вместо этого интернализуются ближе к центру хрусталика. Этот процесс приводит к полной временной многоуровневой записи процесса дифференцировки от начала на поверхности хрусталика до конца в центре хрусталика. Поэтому линза представляет ценность для ученых, изучающих процесс дифференцировки клеток. [21]
Различия в конструкции линз
[ редактировать ]
У многих водных позвоночных хрусталик значительно толще, почти сферический, что приводит к увеличению преломления света. Эта разница помогает компенсировать меньший угол преломления между роговицей глаза и водянистой средой, поскольку они имеют более схожие показатели преломления, чем роговица и воздух. [22] Волокнистые клетки рыб обычно значительно тоньше, чем у наземных позвоночных, и, по-видимому, белки-кристаллины транспортируются к свободным клеткам органелл на внешней стороне хрусталика к внутренним клеткам через многие слои клеток. [23] Некоторым позвоночным иногда необходимо хорошо видеть как над, так и под водой. Одним из примеров являются ныряющие птицы, способные менять фокус на 50–80 диоптрий. По сравнению с животными, приспособленными только к одной среде, ныряющие птицы имеют несколько измененную структуру хрусталика и роговицы с механизмами фокусировки, позволяющими работать в обеих средах. [24] [25] Даже среди наземных животных хрусталик приматов, таких как человек, необычно плоский, что в некоторой степени объясняет, почему наше зрение, в отличие от ныряющих птиц, особенно размыто под водой. [26]
Функция
[ редактировать ]Фокусировка
[ редактировать ]

У людей широко цитируемый механизм фокусировки Гельмгольца, также называемый аккомодацией , часто называют «моделью». [27] Прямое экспериментальное доказательство любой модели хрусталика обязательно затруднено, поскольку хрусталик позвоночных прозрачен и хорошо функционирует только у живых животных. При рассмотрении всех позвоночных аспекты всех моделей могут играть разную роль в фокусе линзы.
Хрусталик, меняющий форму, у многих наземных позвоночных.
[ редактировать ]Внешние силы
[ редактировать ]
Модель человеческой линзы, меняющей форму, была предложена Янгом в лекции 27 ноября 1800 года. [28] Другие, такие как Гельмгольц и Хаксли, усовершенствовали модель в середине 1800-х годов, объяснив, как сокращается цилиарная мышца, округляющая хрусталик, чтобы фокусироваться вблизи. [29] и эта модель была популяризирована Гельмгольцем в 1909 году. [30] [31] Модель можно резюмировать следующим образом. Обычно хрусталик удерживается под напряжением за счет того, что его подвешивающие связки туго натягиваются под давлением глазного яблока. На коротком фокусном расстоянии цилиарная мышца сокращается, снимая часть напряжения со связок, позволяя хрусталику немного эластично округляться, увеличивая преломляющую силу. Для изменения фокуса на объекте, находящемся на большем расстоянии, требуется более тонкая и менее изогнутая линза. Это достигается за счет расслабления некоторых сфинктеров, например цилиарных мышц. Хотя это и не упоминается, это, по-видимому, позволяет давлению в глазном яблоке снова расширить его наружу, сильнее натягивая хрусталик, делая его менее изогнутым и тоньше, тем самым увеличивая фокусное расстояние . Проблема с моделью Гельмгольца заключается в том, что, несмотря на опробование математических моделей, ни одна из них не приблизилась к работе с использованием только механизмов Гельмгольца. [32]

Шахар предложил модель наземных позвоночных, которая не получила одобрения. [33] Теория позволяет математическому моделированию более точно отразить способ фокусировки хрусталика, принимая во внимание сложность поддерживающих связок и наличие радиальных, а также круговых мышц в цилиарном теле. [34] [35] В этой модели связки могут в разной степени натягивать хрусталик на экваторе с помощью радиальных мышц, в то время как связки смещаются от экватора вперед и назад. [36] расслабляются в разной степени за счет сокращения круговых мышц. [37] Эти многочисленные действия [38] работа с эластичной линзой позволяет более тонко менять форму линзы спереди. Не только изменение фокуса, но и коррекция аберраций объектива, которые в противном случае могли бы возникнуть в результате изменения формы, и при этом лучше подходящая для математического моделирования. [32]
« Цепная » модель фокуса линзы, предложенная Коулманом. [39] требует меньшего напряжения связок, удерживающих хрусталик. Вместо того, чтобы хрусталик в целом растягивался тоньше для зрения вдаль и позволял расслабиться для фокусировки вблизи, сокращение круговых цилиарных мышц приводит к тому, что хрусталик испытывает меньшее гидростатическое давление на свою переднюю часть. Передняя часть хрусталика может затем изменить свою форму между поддерживающими связками, подобно тому, как провисшая цепь, висящая между двумя полюсами, может изменить свою кривую, когда полюса сближаются. Эта модель требует плавного движения только передней части линзы, а не пытается изменить форму линзы в целом.
Внутренние силы
[ редактировать ]

Когда в 1801 году Томас Янг предложил изменение формы хрусталика человека в качестве механизма фокальной аккомодации, он думал, что хрусталик может представлять собой мышцу, способную сокращаться. Этот тип модели называется внутрикапсулярной аккомодацией, поскольку он основан на активности внутри хрусталика. В Нобелевской лекции 1911 года Аллвар Гулстранд говорил на тему «Как я нашел внутрикапсулярный механизм аккомодации», и этот аспект фокусировки хрусталика продолжает исследоваться. [40] [41] [42] Янг потратил время на поиск нервов, которые могли бы стимулировать сокращение хрусталика, но безуспешно. С тех пор стало ясно, что хрусталик — это не простая мышца, стимулируемая нервом, поэтому модель Гельмгольца 1909 года взяла верх. Исследователи до двадцатого века не имели возможности воспользоваться многими более поздними открытиями и методами. Мембранные белки, такие как аквапорины , которые позволяют воде проникать в клетки и выходить из них, являются наиболее распространенным мембранным белком в хрусталике. [43] [44] Также широко распространены коннексины , которые обеспечивают электрическое соединение клеток. Электронная и иммунофлуоресцентная микроскопия показывают, что волокнистые клетки сильно различаются по структуре и составу. [45] [46] [47] Магнитно-резонансная томография подтверждает наличие слоев в хрусталике, которые могут обеспечивать различные планы рефракции внутри него. [48] Показатель преломления хрусталика человека варьируется примерно от 1,406 в центральных слоях до 1,386 в менее плотных слоях хрусталика. [49] Этот градиент индекса увеличивает оптическую силу линзы. становится известно о структуре хрусталика млекопитающих in situ По мере того, как благодаря фотографиям Шаймпфлюга , МРТ [50] [51] В результате физиологических исследований становится очевидным, что хрусталик сам по себе не реагирует полностью пассивно на окружающую цилиарную мышцу, но может быть способен изменять свой общий показатель преломления посредством механизмов, включающих динамику воды в хрусталике, которые еще предстоит выяснить. [52] [53] [54] На прилагаемой микрофотографии показаны сморщенные волокна расслабленной овечьей линзы после ее удаления из животного, что указывает на укорочение волокон хрусталика во время аккомодации вблизи фокуса. Возрастные изменения хрусталика человека также могут быть связаны с изменениями динамики воды в хрусталике. [55] [56]
Линзы птиц, рептилий, земноводных, рыб и др.
[ редактировать ]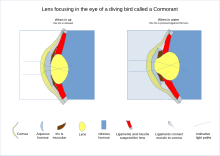
У рептилий и птиц цилиарное тело, поддерживающее хрусталик посредством поддерживающих связок, также соприкасается с хрусталиком множеством подушечек на его внутренней поверхности. Эти подушечки сжимают и отпускают линзу, изменяя ее форму при фокусировке на объектах на разных расстояниях; эту функцию обычно выполняют поддерживающие связки у млекопитающих . При зрении у рыб и амфибий форма хрусталика фиксируется, и вместо этого фокусировка достигается за счет перемещения хрусталика вперед или назад внутри глаза с помощью мышцы, называемой ретрактором линзы. [26]
У хрящевых рыб поддерживающие связки заменены мембраной, включающей небольшую мышцу на нижней стороне хрусталика. Эта мышца вытягивает хрусталик вперед из расслабленного положения при фокусировке на близлежащих объектах. У костистых , напротив, из сосудистой структуры на дне глаза выступает мышца, называемая серповидным отростком , и служит для оттягивания хрусталика назад из расслабленного положения для фокусировки на удаленных объектах. Хотя амфибии, как и хрящевые рыбы, перемещают хрусталик вперед, задействованные мышцы не одинаковы ни у одного из типов животных. У лягушек имеются две мышцы: одна над хрусталиком и одна под хрусталиком, тогда как у других земноводных имеется только нижняя мышца. [26]
У простейших позвоночных — миног и миксин — хрусталик вообще не прикрепляется к внешней поверхности глазного яблока. Водянистой влаги у этих рыб нет, и стекловидное тело просто прижимает хрусталик к поверхности роговицы. Чтобы сфокусировать взгляд, минога сглаживает роговицу с помощью мышц снаружи глаза и толкает хрусталик назад. [26]
Хотя это и не позвоночные, здесь кратко упоминается конвергентная эволюция глаз позвоночных и моллюсков . Самый сложный глаз моллюсков - это глаз головоногих моллюсков , который внешне похож по структуре и функциям на глаз позвоночных, включая аккомодацию, но отличается в основных чертах, таких как наличие двухчастной линзы и отсутствие роговицы. [57] [58] Фундаментальные требования оптики должны быть выполнены всеми глазами с помощью линз с использованием имеющихся в их распоряжении тканей, поэтому на первый взгляд все глаза имеют тенденцию выглядеть одинаково. Это способ удовлетворения оптических потребностей с использованием различных типов клеток и структурных механизмов, который варьируется у разных животных.
Кристаллины и прозрачность
[ редактировать ]
Кристаллины — это водорастворимые белки , которые составляют более 90% белка хрусталика. [59] Три основных типа кристаллинов, обнаруженных в человеческом глазу, — это α-, β- и γ-кристаллины. Кристаллины имеют тенденцию образовывать растворимые высокомолекулярные агрегаты, которые плотно упаковываются в волокна хрусталика, тем самым увеличивая показатель преломления хрусталика, сохраняя при этом его прозрачность. Кристаллины β и γ обнаруживаются преимущественно в хрусталике, тогда как субъединицы α-кристаллина изолированы из других частей глаза и тела. Белки α-кристаллины принадлежат к более крупному суперсемейству молекулярных белков-шаперонов , поэтому считается, что белки-кристаллины эволюционно рекрутировались из белков-шаперонов для оптических целей. [60] Шаперонные функции α-кристаллина могут также способствовать поддержанию белков хрусталика, которые должны сохраняться у человека на протяжении всей его жизни. [60]
светорассеивающих органелл, таких как ядро , эндоплазматическая сеть и митохондрии . Другим важным фактором поддержания прозрачности хрусталика является отсутствие в зрелых волокнах хрусталика [61] Волокна хрусталика также имеют очень обширный цитоскелет , который поддерживает точную форму и упаковку волокон хрусталика; нарушения/мутации в определенных элементах цитоскелета могут привести к потере прозрачности. [62]
Линза блокирует большую часть ультрафиолетового света в диапазоне длин волн 300–400 нм; более короткие волны блокируются роговицей. Пигментом, ответственным за блокировку света, является 3-гидроксикинуренин глюкозид, продукт катаболизма триптофана в эпителии хрусталика. [63] Ультрафиолетовый свет высокой интенсивности может повредить сетчатку, искусственные интраокулярные линзы , которые также блокируют ультрафиолет. поэтому изготавливаются [64] Люди, у которых нет хрусталика (состояние, известное как афакия ), воспринимают ультрафиолетовый свет как беловато-синий или беловато-фиолетовый. [65] [66]
Питание
[ редактировать ]Хрусталик метаболически активен и требует питания для поддержания своего роста и прозрачности. Однако по сравнению с другими тканями глаза хрусталик потребляет значительно меньше энергии. [67]
К девяти неделям развития человека хрусталик окружен и питается сетью сосудов — tunica vasculosa lentis , которая происходит из гиалоидной артерии . [17] Начиная с четвертого месяца развития, гиалоидная артерия и связанная с ней сосудистая сеть начинают атрофироваться и к рождению полностью исчезают. [68] В послеродовом глазу канал Клоке отмечает прежнее расположение гиалоидной артерии.

После регрессии гиалоидной артерии хрусталик получает все питание за счет водянистой влаги. Питательные вещества диффундируют внутрь, а отходы — наружу посредством постоянного потока жидкости от переднего/заднего полюсов хрусталика и из экваториальных областей. Эту динамику поддерживает Na. + /К + -АТФазные насосы расположены в экваториально расположенных клетках эпителия хрусталика. [9] Взаимодействие этих насосов с водными каналами в клетках, называемыми аквапоринами, молекулами размером менее 100 дальтон между клетками через щелевые соединения и кальцием с помощью транспортеров/регуляторов (каналы TRPV) приводит к потоку питательных веществ через хрусталик. [69] [70]
Глюкоза является основным источником энергии для хрусталика. Поскольку зрелые волокна хрусталика не имеют митохондрий , примерно 80% глюкозы метаболизируется посредством анаэробного метаболизма . [71] Оставшаяся часть глюкозы направляется преимущественно по пентозофосфатному пути . [71] Отсутствие аэробного дыхания означает, что хрусталик потребляет очень мало кислорода. [71]
Клиническое значение
[ редактировать ]- Катаракта – это помутнение хрусталика. Некоторые из них малы и не требуют никакого лечения, другие могут быть достаточно большими, чтобы блокировать свет и ухудшать зрение. Катаракта обычно развивается по мере того, как стареющий хрусталик становится все более непрозрачным, но катаракта также может образоваться врожденно или после повреждения хрусталика. Ядерный склероз — это разновидность возрастной катаракты. Диабет является еще одним фактором риска развития катаракты. Операция по удалению катаракты включает удаление хрусталика и установку искусственной интраокулярной линзы.
- Пресбиопия – это возрастная потеря аккомодации, характеризующаяся неспособностью глаза фокусироваться на близлежащих объектах. Точный механизм до сих пор неизвестен, но возрастные изменения твердости, формы и размера хрусталика связаны с этим заболеванием.
- Эктопия хрусталика – это смещение хрусталика от нормального положения.
- Афакия – отсутствие хрусталика в глазу. Афакия может быть результатом хирургического вмешательства или травмы, а может быть врожденной.
Дополнительные изображения
[ редактировать ] МРТ человеческого глаза, показывающая линзу.
МРТ человеческого глаза, показывающая линзу.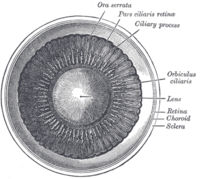 Внутренняя часть передней камеры глаза.
Внутренняя часть передней камеры глаза. Хрусталик затвердел и разделился.
Хрусталик затвердел и разделился. Разрез края хрусталика, показывающий переход эпителия в волокна хрусталика, известный как дуговая область.
Разрез края хрусталика, показывающий переход эпителия в волокна хрусталика, известный как дуговая область.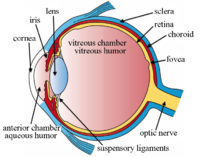 Структуры глаза, обозначенные
Структуры глаза, обозначенные Другой вид глаза и его структур с надписью
Другой вид глаза и его структур с надписью![Этот svg-файл был настроен так, чтобы лучи, диафрагма и хрусталик можно было легко изменить[72]](//upload.wikimedia.org/wikipedia/commons/thumb/5/5e/Editable_ray_diagram_of_eye_v0.svg/200px-Editable_ray_diagram_of_eye_v0.svg.png) Этот svg-файл был настроен так, чтобы лучи, диафрагма и хрусталик можно было легко изменить. [72]
Этот svg-файл был настроен так, чтобы лучи, диафрагма и хрусталик можно было легко изменить. [72]





![Этот svg-файл был настроен так, чтобы лучи, диафрагма и хрусталик можно было легко изменить[72]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/5e/Editable_ray_diagram_of_eye_v0.svg/200px-Editable_ray_diagram_of_eye_v0.svg.png)
См. также
[ редактировать ]- Рефлекс аккомодации
- Кристаллин
- Эволюция глаза : как развивался хрусталик
- Интраокулярные линзы
- Ирис
- Капсула линзы
- Факоэмульсификация
- Визуальное восприятие
- Зонулы из Зинна
Ссылки
[ редактировать ]- ^ Басснетт, Стивен (май 2021 г.). «Зонула Зинна» . Прогресс в исследованиях сетчатки и глаз . 82 : 100902. doi : 10.1016/j.preteyeres.2020.100902 . ПМК 8139560 . ПМИД 32980533 .
- ^ "Экватор линзы - определение из" . Биология-Online.org. Архивировано из оригинала 22 марта 2012 г. Проверено 25 ноября 2012 г.
- ^ «Эватор хрусталика - определение экватора хрусталика в Медицинском словаре - в Бесплатном онлайн-медицинском словаре, тезаурусе и энциклопедии» . Medical-dictionary.thefreedictionary.com . Проверено 25 ноября 2012 г.
- ^ Jump up to: а б с д и Джон Форрестер, Эндрю Дик, Пол МакМенамин, Уильям Ли (1996). Глаз: фундаментальные науки на практике . Лондон: WB Saunders Company Ltd., с. 28 ISBN 0-7020-1790-6
- ^ Янофф, Мирон; Дукер, Джей С.; Аугсбургер, Джеймс Дж. (2008). Офтальмология (3-е изд.). Эдинбург: Мосби. п. 382. ИСБН 978-0323057516 .
- ^ Мохан, PS; Спиро, Р.Г. (25 марта 1986 г.). «Макромолекулярная организация базальных мембран. Характеристика и сравнение компонентов клубочковой базальной мембраны и капсулы хрусталика с помощью иммунохимических процедур и процедур сродства к лектинам» . Журнал биологической химии . 261 (9): 4328–36. дои : 10.1016/S0021-9258(17)35665-X . ПМИД 3512568 .
- ^ Jump up to: а б Грюйтерс, Wouterus TM (1 июля 2024 г.). «Новая структура глазного хрусталика, связанная с ростом капсулы / базальной мембраны». Микропубликация по биологии . doi : 10.17912/micropub.biology.000828 .
- ^ Ши, Янжун; Ту, Идун; Де Мария, Алисия; Мечам, Роберт П.; Басснетт, Стивен (1 апреля 2013 г.). «Развитие, состав и структурное устройство цилиарной зоны мыши» . Исследовательская офтальмология и визуальные науки . 54 (4): 2504–2515. дои : 10.1167/iovs.13-11619 . ПМЦ 3621578 . ПМИД 23493297 .
- ^ Jump up to: а б Кандия, Оскар А. (2004). «Перенос электролитов и жидкости через эпителий роговицы, конъюнктивы и хрусталика». Экспериментальное исследование глаз . 78 (3): 527–535. дои : 10.1016/j.exer.2003.08.015 . ПМИД 15106931 .
- ^ Jump up to: а б Ши, Янжун; Бартон, Келли; Де Мария, Алисия; Петраш, Дж. Марк; Шилс, Алан; Басснетт, Стивен (15 мая 2009 г.). «Многослойный синцитий хрусталика позвоночных» . Журнал клеточной науки . 122 (10): 1607–1615. дои : 10.1242/jcs.045203 . ПМК 2680101 . ПМИД 19401333 .
- ^ Jump up to: а б «глаз, человек». Британская энциклопедия из Британской энциклопедии 2006 г., DVD Ultimate Reference Suite 2009 г.
- ^ Он, Миньян; Чжоу, Гуанкай; Линь, Цинхун; Чжоу, Нань (сентябрь 2023 г.). «Роль mip в развитии хрусталика у рыбок данио» . Паттерны экспрессии генов . 49 : 119330. doi : 10.1016/j.gep.2023.119330 . ПМИД 37369320 .
- ^ Пайди, Сантош Кумар; Чжан, Циньжун; Ян, Юхан; Ся, Чун-Хонг; Джи, На; Гун, Сяохуа (19 января 2023 г.). «Адаптивная оптическая двухфотонная флуоресцентная микроскопия исследует клеточную организацию хрусталика глаза in vivo». bioRxiv 10.1101/2023.01.17.524320 .
- ^ Митчелл, ПК (апрель 1891 г.). «Двойной куриный эмбрион» . Журнал анатомии и физиологии . 25 (Часть 3): 316–324,1. ПМЦ 1328169 . ПМИД 17231922 .
- ^ Чаухан, Б; Плагеман, Т; Лу, М; Ланг, Р. (2015). «Эпителиальный морфогенез: глаз мыши как модельная система» . Актуальные темы биологии развития . 111 : 375–99. дои : 10.1016/bs.ctdb.2014.11.011 . ПМК 6014593 . ПМИД 25662266 .
- ^ Jump up to: а б Муччиоли, Мария; Кайси, Даля; Герман, Кен; Плагеман, Тимоти Ф. (апрель 2016 г.). «Полярность плоских клеток хрустальной плакоды зависит от Cdc42-опосредованного ингибирования сокращения соединений» . Биология развития . 412 (1): 32–43. дои : 10.1016/j.ydbio.2016.02.016 . ПМЦ 7370377 . ПМИД 26902112 .
- ^ Jump up to: а б Глаз: фундаментальные науки на практике , с. 102, ISBN 0-7020-1790-6
- ^ Маддала, Рупалата; Чаухан, Бхареш К.; Уокер, Кристофер; Чжэн, И; Робинсон, Майкл Л.; Ланг, Ричард А.; Рао, Понуготи В. (декабрь 2011 г.). «Хрусталик мыши с дефицитом Rac1 GTPase демонстрирует дефекты формы, формирования швов, миграции и выживания волоконных клеток» . Биология развития . 360 (1): 30–43. дои : 10.1016/j.ydbio.2011.09.004 . ПМЦ 3215831 . ПМИД 21945075 .
- ^ Цвекл, А.; Эшери-Падан, Р. (2014). «Клеточные и молекулярные механизмы развития хрусталика позвоночных» . Разработка . 141 (23): 4432–4447. дои : 10.1242/dev.107953 . ПМК 4302924 . ПМИД 25406393 .
- ^ Канту, Клаудио; Циммерли, Дарио; Хаусманн, Джордж; Валента, Томас; Мавр, Андреас; Аге, Мишель; Баслер, Конрад (2014). «Pax6-зависимая, но β-катенин-независимая функция белков Bcl9 в развитии хрусталика мыши» . Гены и развитие . 28 (17): 1879–1884. дои : 10.1101/gad.246140.114 . ПМК 4197948 . ПМИД 25184676 .
- ^ Лими, Сайма; Сенекаль, Адриан; Коулман, Роберт; Лопес-Джонс, Мелисса; Го, Пэн; Полумбо, Кристина; Певец, Роберт Х.; Скулчи, Артур И.; Цвекл, Алесь (август 2018 г.). «Динамика фракции и размера транскрипционного взрыва во время дифференцировки клеток волокон хрусталика и детальное понимание процесса денуклеации» . Журнал биологической химии . 293 (34): 13176–13190. дои : 10.1074/jbc.RA118.001927 . ПМК 6109918 . ПМИД 29959226 .
- ^ Кардонг, К. (2008). Позвоночные животные: Сравнительная анатомия, функции, эволюция (5-е изд.). (стр. 676–677). Бостон: МакГроу-Хилл
- ^ Козловский, Томаш М.; Крегер, Рональд Х.Х. (сентябрь 2019 г.). «Постоянная толщина клеток хрусталика у рыб предполагает транспорт кристаллина к лишенным ядра клеткам» . Исследование зрения . 162 : 29–34. doi : 10.1016/j.visres.2019.06.008 . ПМИД 31278970 . S2CID 195820065 .
- ^ Кацир, Гади; Хауленд, Ховард К. (1 марта 2003 г.). «Сила роговицы и аккомодация под водой у больших бакланов (Phalacrocorax carbo sinensis)». Журнал экспериментальной биологии . 206 (5): 833–841. дои : 10.1242/jeb.00142 . ПМИД 12547938 . S2CID 3096767 .
- ^ Сивак, Дж.Г.; Хильдебранд, Т.; Леберт, К. (январь 1985 г.). «Величина и скорость аккомодации у ныряющих и неныряющих птиц». Исследование зрения . 25 (7): 925–933. дои : 10.1016/0042-6989(85)90203-2 . ПМИД 4049742 . S2CID 42368520 .
- ^ Jump up to: а б с д Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 463–464. ISBN 978-0-03-910284-5 .
- ^ Лэнд, Майкл (19 апреля 2015 г.). «Фокусировка путем изменения формы хрусталика глаза: комментарий к книге Янга (1801) «О механизме глаза» » . Философские труды Королевского общества B: Биологические науки . 370 : 20140308.doi : (1666 ) 10.1098/rstb.2014.0308 . ПМК 4360117 . ПМИД 25750232 .
- ^ Лэнд, М (19 апреля 2015 г.). «Фокусировка путем изменения формы хрусталика глаза: комментарий к книге Янга (1801) «О механизме глаза» » . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 370 (1666). дои : 10.1098/rstb.2014.0308 . ПМК 4360117 . ПМИД 25750232 .
- ^ Хаксли, Томас Х (1871). Уроки элементарной физиологии (5-е изд.). Лондон и Нью-Йорк: МАКМИЛЛАН И КО., стр. 256–258.
- ^ Гельмгольц, Х. фон (1962). Трактат по физиологической оптике (английский перевод под редакцией JPC Southall. Оптическое общество Америки. Из третьего немецкого издания Handbuch der Physiologischen Optik (1909), Леопольд Восс, Лейпциг. Переиздание Дувра). Нью-Йорк, штат Нью-Йорк: Dover Publications Inc.
- ^ Фишер, РФ (1 августа 1977 г.). «Сила сокращения цилиарной мышцы человека при аккомодации» . Журнал физиологии . 270 (1): 51–74. дои : 10.1113/jphysicalol.1977.sp011938 . ПМЦ 1353417 . ПМИД 915798 .
- ^ Jump up to: а б Шачар, Рональд А.; Бакс, Эндрю Дж. (июнь 2001 г.). «Механизм аккомодации человека, анализируемый с помощью нелинейного анализа методом конечных элементов». Комплексная терапия . 27 (2): 122–132. дои : 10.1007/s12019-996-0006-5 . ПМИД 11430259 . S2CID 71369369 .
- ^ Атчисон, Д.А. (июль 1995 г.). «Аккомодация и пресбиопия». Офтальмологическая и физиологическая оптика . 15 (4): 255–72. doi : 10.1046/j.1475-1313.1995.9500020e.x . ПМИД 7667018 . S2CID 24282106 .
- ^ Мэйсяо; Сюй, Чжэ; Карп, Ван, Цзяньхуа (1 июня 2015 г.). Шао, Илей; Тао, Цзян, Хун; Чжун, Цзянгуан , Связанные изменения в биометрии переднего сегмента во время аккомодации» . Investigative Ophthalmology & Visual Science . 56 (6): 3522–3530. : 10.1167 /iovs.15-16825 . PMC 4464043. doi PMID 26030106 .
- ^ Шачар, Рональд А. (22 сентября 2015 г.). «Изменения конфигурации аккомодационных цилиарных мышц человека соответствуют механизму аккомодации Шахара». Исследовательская офтальмология и визуальные науки . 56 (10): 6075. doi : 10.1167/iovs.15-17452 . ПМИД 26393665 .
- ^ Стритен, BW (1977). «Б.В. Стритен; Зональная вставка: исследование сканирующей электронной микроскопии». Инвестируйте. Офтальмол. Вис. Наука . 16 (4): 364–375.
- ^ Шачар, РА (март 1994 г.). «Зонулярная функция: новая гипотеза с клиническим значением». Анналы офтальмологии . 26 (2): 36–8. ПМИД 8010701 .
- ^ Кнаус, Кэтрин Р.; Хипсли, ЭннМари; Блемкер, Сильвия С. (июнь 2021 г.). «Влияние сокращения цилиарной мышцы на аккомодацию хрусталика, исследованное с помощью 3D-модели». Биомеханика и моделирование в механобиологии . 20 (3): 879–894. дои : 10.1007/s10237-021-01417-9 . ПМИД 33491156 . S2CID 231704221 .
- ^ Коулман, Д. Джексон (июнь 1970 г.). «Единая модель аккомодационного механизма». Американский журнал офтальмологии . 69 (6): 1063–1079. дои : 10.1016/0002-9394(70)91057-3 . ПМИД 5423772 .
- ^ ПАУ, Ч (1952). «[Аккомодационное смещение ядра хрусталика при внутрикапсулярной аккомодации]». Клинические ежемесячные информационные бюллетени по офтальмологии и офтальмологическому обучению . 121 (2): 224–6. ПМИД 14955961 .
- ^ Хуггерт, Арне (27 мая 2009 г.). «Внутрикапсулярный механизм аккомодации». Акта офтальмологическая . 42 (2): 389–397. дои : 10.1111/j.1755-3768.1964.tb03627.x . ПМИД 14213923 . S2CID 37325357 .
- ^ Лопес-Хиль, Норберто (3 марта 2022 г.). «Пересмотр механизма внутрикапсульного размещения Галлстранда» . Фотоника . 9 (3): 152. Бибкод : 2022Фото...9..152Л . doi : 10.3390/photonics9030152 .
- ^ Броехейсе, РМ; Кульманн, Эд; Столс, А.Л. (сентябрь 1976 г.). «Мембраны хрусталика II. Выделение и характеристика основного внутреннего полипептида (MIP) мембран бычьих волокон хрусталика». Экспериментальное исследование глаз . 23 (3): 365–371. дои : 10.1016/0014-4835(76)90135-4 . ПМИД 976377 .
- ^ Малдерс, С.М.; Престон, генеральный менеджер; Дин, премьер-министр; Гуджино, ВБ; ван Ос, Швейцария; Агре, П. (14 апреля 1995 г.). «Свойства водного канала основного внутреннего белка хрусталика» . Журнал биологической химии . 270 (15): 9010–16. дои : 10.1074/jbc.270.15.9010 . hdl : 2066/216433 . ПМИД 7536742 .
- ^ Кушак, Дж; Алькала, Дж; Мейзел, Х. (декабрь 1980 г.). «Поверхностная морфология клеток хрусталика эмбрионов и взрослых кур». Американский журнал анатомии . 159 (4): 395–410. дои : 10.1002/aja.1001590406 . ПМИД 7223675 .
- ^ Грюйтерс, WT; Кистлер, Дж; Булливант, С. (октябрь 1987 г.). «Формирование, распределение и диссоциация межклеточных соединений в хрусталике». Журнал клеточной науки . 88 (Часть 3) (3): 351–9. дои : 10.1242/jcs.88.3.351 . ПМИД 3448099 .
- ^ Грюйтерс, WT (июль 1989 г.). «Белок, не являющийся коннексоном (MIP), участвует в формировании щелевых соединений хрусталика». Журнал клеточной науки . 93 (Часть 3) (3): 509–13. дои : 10.1242/jcs.93.3.509 . ПМИД 2691517 .
- ^ Германс, Э.А.; Дуббельман, М; Ван дер Хейде, Р.; Хитаар, РМ (декабрь 2008 г.). «Эквивалентный показатель преломления хрусталика человека при аккомодационной реакции». Оптометрия и наука о зрении . 85 (12): 1179–84. дои : 10.1097/OPX.0b013e31818e8d57 . ПМИД 19050472 . S2CID 205907383 .
- ^ Хехт, Юджин. Оптика , 2-е изд. (1987), Эддисон Уэсли, ISBN 0-201-11609-X . п. 178.
- ^ Херманс, Эрик А.; Пауэлс, Петра Дж.В.; Дуббельман, Мишель; Куйер, Йост, Пенсильвания; ван дер Хейде, Роб Г.Л.; Хитаар, Роб М. (1 января 2009 г.). «Постоянный объем хрусталика человека и уменьшение площади поверхности капсульного мешка во время аккомодации: исследование МРТ и Шаймпфлюга» . Исследовательская офтальмология и визуальные науки . 50 (1): 281–289. дои : 10.1167/iovs.08-2124 . ПМИД 18676625 .
- ^ Станке, Т.; Хадлич, С.; Ври, А.; Гутхофф, Р.; Стахс, О.; Лангнер, С. (16 декабря 2016 г.). «Магнитно-резонансная микроскопия аппарата аккомодации». Клинические ежемесячные бюллетени по офтальмологии . 233 (12): 1320–1323. дои : 10.1055/s-0042-118599 . ПМИД 27984837 . S2CID 78808282 .
- ^ Вагефи, Э; Понтре, BP; Джейкобс, доктор медицины; Дональдсон, П.Дж. (август 2011 г.). «Визуализация динамики жидкости в хрусталике глаза с помощью МРТ: манипулирование стабильным содержанием воды и потоками воды». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 301 (2): Р335-42. дои : 10.1152/ajpregu.00173.2011 . ПМИД 21593426 . S2CID 9525037 .
- ^ Дональдсон, Пол Дж.; Чен, Яди; Петрова, Росица С.; Грей, Ангус К.; Лим, Джули К. (декабрь 2022 г.). «Регулирование содержания воды в хрусталике: влияние на физиологическую оптику хрусталика». Прогресс в исследованиях сетчатки и глаз . 95 : 101152. doi : 10.1016/j.preteyeres.2022.101152 . ПМИД 36470825 . S2CID 254243790 .
- ^ Сузуки-Керр, Х; Уокер, КЛ; Хан, Миннесота; Лим, Джей Си; Дональдсон, Пи Джей (2022). «Гипосмотический стресс вызывает высвобождение АТФ в отдельной зоне внешней коры хрусталика крысы» . Молекулярное видение . 28 : 245–256. ПМЦ 9514545 . ПМИД 36284672 .
- ^ Моффат, бакалавр; Ландман, штат Калифорния; Траскотт, Р.Дж.; Суини, Миннесота; Поуп, Дж. М. (декабрь 1999 г.). «Возрастные изменения кинетики транспорта воды в хрусталике нормального человека». Экспериментальное исследование глаз . 69 (6): 663–9. дои : 10.1006/exer.1999.0747 . ПМИД 10620395 .
- ^ Джонс, CE; Атчисон, округ Колумбия; Медер, Р.; Поуп, Дж. М. (август 2005 г.). «Распределение показателя преломления и оптические свойства изолированного хрусталика человека, измеренные с помощью магнитно-резонансной томографии (МРТ)». Исследование зрения . 45 (18): 2352–2366. дои : 10.1016/j.visres.2005.03.008 . ПМИД 15979462 . S2CID 8894700 .
- ^ Джаггер, WS; Сэндс, П.Дж. (1 августа 1999 г.). «Широкоугольная градиентно-индексная оптическая модель хрусталика и глаза осьминога». Исследование зрения . 39 (17): 2841–2852. дои : 10.1016/S0042-6989(99)00012-7 . ПМИД 10492814 . S2CID 17808919 .
- ^ Шеффель, Ф.; Мерфи, CJ; Хауленд, ХК (15 ноября 1999 г.). «Размещение у каракатицы (Sepia officinalis)» . Журнал экспериментальной биологии . 202 (22): 3127–3134. дои : 10.1242/jeb.202.22.3127 . ПМИД 10539961 .
- ^ Хоэнвартер, В.; Клозе, Дж.; Юнгблут, PR (2006). «Протеомика хрусталика глаза». Аминокислоты . 30 (4): 369–389. дои : 10.1007/s00726-005-0283-9 . ПМИД 16583312 . S2CID 19978371 .
- ^ Jump up to: а б Эндли, Уша П. (2007). «Кристаллины в глазу: функция и патология». Прогресс в исследованиях сетчатки и глаз . 26 (1): 78–98. doi : 10.1016/j.preteyeres.2006.10.003 . ПМИД 17166758 . S2CID 29317220 .
- ^ Ланг, Ричард А. (январь 1997 г.). «Апоптоз в развитии глаз млекопитающих: морфогенез хрусталика, сосудистая регрессия и иммунная привилегия» . Смерть клеток и дифференциация . 4 (1): 12–20. дои : 10.1038/sj.cdd.4400211 . ПМИД 16465205 .
- ^ Блумендаль, Ганс; Де Йонг, Уилфрид; Йенике, Райнер; Лубсен, Николетт Х.; Слингсби, Кристина; Тардье, Аннет (2004). «Старение и зрение: структура, стабильность и функция кристаллинов хрусталика» . Прогресс биофизики и молекулярной биологии . 86 (3): 407–485. doi : 10.1016/j.pbiomolbio.2003.11.012 . ПМИД 15302206 .
- ^ Эндрю М. Вуд и Роджер Дж. В. Траскотт (март 1993 г.). «УФ-фильтры в человеческих линзах: катаболизм триптофана». Экспериментальное исследование глаз . 56 (3): 317–325. дои : 10.1006/exer.1993.1041 . ПМИД 8472787 .
- ^ Мейнстер, Массачусетс (2006). «Фиолетовый и синий свет, блокирующий интраокулярные линзы: фотозащита и фоторецепция» . Британский журнал офтальмологии . 90 (6): 784–792. дои : 10.1136/bjo.2005.086553 . ПМК 1860240 . ПМИД 16714268 .
- ^ Андерсон, Роберт М. (1983). «Визуальное восприятие и наблюдения афакического хирурга». Перцептивные и моторные навыки . 57 (3_приложение): 1211–1218. дои : 10.2466/pms.1983.57.3f.1211 . ПМИД 6664798 . S2CID 20005737 .
- ^ Хэмблинг, Дэвид (29 мая 2002 г.). «Пусть свет сияет» . Хранитель .
- ^ Уайкхарт, Дэвид Р. (2003). Биохимия глаза, 2-е изд. 2003. Филадельфия: Баттерворт Хайнеманн, с. 107–8 ISBN 0-7506-7152-1
- ^ Глаз: фундаментальные науки на практике, с. 104, ISBN 0-7020-1790-6
- ^ Джанноне, Адриенн А.; Ли, Лепинг; Селитто, Катерина; Уайт, Томас В. (23 декабря 2021 г.). «Физиологические механизмы, регулирующие транспорт хрусталика» . Границы в физиологии . 12 : 818649. doi : 10.3389/fphys.2021.818649 . ПМЦ 8735835 . ПМИД 35002784 .
- ^ Деламер, Николас А.; Шахидулла, Мохаммед (31 января 2022 г.). «Регуляция транспорта ионов с помощью TRPV4 и TRPV1 в хрусталике и мерцательном эпителии» . Границы в физиологии . 12 : 834916. doi : 10.3389/fphys.2021.834916 . ПМЦ 8841554 . ПМИД 35173627 .
- ^ Jump up to: а б с Биохимия глаза , 2-е изд., с. 107–8, ISBN 0-7506-7152-1
- ^ Загрузите и откройте с помощью Inkscape 9.1. Отдельные компоненты располагаются на разных «слоях», что облегчает редактирование.
Внешние ссылки
[ редактировать ]- Гистологическое изображение: 08001loa - Система обучения гистологии в Бостонском университете.
