Heterodontosauridae
| Гетеродонтозавриды Временный диапазон: ранняя юрская величина - ранняя меловая , Возможная поздняя запись
| |
|---|---|

| |
| Актеры образца SAM-PK-K1332 из Heterodontosaurus Tucki , Музей палеонтологии Университета Калифорнийского университета | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Динозаурия |
| Клада : | † Ornithischia |
| Клада : | † Saphornithischia |
| Семья: | † heterodontosauridae Romer , 1966 (Kuhn, 1966 ) |
| Подгруппы [ 1 ] | |
| |
Heterodontosauridae - это семейство динозавров орнитишских , которые, вероятно, были одними из самых базальных (примитивных) членов группы. Их филогенетическое размещение является неопределенным, но чаще всего их можно найти примитивным за пределами группы Genasauria . [ 2 ] Хотя их окаменелости являются относительно редкими, а их группа не мала, они были обнаружены на всех континентах, кроме Австралии и Антарктиды , с диапазоном, охватывающей раннюю юридическую больную до раннего мелового .
Гетеродонтозавриды были динозаврами размером с лисы менее 2 метров (6,6 фута), включая длинный хвост. Они известны в основном своими характерными зубами, включая увеличенные клыки, похожие на собаки , и щечные зубы, адаптированные для жевания, аналогичные тем, что у мелородных хадрозавридов . Их диета была травоядным или, возможно, всеядным .
Описание
[ редактировать ]

Среди гетеродонтозавридов только сам гетеродонтозавр известен из полного скелета. Фрагментарные скелетные остатки аберктозавра известны, но не были полностью описаны, в то время как большинство других гетеродонтозавридов известны только из фрагментов челюсти и зубов. Следовательно, большинство синапоморфий гетеродонтозавридов (определяющие особенности) были описаны из костей зубов и челюсти. [ 3 ] [ 4 ] Гетеродонтозавр измерял чуть более 1 метра (3,3 фута) в длину, [ 5 ] в то время как фрагментарные остатки ликорхина могут указывать на более крупного человека. [ 6 ]
Tianyuulong из Китая, по-видимому, сохраняет нитевидное целое, который был интерпретирован как вариант прото-пегенчков, обнаруженных в некоторых тероподах. Эти нити включают гребень вдоль его хвоста. Присутствие этого нитчатого покровного положения использовалось, чтобы предположить, что как орнитишки, так и сауришцы были эндотермическими . [ 7 ]
Череп и зубы
[ редактировать ]У Abrictosaurus и Heterodontosaurus были очень большие глаза . Под глазами Jugal Bone проецировалась вбок, а также особенность, также присутствующая в цератопсах . Как и в челюстях большинства орнитишчианцев, передний край предчезгиллы кератиновый (кость на кончике верхней челюсти) был беззубым и, вероятно, поддержал клюв ( рампхотека ), хотя у гетеродонтозавридов были зубы в заднем участке премаксилья. Большой промежуток, называемый диастемой , отделял эти предчелюстные зубы от зубов верхней челюсти (главная верхняя челюстная кость) во многих орнитишайцах, но эта диастема была характерна для гетеродонтозавридов. Манкюль . (нижняя челюсть) была опрокинута предшественником , костью, уникальной для орнитишчианцев Эта кость также поддерживала клюв, похожий на тот, который был найден на предчецилле . Все зубы в нижней челюсти были обнаружены на зубной кости. [ 3 ]

Гетеродонтозавриды названы в честь их сильно гетеродонтного зубного зуба . Там было три предчелюстных зуба. В ранней юрской игре Абриктозавр , Гетеродонтозавр и Ликорхин первые два предчелюстных зуба были небольшими и коническими, в то время как гораздо более крупный третий зуб напоминал клыков карниворских млекопитающих и часто называют канинообразными или «туск». Нижняя канихообразная, больше, чем верхняя, заняла первое положение в зубном состоянии и была приспособлена арочной диастемой верхней челюсти, когда рот был закрыт. [ 3 ] Эти канины были зазубрены как на переднем, так и на заднем краях у гетеродонтозавра и ликорхина , в то время как у абрактозавра были зубцы только на переднем крае. [ 8 ] [ 9 ] В раннемеловом эхинодоне , возможно, было две верхние канины, которые были на верхней челюсти, а не на предчезгилле, [ 10 ] и фрукты из позднего юрского периода, возможно, имели две более низкие канины на каждой зубной зубе. [ 11 ] [ 12 ]

Как и характерные клыки, щечные зубы полученных гетеродонтозавридов также были уникальными среди ранних орнитишхов. Небольшие хребты или зубные зубцы выровняли края орнитишских щечных зубов для обрезки растительности. Эти зубцы простираются только на треть пути вниз по зубной короне от кончика во всех гетеродонтозавридах; У других орнитишхов зубцы распространяются дальше в сторону корня. Базальные формы, такие как Абриктозавр, имели щечные зубы как в верхней челюсти, так и в зубном состоянии, которые, как правило, были похожи на другие орнитишки: широко расставленные, каждый из которых имел низкую корону и сильно развитый хребет (пояс), отделяющий корону от корня. В более производительных формах, таких как Lycorhinus и Heterodontosaurus , зубы были в обработке зубчатой доли, с гораздо более высокими коронками и отсутствием cingula, так что не было никакой разницы в ширине между коронами и корнями. [ 3 ]
Эти полученные щечные зубы перекрывались, так что их короны образовали непрерывную поверхность, на которой можно разжечь пищу. Зубные ряды были слегка вставлены со стороны рта, оставляя пространство за зубами, которое могло быть ограничено мускулистой щекой , которая была бы необходима для жевания. Хадрозавры и цератопсианцы мела, а также многие травоядные млекопитающие, сходящимися в виду , будут развиваться несколько аналогичные стоматологические батареи . В отличие от адрозавров, которые постоянно заменяли сотни зубов, замена зубов в гетеродонтозавридах происходила гораздо медленнее, и несколько образцов были обнаружены без одного заменного зуба в ожидании. Характерно, что у гетеродонтозавридов не хватало небольших отверстий (отверстия) на внутренней стороне костей челюсти, которые, как считается, помогали в развитии зубов у большинства других орнитишхов. Гетеродонтозавриды также имели уникальный сфероидальный сустав между зубными зубцами и предшественником, что позволило нижним челюстям вращаться наружу, когда рот был закрыт, шлифовав зубы щека друг к другу. Из -за медленной скорости замены это шлифовальное масштаб производило экстремальный износ зубов, который обычно уничтожил большую часть зубных зубьев у более старых зубов, хотя повышенная высота крон дала каждому зубу долгий срок службы. [ 13 ]
Скелет
[ редактировать ]
Посткраниальная анатомия Heterodontosaurus Tucki была хорошо описана, хотя H. Tucki , как правило, считается наиболее полученными из ранних гетеродонтозавридов, поэтому невозможно узнать, сколько его особенностей было разделено с другими видами. [ 3 ] Передатки были длинными для динозавра, более 70% длины задних конечностей. Хорошо развитый дельтопекторный гребень (гребень для прикрепления мышц грудной клетки и плеч ) плечевой кости и заметный процесс олекранона где были прикреплены мышцы, которые расширяют предплечье ), ( указывают на то, что передняя конечность также была мощной. было пять цифр На Manus («рука»). Первый был большим, охваченным с резким изогнутым когтиком, и он вращался внутрь при сгибке; Роберт Баккер назвал это «Twist-Thumb». [ 14 ] Вторая цифра была самой длинной, немного длиннее третьей. Обе эти цифры носили когти, в то время как когтя Четвертые и пятые цифры были очень маленькими и простыми по сравнению. В задних конечностях голени была на 30% длиннее бедренной кости , которая обычно считается адаптацией для скорости. Цвета и малоберцовая кость нижней ноги были слиты с астрагалом и кальциной лодыжки, образуя « тибиофиофиотарс » с современными птицами . Также аналогично птицам, нижние кости нижних лапков кости и плюсневые (лодыжка) были слиты, чтобы сформировать « tarsometatarsus ». есть четыре цифры В PES (задних цветов), только второй, третий и четвертый контакт с землей. Хвост, в отличие от многих других орнитишхов, не имел окостененных сухожилий для поддержания жесткой осанки и, вероятно, был гибким. [ 5 ] Фрагментарный скелет, известный своим абиктозавром, никогда не был полностью описан, хотя передние мыши и маны были меньше, чем у гетеродонтозавра . Кроме того, четвертая и пятая цифра передней основы каждая равен по одной кости фаланги . [ 15 ]
Классификация
[ редактировать ]
Южноафриканский палеонтолог Роберт Брум создал название Geranosaurus в 1911 году для костей челюсти динозавров, пропускающих все зубы и некоторые частичные связанные с ними кости конечностей. [ 16 ] В 1924 году был назван Ликорхин и классифицирован как Cynodont , Сидни Хаутон . [ 17 ] Heterodontosaurus был назван в 1962 году, и IT, Lycorhinus и Geranosaurus были признаны тесно связанными орнитишскими динозаврами. [ 18 ] Альфред Ромер назвал Heterodontosauridae в 1966 году в качестве семейства орнитишских динозавров, включая Heterodontosaurus и Lycorhinus . [ 19 ] Кун независимо предложил Heterodontosauridae в том же году и иногда называется его главным автором. [ 20 ] Он был определен как клада в 1998 году Полом Серено [ 21 ] и переопределенный им в 2005 году как клада стебля, состоящая из гетеродонтозавра и всех видов, более тесно связанных с гетеродонтозавром , чем с парасауролофусом Уокери , Пахицефалозавром Вайомингенсис , Triceratops Horridus или Ankylosaurus Magniventris . [ 22 ] Heterodontosaurinae-это таксон, основанный на стебле, впервые определяемый филогенетически, Пол Серено в 2012 году как «самая инклюзивная клада, содержащая гетеродонтозавра Tuki , но не Tianyulong Confuciusi , Fruitadens Haagarorum , Echinodon Beclesii ». [ 1 ]
Heterodontosauridae включает в себя роды Abrictosaurus , Lycorhinus и Heterodontosaurus , все из Южной Африки. В то время как Ричард Тулборн однажды переназнал все три на Ликорхин , [ 15 ] Все остальные авторы считают три рода различными. [ 9 ] В семье гетеродонтозавр и ликорхин считаются родственными таксонами , а Аббиктозавр в качестве базального члена. [ 4 ] Геранозавр также является гетеродонтозавридом, но обычно считается номенем дубия, потому что тип образца отсутствует все свои зубы, что делает его неотличимым от любого другого рода в семье. [ 3 ] Совсем недавно род Echinodon считался гетеродонтозавридом в нескольких исследованиях. [ 10 ] [ 11 ] Lanasaurus был назван в честь верхней челюсти в 1975 году, [ 23 ] Но более поздние открытия показали, что вместо этого он принадлежит Lycorhinus , что делает Lanasaurus младшим синонимом этого рода. [ 6 ] Dianchungosaurus когда -то считался гетеродонтозавридом из Азии , [ 24 ] Но с тех пор было показано, что останки были просауропод и химерой мезоукрокодийских останков . [ 25 ] Жозе Бонапарт также классифицировал южноамериканский пизанозавр как гетеродонтозаврид в один раз, [ 26 ] Но это животное в настоящее время известно как более базальный орнитишский. [ 27 ]

Членство в Heterodontosauridae хорошо известно по сравнению с его неопределенным филогенетическим положением. Несколько ранних исследований показали, что гетеродонтозавриды были очень примитивными орнитишами. [ 5 ] [ 18 ] Из -за предполагаемого сходства в морфологии передних конечностей Роберт Баккер предложил взаимосвязь между гетеродонтозавридами и ранними сауроподоморфами , такими как Анчисавр , соединяя приказы Сауришиа и Орнитишиа . [ 14 ] Доминирующая гипотеза за последние несколько десятилетий поместила гетеродонтозавриды в качестве базальных орнитопод. [ 3 ] [ 4 ] [ 8 ] [ 28 ] Тем не менее, другие предположили, что гетеродонтозавриды вместо этого имеют общего предка с маргиноцефалией ( цератопсианцы и пахицефалозавры ), [ 29 ] [ 30 ] Гипотеза, которая обнаружила поддержку в некоторых исследованиях начала 21 -го века. [ 31 ] [ 32 ] Клада, содержащая гетеродонтозавриды и марниноцефальцы, была названа Heterodontosauriformes . [ 33 ] Гетеродонтозавриды также рассматривались как базальные как для орнитопод, так и по маргиницефалиям. [ 34 ] [ 35 ] В 2007 году кладистический анализ показал, что гетеродонтозавриды являются базальными ко всем известным орнитишам, кроме Pisanosaurus , что повторяет одну из самых ранних работ по семье. [ 36 ] [ 37 ] Тем не менее, исследование, проведенное Bonaparte, показало, что Pisanosauridae были синонимом Heterodontosauridae, а не отдельной семьей само по себе, включая Pisanosaurus в качестве гетеродонтозавра. [ 38 ] Butler et al. (2010) обнаружили, что Heterodontosauridae является наиболее базальным известным значимым орнитишским излучением. [ 39 ]
Кладограмма ниже показывает взаимосвязи в Heterodontosauridae и следует анализу Sereno, 2012: [ 40 ]
| Heterodontosauridae | |
Переработка Cerapoda в 2020 году от Dieudonné и коллег восстановила животных, традиционно считавшихся «гетеродонтозавридами» как базальную группировку в пахицефалозавре, парафилетической по отношению к традиционным, головаемым куполовым пахицефалозаврами. Этот результат был основан на многочисленных характеристиках черепа, включая зубное утепление , а также для того, чтобы объяснить тот факт, что окаменелости пахицефалозавра совершенно неизвестны из юрского периода. Современное понимание орнитишской филогении подразумевает, что юрские пахицефалозавры должны существовать, потому что были обнаружены многочисленные юрские цератопсийцы , однако не было идентифицировано таких пахицефалозавров. Этот анализ был сделан, чтобы уточнить выводы Барона и коллег (2017), которые считали, что Чилесура является базальным орнитишким . [ 41 ] Филогенетический анализ проводился с чилс -явром, кодируемым как орнитиша, который также имел значение для филогения орнитоподов .
Кладограмма ниже представляет собой сокращенную версию результатов Дидиудонны и коллег: [ 42 ]
| Ornithischia |
| |||||||||||||||||||||
Распределение
[ редактировать ]
Несмотря на то, что он первоначально известен только с ранней юры Южной Африки, останки гетеродонтозавридов в настоящее время известны с четырех континентов . В начале истории гетеродонтозавридов суперконтинент Pangea все еще был в значительной степени нетронутой, что позволило семье достичь почти в течение всего мира . [ 10 ] Самым старым известным возможным останками гетеродонтозаврида являются фрагмент челюсти и изолированные зубы из Колада Лагуна формации Аргентины , которая восходит к позднему триасу . Эти останки имеют полученную морфологию, аналогичную гетеродонтозавру , в том числе канинообразную с зубцами как по передним, так и на заднем краях, а также с высоким содержанием верхнечелюстных зубов, в которых отсутствует сингулу. [ 43 ] Irmis et al. (2007) предварительно согласились с тем, что этот ископаемый материал представляет собой гетеродонтозаврид, но заявил, что для подтверждения этого назначения необходимы дополнительные материалы, поскольку образец плохо сохранился, [ 44 ] В то время как Sereno (2012) только заявил, что этот материал может представлять собой орнитиша или даже специально гетеродонтозаврид. [ 1 ] Olsen , Kent & Whiteside (2010) отметили, что сама возраст формирования Лагуна Колада плохо ограничен, и, таким образом, он не был окончательно определил, имеет ли предполагаемый гетеродонтозаврид из этой формирования триаса или юрского возраста. [ 45 ] Самая разнообразная фауна гетеродонтозавридов происходит из ранней юры Южной Африки, где ископаемые гетеродонтозавра , абриктозавр , ликорхин и сомнительный геранозавр . обнаружены [ 3 ]
Незаписные ранние юрские гетеродонтозавриды также известны из Соединенных Штатов [ 46 ] и Мексика , [ 47 ] соответственно. Кроме того, начиная с 1970 -х годов, было обнаружено множество ископаемых материалов из формирования поздней юры Моррисон недалеко от Фрукты , штат Колорадо , в Соединенных Штатах. [ 11 ] Описанный в печати в 2009 году, этот материал был помещен в род Fruitadens . [ 12 ] Зубы гетеродонтозавридов, в которых отсутствуют поясные, также были описаны из поздних юрских и ранних меловых образований в Испании и Португалии . [ 48 ] Остатки Эхинодона были перестроены в 2002 году, показывая, что это может представлять собой позднюю гетеродонтозаврид с Берриасской стадии раннего мела на юге Англии. [ 10 ] Dianchungosaurus из ранней юры Китая больше не считается гетеродонтозавридом; хотя известно одна азиатская форма юрского периода ( Tianyulong ). [ 7 ] Неопределенные щеки зубы, возможно, представляющие гетеродонтозавриды, также известны из Barremian Wessex формирования в южной Англии, что, если это будет подтверждено, будет представлять собой самый молодой отчет о группе. [ 49 ]
Палеобиология
[ редактировать ]
Большинство окаменелостей гетеродонтозавридов встречаются в геологических формациях , которые представляют засушливые и полузасушливые среды, включая формирование верхнего эллиота Южной Африки и ложе Пурбека на юге Англии. [ 4 ] Было высказано предположение, что гетеродонтозавриды перенесли сезонную аспект или спячку в самые сухие времена года. Из -за отсутствия замены зубов в большинстве гетеродонтозавридов было предположено, чтобы весь набор зубов был заменен в течение этого спящего периода, поскольку казалось, что постоянная и спорадическая замена зубов прервела бы функцию зубного ряда в качестве одной жевательной поверхности Полем [ 15 ] Тем не менее, это было основано на недопонимании механики челюсти гетеродонтозавридов. [ 50 ] Считалось, что гетеродонтозавриды действительно постоянно заменяли свои зубы, хотя и медленнее, чем в других рептилиях , но компьютерная томография черепа из ювенильных и зрелых гетеродонтозавра не показывает заменяющих зубов. [ 51 ] В настоящее время нет никаких доказательств, которые подтверждают гипотезу о эстиции в гетеродонтозавридах, [ 3 ] Но это не может быть отвергнуто, основываясь на сканировании черепа. [ 51 ]
В то время как щеки -зубы гетеродонтозавридов явно адаптированы для измельчения жесткого растительного материала, их диета могла быть всеядшей. Опознанные предчелюстные зубы и острые изогнутые когти на передних конечностях предполагают некоторую степень хищного поведения. Было высказано предположение, что длинные, мощные передние конечности гетеродонтозавра могли быть полезны для разрыва в гнездах насекомых, аналогично современным антитетерам . Эти передние конечности могли также функционировать как инструменты копания, возможно, для корней и клубней . [ 3 ]
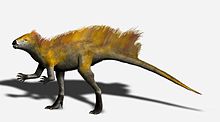
Длина передней передней части по сравнению с задними конечностями предполагает, что гетеродонтозавра могли быть частично четвероногими , а выдающийся процесс олекранона и гиперэкстратируемые цифры передней части конечности обнаруживаются во многих четвероногих. Тем не менее, Manus четко предназначен для захвата, а не поддержки веса. Многие особенности задних конечностей, в том числе длинная большеберцовая кость и нога, а также слияние тибиофибиотарса и тарсометатарса, указывают на то, что гетеродонтозавриды были адаптированы для быстрого бега на заджебках, поэтому маловероятно, что гетеродонтозавр двинулся на все четыре, за исключением, возможно, в течение, возможно, в течение, возможно, когда, возможно, когда, возможно. кормление. [ 5 ]
Короткие клыки, найденные во всех известных гетеродонтозавридах, сильно напоминают клыки, найденные у современных мускусных оленей , пекари и свиней . У многих из этих животных (а также с более длинным моржом и азиатскими слонами ) это сексуально диморфная черта, с клыками только у мужчин. Типовому образцу абриктозавра отсутствует клыки и первоначально был описан как самка. [ 15 ] Хотя это остается возможным, невзванные сакральные позвонки и короткие лица указывают на то, что этот образец представляет юношеское животное. Второй, более крупный образец, который первоначально предполагал, что он принадлежит к абриктозавру, явно обладает клыками, которые использовались для поддержки идеи, что клыки встречаются только у взрослых, а не вторичной сексуальной характеристики мужчин. Эти клыки могли быть использованы для боя или демонстрации с членами одного и того же вида или с другими видами. [ 3 ] Отсутствие клыков в ювенильных абриктозаврах также может быть еще одной характеристикой, отделяющей его и от других гетеродонтозавридов, как известны клыки в ювенильных гетеродонтозавра . Другие предлагаемые функции для клыков включают защиту и использование в иногда всеядную диету. [ 51 ] Тем не менее, этот образец был альтернативно переназначен на Lycorhinus Sereno в 2012 году, который, как известно, уже имел клыки, и, следовательно, их отсутствие в Abrictosaurus, возможно, не было результатом возраста. [ 1 ]
был обнаружен небольшой полный ископаемый скелет гетеродонтозавра более 200 миллионов лет В 2005 году в Южной Африке . В июле 2016 года он был отсканирован командой южноафриканских исследователей, использующих европейский синхротронный радиационный объект ; Сканирование зубного зуба выявило кости неба толщиной менее миллиметра. [ 52 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и Серено, Пол С. (2012). «Таксономия, морфология, жевательная функция и филогения гетеродонтозавридов динозавров» . Зоуки (226): 1–225. doi : 10.3897/Zookeys.226.2840 . PMC 3491919 . PMID 23166462 .
- ^ Мадзия, Даниэль; Арбор, Виктория М.; Бойд, Клинт А.; Фарке, Эндрю А.; Cruzado-Caballero, Penélope; Эванс, Дэвид С. (2021-12-09). «Филогенетическая номенклатура орнитишских динозавров» . ПЕРЕЙ . 9 : E12362. doi : 10.7717/peerj.12362 . ISSN 2167-8359 . PMC 8667728 . PMID 34966571 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Weshampel, David B .; Витмер, Лоуренс М. (1990). "Gererododosasasuredae" В Weshampel, David B.; Додсон, Питер; Osmóllska, Halszka (ред.). Динария Беркли: Университет Калифорнийской прессы. стр. 486–4 ISBN 978-0-520-06727-1 .
- ^ Jump up to: а беременный в дюймовый Норман, Дэвид Б .; Суды, Ганс-Дитер; Витмер, Лоуренс М.; Кория, Родольфо А. (2004). «Базальная орнитопода». В Вайшампеле, Дэвид Б.; Додсон, Питер; Osmólska, Halszka (ред.). Динозаврия (второе изд.). Беркли: Университет Калифорнийской прессы. С. 393–412. ISBN 978-0-520-24209-8 .
- ^ Jump up to: а беременный в дюймовый Санта -Лука, Альберт П. (1980). «Посткраниальный скелет Гетеродонтозавра Туки (Рептилия, Орнитиша) из Стормберга Южной Африки». Анналы Южноафриканского музея . 79 (7): 159–211.
- ^ Jump up to: а беременный Гоу, Кристофер Э. (1990). «Зубная верхняя челюсть, относящаяся к Lycorhinus angustidens Haughton, 1924 (Dinosauria, Ornithischia)». Анналы Южноафриканского музея . 99 (10): 367–380.
- ^ Jump up to: а беременный Чжэн, Ся-Тин; Ты, Хай-Лу; Сюй, Син; Донг, Чжи-Мин (19 марта 2009 г.). «Ранний меловой гетеродонтозавридной динозавр с нитевидными покровными структурами». Природа . 458 (7236): 333–336. Bibcode : 2009natur.458..333Z . doi : 10.1038/nature07856 . PMID 19295609 . S2CID 4423110 .
- ^ Jump up to: а беременный Тулборн, Ричард А. (1970). «Систематическое положение триасового Ornithischian Dinosaur Lycorhinus angustidens ». Зоологический журнал Линневого общества . 49 (3): 235–245. doi : 10.1111/j.1096-3642.1970.tb00739.x .
- ^ Jump up to: а беременный Хопсон, Джеймс А. (1975). «О общем отделении орнитишских динозавров Lycorhinus и Heterodontosaurus из серии Stormberg (Верхний триасы) Южной Африки». Южноафриканский журнал науки . 71 : 302–305.
- ^ Jump up to: а беременный в дюймовый Норман, Дэвид Б .; Барретт, Пол М. (2002). «Орнитишайские динозавры из нижнего мела (Берриаса) Англии». В Милнере, Эндрю; Баттен, Дэвид Дж. (Ред.). Жизнь и окружающая среда в Purbeck Times . Специальные документы в палеонтологии 68 . Лондон: палеонтологическая ассоциация. С. 161–189. ISBN 978-0-901702-73-9 .
- ^ Jump up to: а беременный в Гальтон, Питер М. (2007). «Зубы орнитишских динозавров (в основном Ornithopoda) из формирования Моррисона (верхняя юрская величина) западных Соединенных Штатов». В Карпентере, Кеннет (ред.). Рога и клювы: цератопсские и орнитоподские динозавры . Блумингтон: издательство Университета Индианы. С. 17–47. ISBN 978-0-253-34817-3 .
- ^ Jump up to: а беременный Батлер, Ричард Дж.; Гальтон, Питер М.; Porro, Laura B.; Chiappe, Luis M.; Хендерсон, DM; Эриксон, Грегори М. (2010). «Нижние пределы орнитишского размера тела динозавров, выведенные из нового верхнего юрского гетеродонтозаврида из Северной Америки» . Труды Королевского общества б . 277 (1680): 375–381. doi : 10.1098/rspb.2009.1494 . PMC 2842649 . PMID 19846460 .
- ^ Weishampel, David B. (1984). "Введение". Эволюция механизмов челюсти у динозавров Ornithopod . Достижения в области анатомии, эмбриологии и клеточной биологии. Тол. 87. Берлин; Нью-Йорк: Springer-Verlag. С. 1–109. doi : 10.1007/978-3-642-69533-9_1 . ISBN 978-0-387-13114-6 Полем PMID 6464809 .
- ^ Jump up to: а беременный Баккер, Роберт Т. (1986). Верни динозавров: новые теории, раскрывающие тайну динозавров и их вымирание . Нью -Йорк: Уильям Морроу. п. 453pp. ISBN 978-0-14-010055-6 .
- ^ Jump up to: а беременный в дюймовый Тулборн, Ричард А. (1974). «Новый гетеродонтозауридный динозавр (Reptilia: Ornithischia) из верхних триасовых красных слоев Лесото». Зоологический журнал Линневого общества . 55 (2): 151–175. doi : 10.1111/j.1096-3642.1974.tb01591.x .
- ^ Метла, Роберт. (1911). «О динозаврах Стормберга, Южная Африка». Анналы Южноафриканского музея . 7 : 291–308.
- ^ Haughton, Sidney H. (1924). «Фауна и стратиграфия серии Stormberg». Анналы Южноафриканского музея . 12 : 323–497.
- ^ Jump up to: а беременный Кромптон, AW; Количество, Алан (1962). «Новый Ornithischian из верхнего триаса Южной Африки». Природа . 196 (4859): 1074–1077. Bibcode : 1962nater.196.1074c . doi : 10.1038/1961074a0 . S2CID 4198113 .
- ^ Ромер, Альфред С. (1966). Палеонтология позвоночных (третье изд.). Чикаго: Университет Чикагской Прессы. п. 468 стр . ISBN 978-0-7167-1822-2 .
- ^ Серено, Пол (2012-03-10). «Таксономия, морфология, жевательная функция и филогения гетеродонтозавридов динозавров» . Зоуки (226): 1–225. doi : 10.3897/Zookeys.226.2840 . ISSN 1313-2970 . PMC 3491919 . PMID 23166462 .
- ^ Серено, Пол С. (1998). «Рациональный для филогенетических определений, с применением к таксономии динозаврии более высокого уровня». Новый ежегодник для геологии и палеонтологии, трактаты . 210 (1): 41–83. Doi : 10.1127/njgpa/210/998/41 .
- ^ Серено, Пол С. (2005-11-07). «Архосаврия стебля - Taxonsearch» . Архивировано из оригинала 2007-02-19 . Получено 2007-02-24 .
- ^ Гоу, Кристофер Э. (1975). «Новый гетеродонтозаврид из красных болтов Южной Африки, показывающий четкие доказательства замены зубов». Зоологический журнал Линневого общества . 57 (4): 335–339. doi : 10.1111/j.1096-3642.1975.tb01895.x .
- ^ Янг, CC (1982). «[Новый род динозавра из округа Луфенг, провинция Юньнань]». В Чжоу М. (ред.). [Собранные произведения Ян Чжунцзян] (на китайском языке). Пекин: Academica Sinica. С. 38–42.
- ^ Барретт, Пол М.; Сюй Син (2005). «Переоценка Dianchungosaurus lufengensis Yang, 1982a, загадочная рептилия из нижней формирования Луфэна (нижняя юра) провинции Юньнань, Китайская Народная Республика». Журнал палеонтологии . 79 (5): 981–986. doi : 10.1666/0022-3360 (2005) 079 [0981: Arodly] 2.0.co; 2 . ISSN 0022-3360 . S2CID 89542580 .
- ^ Родился, Джозеф Ф. (1976). Полем Журнал палеонологии 50 : 808–8
- ^ Вайшампл, Дэвид Б .; Витмер, Лоуренс М. (1990). « Lesothosaurus , Pisanosaurus и Technosaurus ». В Вайшампеле, Дэвид Б.; Додсон, Питер; Osmólska, Halszka (ред.). Динозаврия (первое изд.). Беркли: Университет Калифорнийской прессы. С. 416–425. ISBN 978-0-520-06727-1 .
- ^ Серено, Пол С. (1986). «Филогения динозавров с птицами». Национальные географические исследования . 2 (2): 234–256.
- ^ Чжао Xijin (1983). Полем Acta Poolonological Польша 28 (1–2): 295–3
- ^ Купер, Майкл А. (1985). «Пересмотр орнитишского динозавра Кангнасавр Кутцеи Хаутон, с классификацией Ornithischia». Анналы Южноафриканского музея . 95 : 281–317.
- ^ Чжао Сиджин ; Ченг Чжэнву; Сюй Син (1999). «Самый ранний кератопс из Тучэнзи Формирования Ляонинг, Китай». Журнал палеонтологии позвоночных . 19 (4): 681–691. Bibcode : 1999jvpal..19..681x . doi : 10.1080/02724634.1999.10011181 .
- ^ Вы приветствуете; Сюй Син ; Ван Сяолин (2003). «Новый род psittacosauridae (Dinosauria: Ornithopoda) и происхождение и ранняя эволюция динозавров Marginocephaly». Acta Geologica Sinica (английское издание) . 77 (1): 15–20. Bibcode : 2003acgls..77 ... 15y . doi : 10.1111/j.1755-6724.2003.tb00105.x . S2CID 89051352 .
- ^ Сюй Син, х ; Форстер, Кэтрин А.; Кларк, Джеймс М.; Мо Джиньу. (2006). «Базальный цератопс с переходными чертами из позднего юрского периода северо -западного Китая» . Труды Королевского общества B: Биологические науки . 273 (1598): 2135–2140. doi : 10.1098/rspb.2006.3566 . PMC 1635516 . PMID 16901832 .
- ^ Мэри, Тереза ; Осмоллска, Халска. (1985)). Филогани " Acta Poolonological Польша 30 : 137–1
- ^ Батлер, Ричард Дж. (2005). «Фаброзаурид» орнитишских динозавров формирования Верхнего Эллиота (Нижняя Юра) Южной Африки и Лесото » . Зоологический журнал Линневого общества . 145 (2): 175–218. doi : 10.1111/j.1096-3642.2005.00182.x .
- ^ Батлер, Ричард Дж .; Смит, Роджер М.Х.; Норман, Дэвид Б. (2007). «Примитивный орнитишский динозавр из позднего триаса Южной Африки, а также ранняя эволюция и диверсификация Ornithischia» . Труды Королевского общества B: Биологические науки . 274 (опубликовано онлайн): 2041–6. doi : 10.1098/rspb.2007.0367 . PMC 2275175 . PMID 17567562 .
- ^ Батлер, Ричард Дж.; Упчерч, Пол; Норман, Дэвид Б. (2008). «Филогения орнитишских динозавров». Журнал систематической палеонтологии . 6 (1): 1–40. Bibcode : 2008jspal ... 6 .... 1b . doi : 10.1017/s1477201907002271 . S2CID 86728076 .
- ^ Bonaparte, JF (1976). «Pisanosaurus mertii Casamiquela и происхождение Ornithischia». Журнал палеонтологии. 50 (5): 808–820. JSTOR 1303575 .
- ^ Rj Батлер. 2010. Анатомия базального орнитишского динозавра эокуруса Парвуса из формирования Нижнего Эллиота (позднего триаса) Южной Африки. Зоологический журнал Линневого общества 160: 648-684
- ^ Серено, ПК (2012). Стр. 193-2
- ^ Барон, Мэтью Г.; Барретт, Пол М. (2018-03-01). «Поддержка размещения Chilesaurus в Ornithischia: ответ Müller et al» . Биологические письма . 14 (3): 20180002. DOI : 10.1098/rsbl.2018.0002 . ISSN 1744-9561 . PMC 5897612 . PMID 29593075 .
- ^ Dieudonné, PE; Cruzed-Caballero, P.; Godefroit, P.; Tortosa, T. (2020). «Новая филогения динозавров Cerapodan» . Биология историческая . 33 (10): 2335–2355. Doi : 10.1080/08912963.2020.1793979 .
- ^ Баез, Ана Мария; Marsican, Claudia A. (2001). Disnosaur от верхнего триссера Trissic Patago Амегина 38 (3): 271–2
- ^ Рэндалл Б. Ирмис; Уильям Г. Паркер; Стерлинг Дж. Несбитт; Джун Лю (2007). «Ранние орнитишские динозавры: Tiasasic Record». Историческая биология . 19 (1): 3–22. Bibcode : 2007hbio ... 19 .... 3i . Citeseerx 10.1.1.539.8311 . doi : 10.1080/08912960600719988 . S2CID 11006994 .
- ^ Пол Э. Олсен; Деннис В. Кент; Джессика Х. Уайтсайд (2010). «Последствия астрохронологии и геомагнитной полярности на основе супергруппы и шкалы времени геомагнитной полярности (Newark-APTS) для темпа и способа ранней диверсификации динозавров». Земля и экологическая наука Сделок Королевского общества Эдинбурга . 101 (3–4): 201–229. Bibcode : 2010eest.101..201o . doi : 10.1017/s1755691011020032 . S2CID 16123732 .
- ^ Суды, Ганс-Дитер; Кларк, Джеймс М.; Дженкинс, Фариш А. (1994). «Обзор ранних юрских тетрапод из группы Глен -Каньон на юго -западе». В Фрейзере, Николас С.; Суды, Ганс-Дитер (ред.). В тени динозавров: ранние мезозойские тетраподы . Кембридж: издательство Кембриджского университета. С. 285–294. ISBN 978-0-521-45899-3 .
- ^ Кларк, Джеймс; Монтеллано, Марисоль; Хопсон, Джеймс А.; Эрнандес, Рене; Фасковский, Дэвид А. (1994). «Ранняя или средняя юрская сборка тетрапода из формации La Boca, Северо -Восточная Мексика». В Фрейзере, Северная Каролина; Суды, H.-D. (ред.). В тени динозавров: ранние мезозойские тетраподы . Кембридж: издательство Кембриджского университета. С. 295–302. ISBN 978-0-521-45899-3 .
- ^ Санчес-Хернандес, Барбара; Бентон, Майкл Дж .; Найш, Даррен (2007). «Динозавры и другие ископаемые позвоночные из позднего юрского периода и раннего мела в области Галве, NE, Испания» (PDF) . Палеогеография, палеоклиматология, палеоэкология . 249 (1–2): 180–215. Bibcode : 2007ppp ... 249..180s . doi : 10.1016/j.palaeo.2007.01.009 .
- ^ Свитман, Стивен С. (март 2016 г.). «Сравнение барремно-ранних аптианских позвоночных сообщений от группы Jehol, северо-восточного Китая и группы Wealden, Южная Великобритания: ценность исследований микровертообразных в неблагоприятных условиях сохранения» . Палеобиоразнообразие и палеоокружение . 96 (1): 149–167. BIBCODE : 2016PDPE ... 96..149S . doi : 10.1007/s12549-015-0217-9 . ISSN 1867-1594 . S2CID 129956539 .
- ^ Хопсон, Джеймс А. (1980). «Функция зубов и замена в ранних мезозойских орнитишских динозаврах: последствия для эстиции». Летая . 13 (1): 93–105. Bibcode : 1980Letha..13 ... 93H . doi : 10.1111/j.1502-3931.1980.tb01035.x .
- ^ Jump up to: а беременный в Батлер, Ричард Дж.; Porro, Laura B.; Норман, Дэвид Б. (2008). «Ювенильный череп примитивного орнитишского динозавра гетеродонтозавра Таки из« Стормберга »Южной Африки». Журнал палеонтологии позвоночных . 28 (3): 702–711. doi : 10.1671/0272-4634 (2008) 28 [702: AJSOTP] 2.0.co; 2 . ISSN 0272-4634 . S2CID 86739244 .
- ^ «Ископаемые в SA, обнаруженные в SA, наконец -то отказываются от своих секретов» . 2016-07-27. Архивировано с оригинала 2016-07-28 . Получено 2016-07-27 .