Европазавр
| Европазавр Временной диапазон: поздняя юра ,
| |
|---|---|

| |
| Реконструированный скелет, Музей динозавров Аатала. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | † Зауроподоморфа |
| Клэйд : | † Зауропода |
| Клэйд : | † Макронария |
| Клэйд : | † Камарасавроморфа |
| Род: | † Европазавр Матеус и др. у Сандера и др. , 2006 г. [ 1 ] |
| Разновидность: | † Э. Хольгери
|
| Биномиальное имя | |
| † Europasaurus holgeri Матеус и др. в Сандере и др. , 2006 г. [ 1 ]
| |
Европазавр (что означает «европейская ящерица») — базальный макронарий зауропод , форма четвероногого травоядного динозавра . Он жил в позднеюрский период ( средний кимеридж , примерно 154–151 миллион лет назад) на севере Германии и был идентифицирован как пример островной карликовости , возникшей в результате изоляции популяции зауроподов на острове в бассейне Нижней Саксонии .
Открытие и присвоение имени
[ редактировать ]
В 1998 году единственный зуб зауропода был обнаружен частным коллекционером окаменелостей Хольгером Людтке в действующем карьере на горе Лангенберг , между общинами Окер , Харлингерод и Геттингерод в Германии. действовал Меловой карьер Лангенберг уже более века; горные породы добываются с помощью взрывных работ и в основном перерабатываются в удобрения. Карьер обнажает почти непрерывную последовательность карбонатных пород толщиной 203 м (666 футов). [ 2 ] принадлежащие формации Сюнтель , возраст которых варьируется от раннего оксфорда до позднего и кимериджа отложились в мелком море с глубиной воды менее 30 м (98 футов). Слои, обнаженные в карьере, ориентированы почти вертикально и слегка опрокинуты, что является результатом подъема прилегающих гор Гарц в течение нижнего мела . Карьер, широко известный среди геологов как классическое обнажение, широко изучался и посещался студентами-геологами на протяжении десятилетий. Хотя окаменелости морских беспозвоночных богаты окаменелостями, окаменелости наземных животных были редки. Зуб зауропода был первым экземпляром динозавра-зауропода из юрского периода на севере Германии. [ 3 ]
После того, как было обнаружено больше ископаемого материала, включая кости, в апреле 1999 года начались раскопки костного слоя, проводимые местной ассоциацией частных коллекционеров окаменелостей. Хотя оператор карьера сотрудничал, раскопки осложнялись почти вертикальной ориентацией слоев, ограничивавшей доступ, а также продолжающейся добычей полезных ископаемых. Материал зауроподов не мог быть извлечен непосредственно из слоя, его приходилось собирать из блоков, образовавшихся в результате взрывных работ. Таким образом, точное происхождение костного материала было неясно, но позже его можно было проследить до одного слоя (слой 83). [ 3 ] [ 4 ] В ходе раскопок, проведенных 20–28 июля 2000 г., было обнаружено ок. 50 т (49 длинных тонн; 55 коротких тонн) костных блоков, содержащих останки позвоночных. [ 5 ] Окаменелости были подготовлены и сохранены в Парке динозавров в Мюнхене (DFMMh), частном музее динозавров под открытым небом, расположенном недалеко от Ганновера . Из-за очень хорошей сохранности костей консолидирующие средства приходилось применять лишь изредка, а подготовку можно было провести сравнительно быстро, поскольку кость легко отделялась от окружающей породы. Кости простой формы иногда можно было подготовить менее чем за час, тогда как подготовка крестца требовала трехнедельного труда. К январю 2001 года уже было подготовлено 200 отдельных костей позвоночных. На данный момент самая высокая плотность кости была обнаружена в блоке размером 70 х 70 см, который содержал ок. 20 костей. [ 4 ] К январю 2002 года при подготовке еще большего блока был обнаружен частичный череп зауропода – первый, обнаруженный в Европе. Перед полным удалением костей из блока из блока была изготовлена силиконовая модель, чтобы зафиксировать точное трехмерное положение отдельных костей. [ 5 ]
Часть ископаемого материала европазавра была повреждена или уничтожена в результате поджога в ночь с 4 на 5 октября 2003 года. Пожар уничтожил лабораторию и выставочный зал Парка динозавров в Мюнхене, в результате чего было потеряно 106 костей, которые составляют 15% костей, приготовленных в то время. Кроме того, пожар затронул большую часть еще неподготовленных блоков, а на раскаленный камень попала пожарная вода, что привело к дополнительному разрушению. [ 6 ] Среди разрушенных экземпляров имеется DFMMh/FV 100, включавший наиболее сохранившуюся серию позвонков и единственный целый таз. [ 7 ] [ 6 ]

В 2006 году новый таксон зауроподов был официально описан как Europasaurus holgeri . Данная этимология названия рода - «рептилия из Европы», а видовое название дано в честь Хольгера Людтке, первооткрывателя первых окаменелостей. [ 1 ] Учитывая сравнительно небольшой размер костей, первоначально предполагалось, что они принадлежат молодым особям. Однако в публикации 2006 года было установлено, что большинство экземпляров были взрослыми особями, а европазавр был островным карликом. [ 3 ] Число отдельных костей зауропод увеличилось до 650 и включает особей с различным сочленением; материал был обнаружен на площади 60–80 м (200–260 футов) в квадрате. [ 2 ] Из этих экземпляров был выбран голотип — разрозненная, но ассоциированная особь (DFMMh/FV 291). [ 2 ] Голотип включает в себя множественные черепа кости ( предчелюстную , верхнюю и квадратно-скуловую кости ), частичную черепную коробку , множественные кости нижней челюсти ( зубные , надугольные и угловые ), большое количество зубов, шейные позвонки , крестцовые позвонки и ребра от шеи и туловища. По меньшей мере еще 10 особей были отнесены к тому же таксону на основании совпадения материалов.
Летом 2012 года начались масштабные раскопки с целью выкопать кости европазавра не только из отдельных блоков, но и непосредственно из слоя горных пород. Доступ к костному пласту потребовал удаления около 600 тонн породы с помощью экскаваторов и колесных погрузчиков, а также постоянной откачки воды из основания карьера. Раскопки продолжались весной и летом 2013 года. В результате кампании были обнаружены новые останки рыб, черепах и крокодилов, а также ценная информация о костном слое; Однако дополнительные кости европазавра обнаружить не удалось. К 2014 году из слоя 83 было получено около 1300 костей позвоночных, большинство из которых принадлежало европазаврам ; еще около 3000 костей ждут подготовки. По костям челюсти было идентифицировано минимум 20 особей. [ 3 ]
Описание
[ редактировать ]Европазавр — очень маленький зауропод, его длина составляет всего 5,7–6,2 м (19–20 футов), а взрослый вес — 750–800 кг (1650–1760 фунтов). [ 1 ] [ 8 ] [ 9 ] Эта длина была оценена на основе частичной бедренной кости, масштабированной до размеров почти полного экземпляра камаразавра . Известны более молодые особи, размером от 3,7 м (12 футов) до самого молодого подростка ростом 1,75 м (5,7 футов). [ 1 ] [ 2 ]
Отличительные характеристики
[ редактировать ]
Помимо того, что был очень маленьким неозауроподом , европазавр он , как считалось, обладал множеством уникальных морфологических особенностей, позволяющих отличать его от близких родственников, согласно его первоначальным описаниям Сандеру и др. (2006 г. [ 1 ] ). Считалось, что носовой отросток предчелюстной кости изгибается вперед и выступает вверх (теперь это известно как сохранение). [ 10 ] имеется выемка ), на верхней поверхности центров шейных позвонков , лопатка имеет выступающий отросток на задней поверхности тела, таранная кость (голеностопная кость) в два раза шире высоты. [ 1 ]
По сравнению с камаразавром , у европазавра иная морфология заглазничной кости , где задний фланец не такой короткий, короткий контакт между носовыми и лобными костями черепа, форма его теменной кости (прямоугольная у европазавра ) и нервных отростков. его позвонки перед тазом не расщеплены. Также были упомянуты сравнения с брахиозавром (ныне называемым жираффатитан ), и было установлено, что у европазавра более короткая морда, контакт между квадратно-скуловой и чешуйчатой костью , а также плечевая кость (верхняя кость передней конечности), которая имеет уплощенные и выровненные проксимальную и дистальную поверхности. Наконец, были проведены быстрые сравнения с потенциальным брахиозавридом Lusotitan , который имеет другую форму подвздошной кости и таранной кости, и Cetiosaurus humerocristatus (названный Duriatitan ), у которого дельтопекторальный гребень менее выражен и простирается на меньшую часть плечевой кости. [ 1 ]
Череп
[ редактировать ]
сохранились почти все наружные кости черепа европазавра У экземпляров , за исключением префронтальных костей . Некоторые дополнительные кости представлены лишь очень фрагментированными и неинформативными окаменелостями, например, слезные кости . Известно восемь предчелюстных костей, обычно с прямоугольной формой морды, как у камаразавра . Передняя проекция предчелюстной кости, выявленная Sander et al. (2006 г. [ 1 ] ) был повторно идентифицирован как артефакт консервации в Marpmann et al. (2014 г. [ 10 ] сходный с анатомией, обнаруженной у Camarasaurus и Euhelopus ), в меньшей степени . Дорсальный выступ предчелюстной кости, тот, который соприкасается с носовой костью, начинается как задне- дорсальный выступ, а затем становится прямым вертикальным в точке поднариального отверстия , пока не достигнет носового отверстия. Этот слабый «шаг» наблюдается у камаразавра и эугелопа и более сильно присутствует у абидозавра , жирафатитана и, возможно, черепа брахиозавра . Эти последние таксоны также имеют более длинную морду с большим расстоянием от первого зуба до носового отростка предчелюстной кости. Кроме того, европазавр разделяет с базальными камаразавроморфами (брахиозавридами, камаразаврами , эвхелопусами и малавизаврами ) орбиту и носовые отверстия аналогичного размера , тогда как носовое отверстие значительно уменьшено у производных титанозавров ( рапетозавр , тапуиазавр и немегтозавр ). [ 10 ]

Единственная верхняя челюсть присутствует в хорошо сохранившемся материале европазавра , DFMMh/FV 291.17. Эта верхняя челюсть имеет длинное тело с двумя удлиненными отростками: носовым и задним. Имеется лишь слабый слезный отросток, как у большинства зауроподов, за исключением рапетозавра . Носовой отросток удлиненный, прикрывает передний и вентральный края анторбитального окна . Этот отросток распространяется примерно на 120° от горизонтального зубного ряда . Основание носового отростка образует также тело слезного отростка, а при их расхождении находится предглазничное отверстие, по форме сходное с таковым у камаразавра , эвгелопуса , абидозавра и жирафатитана , но примерно на 1/2 выше пропорционально. Преанторбитальное отверстие, небольшое отверстие перед или под анторбитальным отверстием, хорошо развито у таких таксонов, как Diplodocus и Tapuiasaurus , и почти отсутствует, как у Camarasaurus и Euhelopus . было около 12–13 зубов Всего в верхней челюсти европазавра , что меньше, чем у более базальных таксонов (16 зубов у Jobaria и 14–25 у Atlasaurus ), но попадающий в диапазон изменчивости Brachiosauridae (от 15 у Brachiosaurus до 10 у Abydosaurus ). крае до четырех мелких зубчиков Все сохранившиеся нестертые зубы имеют на медиальном . Небольшое количество коронок задних зубов слегка искривлено (~15°), но значительно меньше, чем у брахиозаврид (30–45°). [ 10 ]

Среди носовых костей европазавра известно несколько, но немногие из них являются целыми или неискаженными. Носовые кости сзади перекрываются лобными костями , а сбоку тупо сочленяются с предлобными костями. В отличие от носовых костей жирафа , у европазавра они выступают горизонтально вперед, образуя небольшую часть крыши черепа над переднеглазничными отверстиями. известны четыре фронтальных изображения У европазавра : три слева и один справа. Из-за их разчленения вполне вероятно, что лобные кости никогда не сливались во время роста, в отличие от камаразавра . Лобные кости образуют часть крыши черепа, сочленяясь с другими костями, такими как носовые, теменные, префронтальные и заднеглазничные кости , и они длиннее в передне-заднем направлении, чем ширина, что является уникальным признаком среди эузавроподов . Как и у диплодокоидов ( Amargasaurus , Dicraeosaurus и Diplodocus ), а также Camarasaurus , лобные кости исключены из лобно-теменных окон (или теменных окон, когда лобные кости исключены). Фронтальные части также исключены из край надвисочного окна (распространенный признак у зауроподов, более продвинутых, чем у шунозавра ), и они имеют лишь небольшое, неорнаментированное участие в орбите . известно несколько теменных костей У европазавра , которые имеют прямоугольную форму, гораздо более широкую, чем длинную. теменные кости также широкие, если смотреть с задней части черепа, они немного выше большого затылочного отверстия (отверстия спинного мозга). Теменные кости составляют примерно половину границы поствисочного окна (отверстия над самой задней частью черепа), а другая область окружена чешуйчатыми костями и некоторыми костями черепной коробки. Теменные кости также образуют часть края надвисочного окна, которое у европазавра шире, чем длинное , как у жираффатитана , камаразавра и спинофорозавра . Помимо вышеупомянутых окон, теменные также имеют «посттеменные окна», которые редко можно увидеть за пределами Dicraeosauridae . присутствует трехлучевая заглазничная кость У европазавра , которая развилась в результате слияния постфронтальной и постосбитальной костей более базальных таксонов. Между передним и вентральным выступающими отростками заглазничная кость образует край орбиты, а между задним и вентральным отростками она граничит с подвисочное окно . [ 10 ]

известны множественные скуловые кости У европазавра , которые по морфологии больше похожи на базальных зауроподоморфов , чем на других макронарий. Он образует часть границы глазницы, подвисочного окна и нижнего края черепа, но не достигает предглазничного окна. Задний отросток скуловой кости очень хрупкий и узкий, на нем виден костный рубец от сочленения с квадратноскуловой костью. Из задней части тела скуловой кости выступают два выступа, которые расходятся под углом 75° и образуют нижний и передний края подвисочного окна. Как и у Riojasaurus и Massospondylus , двух зауроподоморфов, не являющихся зауроподами, скуловая кость образует большую часть края орбиты, от заднего до переднего нижнего угла. Эта особенность наблюдалась у эмбрионов титанозавров, но не у взрослых особей. Квадратоскуловая кость представляет собой удлиненный элемент, имеющий два выступающих плеча: переднее и дорсальное. Как и у других зауроподов, передний отросток длиннее спинного, но у европазавра руки более одинаковой длины. Горизонтальный отросток параллелен зубному ряду европазавра , сходен с камаразавром, но в отличие от жираффатитана и абидозавра . На переднем плече кости имеется выступающий вентральный выступ, который, возможно, является синапоморфией Brachiosauridae, хотя он также встречается у некоторых Camarasaurus особей . Два квадратно-скуловых отростка расходятся почти под прямым углом (90°), хотя дорсальный отросток изгибается, повторяя форму квадратной кости . Чешуйчатые кости, найденные у европазавра, имеют ту же форму при взгляде сбоку, что и камаразавр , то есть вопросительный знак . Чешуйчатые кости сочленяются со многими костями черепа, в том числе с костями крыши черепа, вентральной части черепа и черепной коробки. Как и заглазничные кости, чешуйчатые кости трехлучевые, с вентральной, передней и медиальной. процесс. [ 10 ]
Сохранилось тринадцать элементов нёба европазавра , кости включая квадратную, крыловидную и эктоптеригоидную . Квадратные кости сочленяются с костями неба и черепной коробки, а также с наружными костями черепа. По форме они похожи на жираффатитов и камаразавров и имеют хорошо развитые суставные поверхности. На большей части длины квадратной кости имеется один стержень с крыловидным крылом вдоль медиальной стороны. Крыловидные кости — самые крупные из небных костей зауроподов, они имеют трехлучевую форму, как и заглазничные кости. Передний выступ контактирует с противоположной крыловидной костью, латеральное крыло - с эктоптеригоидной, а заднее крыло поддерживает квадратную и базиптеригоидную кости (кость, обеспечивающая соединение неба с черепной коробкой). Эктоптеригоид — небольшая кость неба, которая соединяет центральные кости неба (крыловидную и небную ) с верхней челюстью. Эктоптеригоиды имеют L-образную форму, передний отросток прикрепляется к верхней челюсти, а дорсальный отросток соединяется с крыловидным отростком. [ 10 ]
Позвонки
[ редактировать ]
Шейные позвонки европазавра лучше всего сохранились и наиболее представлены из позвоночного столба. Однако не вся шея известна, поэтому число шейных позвонков может находиться между камаразавром (12 позвонков) и рапетозавром (17 позвонков). Кроме того, несколько шейных позвонков происходят от людей разного возраста, а длина центра и внутренняя структура, как известно, меняются на протяжении развития. У взрослых шейные центры удлиненные, опистоцельные (передний конец шаровидный), с выемкой на вершине заднего конца центра. Эта особенность была описана как характерная для европазавра, но также известна у Euhelopus и Giraffatitan . В боковой части центра европазавра имеется углубление, открывающееся во внутреннюю часть позвонков. В отличие от жираффатитов и брахиозавридов, у европазавра нет тонких гребней ( пластин ), разделяющих это отверстие. Европазавр имеет общие черты пластинок на верхних позвонках с базальными макронариями и брахиозавридами. В отличие от передне- и среднешейных, задние шейные позвонки менее удлинены и пропорционально выше, как и у других макронарий, со значительными изменениями в положении суставных поверхностей. [ 7 ]

Передние спинные позвонки сильно опистоцельные, как и шейные, и могут быть объединены в ряды на основании отсутствия гипантрума и низкого расположения парапофизов . Внутренние структуры открытые и камерытные , как у Camarasaurus , Giraffatitan и Galvesaurus , но в отличие от этих таксонов эта пневмоность не распространяется на средние и задние спинные позвонки. Расположение и наличие передних пластинок у европазавра сходно с другими базальными макронариями, но в отличие от более базальных таксонов (например, Mamenchisaurus , Haplocanthosaurus ) и более производных таксонов (например, Giraffatitan ). Средние спинные кости имеют пневматическую полость, которая простирается вверх в нервную дугу , как у Barapasaurus , Cetiosaurus , Tehuelchesaurus и Camarasaurus . Вентральный край этого отверстия ромбовидный, четко выраженный. В задних позвонках латеральная пневматическая полость сместилась выше по центру, изменение наблюдается и у других базальных макронарий. Эти плевроцели широкие спереди и узкие, а сзади становятся остроугольными. нервный отросток европазавра брахиозавра стоит вертикально - базальная особенность, не встречающаяся у или более производных зауроподов. [ 7 ]
Ряд всех полных крестцовых позвонков известен только по одному экземпляру, DFMMh/FV 100, который был уничтожен во время пожара в 2003 году. Все пять позвонков, характерное количество более базальных неозауроподов, включены в крестец . Третий и четвертый сакралы представляют собой изначальные сакралы, присутствующие во всех группах динозавров. Второй, S2, является предковым сакральным зауроподоморфом, который был добавлен к базальным зауроподоморфам, у которых у всех есть три сакральных, за исключением Платеозавра . Пятый крестец, сросшийся позади первичной пары, представляет собой каудосакраль, мигрировавший из хвоста в таз у таксонов вокруг Leonerasaurus . Первый крестцовый отдел, сочлененный с подвздошной костью, но не сросшийся с другими позвонками, представляет собой спинно-крестцовый отдел, в результате чего количество позвонков, обнаруженных у всех неозауроподов, достигло пяти. Уровень слияния дорсо-крестцового отдела подтверждает эволюционную историю крестцового счета зауроподов: первичная пара включала сначала спинной (всего три), затем хвостовой (всего четыре), а затем еще один спинной позвонок, в результате чего в общей сложности образовалось пять позвонков. [ 7 ]
Кожа
[ редактировать ]Среди макронарий окаменелые отпечатки кожи известны только у Haestasaurus , Tehuelchesaurus и Saltasaurus . И Tehuelchesaurus , и Haestasaurus могут быть тесно связаны с Europasaurus , а характеристики отпечатков кожи всех зауроподов схожи. Гестазавр , первый динозавр, известный по отпечаткам кожи, сохранил покровы на части руки вокруг локтевого сустава. [ 11 ] Кожные отпечатки более распространены в материале Tehuelchesaurus , где они известны из областей передних конечностей, лопаток и туловища. Костных пластинок или узелков, указывающих на броню, нет, но есть несколько типов чешуек. Типы кожи Tehuelchesaurus в целом больше похожи на те, что встречаются у диплодоцидов и Haestasaurus, чем у эмбрионов титанозавров из Аука Мауево . [ 12 ] Поскольку форма и сочленение сохранившихся бугорков у этих базальных макронарий аналогичны другим таксонам, где сохранилась кожа, включая экземпляры Brontosaurus excelsus и промежуточных диплодокоидов, такие кожные структуры, вероятно, широко распространены среди Neosauropoda. [ 11 ]
Классификация
[ редактировать ]был впервые назван, Когда европазавр он считался таксоном Макронарии, который не входил в семейство Brachiosauridae или кладу Titanosauromorpha . Это указывало на то, что карликовость таксона была результатом эволюции, а не характеристикой группы. [ 1 ] Были проанализированы три матрицы с включением европазавра , матрицы Уилсона (2002 г.) [ 13 ] ) и Апчерч (1998 г.) [ 14 ] ) и Upchurch et al. (2004 г. [ 15 ] ). Все анализы привели к сходной филогенетике, где европазавр оказался более производным, чем камаразавр , но вне клады Brachiosauridae и Titanosauromorpha (теперь называемой Titanosauriformes ). Результаты предпочтительного анализа Sander et al. (2006) показаны ниже слева: [ 2 ]

Во время описания позвонков европазавра Carballido & Sander (2013) был проведен еще один филогенетический анализ (правый столбец выше). Кладистическая матрица была расширена за счет включения большего количества таксонов зауропод, таких как Bellusaurus , Cedarosaurus и Tapuiasaurus . Таксон Brachiosaurus также был разделен на настоящих Brachiosaurus ( B. altithorax ) и Giraffatitan ( B. brancai ), на основании Тейлора (2009). [ 16 ] ). На основании этого нового и более обширного анализа было обнаружено, что европазавр находится в аналогичном положении, поскольку базальный камаразавроморф ближе к титанозаврам, чем камаразавр . Однако Euhelopus , Tehuelchesaurus , Tastavinsaurus и Galvesaurus были помещены между Europasaurus и Brachiosauridae. [ 7 ]
Размещение как брахиозаврид
[ редактировать ]В анализе филогении Titanosauriformes в 2012 году Д'Эмик (2012) считал, что Europasaurus принадлежит к Brachiosauridae, а не является базальным по отношению к самым ранним брахиозавридам. Филогения определила наиболее истинных брахиозавридов на сегодняшний день, хотя вместо этого было установлено, что несколько потенциальных брахиозавридов принадлежали к Somphospondyli ( Paluxysaurus , Sauroposeidon и Qiaowanlong ). Однако Д'Эмик неуверенно считал европазавра подтвержденным брахиозавридом. Хотя в филогении была сильная поддержка его размещения, европазавр , один из немногих базальных макронарий с черепом, не имеет нескольких костей, которые демонстрируют характерные особенности группы, такие как хвостовые позвонки. Кладограмма ниже слева иллюстрирует филогенетические результаты Д'Эмика (2012) с разрушенными Euhelopodidae и Titanosauria. [ 17 ]

Более поздний анализ титанозаврообразных согласился с Д'Эмиком (2012) относительно места европазавра . Он образовал политомию с брахиозавром и «французским ботриоспондилом » (названным Vouivria ) в качестве основных брахиозавридов. Следующим по степени происхождения в кладе был Lusotitan , а Giraffatitan , Abydosaurus , Cedarosaurus и Venenosaurus образовывали более производную кладу брахиозавридов. «Искривленные» зубы европазавра оказались одной из уникальных особенностей Brachiosauridae, что могло означать уверенную принадлежность изолированных зубов зауроподов к кладе. [ 18 ]
Дальнейший филогенетический анализ был проведен на Brachiosauridae на основе анализа Д'Эмика (2012). Эта филогения, проведенная Д'Эмиком и соавт. (2016) разрешили очень похожее размещение европазавра внутри Brachiosauridae, хотя Sonorasaurus был помещен в кладу с Giraffatitan , а Lusotitan был помещен в политомию с Abydosaurus и Cedarosaurus . Оставшееся дерево было таким же, как у Д'Эмика (2012), хотя брахиозавр превратился в политомию с более производными брахиозавридами. [ 19 ] Другая филогения, Mannion et al. (2017) получили результаты, аналогичные данным D'Emic (2012) и D'Emic et al. (2016). Europasaurus был самым базальным брахиозавридом, а «французский Bothriospondylus», или Vouivria , был следующим по базальному брахиозавриду. Брахиозавр был помещен вне полтомии всех других брахиозавридов, жираффатитана , абидозавра , соноразавра , цедарозавра и вененозавра . [ 20 ] Филогения 2017 года, предложенная Ройо-Торресом и др. (2017) разрешили более сложные отношения внутри Brachiosauridae. Помимо европазавра как самого базового брахиозаврида, внутри клады существовало две подгруппы: одна включала Giraffatitan , Sonorasaurus и Lusotitan , а другая включала почти всех других брахиозавридов, а также Tastavinsaurus . эту вторую кладу можно было бы назвать Laurasiformes Согласно определению группы, . Брахиозавр находился в политомии с двумя субкладами Brachiosauridae. Филогения Ройо-Торреса и др. (2017) можно увидеть выше, в правом столбце. [ 21 ]
Палеобиология
[ редактировать ]Рост
[ редактировать ]
было установлено, что европазавр был уникальным карликовым видом, а не молодым представителем существующего таксона, такого как камаразавр Путем гистологического анализа нескольких экземпляров европазавра . Этот анализ показал, что самый молодой экземпляр (DFMMh/FV 009) не имеет признаков старения, таких как следы роста или пластинчатая костная ткань, а также является самым маленьким экземпляром - 1,75 м (5,7 футов) в длину. Такая костная ткань является показателем быстрого роста, поэтому, вероятно, это молодой молодой особь. Более крупный экземпляр (DFMMh/FV 291,9) на высоте 2 м (6,6 футов) демонстрирует большое количество пластинчатой ткани без следов роста, поэтому, вероятно, это тоже молодой особь. Следующий по размеру экземпляр (DFMMh/FV 001) имеет следы роста (в частности, кольца ), и при длине 3,5 м (11 футов), возможно, является полувзрослым экземпляром. Еще более крупный DFMMh/FV 495 имеет зрелые остеоны и кольца, его длина составляет 3,7 м (12 футов). У второго по величине проанализированного экземпляра (DFMMh/FV 153) также наблюдаются следы роста, но они встречаются чаще. Размер этого экземпляра составляет 4,2 м (14 футов). Единственная неполная бедренная кость представляет собой самый крупный из известных Особь европазавра , длина тела 6,2 м (20 футов). В отличие от всех других экземпляров, у этого экземпляра (DFMMh/FV 415) наблюдаются линии остановки роста , что указывает на то, что он погиб после достижения полного размера тела. Внутренняя кость также частично пластинчатая , что указывает на то, что она недавно перестала расти. [ 1 ] [ 2 ]
Эти комбинации факторов роста показывают, что Europasaurus развил свои небольшие размеры из-за значительно сниженной скорости роста, набирая размер медленнее, чем более крупные таксоны, такие как Camarasaurus . Это замедление темпов роста противоположно общей тенденции зауроподов и теропод , которые достигли больших размеров с увеличением темпов роста. [ 1 ] Некоторые из близких родственников европазавра представляют собой крупнейших известных динозавров , в том числе брахиозавра и зауропосейдона . Марпманн и др. (2014) предположили, что небольшой размер и сниженная скорость роста европазавра были следствием педоморфизма , при котором взрослые особи таксонов сохраняют ювенильные характеристики, такие как размер. [ 10 ]
Исследования внутренних ушей детенышей европазавра показывают, что они были рано развитыми, и предполагается, что они в некоторой степени зависели от защиты взрослых в стаде, чего не наблюдается у более крупных зауроподов из-за огромной разницы в размерах родителей и потомство. , обладали хорошим слухом Строение и большая длина внутреннего уха представителей этого рода также позволяют предположить, что они, как и европазавр . Внутривидовое общение также, по-видимому, было важно для этого зауропода, основываясь на этих исследованиях, позволяющих предположить, что этот зауропод демонстрировал четкое, стадное поведение. [ 22 ]
карликовость
[ редактировать ]
Было высказано предположение, что предок европазавра должен был быстро уменьшиться в размерах после эмиграции на остров, существовавший в то время, поскольку самый большой из островов в регионе вокруг северной Германии имел площадь менее 200 000 км (120 000 миль) в квадрате. которые, возможно, не могли поддерживать сообщество крупных зауроподов. С другой стороны, макронария могла уменьшаться одновременно с большей территорией суши, пока не достигла размеров европазавра . [ 1 ] Предыдущие исследования островной карликовости в основном ограничивались маастрихтом острова Хацег в Румынии , где обитают карликовый титанозавр Magyarosaurus и карликовый гадрозавр Telmatosaurus . [ 1 ] [ 23 ] [ 24 ] тельматозавр Известно, что произошел от маленькой взрослой особи, и хотя он очень маленький, известно, что экземпляры мадьярозавра небольших размеров происходят от взрослых до старых особей. [ 24 ] [ 25 ] Взрослые особи Magyarosaurus dacus имели размер тела, аналогичный Europasaurus , но у самого крупного из последних были более длинные бедра, чем у самого крупного из первых, в то время как Magyarosaurus hungaricus был значительно крупнее любого из таксонов. [ 25 ] Карликовость у европазавра представляет собой единственное значительное быстрое изменение массы тела у производных зауроподоморф , при этом общей тенденцией таксонов является рост общего размера в других группах. [ 26 ]
Палеоэкология
[ редактировать ]
Район Лангенберг в Германии, от раннего оксфорда до позднего кимериджа, демонстрирует разнообразие растительного и животного мира островной экосистемы поздней юры. [ 27 ] В кимериджском периоде эта местность была морской и располагалась между Рейнишским, Богемским и Лондонско-Брабантским массивами . Это не указывает на то, что присутствующие таксоны были морскими , поскольку животные и растения могли быть отложены аллохтонно (хотя и на небольшом расстоянии) от окружающих островов. [ 28 ] Отложения показывают, что время от времени происходил приток пресной или солоноватой воды, и сохранившиеся окаменелости подтверждают это. большое количество окаменелостей морских двустворчатых моллюсков , а также иглокожих В известняке карьера присутствует и микрокаменелостей, хотя многие животные и растения были наземными. [ 29 ]
Многие морские таксоны сохранились в Лангенберге, хотя они не часто сосуществовали с европазавром . Существует как минимум три рода черепах : Plesiochelys , Thalassemys и до двух неназванных таксонов. [ 27 ] [ 30 ] [ 31 ] Обильны актиноптеригии , представленные Lepidotes , Macromesodon , Proscinetes , Coelodus , Macrosemius , Notagogus , Histionotus , Ionoscopus , Callopterus , Caturus , Sauropsis , Belonostomus и Thrissops . [ 32 ] Также присутствуют по крайней мере пять различных морфологий гибодонтовых акул: неоселахий Palaeoscyllium , Synechodus и Asterodermus . [ 33 ] два морских крокодилоформных вида Из Лангенберга известны , Machimosaurus и Steneosaurus крокодилообразный земноводный Goniopholis . , которые, вероятно, питались черепахами и рыбой, а также был обнаружен [ 34 ]

хвойные шишки и ветки можно идентифицировать как араукария Brachyphyllum По наземным окаменелостям, обнаруженным на этом участке, . В этом месте хранится множество таксонов, в том числе большое скопление костей и особей европазавра . не менее 450 костей европазавра , около 1/3 из которых имела следы зубов. В карьере Лангенберг было обнаружено [ 27 ] Размеры и форма этих следов зубов хорошо совпадают с зубами рыб , крокодилообразных или других падальщиков, но подтвержденных следов теропод нет. [ 27 ] [ 35 ] Большое количество присутствующих особей позволяет предположить, что стадо европазавров пересекало приливную зону и утонуло. [ 27 ] Хотя доминирующим крупнотелым животным является европазавр , есть также материал от диплодоцидного зауропода, стегозавра и нескольких теропод. У диплодоцида сохранились три шейки матки, и по их размеру можно предположить, что таксон также был карликовым. Стегозавры и различные тероподы сохраняют только зубы, за исключением нескольких костей, возможно, принадлежащих таксону Ceratosauridae . [ 27 ] как минимум четыре различных типа теропод Отдельные зубы показывают, что в этом месте обитало , включая мегалозаврид Torvosaurus sp. а также еще один мегалозаврид и неопределенные представители Allosauridae и Ceratosauria ; а также есть самые старые зубы, известные у Velociraptorinae . [ 36 ] [ 37 ]
Помимо динозавров, в карьере Лангенберг сохранилось и множество мелких наземных позвоночных. К таким животным относятся хорошо сохранившийся трехмерный скелет птерозавра из Dsungaripteridae и изолированные останки из Ornithocheiroidea и Ctenochasmatidae ; ящерица парамацеллодидная ; а также частичные скелеты и черепа родственника Theriosuchus, теперь известного как род Knoetschkesuchus . [ 27 ] [ 28 ] [ 30 ] Также сохранились зубы дриолестидных млекопитающих, а также докодонта , таксона Eobaataridae , и мультитуберкулята , имеющего сходство с Proalbionbataar (ныне называемого Teutonodon ). [ 27 ] [ 38 ]
Вымирание
[ редактировать ]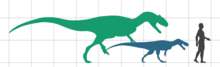
Следы динозавров, сохранившиеся в карьере Лангенберг, указывают на возможную причину вымирания европазавра и, возможно, других островных карликов, присутствующих на островах региона. Следы расположены на высоте 5 м (16 футов) над отложениями особей европазавра , что показывает, что по крайней мере через 35 000 лет после этого отложения произошло падение уровня моря, что привело к перевороту фауны. Тероподы, населяющие остров, сосуществовавшие с европазаврами , имели рост около 4 м (13 футов), но тероподы, прибывшие по сухопутному мосту, сохранили следы. до 54 см (21 дюйм), что указывает на размер тела от 7 до 8 м (23 и 26 футов), если реконструировать его как аллозавра . Описатели этих следов (Йенс Лалленсак и его коллеги) предположили, что эти таксоны теропод, вероятно, привели к вымиранию специализированной карликовой фауны, а слой, из которого произошли следы (слой Лангенберг 92), вероятно, самый молодой, в котором европазавр присутствует . . [ 39 ] [ 40 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н Сандер, премьер-министр; Матеус, О.В.; Лавен, Т.; Кнётшке, Н. (2006). «Гистология костей указывает на островную карликовость у нового позднеюрского динозавра-зауропода». Природа . 441 (7094): 739–741. Бибкод : 2006Natur.441..739M . дои : 10.1038/nature04633 . ПМИД 16760975 . S2CID 4361820 .
- ^ Jump up to: а б с д и ж Сандер, премьер-министр; Матеус, О.В.; Лавен, Т.; Кнётшке, Н. (2006). «Дополнительная информация для: гистология костей указывает на островную карликовость у нового позднеюрского динозавра-зауропода». Природа . 441 (7094): 739–741. Бибкод : 2006Natur.441..739M . дои : 10.1038/nature04633 . ПМИД 16760975 . S2CID 4361820 .
- ^ Jump up to: а б с д Крылья, Оливер (2014). «Об охоте на карликовых динозавров в карьере Лангенберг под Госларом». Окаменелости (на немецком языке). 2 :44-50.
- ^ Jump up to: а б Кнетшке, Нильс (2001). «Первые результаты и опыт восстановления и подготовки фауны верхней юры (Киммериды)». Таксидермист (на немецком языке). 47 (1). Ганновер: 7–13.
- ^ Jump up to: а б Кнетшке, Нильс; Рёсслер, Деннис (2002). «Новые результаты «Ассоциации содействия палеонтологии Нижней Саксонии eV». Подготовитель (на немецком языке). 48 (1). Ганновер: 21–30.
- ^ Jump up to: а б Уингз, О.; Кнетшке Н. «Песнь взрыва и огня: Europasaurus holgeri ». 74-е ежегодное собрание Общества палеонтологии позвоночных: программа и тезисы .
- ^ Jump up to: а б с д и Карбаллидо, JL; Сандер, член парламента (2013). «Посткраниальный осевой скелет Europasaurus holgeri (Dinosauria, Sauropoda) из верхней юры Германии: значение для онтогенеза зауропод и филогенетических взаимоотношений базальных макронарий» (PDF) . Журнал систематической палеонтологии . 12 (3): 335–387. дои : 10.1080/14772019.2013.764935 . hdl : 11336/19199 . ISSN 1477-2019 . S2CID 85087382 .
- ^ Пол, Грегори С. (2016). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. п. 226. ИСБН 978-1-78684-190-2 . OCLC 985402380 .
- ^ Штейн, К.; Чики, З.; Карри Роджерс, К.; Вейшампель, Д.Б.; Редельсторфф, Р.; Карбаллидоа, JL; Сандер, премьер-министр (2010). «Небольшой размер тела и выраженное ремоделирование кортикальной кости указывают на филетическую карликовость у Magyarosaurus dacus (Sauropoda: Titanosauria)» . Труды Национальной академии наук Соединенных Штатов Америки . 20. 107 (20): 9258–9263. Бибкод : 2010PNAS..107.9258S . дои : 10.1073/pnas.1000781107 . ПМК 2889090 . ПМИД 20435913 .
- ^ Jump up to: а б с д и ж г час Марпманн, Дж. С.; Карбаллидо, JL; Сандер, премьер-министр; Кнетшке, Н. (2014). «Краниальная анатомия позднеюрского карликового зауропода Europasaurus holgeri (Dinosauria, Camarasauromorpha): онтогенетические изменения и размерный диморфизм». Журнал систематической палеонтологии . 13 (3): 1–43. дои : 10.1080/14772019.2013.875074 . S2CID 84076044 .
- ^ Jump up to: а б Апчерч, П.; Мэннион, Вашингтон; Тейлор, член парламента (2015). «Анатомия и филогенетические связи «Pelorosaurus» becklesii (Neosauropoda, Macronaria) из раннего мела Англии» . ПЛОС ОДИН . 10 (6): e0125819. Бибкод : 2015PLoSO..1025819U . дои : 10.1371/journal.pone.0125819 . ISSN 1932-6203 . ПМЦ 4454574 . ПМИД 26039587 .
- ^ Хименес, О. дель В. (2007). «Отпечатки кожи Tehuelchesaurus (Sauropoda) из верхней юры Патагонии» (PDF) . Журнал Аргентинского музея естественных наук . 9 (2): 119–124. дои : 10.22179/revmacn.9.303 . S2CID 132980503 . Архивировано из оригинала (PDF) 7 октября 2012 г. Проверено 25 октября 2017 г.
- ^ Уилсон, Дж. А. (2002). «Филогения динозавров зауроподов: критика и кладистический анализ» (PDF) . Зоологический журнал Линнеевского общества . 136 (2): 217–276. дои : 10.1046/j.1096-3642.2002.00029.x .
- ^ Апчерч, премьер-министр (1998). «Филогенетические взаимоотношения динозавров-зауроподов» (PDF) . Зоологический журнал Линнеевского общества . 124 : 43–103. дои : 10.1111/j.1096-3642.1998.tb00569.x .
- ^ Апчерч, Премьер-министр; Барретт, премьер-министр; Додсон, П. (2004). «Зауропода». В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 259–322 . ISBN 978-0-520-94143-4 .
- ^ Тейлор, член парламента (2009). «Переоценка Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) и его родовое отделение от Giraffatitan Branchai (Janensch 1914)» Журнал палеонтологии позвоночных . 29 (3): 787–806. CiteSeerX 10.1.1.441.3748 . дои : 10.1671/039.029.0309 . S2CID 15220647 .
- ^ Д'Эмик, доктор медицины (2012). «Ранняя эволюция титанозаврообразных динозавров-зауроподов» (PDF) . Зоологический журнал Линнеевского общества . 166 (3): 624–671. дои : 10.1111/j.1096-3642.2012.00853.x . S2CID 54752135 .
- ^ Мэннион, Вашингтон; Апчерч, П.; Барнс, Р.Н.; Матеус, О. (2013). «Остеология позднеюрского португальского динозавра-зауропода Lusotitan atalaiensis (Macronaria) и история эволюции базальных титанозавроформ» . Зоологический журнал Линнеевского общества . 168 (1): 98–206. дои : 10.1111/zoj.12029 . ISSN 0024-4082 .

- ^ Д'Эмик, доктор медицины; Бригадир, БЗ; Джуд, Н.А. (2016). «Анатомия, систематика, палеосреда, рост и возраст динозавра-зауропода Sonorasaurus thompsoni из мелового периода Аризоны, США». Журнал палеонтологии . 90 (1): 102–132. дои : 10.1017/jpa.2015.67 . ISSN 0022-3360 . S2CID 133054430 .
- ^ Мэннион, Вашингтон; Аллен, Р.; Мойн, О. (2017). «Самый ранний известный титанозавроподобный динозавр-зауропод и эволюция брахиозаврид» . ПерДж . 5 : е3217. дои : 10.7717/peerj.3217 . ISSN 2167-8359 . ПМК 5417094 . ПМИД 28480136 .
- ^ Ройо-Торрес, Р.; Фуэнтес, К.; Мейджиде, М.; Мейхиде-Фуэнтес, Ф.; Мейхиде-Фуэнтес, М. (2017). «Новый динозавр Brachiosauridae Sauropod из нижнего мела Европы (провинция Сория, Испания)». Меловые исследования . 80 : 38–55. Бибкод : 2017CrRes..80...38R . дои : 10.1016/j.cretres.2017.08.012 .
- ^ «Что внутреннее ухо европазавра говорит о его жизни: динозавр с длинной шеей из северной Германии был преждевременным» .
- ^ Нопца, Ф. (1914). «О появлении динозавров в Трансильвании». Вер. Зоол. Бот . 54 :12-14.
- ^ Jump up to: а б Вейшампель, Д.Б.; Норман, Д.Б.; Григореску, Д. (1993). « Telmatosaurus transsylvanicus из позднего мела Румынии: самый базальный гадрозавридный динозавр». Палеонтология . 36 : 361–385.
- ^ Jump up to: а б Штейн, К.; Чики, З.; Карри-Роджерс, К.; Вейшампель, Д.Б.; Редельсторфф, Р.; Карбаллидо, JL; Сандер, премьер-министр (2010). «Небольшой размер тела и выраженное ремоделирование кортикальной кости указывают на филетическую карликовость у Magyarosaurus dacus (Sauropoda: Titanosauria)» . Труды Национальной академии наук . 107 (20): 9258–9263. Бибкод : 2010PNAS..107.9258S . дои : 10.1073/pnas.1000781107 . ISSN 0027-8424 . ПМК 2889090 . ПМИД 20435913 .
- ^ Бенсон, РБЖ; Кампионе, Невада; Каррано, Монтана; Мэннион, Вашингтон; Салливан, К.; Апчерч, П.; Эванс, округ Колумбия (2014). «Скорости эволюции массы тела динозавров указывают на 170 миллионов лет устойчивых экологических инноваций в линии стволовых птиц» . ПЛОС Биология . 12 (5): e1001853. дои : 10.1371/journal.pbio.1001853 . ПМК 4011683 . ПМИД 24802911 .
- ^ Jump up to: а б с д и ж г час Крылья, О. (2015). «Карьер Лангенберг возле Гослара: уникальное окно в наземную экосистему поздней юры в Северной Германии» . Тезисы докладов 12-го симпозиума по мезозойским наземным экосистемам : 99–100.
- ^ Jump up to: а б Шварц, Д.; Раддац, М.; Крылья, О. (2017). « Knoetschkesurus langenbergensis gen. nov. sp. nov., новый крокодилообразный атопозавр из верхнеюрского карьера Лангенберг (Нижняя Саксония, северо-запад Германии) и его родство с Theriosuchus » . ПЛОС ОДИН . 12 (2): e0160617. Бибкод : 2017PLoSO..1260617S . дои : 10.1371/journal.pone.0160617 . ПМК 5310792 . ПМИД 28199316 .
- ^ Крылья, О. (2013). «Уникальная тафономия карликового зауропода Europasaurus из позднеюрских морских толщ Лангенбергского карьера (Нижняя Саксония, Северная Германия)» . Журнал палеонтологии позвоночных, программа и рефераты. 73-е ежегодное собрание Общества палеонтологии позвоночных . 2013 : 239.
- ^ Jump up to: а б Уингз, О.; Сандер, член парламента (2012). «Позднеюрский комплекс позвоночных из карьера Лангенберг, Окер, Северная Германия» . 10-е ежегодное собрание Европейской ассоциации палеонтологов позвоночных . 20 : 281–284.
- ^ Маринейру, Дж.; Матеус, О. (2011). «Обнаружение морской черепахи Thalassemys в кимеридже Окера, Германия» (PDF) . 71-е ежегодное собрание Общества палеонтологии позвоночных. Тезисы 71-го ежегодного собрания Общества палеонтологии позвоночных : 151.
- ^ Мудрох, А.; Тис, Д. (1996). «Костные рыбьи зубы (Osteichthyeso Actinopterygii) из верхней юры (кимериджа) Лангенберга близ Окера (Северная Германия)» . Geologica et Palaeontologica (на немецком языке). 30 : 239–265.
- ^ Тис, Д. (1995). «Плакоидные чешуи (Chondrichthyes: Elasmobranchii) из поздней юры (кимериджа) северной Германии». Журнал палеонтологии позвоночных . 15 (3): 463–481. дои : 10.1080/02724634.1995.10011242 .
- ^ Карл, Х.-В.; Грёнинг, Э.; Браукманн, К.; Шварц, Д.; Кнётшке, Н. (2006). «Позднеюрские крокодилы Лангенберга близ Окера, Нижняя Саксония (Германия), и описание сопутствующих материалов (с замечаниями по истории добычи «Лангенбергского известняка» и «Обернкирхенского песчаника»)» . Клаусталер Геовиссеншафтен . 5 : 59–77.
- ^ Слодоник, М.; Крылья, О. (2015). «Следы укусов на костях европазавра из карьера Лангенберг близ Гослара (Нижняя Саксония, Германия)» . 13-е ежегодное собрание Европейской ассоциации палеонтологов позвоночных Ополе, Польша, Тезисы докладов : 141.
- ^ Герке, О.; Крылья, О. (2016). «Многомерный и кладистический анализ изолированных зубов выявляет симпатию тероподовых динозавров в поздней юре Северной Германии» . ПЛОС ОДИН . 11 (7): e0158334. Бибкод : 2016PLoSO..1158334G . дои : 10.1371/journal.pone.0158334 . ПМЦ 4934775 . ПМИД 27383054 .
- ^ Люббе, Т. ван Дер; Рихтер, У.; Кнетшке, Н. (2009). «Зубы велоцирапторина дромеозаврид из кимериджа (поздней юры) Германии» (PDF) . Acta Palaeontologica Polonica . 54 (3): 401–408. дои : 10.4202/app.2008.0007 . S2CID 38105783 .
- ^ Мартин, Т.; Шульц, Дж. А.; Шверманн, АХ; Крылья, О. (2016). «Первые юрские млекопитающие Германии: многобугорковые зубы из карьера Лангенберг (Нижняя Саксония)» (PDF) . Палеонтология Полоника . 67 : 171–179. doi : 10.4202/pp.2016.67_171 (неактивен 31 января 2024 г.). Архивировано из оригинала (PDF) 10 августа 2016 года . Проверено 27 октября 2017 г.
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Лалленсак, JN; Сандер, член парламента; Кнетшке, Н.; Крылья, О. (2015). «Следы динозавров из карьера Лангенберг (поздняя юра, Германия), реконструированные с помощью исторической фотограмметрии: свидетельства существования крупных теропод вскоре после островной карликовости» . Электронная палеонтология . 18 (2): 1–34.
- ^ Гарланд, Ник. «Карликовые динозавры вымерли на фоне блицкрига хищников» . Архивы Земли . Проверено 10 ноября 2022 г.
Дальнейшее чтение
[ редактировать ]- Холл, Британская Колумбия; Халлгримссон, Б. (2011). Эволюция Стрикбергера . Издательство Джонс и Бартлетт. п. 446. ИСБН 978-1-4496-4722-3 .
- Брусатте, СЛ (2012). Палеобиология динозавров . Джон Уайли и сыновья. п. 210. ИСБН 978-0-470-65658-7 .
- Болечек, К.; Крылья, О. (2013). Europasaurus holgeri — карликовый динозавр из поздней юры Германии . стр. 46–47. ISBN 978-84-695-8567-2 .
{{cite book}}:|journal=игнорируется ( помогите ) - Орган, CL; Брусатте, СЛ; Штейн, К. (2009). «Динозавры-зауроподы развили геномы среднего размера, не связанные с размером тела» . Труды Лондонского королевского общества B: Биологические науки . 276 (1677): 4303–4308. дои : 10.1098/rspb.2009.1343 . ISSN 0962-8452 . ПМК 2817110 . ПМИД 19793755 .
- Филиппини, ФС; Отеро, А.; Гаспарини, З. (2016). «Филогенетическая значимость крестца среди макронарий зауроподов: данные по тазу из верхнего мела Патагонии, Аргентина». Алчеринга . 41 : 69–78. дои : 10.1080/03115518.2016.1180806 . ISSN 0311-5518 . S2CID 131785420 .











