Гляциализавр
| Гляциализавр Временной диапазон: плинсбах ,
| |
|---|---|
| |
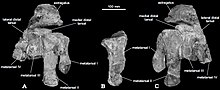
| Голотип костей стопы ( образец голотипа FMNH PR1823) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | † Зауроподоморфа |
| Семья: | † Массоспондилиды |
| Род: | † Гляциализавр Смит и Пол, 2007 г. |
| Типовой вид | |
| † Glacialisaurus Hammeri Смит и Пол, 2007 г.
| |
Glacialisaurus) — род динозавров зауроподоморфов - . Он жил на плинсбахском этапе раннего юрского периода около 186–182 миллионов лет назад на территории, которая сейчас является центральной областью гор Антарктиды Трансантарктических . Известен по двум экземплярам; голотип , а (имя-образец), частичная предплюсна (лодыжка) и плюсна также частичная левая бедренная кость. Окаменелости были собраны группой под руководством палеонтолога Уильяма Р. Хаммера во время полевой экспедиции 1990–91 годов к формации Хансон в Антарктиде. Они были описаны в 2007 году и легли в основу нового рода и вида Glacialisaurus Hammeri . Название рода переводится как «ледяная» или «замороженная ящерица», а видовое название чтит Хаммера.
Этот динозавр был классифицирован как массоспондилид , группа базальных (раннерасходящихся или «примитивных») зауроподоморфов среднего размера, существовавших в позднем триасе и ранней юре на всех континентах, кроме Австралии. Его длина оценивается в 6,2 м (20 футов). Гляциализавр был крупным травоядным динозавром, хотя по размеру он был средним для массоспондилид. Glacialisaurus отличался от других зауроподоморфов такими особенностями, как наличие мощного медиального надмыщелкового гребня на нижней части бедренной кости, мощного приводящего гребня, идущего от верхнего конца медиального мыщелка бедренной кости, и второй плюсневой кости со слабо выпуклым передним краем на нижней бедренной кости. верхний конец.
Открытие и присвоение имени
[ редактировать ]
Окаменелости динозавра -зауроподоморфа группой из колледжа Огастана под руководством палеонтолога Уильяма Р. Хаммера во время полевых исследований 1990–91 годов в нижней части формации Хэнсон горы Киркпатрик в Центральных Трансантарктических горах Антарктиды были обнаружены полевой , относящихся к ранней юре . [ 1 ] [ 2 ] [ 3 ] Окаменелости принадлежали двум разным людям: элементы правой лодыжки и плюсневой кости , такие как таранная кость , две предплюсны и четыре плюсневые кости, сохранившиеся в сочленении (образец FMNH PR1823), и нижняя часть левой бедренной кости (FMNH PR1822, бедренная кость). , заканчивающийся сразу после рассасывания медиального надмыщелкового гребня. [ 3 ]
На том же месте было собрано несколько других окаменелостей, в том числе окаменелости хищного тероподного динозавра криолофозавра , плечевой кости птерозавра (кость плеча) и большого зуба тритилодонта . [ 4 ] [ 2 ] [ 1 ] все они найдены на высоте около 4100 м (13 500 футов). [ 3 ] Правая лодыжка и предплюсна сохранились в слое слоев толщиной 1 м (3,3 фута) , а бедренная кость сохранилась при поверхностном выветривании рядом с экземпляром криолофозавра . [ 3 ] [ 4 ] Окаменелости были отправлены в Полевой музей естественной истории в Чикаго, США , и впервые о них сообщалось в 1994 году. [ 2 ] В этом отчете предполагалось, что шейные позвонки криолофозавра, найденные поблизости, также принадлежали зауроподоморфу. [ 2 ] но с тех пор это было опровергнуто. [ 3 ]
Окаменелости были описаны палеонтологами Натаном Смитом и Диего Полом, которые назвали новый род и вид Glacialisaurus Hammeri , используя FMNH PR1823 в качестве образца голотипа . Родовое название происходит от латинского корня glacialis, означающего «ледяной», после его открытия в районе ледника Бердмор в Центральных Трансантарктических горах, и слова sauros, означающего «ящерица». [ 3 ] Видовое название дано в честь Хаммера за его вклад в антарктическую палеонтологию. [ 3 ]
Описание
[ редактировать ]
известно немного останков Хотя о Glacialisaurus , кости его ног показывают, что это был крепкий базальный (ранний дивергентный или «примитивный») зауроподоморф. Фрагмент бедренной кости является более крупным из двух известных экземпляров, его размер в сохранившемся виде составляет 300 мм (0,98 фута), а предполагаемая общая длина в неповрежденном состоянии составляет 600 мм (2,0 фута). [ 3 ] По оценкам, гляциализавр имел длину около 6,2 м (20 футов). [ 5 ] Будучи базальным зауроподоморфом, Glacialisaurus имел длинную шею и пропорционально небольшую голову с листовидными зубами. Рука должна была быть короткой, широкой и крепкой, с большим когтем на большом пальце. [ 6 ]
Кости ног
[ редактировать ]

Поперечное сечение мощного стержня бедренной кости немного шире из стороны в сторону, чем спереди назад, хотя и не так сильно, как у эузавропод. Медиальный надмыщелковый гребень простирается от медиальной поверхности нижней части тела бедренной кости и отличается от всех других зауроподоморфов тем, что он прочный - признак, конвергентно развившийся у базальных теропод. Передняя поверхность бедренной кости плоская, а не выпуклая из стороны в сторону, что характерно для других базальных зауроподоморфов. На верхней поверхности верхней части бедра отсутствует передняя разгибательная борозда. На нижнем конце латеральный и медиальный мыщелки разделены краниокаудальной бороздой, которая резко заканчивается подколенной ямкой (отверстием в кости). Glacialisaurus также отличается мощным приводящим гребнем, идущим от верхнего конца медиального мыщелка бедренной кости. Этот гребень начинается на конце медиального мыщелка и имеет почковидную форму с длинной осью, проходящей проксимолатерально-дистомедиально. [ 3 ]


Таранная кость низкая и вытянутая из стороны в сторону, а медиальная часть не имеет краниокаудального расширения по сравнению с латеральной частью, что характерно для большинства неэузавропод . Астрагал на нижнем конце слабо выпуклый, хотя это не так сильно, как у Blikanasaurus и Lessemsaurus . Верхняя поверхность таранной кости слегка выпуклая, потому что именно здесь нижний конец большеберцовой кости (голени) сочленяется с таранной костью. Эта поверхность пронизана двумя ямками (небольшими отверстиями в кости), которые интерпретируются как сосудистые ямки . Восходящий отросток (выступ кости) имеет форму холмика и его верхняя суставная поверхность обращена проксимомедиально. Дистальные лапки имеют вытянутую вбок треугольную форму, если смотреть с верхних концов. Углы предплюсн округлые и выпуклые, особенно в заднемедиальном углу. Медиальный дистальный отдел предплюсны не ограничивается только III плюсневой костью, но едва прилегает к проксимальному концу II плюсневой кости, как у Сатурналии . Латеральная дистальная предплюсна имеет четырехугольную форму и, вероятно, была длиннее в медиолатеральном направлении, чем в проксимодистальном. [ 3 ]
Плюсневая кость I составляет примерно 3/4 длины плюсневой кости II, как и у большинства базальных зауроподоморфов. Плюсневая кость I имеет широкий и короткий стержень эллиптической формы, в большей степени, чем у большинства других базальных зауроподоморфов. Верхняя часть небольшой задней борозды, разделяющей два дистальных мыщелка, аналогична таковой у платеозавра . Медиальный мыщелок менее крепкий и расположен более проксимально, чем латеральный. Это могло бы вызвать медиальное смещение первого пальца, характерное для большинства ящеротазовых динозавров (группа, в которую входят зауроподоморфы и тероподы). Верхний конец II плюсневой кости имеет форму песочных часов и имеет вогнутые медиально-латеральные концы, сочленяющиеся с другими плюсневыми костями. Медиальная вогнутость хорошо развита, латеральная — слабее. [ 3 ]

К диагностическим признакам (признакам, отличающим таксон от других) вторых плюсневых костей относятся: передний край слабовыпуклый в проксимальном направлении; гипертрофированный латеральный подошвенный фланец на проксимальном конце (присутствует, но менее развит у многих базальных зауроподоморфов, например, Saturnalia , Plateosaurus ); и медиальный дистальный мыщелок, который более крепок и хорошо развит, чем латеральный дистальный мыщелок. Третьей плюсневой кости не хватает многих сохранившихся деталей, но она имеет трапециевидный верхний конец с прямым или вогнутым передним краем и слегка выпуклым медиальным краем для сочленения со II плюсневой костью. Задний край сбоку уже, чем передний, но не острый и не закругленный, в результате чего верхний контур III плюсневой кости почти трапециевидный, как у Lufengosaurus , Gyposaurus и Coloradisaurus . Напротив, большинство зауроподоморфов, не относящихся к эузавроподам, имеют почти треугольные верхние очертания. Сохранилась только верхняя часть IV плюсневой кости, но ее верхний контур аналогичен таковому у Люфенгозавр с широкой передней стороной и пальцеобразным заднемедиальным выступом. Этот пальцеобразный отросток слегка выпуклый и должен был сочленяться с III плюсневой костью. [ 3 ]
Классификация
[ редактировать ]Филогенетическое положение Glacialisaurus нестабильно из-за его фрагментарного характера, но он часто встречается как член семейства Massospondylidae . [ 7 ] [ 8 ] [ 9 ] [ 10 ] [ 11 ] [ 3 ] Массоспондилиды — это группа зауроподоморфов, не являющихся эузавроподами , которые существовали в период от позднего триаса до ранней юры в Африке, Антарктиде, Азии и Америке. [ 3 ] Массоспондилиды были обнаружены как сестринская группа (наиболее близкородственная группа) для более производных зауроподоморфов, включая самих Sauropoda , и более производных, чем такие группы, как Plateosauridae. [ 9 ] [ 3 ] В своем филогенетическом анализе взаимоотношений Glacialisaurus в 2007 году Смит и Пол обнаружили, что это был массоспондилид. [ 3 ] Особенности его стопы схожи с люфенгозавром (из ранней юры Китая), и филогенетическое исследование показало, что эти динозавры были близкими родственниками, тогда как Massospondylus оказался более базовой формой. [ 3 ] Это было подтверждено более поздними анализами, [ 12 ] [ 8 ] [ 9 ] [ 10 ] [ 7 ] включая Мюллера (2019), который нашел его в кладе с Coloradisaurus и Lufengosaurus , в то время как Massospondylus , Sarahsaurus , Pradhania и Xingxiulong были более базальными в семействе. [ 11 ]


Палеонтолог Оливер В.М. Раухут и его коллеги обнаружили, что Lufengosaurus является сестринским таксоном Glacialisaurus в 2020 году, а следующая кладограмма показывает место, которое они установили для Glacialisaurus в группе зауроподоморфов Massopoda: [ 10 ]
Открытие Glacialisaurus важно для изучения раннего распространения динозавров-зауроподов. [ 3 ] Присутствие этого примитивного зауроподоморфа в формации Хансон (где также были обнаружены останки, приписываемые настоящим зауроподам) показывает, что как примитивные, так и продвинутые представители этой линии существовали бок о бок в раннем юрском периоде. [ 13 ] [ 3 ]
Палеосреда
[ редактировать ]
Glacialisaurus известен из формации Хансон, которая является одной из двух основных скальных образований, содержащих динозавров, обнаруженных в Антарктиде. Образцы были обнаружены в туфоалевролитах , отложенных в синемюрско-плинсбахском ярусе ранней юры. [ 2 ] [ 14 ] датируемый примерно 194-188 миллионами лет назад. [ 3 ] [ 15 ] Это геологическое образование является частью группы Виктория Трансантарктических гор, которая находится на высоте примерно 4000 м (13 000 футов) над уровнем моря. [ 14 ] Большая высота этого места подтверждает идею о том, что в ранней юрской Антарктиде были леса, населенные разнообразными видами, по крайней мере, вдоль побережья. [ 16 ] Формация Хансон отложилась в активной вулкано-тектонической рифтовой системе, образовавшейся во время распада суперконтинента Гондвана . [ 15 ] Местный вулканизм и свидетельства лесных пожаров известны из некоторых палеоботанических памятников формации Хансон. [ 17 ]
Модели потоков воздуха в юрском периоде показывают, что прибрежные районы, вероятно, никогда не опускались ниже нуля, хотя внутри страны существовали более экстремальные условия. [ 18 ] Гляциализавр был найден примерно в 650 километрах (400 миль) от Южного полюса . [ 14 ] который в то время находился примерно на 1000 км (621 миль) или чуть дальше на север. [ 16 ] В этом образовании обнаружены останки крупного теропода криолофозавра птерозавра размером с ворону , диморфодонтидного размером с крысу , тритилодонтного синапсида и двух маленьких безымянных зауроподоморфов. [ 15 ] [ 3 ] [ 4 ] Многие роды растений также были обнаружены в районе пика Шафер формации Хансон, что позволяет предположить, что леса похожи на открытые леса Северного острова в Новой Зеландии. [ 17 ] Известные растения включают хвойные деревья Cheirolepidiaceae , хвощи Equisetites и папоротники Cladophlebis , которые также были обнаружены или похожи на растения, обнаруженные в других раннеюрских участках, представляющих теплый климат. [ 19 ] [ 20 ] [ 17 ] Базальные зауроподоморфы, такие как Glacialisaurus, были первыми очень крупными динозаврами и, из-за своего роста, первыми травоядными животными, способными питаться высоко. [ 6 ] [ 21 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Хаммер, В.Р., и Хикерсон, В.Дж. (1996). Значение фауны раннеюрских позвоночных Антарктиды. Континентальная юра , 215–218.
- ^ Перейти обратно: а б с д и Хаммер, Уильям Р.; Хикерсон, Уильям Дж. (1994). «Хохлатый теропод-динозавр из Антарктиды» . Наука . 264 (5160): 828–830. Бибкод : 1994Sci...264..828H . дои : 10.1126/science.264.5160.828 . ISSN 0036-8075 . ПМИД 17794724 . S2CID 38933265 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в Смит, Натан Д.; Пол, Диего (2007). «Анатомия базального динозавра-зауроподоморфа из раннеюрской формации Хансон в Антарктиде» . Acta Palaeontologica Polonica . 52 (4): 657–674.
- ^ Перейти обратно: а б с Смит, Н.Д., Маковицкий, П.Дж., Хаммер, В.Р., и Карри, П.Дж. (2007). Остеология Cryolophosaurus ellioti (Dinosauria: Theropoda) из ранней юры Антарктиды и значение для ранней эволюции теропод. Зоологический журнал Линнеевского общества , 151 (2), 377–421.
- ^ Хольц-младший, Томас Р. (2012). «Динозавры: наиболее полная и современная энциклопедия для любителей динозавров всех возрастов, приложение» (PDF) . Проверено 12 января 2012 г.
- ^ Перейти обратно: а б Хольц-младший, Томас Р. (2007). Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов . Нью-Йорк: Рэндом Хаус . п. 177-178. ISBN 978-0-375-82419-7 .
- ^ Перейти обратно: а б Апалдетти, Сесилия; Мартинес, Рикардо Н. (2010). Новая информация о черепе Coloradisaurus brevis Bonaparte (Dinosauria, Sauropodomorpha) из верхних слоев формации Лос-Колорадос (поздний триас), северо-запад Аргентины . ISBN 978-987-95849-7-2 .
- ^ Перейти обратно: а б Апалдетти, Сесилия; Пол, Диего; Йейтс, Адам (2013). «Посткраниальная анатомия Coloradisaurus brevis (Dinosauria: Sauropodomorpha) из позднего триаса Аргентины и ее филогенетическое значение: ПОСТЧРЕВНАЯ АНАТОМИЯ COLORADISAURUS BREVIS» . Палеонтология . 56 (2): 277–301. дои : 10.1111/j.1475-4983.2012.01198.x . hdl : 11336/3499 . S2CID 85851161 .
- ^ Перейти обратно: а б с Роу, Тимоти Б.; Сьюс, Ханс-Дитер; Рейс, Роберт Р. (2011). «Распространение и разнообразие самых ранних североамериканских динозавров-зауроподоморфов с описанием нового таксона» . Труды Королевского общества B: Биологические науки . 278 (1708): 1044–1053. дои : 10.1098/rspb.2010.1867 . ПМК 3049036 . ПМИД 20926438 .
- ^ Перейти обратно: а б с Раухут, OWM; Холверда, FM; Фуррер, Х. (2020). «Производный зауроподиформный динозавр и другой материал зауроподоморфов из позднего триаса кантона Шаффхаузен, Швейцария» . Швейцарский журнал геонаук . 113 (1): 8. дои : 10.1186/s00015-020-00360-8 . S2CID 220294939 .
- ^ Перейти обратно: а б Мюллер, Родриго Темп (2020). «Краниомандибулярная остеология Macrocollum itaquii (Dinosauria: Sauropodomorpha) из позднего триаса южной Бразилии» . Журнал систематической палеонтологии . 18 (10): 805–841. дои : 10.1080/14772019.2019.1683902 . ISSN 1477-2019 . S2CID 209575985 .
- ^ Эскурра, Мартин Д. (2010). «Новый ранний динозавр (Saurischia: Sauropodomorpha) из позднего триаса Аргентины: переоценка происхождения и филогении динозавров» . Журнал систематической палеонтологии . 8 (3): 371–425. дои : 10.1080/14772019.2010.484650 . ISSN 1477-2019 . S2CID 129244872 .
- ^ СЕРДА, Игнасио А.; Полина Карабахал, Ариана; САЛЬГАДО, Леонардо; Кориа, Родольфо А.; Регеро, Марсело А.; Тамбусси, Клаудия П.; Моли, Хуан Дж. (2012). «Первая запись динозавра-зауропода из Антарктиды» . Naturwissenschaften . 99 (1): 83–87. Бибкод : 2012NW.....99...83C . дои : 10.1007/s00114-011-0869-x . hdl : 11336/52393 . ISSN 1432-1904 . ПМИД 22173579 . S2CID 18921496 .
- ^ Перейти обратно: а б с Хаммер, WR; Хикерсон, WJ (1999). Томида, Ю.; Рич, TH; Викерс-Рич, Ю. (ред.). «Гондванские динозавры из юры Антарктиды». Материалы Второго симпозиума по динозаврам Гондваны. Монографии Национального научного музея. 15: 211–217.
- ^ Перейти обратно: а б с Смит, Северная Дакота; Хаммер, WR; Маковицкий, П.Дж. (2013). «Новые динозавры из раннеюрской формации Хансон в Антарктиде, а также закономерности разнообразия и биогеографии раннеюрских зауроподоморфов» . Геологическое общество Америки Рефераты с программами . 45 (7): 405–406.
- ^ Перейти обратно: а б Додсон, П. (1997). «Распространение и разнообразие». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров. Академическая пресса. стр. 10–13. ISBN 978-0-12-226810-6 .
- ^ Перейти обратно: а б с Бомфлер, Бенджамин; Потт, Кристиан; Керп, Ганс (2011). «Растительные комплексы из формации Шафер-Пик (нижняя юра), северная часть Земли Виктории, Трансантарктические горы» . Антарктическая наука . 23 (2): 188–208. Бибкод : 2011AntSc..23..188B . дои : 10.1017/S0954102010000866 . ISSN 1365-2079 . S2CID 130084588 .
- ^ Чендлер, Массачусетс; Ринд, Д.; Руди, Р. (1992). «Пангейский климат в ранней юре: моделирование GCM и осадочная летопись палеоклимата». Бюллетень Геологического общества Америки. 104 (5): 543.
- ^ Кантрилл, Дэвид Дж.; Пул, Имоджен (2012). Растительность Антарктиды в геологическом времени . Кембридж: Издательство Кембриджского университета. дои : 10.1017/cbo9781139024990 . ISBN 978-0-521-85598-3 .
- ^ Кантрилл, Дэвид Дж.; Хантер, Мораг А. (2005). «Макрофоссильные флоры бассейна Латади, Антарктический полуостров» . Новозеландский журнал геологии и геофизики . 48 (3): 537–553. дои : 10.1080/00288306.2005.9515132 . ISSN 0028-8306 . S2CID 129854482 .
- ^ Пол, Грегори С. (2016). Принстонский полевой справочник по динозаврам (2-е изд.). Издательство Принстонского университета . стр. 182–183. ISBN 978-0-691-16766-4 .











