Восточная Гондвана

Южный полярный регион мелового периода включал в себя континент Восточная Гондвана – современные Австралия, Зеландия и Антарктида – продукт распада Гондваны в меловой период . В южном регионе в это время было намного теплее, чем сегодня: температура колебалась примерно в диапазоне 4–8 ° C (39–46 ° F) в позднем меловом периоде Маастрихта на территории нынешней юго-восточной Австралии. Это препятствовало развитию постоянных ледниковых щитов и способствовало развитию полярных лесов , в которых в основном преобладали хвойные деревья , саговники и папоротники , и которые зависели от умеренного климата и сильных дождей. содержащими окаменелости Основными геологическими образованиями, , которые отмечают эту область, являются: формации Санта-Марта и Собрал на острове Сеймур у Антарктического полуострова ; остров Сноу-Хилл , Лопес-де-Бертодано и формации Скрытого озера на острове Джеймса Росса, также недалеко от Антарктического полуострова; и формации Эумералла и Вонтагги в Австралии.
В южнополярном регионе обитало множество эндемичных видов, в том числе несколько реликтовых форм, вымерших в других местах к меловому периоду. Из комплекса динозавров наиболее разнообразными были мелкие гипсилофодонтоподобные динозавры. Южный полярный регион также был домом для последней амфибии -лабиринтодонта , Koolasurus . Изоляция Антарктиды создала особую экосистему морской жизни, названную Провинцией Уэдделлиана . [ 1 ]
Пейзаж
[ редактировать ]Геология
[ редактировать ]
Бассейн Латади на юге Антарктического полуострова ( Земля Палмера) содержит вулканические породы , что свидетельствует о крупном магматическом событии в среднем меловом периоде, известном как событие Земли Палмера. В этой области обнаружено множество окаменелостей макро- и микрорастений , а также моллюсков, характерных для раннего мела , а также среднего и позднего юрского периода . [ 3 ] [ 4 ] Область, которая сейчас является Антарктическим полуостровом, на протяжении всего мезозоя погружала прото-Тихоокеанскую плиту , вызывая вулканическую активность. [ 5 ]
Важным ископаемым образованием является формация Санта-Марта на острове Джеймса Росс у Антарктического полуострова , одна из наиболее значительных формаций Южного полушария , представляющая большое разнообразие наземной флоры и фауны полярного региона позднего мела . Остров Сеймур похож в этом отношении: это область открытия позвоночных и беспозвоночных существ, таких как плезиозавры , позднего мела, простирающегося до эоцена . [ 4 ] Другими меловыми образованиями, содержащими окаменелости, в Антарктиде являются формация острова Сноу-Хилл , формация Лопес де Бертодано и формация Хидден-Лейк , также на острове Джеймса Росса. [ 6 ] [ 7 ]
в Формация Эумералла бухте динозавров и формация Вонтагги в Виктории, Австралия, содержат ряд окаменелостей и следов динозавров раннего мелового периода. Формация Тахора представляет собой совокупность рептилий меловой Новой Зеландии . [ 7 ]
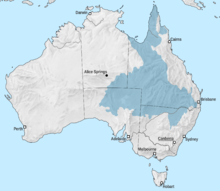
Море Эроманга было внутренним морем на территории будущей Австралии, образовавшимся в раннем меловом периоде. Море достигло бассейна Эроманга с севера через бассейн Карпентари . Южный конец моря включал лагуны и реки, а на востоке, в бассейне Сурата, был залив. Когда в раннем меловом периоде Индия отошла от Австралии, Пертский бассейн также наполнился морской водой. Область, которая должна была стать Коралловым морем, представляла собой рифтовую долину . Когда в течение мелового периода Австралия и Антарктида разошлись, между ними образовалось море. [ 8 ]
Растительность
[ редактировать ]Меловой период характеризуется теплыми глобальными температурами, вызванными большим количеством углекислого газа и, возможно, метана, парниковых газов в атмосфере. Это привело к отсутствию постоянного ледяного покрова в полярных регионах, хотя уровень углекислого газа упал между 115 и 66 миллионами лет назад (млн лет назад), что, возможно, привело к появлению некоторого постоянного ледяного покрова. Возможно, на протяжении мелового периода образовалось несколько небольших ледниковых щитов. [ 9 ] Повышенная тектоническая активность , вызывающая усиление геотермального тепла , возможно, предотвратила оледенение и повышение глобальной температуры. Температура могла быть на 15 ° C (27 ° F) выше, чем в наше время. [ 10 ] [ 11 ] [ 12 ]
юрский период
[ редактировать ]Ландшафт среднеюрской полярной области реконструирован по остаткам обнаженных в Новой Зеландии лесов, которые в юрский период находились между 70 и 80° ю.ш. Растительность в основном состояла из хвойных деревьев, саговников и других голосеменных растений , а также папоротников; на лесной подстилке росли плауны , мохообразные , грибы и водоросли. Здесь был умеренный климат с обильными осадками. [ 13 ] [ 14 ]
Ранний мел
[ редактировать ]В раннем меловом периоде Восточная Гондвана (Австралия, Антарктида и Зеландия ) начала отделяться от Южной Америки, а Индия и Мадагаскар также начали отделяться примерно в то же время. [ 9 ] Тропическая зона , возможно, расширилась в меловой период до 32° ю.ш. , что позволило круглый год расти деревьям в Антарктике в полярных лесах . Обнаружение нескольких взрослых вечнозеленых и лиственных деревьев указывает на температуру от теплой до прохладной с умеренными сезонами без повсеместного замерзания, по крайней мере, между южной широты 70 и 85 ° . [ 15 ] [ 16 ] Однако также возможно, что растительная жизнь может быть характерна только для теплых летних месяцев. [ 17 ] Большая часть того, что известно о растительной жизни Восточной Гондваны в меловой период, состоит из остатков пыльцы и компрессов листьев с северной части Антарктического полуострова . [ 18 ] В зависимости от широты полярные зимы могли длиться от шести недель до четырех с половиной месяцев. [ 19 ]

Однако породы из раннемеловой формации Вонтхаджи на юго-востоке Австралии свидетельствуют о сезонно замерзшей почве. [ 21 ] Его географическое положение в раннем меловом периоде на отметке около 78° ю.ш. указывает на то, что зимой в этой области было от одного до трех месяцев темноты, и эта область представляет собой пойму с ледниковым питанием . [ 22 ] Свидетельства раннемелового оледенения и холодного климата были обнаружены в отложениях в бассейне Эроманга на территории современной центральной Австралии, или от 60 до 80 ° ю.ш. в раннем мелу. этот бассейн представлял собой большое внутреннее море В меловой период . Тем не менее, подобные образования могли быть созданы просто селевым потоком, поэтому вполне возможно, что оледенения здесь никогда не было. [ 23 ] [ 24 ] Общий покров полярных льдов в мезозое, возможно, составлял треть того размера, который есть в наше время, хотя резкие похолодания с отрицательными температурами, возможно, случались в течение всего раннего мела. [ 25 ]
Поздний мел
[ редактировать ]Полог полярных лесов вокруг современного острова Александра , который в среднем мелу находился примерно на 75° ю.ш. , был преимущественно вечнозеленым, и, вероятно, большинство южнополярных лесов тоже были такими же и состояли в основном из араукариевых и подокарповых хвойных пород. [ 4 ] [ 26 ] [ 16 ] Считается, что эти деревья оставались в состоянии покоя на протяжении всей полярной зимы до лета под полуночным солнцем . [ 27 ]
Свидетельства цветения растений, датируемые примерно 80 млн лет назад в позднем меловом периоде, позволяют предположить существование лесов умеренного пояса , подобных тем, что существуют в современной Австралии, Новой Зеландии и на юге Южной Америки. [ 20 ] Некоторые остатки цветов были обнаружены около 60° ю . F) и дождливый климат. [ 16 ] [ 18 ]
Остатки пыльцы из юго-восточной Австралии идентичны живым видам растений Австралии: хвойным, цветковым растениям, населяющим районы с большим количеством осадков и средиземноморским климатом , а также склерофильным кустарникам , что указывает на уникальный ландшафт тропических лесов и открытых кустарников. [ 28 ] Судя по размеру годичных колец на ископаемых деревьях, в антарктических полярных лесах наблюдалась тенденция к похолоданию на протяжении всего маастрихтского периода ( 72–66 млн лет назад), от среднегодовой температуры 7 ° C (45 ° F) до более сезонно экстремальных 4–6 млн лет назад. 8 ° C (39–46 ° F). [ 29 ] Эти растения, вероятно, пережили мел-палеогеновое вымирание , которое уничтожило большую часть жизни 66 млн лет назад на вулканическом Антарктическом полуострове ; окаменелости растений, датируемые 60 млн лет назад в палеоцене с острова Сеймур у этого полуострова, признаны предками растений умеренного пояса, населяющих современную Австралию и Южную Америку. [ 27 ]
Позднемеловые ( кампанские ) формации Замек и Половина Трех Пойнт острова Кинг-Джордж были расположены на 60 ° ю.ш. и демонстрируют богатую совокупность ископаемой флоры, такой как Podocarpus ; Араукария ; лептоспорангиатные папоротники Cladophlebis и Clavifera ; и различные цветковые растения Magnoliopsida , Dicotylophyllum , Myrciophyllum santacruzensis , Nothofagus , Sterculiaephyllum australis , Monimiophyllum и так далее. [ 30 ] [ 31 ] [ 32 ] Формация Собрал на острове Сеймур, охватывающая границу мела и палеогена на палеошироте 63 ° южной широты, предоставила новый род ископаемых цветов семейства Cunoniaceae , Eucryphiaceoxylon eucryphioides . [ 33 ]
Экология
[ редактировать ]Как и в сегодняшней Австралии, Восточная Гондвана была домом для многих эндемичных животных, в том числе многих реликтовых видов семейств, вымерших в остальной части мелового периода. Возможно, полярные регионы позднего мела были заселены группами растений и животных, происхождение которых можно проследить до ордовика . [ 16 ] [ 34 ] Постепенная изоляция Антарктиды в позднем меловом периоде привела к появлению отдельной группы водных существ, названной Провинцией Уэдделлиана. [ 35 ]
Динозавры
[ редактировать ]Птицы
[ редактировать ]
Останки предка современных птиц, неорнитов , редко встречаются в мезозое, при этом большая радиация приходится на неоген Антарктиды. позднемеловой Vegavis , птицы, похожей на гуся Однако обнаружение на острове Вега , указывает на то, что основные современные группы птиц уже были распространены в меловом периоде. На острове Вега также была обнаружена бедренная кость, принадлежащая неопознанной сериемоподобной птице. Следы птиц сохранились в бухте динозавров и, поскольку они крупнее, чем у большинства видов птиц мелового периода, указывают на обилие более крупных птиц-энантиорнитов или птиц -орнитуринов в раннем меловом периоде. [ 36 ]
две ныряющие птицы, возможные примитивные ныряльщики В позднемеловом Чили и Антарктиде были обнаружены : Neogaeornis и Polarornis . Полярорнис, возможно, был способен как нырять, так и летать. [ 36 ] Самые ранние пингвины , Кроссваллия и Вайману , известны в период 61–62 млн лет назад в палеоцене , однако молекулярные данные позволяют предположить, что пингвины впервые появились в позднем меловом периоде . Учитывая, что эти пингвины были датированы очень близко к мел-палеогеновому вымиранию , группа эволюционировала либо до этого события, либо очень быстро после него. [ 36 ] [ 37 ]
нептичий
[ редактировать ]
Окаменелости динозавров редки в Южном полярном регионе, а основными местами, содержащими окаменелости, являются группа островов Джеймса Росса ; Ледник Бердмор в Антарктиде; Рома, Квинсленд ; Ручей Мангахуанга в Новой Зеландии; и Бухта Динозавров в Виктории, Австралия . [ 39 ] Останки динозавров этого региона, например, найденные в Виктории, состоят лишь из фрагментарных частей, что делает идентификацию спорной. Например, спорные идентификации аллозаврид , который может представлять собой абелизаврида , цератопсового серендипацератопса , который мог быть анкилозавром трудно классифицировать . , и теропода Тимимуса, которого были сделаны [ 38 ]
Суперконтинент Пангея юрского периода позволил основным кладам динозавров достичь глобального распространения перед распадом, и между южнополярными формами и формами, обнаруженными в других местах, несмотря на разделение океаном Тетис , существовало несколько близкородственных родственников . Однако группам динозавров, достигшим пан-Гондванского распространения в течение мелового периода, пришлось бы использовать сухопутный мост, соединяющий Австралию с Южной Америкой через Антарктиду в южнополярном регионе. [ 34 ] Южнополярный игуанодонтий Muttaburrasaurus наиболее тесно связан с европейскими рабдодонтидами , которые были доминирующей группой в Европе в позднем меловом периоде. Меловой южнополярный кунбарразавр идентифицирован как часть уникальной линии гондванских анкилозавров, что предполагает однородность фауны Гондваны. [ 40 ] [ 41 ] Дромеозавриды . известны из Антарктиды и представляют собой реликтовую популяцию, ранее распространенную по всему миру [ 42 ] Несмотря на эти очевидные межконтинентальные миграции, маловероятно, что южнополярные динозавры мигрировали из полярных лесов зимой, поскольку они были либо слишком массивными, как анкилозавры, либо слишком маленькими, как троодонтиды , чтобы путешествовать на большие расстояния, и большое море между Восточной Гондваной и другими континентами препятствовало любым таким миграциям в позднем мелу. [ 43 ] [ 44 ] Возможно, чтобы справиться с зимними условиями, некоторые динозавры впадали в спячку , например теропод Тимимус . [ 45 ]

Наиболее распространенной и разнообразной группой, обнаруженной на данный момент, являются гипсилофодонтоподобные динозавры, составляющие половину таксонов динозавров, обнаруженных в юго-восточной Австралии, что не встречается в более тропических регионах, что, возможно, указывает на какое-то преимущество перед другими динозаврами на полюсах. [ 16 ] Будучи маленькими и со скрежетающими зубами, они, вероятно, питались низменной растительностью, такой как плауноногие и семенные коробочки подокарпов . [ 46 ] Гипсилофодонтоподобный Leaellynasaura имел большие глазницы , большие, чем у большинства тропических гипсилофодонтоподобных динозавров, и, возможно, обладал острым ночным зрением , что позволяет предположить, что Leaellynasaura и, возможно, другие гипсилофодонтоподобные динозавры жили в полярных районах круглый год или большую часть времени. года, включая полярные зимы. Рост костей был непрерывным на протяжении всей его жизни, что указывает на то, что он не впадал в спячку, возможно, потому, что он был эндотермическим или пойкилотермным . [ 39 ] [ 47 ] или рыть норы. [ 48 ] Однако возможно, что большие глаза возникли просто в результате онтогенеза , то есть относительно большие глазницы могли быть особенностью, наблюдаемой только у молодых особей, или, возможно, были врожденным дефектом, поскольку известен только один экземпляр. [ 43 ] [ 19 ]
Хотя останки скудны и, следовательно, таксономические описания могут быть сомнительными, [ 49 ] останки викторианских теропод были отнесены к семи различным кладам: Ceratosauria , Megaraptora , Tyrannosauroidea и Maniraptora . Однако тираннозавроиды не известны на других континентах Гондваны и более известны из северной Лавразии . [ 50 ] В отличие от других континентов Гондваны, высшими хищниками которых были абелизавриды и кархародонтозавриды , открытие австраловенатора , рапатора и неназванного вида в Австралии позволяет предположить, что мегараптораны были главными хищниками Восточной Гондваны. Хвостовые позвонки неизвестного теропода, получившего название « теропод Джоан Виффен », были обнаружены в позднеюрских /раннемеловых породах Новой Зеландии. [ 51 ]

Четыре титанозавра — австралотитан , саванназавр , диамантиназавр и винтонотитан — обнаруженные в формации Винтон , составляют комплекс зауроподов меловой Австралии, однако эти существа, вероятно, избегали полярных регионов, поскольку их останки полностью отсутствуют в Юго-Восточной Австралии, которая находилась в пределах Южной полярной зоны. регион в меловом периоде. Однако вполне вероятно, что, по крайней мере, титанозавры мигрировали в Австралию из Южной Америки, что потребовало бы от них прохождения через Антарктиду, поскольку титанозавры появились в меловом периоде после распада Пангеи. Вполне возможно, что событие Бонарелли в среднем меловом периоде сделало Антарктиду более теплой и, следовательно, более гостеприимной для зауроподов. [ 50 ] Эти динозавры, вероятно, питались мясистыми семенами подокарповых и тисовых деревьев, а также обычными раздвоенными папоротниками того времени. [ 46 ]
Палеоцен
[ редактировать ]Считается, что после удара астероида наступившая ударная зима уничтожила динозавров вместе с большей частью мезозойской жизни в результате мел-палеогенового вымирания . Однако отсутствие горизонта резкого вымирания в отложениях Антарктики или Австралии для окаменелостей растений и двустворчатых моллюсков в этот период времени указывает на менее мощное воздействие в южнополярном регионе. [ 16 ] Учитывая, что динозавры и другая фауна полярных регионов мелового периода были хорошо приспособлены к жизни в длительные периоды темноты и холодной погоды, было высказано предположение, что это сообщество могло пережить это событие. [ 19 ]
Реки и озера
[ редактировать ]Последние темноспондилы – группа гигантских земноводных , вымерших преимущественно после триаса – населяли Южную полярную область в раннем мелу. Koolasurus , возможно, последний из темноспондилов, как полагают, выжил в регионах, где было слишком холодно для их конкурентов, неозухов – группы рептилий, в состав которой входят современные крокодилы , – которые неактивны в воде при температуре ниже 10 °C (50 °F). , чтобы выжить. [ 52 ] Хотя неозухи известны из меловой Австралии, считается, что они держались подальше от полярного региона, прибывая в Австралию по морю, а не по суше. [ 34 ]

Вполне вероятно, что темноспондилы населяли пресноводные системы полярной Австралии до тех пор, пока событие Бонарелли в среднем мелу около 100 млн лет назад не повысило температуру и не позволило неозухиам заселить Антарктиду. Эти неозухи, взрослая особь которых не превышала 2,5 метра (8,2 фута) в длину, вероятно, привели к вымиранию темноспондилов в тандеме с более развитыми лучеперыми рыбами , которые, возможно, нацелились на их личинки. Миграция неозухий в регион предполагает, что средние зимние температуры превышали 5,5 ° C (41,9 ° F), а среднегодовая температура превышала 14,2 ° C (57,6 ° F). Однако полярные неозухи известны только по почти полному скелету Isisfordia , а другие останки неозухий относятся к неопределенным видам. [ 52 ]
Плезиозавры населяли системы пресноводных рек и эстуариев, учитывая расположение их останков, вероятно, колонизируя Австралию в ранней и средней юре. Их останки, в основном зубы, были зарегистрированы в юго-восточной Австралии и относятся к позднему мелу, хотя они никогда не были описаны, поскольку останки слишком редки, чтобы это можно было сделать. Зубы имеют некоторое сходство с плиозаврами , особенно с ромалеозавридами и лептоклеидами , которые вымерли в раннем меловом периоде, что указывает на то, что полярные пресноводные системы могли быть убежищем для плиозавров мелового периода. В отличие от современных морских рептилий, эти южнополярные плезиозавры, вероятно, лучше переносили более холодные воды. [ 53 ] [ 24 ]
Океаны
[ редактировать ]
раннего и среднего мела Останки морских рептилий Южной Австралии включают пять семейств плезиозавров — Cryptoclididae , Elasmosauridae , Polycotylidae , Rhomaleosauridae и Pliosauridae — и ихтиозавров семейство Ophthalmosauridae . Обнаружение нескольких останков молодых плезиозавров позволяет предположить, что они использовали богатые питательными веществами воды побережья в качестве защищенных мест для отела, а холод отпугивал хищников, таких как акулы. Большинство обнаруженных плезиозавров имели космополитическое распространение , однако там существовали эндемичные формы, такие как Opallionectes и возможный новый вид криптоклидидов. вид Сомнительный эласмозавра Woolungasaurus , названный в 1928 году, является одним из самых ранних описаний австралийской морской рептилии. нескольких моллюсков , брюхоногих моллюсков , аммонитов , костистых рыб , химерид и кальмароподобных белемнитов . Также были обнаружены останки [ 53 ] Прибрежная зона могла подвергнуться зимнему замерзанию, и эти рептилии в ответ могли мигрировать зимой на север, иметь более активный обмен веществ , чем тропические рептилии, впадать в спячку в пресноводных районах, подобно современному американскому аллигатору ( Alligator Mississippiensis ), или были эндотермическими, как современные кожистые морские черепахи ( Dermochelys coriacea ). Меньшее количество плезиозавров и большее количество ихтиозавров и морских черепах в более северных районах Австралии указывает на предпочтение плезиозаврами более холодных районов. [ 53 ]

Несколько позднемеловых океанических плезиозавров и мозазавров были обнаружены в Новой Зеландии и Антарктиде, причем некоторые из них, такие как Mauisaurus , являются эндемичными, а другие, такие как Prognathodon , имеют космополитическое распространение. [ 54 ] [ 55 ] Эласмозавры и плиозавры известны от одного до трех видов из этого района. Открытие трёх криптоклидид в Южном полушарии — Morturneria из Антарктиды, Aristonectes из Южной Америки и Kaiwhekea из Новой Зеландии — указывает на диверсификацию семейства в позднем мелу этого региона и, возможно, на увеличение продуктивности раннего Южного океана . [ 56 ]
Птерозавры
[ редактировать ]
Две клады птерозавров представлены в раннемеловой Австралии, Pteranodontoidea и Ctenochasmatoidea , останки которых происходят в основном из формации Тулебук и районов Квинсленда и Нового Южного Уэльса. Считается, что в меловой Австралии существовало как минимум шесть таксонов птерозавров, однако, учитывая фрагментарный характер останков, многие обнаруженные окаменелости происходят от неустановленного птерозавра. Окаменелости были обнаружены на мелководье и в лагунах, что указывает на рацион рыб и других водных обитателей. Ктенохасматиды дожившими были единственными археоптеродактилоидами, до мелового периода. Единственные останки зубов птерозавров, обнаруженные в Австралии и относящиеся к раннему меловому периоду, принадлежат Mythunga и, возможно, позднемеловому анангериду . По оценкам, мифунга имел размах крыльев 4,5 метра (15 футов), что намного больше, чем у любого другого обнаруженного археоптеродактилоида, хотя возможно, что птерозавр больше связан с Anhangueridae или Ornithocheiridae . [ 57 ] [ 58 ] [ 59 ] Тем не менее, птерозавры по-прежнему существуют в неполярных регионах Австралии, учитывая их способность мигрировать по воздуху, возможно, им не нужно было пересекать сухопутный мост через полярные регионы, чтобы добраться туда, то есть они никогда не населяли южную полярную область. область. [ 16 ] [ 34 ]
Из позднемеловых птерозавров только останки, принадлежащие к семейству Azhdarchidae , найденные в бассейнах Карнарвона и Перта в Западной Австралии были отнесены к таксону . Возможный представитель Ornithocheiridae был обнаружен в позднем мелу Западной Австралии, хотя ранее считалось, что это семейство вымерло в раннем мелу. [ 57 ] [ 58 ] [ 59 ]
Млекопитающие
[ редактировать ]
В раннемеловой Австралии обнаружено семь млекопитающих: неописанный орниторинхид , Kryoryctes , Kollikodon , Ausktribosphenos , Bishops , Steropodon и Corriebaatar ; все они в то время были эндемиками Австралии. Вероятно, млекопитающие пересекли антарктический сухопутный мост между Австралией и Южной Америкой в раннем меловом периоде, и, вероятно, предки эндемичных млекопитающих Австралии прибыли в юрский период через суперконтинент Пангея . [ 60 ] [ 61 ]
Беспозвоночные
[ редактировать ]несколько окаменелостей насекомых и ракообразных Из южнополярных меловых отложений Новой Зеландии известно . Позднемеловой мангаотанский конгломерат Монро располагался на 68° ю.ш. и содержал окаменелости Helastia sp. , [ 62 ] а краб Hemioon novozelandicum был обнаружен в алевролитах Суэйла , расположенных на 76 ° ю.ш., в позднем альбе . [ 63 ] Несколько экземпляров насекомых были также обнаружены в формации Тупуанги островов Чатем на широте 79° ю.ш. в период сеномана - турона . [ 64 ]
См. также
[ редактировать ]- Бухта Динозавров
- Динозавр мечтает
- Геология Антарктиды , включая палеонтологию
- Список австралийских и антарктических динозавров
- Тропические леса Гондваны
- Полярные леса мелового периода.
- « Духи ледяного леса », 5 серия « Прогулок с динозаврами».
Ссылки
[ редактировать ]- ^ Отеро, Родриго А.; Сото-Коин, Серджио; С, Кристиан Салазар; Оярсун, Хосе Луис (27 марта 2015 г.). «Новые эласмозавриды (Sauropterygia, Plesiosauria) из позднего мела бассейна Магалланес, Чилийская Патагония: свидетельства круговорота фауны в течение маастрихта вдоль биогеографической провинции Уэдделл» . Андская геология . 42 (2): 237–267. дои : 10.5027/andgeoV42n2-a05 . ISSN 0718-7106 .
- ^ «Бухта динозавров, Австралия» . Эта Динамическая Земля . Геологическая служба США . Проверено 14 марта 2015 г.
- ^ Воган, APM; Иглз, Г.; Флауэрдью, MJ (2012). «Свидетельства двухфазного события на Земле Палмера на основе пересекающихся структурных связей и времени размещения интрузивной свиты побережья Ласситера, Антарктический полуостров: последствия для конфигурации плит Южного океана в середине мелового периода» (PDF) . Тектоника . 3 (1): TC1010. Бибкод : 2012Tecto..31.1010V . дои : 10.1029/2011TC003006 . S2CID 129316703 .
- ^ Jump up to: а б с Риффенбург 2007 , с. 72.
- ^ Риффенбург 2007 , с. 551.
- ^ Пирри, Д.; Крам, Дж.А.; Ломас, ЮАР; Верховая езда, Джей Би (1997). «Позднемеловая стратиграфия региона Адмиралти-Саунд, бассейн Джеймса Росс, Антарктида». Меловые исследования . 18 (1): 109–137. дои : 10.1006/cres.1996.0052 .
- ^ Jump up to: а б Вейшампель, Дэвид Б. (2004). «Распространение динозавров (ранний мел, Австралазия)». В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 573–574 . ISBN 978-0-520-24209-8 .
- ^ Деттманн, Мэн; Мольнар, Р.Э.; Дуглас, Дж. Г. (1992). «Австралийская меловая наземная фауна и флора: биостратиграфические и биогеографические значения». Меловые исследования . 13 (3): 207–262. дои : 10.1016/0195-6671(92)90001-7 .
- ^ Jump up to: а б Хэй, WW (2008). «Развивающиеся представления о меловом климате и циркуляции океана». Меловые исследования . 29 (5): 725–753. дои : 10.1016/j.cretres.2008.05.025 .
- ^ Ладан, Дж.; Доннадье, Ю. (2016). «Палеогеографическая регламентация ледниковых событий во время меловой супероранжереи» . Природные коммуникации . 7 : 12771. Бибкод : 2016NatCo...712771L . дои : 10.1038/ncomms12771 . ПМК 5036002 . ПМИД 27650167 .
- ^ Берлинг, диджей; Осборн, CP (2002). «Физиологическая экология мезозойских полярных лесов в среде с высоким содержанием CO2» . Анналы ботаники . 89 (3): 329–339. дои : 10.1093/aob/mcf045 . ПМЦ 4233824 . ПМИД 12096745 .
- ^ Перальта-Медина, Э.; Фалькон-Ланг, HJ (2012). «Состав и продуктивность меловых лесов получены на основе глобальной базы данных ископаемой древесины». Геология . 40 (3): 219–222. Бибкод : 2012Geo....40..219P . дои : 10.1130/G32733.1 .
- ^ Торн, В. (2001). «Растительные сообщества среднеюрских лесов высокой палеошироты в Новой Зеландии». Палеогеография, Палеоклиматология, Палеоэкология . 168 (3–4): 273–289. Бибкод : 2001PPP...168..273T . дои : 10.1016/S0031-0182(01)00203-6 .
- ^ Поул, М. (1999). «Структура леса приполярных широт новозеландской юры». Палеогеография, Палеоклиматология, Палеоэкология . 147 (1–2): 121–139. Бибкод : 1999PPP...147..121P . дои : 10.1016/S0031-0182(98)00151-5 .
- ^ Дуглас, Дж. Г.; Уильямс, GE (1982). «Южные полярные леса: раннемеловые флоры Виктории и их палеоклиматическое значение». Палеогеография, Палеоклиматология, Палеоэкология . 39 (3–4): 171–185. Бибкод : 1982PPP....39..171D . дои : 10.1016/0031-0182(82)90021-9 .
- ^ Jump up to: а б с д и ж г Крейм, Дж. А. (1992). «Позднемеловая палеосреда и биота: антарктическая перспектива». Антарктическая наука . 4 (4): 371–382. Бибкод : 1992AntSc...4..371C . дои : 10.1017/S0954102092000555 . S2CID 140637028 .
- ^ Пирес, ЭФ; Герра-Соммер, М. (2011). «Анализ годовых колец ископаемых хвойных пород раннего мела бассейна Арарипе (Бразилия)» (PDF) . Анналы Бразильской академии наук . 83 (2): 409–423. дои : 10.1590/S0001-37652011005000005 . ПМИД 21625801 .
- ^ Jump up to: а б Пул, И.; Готвальд, Х.; Фрэнсис, JE (2000). « Иллисиоксилон , элемент полярных лесов Гондваны? Позднемеловые и раннетретичные леса Антарктиды» . Анналы ботаники . 86 (2): 421–432. дои : 10.1006/anbo.2000.1208 .
- ^ Jump up to: а б с Викерс-Рич, П .; Рич, TH (1993). «Полярные динозавры Австралии». Научный американец . 269 (1): 50–55. Бибкод : 1993SciAm.269a..50V . doi : 10.1038/scientificamerican0793-50 . JSTOR 24941547 .
- ^ Jump up to: а б Готвальд, Х. (2001). « Monimiaceae sensu lato , элемент полярных лесов Гондваны: данные из позднемеловой-раннетретичной древесной флоры Антарктиды». Австралийская систематическая ботаника . 14 (2): 207–230. дои : 10.1071/SB00022 .
- ^ Константин, А.; Чинсами-Туран, А.; Рич, П.В.; Рич, Т. (1998). «Перигляциальная среда и полярные динозавры». Южноафриканский научный журнал . 94 (3): 137–141.
- ^ Клоуз, РА; Викер-Рич, П.; Труслер, П.; Кьяппе, LM; О'Коннор, Цзинмай; Рич, TH; Кул, Л.; Комарауэр, П. (2008). «Самая ранняя гондванская птица из мелового периода юго-востока Австралии». Журнал палеонтологии позвоночных . 29 (2): 616–619. дои : 10.1671/039.029.0214 . S2CID 130306474 .
- ^ Элли, Северная Каролина; Фрейкс, Луизиана (2003). «Первое известное меловое оледенение: член Ливингстон-Тиллит формации Кадна-Оуи, Южная Австралия». Австралийский журнал наук о Земле . 50 (2): 139–144. Бибкод : 2003AuJES..50..139A . дои : 10.1046/j.1440-0952.2003.00984.x . S2CID 128739024 .
- ^ Jump up to: а б Кир, BP (2004). «Останки плезиозавра из меловых высокоширотных неморских отложений Юго-Восточной Австралии». Журнал палеонтологии позвоночных . 26 (1): 196–199. doi : 10.1671/0272-4634(2006)26[196:PRFCHN]2.0.CO;2 . S2CID 130794388 .
- ^ Прайс, ГД (1999). «Свидетельства и последствия существования полярных льдов в мезозое» (PDF) . Обзоры наук о Земле . 48 (3): 183–210. Бибкод : 1999ESRv...48..183P . дои : 10.1016/S0012-8252(99)00048-3 .
- ^ Фалькон-Ланг, HJ; Кантрилл, диджей (2001). «Фенология листьев некоторых среднемеловых полярных лесов, остров Александра, Антарктида». Геологический журнал . 138 (1): 39–52. Бибкод : 2001GeoM..138...39F . дои : 10.1017/S0016756801004927 . S2CID 129461955 .
- ^ Jump up to: а б Риффенбург 2007 , с. 413.
- ^ Деттманн, Мэн; Ярзен, DM (1991). «Пыльцевые доказательства позднемеловой дифференциации протейных в южных полярных лесах». Канадский журнал ботаники . 69 (4): 901–906. дои : 10.1139/b91-116 .
- ^ Фрэнсис, JE; Пул, И. (2002). «Меловой и раннетретичный климат Антарктиды: данные из ископаемой древесины» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 182 (1–2): 47–64. Бибкод : 2002PPP...182...47F . дои : 10.1016/S0031-0182(01)00452-7 .
- ^ Сборка на Fossilworks.org Касл- Хилл
- ^ Сборка Skua на Fossilworks.org Bay
- ^ Сборка Half Three на Fossilworks.org Point
- ^ Sobral Formation at Fossilworks .org
- ^ Jump up to: а б с д Мольнар, Р.Э. (1989). «Наземные четвероногие в меловой Антарктиде». В Крейн, Дж. А. (ред.). Происхождение и эволюция антарктической биоты . Том. 47. Специальные публикации Геологического общества. стр. 131–140. Бибкод : 1989ГСЛСП..47..131М . CiteSeerX 10.1.1.1010.6989 . дои : 10.1144/ГСЛ.СП.1989.047.01.11 . S2CID 140579588 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Риффенбург 2007 , с. 146.
- ^ Jump up to: а б с д Тамбусси, К.; Госпиталече, Калифорния (2007). «Антарктические птицы (неорниты) в мел-эоценовое время» (PDF) . Журнал Геологической ассоциации . 62 (4): 604–617.
- ^ Хакетт, С.Дж.; Кимбалл, RT; Редди, С.; Боуи, RCK; Браун, Эль; Браун, MJ (2008). «Филогеномное исследование птиц раскрывает их эволюционную историю». Наука . 320 (5884): 1763–1768. Бибкод : 2008Sci...320.1763H . дои : 10.1126/science.1157704 . ПМИД 18583609 . S2CID 6472805 .
- ^ Jump up to: а б Бенсон, РБЖ; Рич, TH; Викерс-Рич, П.; Холл, М. (2012). «Фауна теропод из Южной Австралии указывает на высокое полярное разнообразие и обусловленную климатом провинциальность динозавров» . ПЛОС ОДИН . 7 (5): e37122. Бибкод : 2012PLoSO...737122B . дои : 10.1371/journal.pone.0037122 . ПМЦ 3353904 . ПМИД 22615916 .
- ^ Jump up to: а б Рич, TH; Викерс-Рич, П.; Ганглофф, Р.А. (2002). «Полярные динозавры» (PDF) . Наука . 295 (5557): 979–980. дои : 10.1126/science.1068920 . ПМИД 11834803 . S2CID 28065814 .
- ^ Сото-Акунья, Серджио; Варгас, Александр О.; Калуза, Джонатан; Леппе, Марсело А.; Ботельо, Жоау Ф.; Пальма-Либерона, Хосе; Саймон-Гутштейн, Кэролайн; ФЕРНАНДЕС, Рой А.; Ортис, Эктор; МИЛА, Вероника; Аравена, Барбара (декабрь 2021 г.). «Причудливое хвостовое вооружение переходного анкилозавра из субантарктического Чили» . Природа . 600 (7888): 259–263. Бибкод : 2021Natur.600..259S . дои : 10.1038/s41586-021-04147-1 . ISSN 1476-4687 . ПМИД 34853468 . S2CID 244799975 .
- ^ Фрауэнфельдер, Тимоти Г.; Белл, Фил Р.; Бруэм, Том; Бевитт, Джозеф Дж.; Бикнелл, Рассел, округ Колумбия; Кир, Бенджамин П.; Горе, Стивен; Кампионе, Николас Э. (28 марта 2022 г.). «Новые черепные остатки анкилозавров из формации Тулебук нижнего мела (верхний альб) в Квинсленде, Австралия» . Границы в науках о Земле . 10 . дои : 10.3389/feart.2022.803505 . ISSN 2296-6463 .
- ^ Кейс, Дж. А.; Мартин, Дж. Э.; Регеро, М. (2007). «Дромеозавр из маастрихта острова Джеймса Росс и позднемеловая антарктическая фауна динозавров». Геологическая служба США и Национальные академии . CiteSeerX 10.1.1.546.3890 .
- ^ Jump up to: а б Ричардс, THV; Ричардс, TH; Викерс-Рич, П. (2000). «Откуда мы знаем, куда мы идем?» . Динозавры тьмы . Издательство Университета Индианы. ISBN 978-0-253-33773-3 .
- ^ Белл, PR; Снавли, Э. (2007). «Полярные динозавры на параде: обзор миграции динозавров» . Алчеринга: Австралазийский журнал палеонтологии . 32 (3): 271–284. дои : 10.1080/03115510802096101 .
- ^ Чинсами, А.; Рич, Т.; Викерс-Рич, П. (1998). «Гистология костей полярных динозавров». Журнал палеонтологии позвоночных . 18 (2): 385–390. дои : 10.1080/02724634.1998.10011066 .
- ^ Jump up to: а б Деттманн, Мэн; Мольнар, Р.Э.; Дуглас, Дж. Г.; Бургер, Д.; Филдинг, К. (1992). «Австралийская меловая наземная фауна и флора: биостратиграфические и биогеографические значения». Меловые исследования . 13 (3): 207–262. дои : 10.1016/0195-6671(92)90001-7 .
- ^ Вудворд, Х.Н.; Рич, TH; Чинсами, А.; Викерс-Рич, П. (2011). «Динамика роста полярных динозавров Австралии» . ПЛОС ОДИН . 6 (8): e23339. Бибкод : 2011PLoSO...623339W . дои : 10.1371/journal.pone.0023339 . ПМК 3149654 . ПМИД 21826250 .
- ^ Мартин, Эй Джей (2009). «Норы динозавров в группе Отвей (Альб) в Виктории, Австралия, и их связь с полярной средой мелового периода». Меловые исследования . 30 (5): 1223–1237. дои : 10.1016/j.cretres.2009.06.003 .
- ^ Уайт, Массачусетс; Белл, PR; Кук, АГ; Поропат, Сан-Франциско; Эллиотт, Д.А. (2015). «Дентария Australovenator wintonensis (Theropoda, Megaraptoridae); значение для зубного ряда мегарапторидов» . ПерДж . 3 : е1512. дои : 10.7717/peerj.1512 . ПМК 4690360 . ПМИД 26713256 .
- ^ Jump up to: а б с Поропат, Сан-Франциско; Мэннион, Вашингтон; Апчерч, П.; Хокнулл, ЮАР; Кир, BP (2016). «Новые австралийские зауроподы проливают свет на палеобиогеографию динозавров мелового периода» . Научные отчеты . 6 (34467): 34467. Бибкод : 2016NatSR...634467P . дои : 10.1038/srep34467 . ПМК 5072287 . ПМИД 27763598 .
- ^ Мольнар, Р.Э.; Виффен, Дж.; Хейс, Б. (1998). «Вероятная кость теропода из поздней юры Новой Зеландии» (PDF) . Новозеландский журнал геологии и геофизики . 41 (2): 145–148. дои : 10.1080/00288306.1998.9514798 .
- ^ Jump up to: а б с Уоррен, А.; Кул, Л.; Клиланд, М.; Викерс-Рич, П. (1991). «Раннемеловой лабиринтодонт» . Алчеринга . 15 (3–4): 327–332. дои : 10.1080/03115519108619027 .
- ^ Jump up to: а б с Кир, BP (2006). «Морские рептилии из нижнего мела Южной Австралии: элементы высокоширотного холодноводного комплекса» . Палеонтология . 49 (4): 837–856. дои : 10.1111/j.1475-4983.2006.00569.x .
- ^ Мартин, JE; Фернандес, М. (2007). «Синонимия позднемелового рода мозазавров (Squamata) Lakumasaurus из Антарктиды с Taniwhasaurus из Новой Зеландии и ее влияние на сходство фауны в провинции Уэдделлиан». Геологический журнал . 42 (2): 203–211. дои : 10.1002/gj.1066 . S2CID 128429649 .
- ^ Виффен, Дж. (1989). «Новые мозазавры (Reptilia; семейство Mosasauridae) из верхнего мела Северного острова, Новая Зеландия» . Новозеландский журнал геологии и геофизики . 33 (1): 67–85. дои : 10.1080/00288306.1990.10427574 .
- ^ Круикшанк, ARI; Фордайс, RW (2003). «Новая морская рептилия (Sauropterygia) из Новой Зеландии: дополнительные доказательства позднемеловой южной радиации криптоклидидных плезиозавров» . Палеонтология . 45 (3): 557–575. дои : 10.1111/1475-4983.00249 . S2CID 129320404 .
- ^ Jump up to: а б Бромман, Т.; Смит, ET; Белл, PR (2017). «Изолированные зубы Анхангерии (Pterosauria: Pterodactyloidea) из нижнего мела Лайтнинг-Ридж, Новый Южный Уэльс, Австралия» . ПерДж . 3 (5): е3256. дои : 10.7717/peerj.3256 . ПМК 5419211 . ПМИД 28480142 .
- ^ Jump up to: а б Келлер, AWA; Рич, TH; Коста, Франция; Викерс-Рич, П.; Кир, BP; Уолтерс, М. (2009). «Новые изолированные кости птеродактилоидов из формации Альбийский Тулебук (западный Квинсленд, Австралия) с комментариями к фауне австралийских птерозавров» . Алчеринга . 34 (3): 219–230. дои : 10.1080/03115511003656552 . S2CID 129915551 .
- ^ Jump up to: а б Кир, BP; Дьякон, Г.Л.; Сиверсон, М. (2009). «Останки позднемелового птерозавра из Molecap Greensand в Западной Австралии». Алчеринга . 34 (3): 273–279. дои : 10.1080/03115511003661651 . S2CID 129729142 .
- ^ Рич, TH; Викерс-Рич, П.; Фланнери, ТФ; Кир, BP (2009). «Австралийский мультитуберкулез и его палеобиогеографические последствия» . Acta Palaeontologica Polonica . 54 (1): 1–6. дои : 10.4202/app.2009.0101 .
- ^ Арчер, М.; Фланнери, ТФ; Ричи, А.; Мольнар, Р.Э. (1985). «Первое мезозойское млекопитающее из Австралии - монотрема раннего мела». Природа . 318 (6044): 363–366. Бибкод : 1985Natur.318..363A . дои : 10.1038/318363a0 . S2CID 4342084 .
- ^ Конгломерат на Fossilworks.org Монро
- ^ Stream на Fossilworks.org Swale
- ^ Залив Уэйхере, остров на Fossilworks.org Питт
Дальнейшее чтение
[ редактировать ]- Риффенбург, Б. (2007). Энциклопедия Антарктики . Том. 1. Тейлор и Фрэнсис. ISBN 978-0-415-97024-2 .
- Дутра, TL; Баттен, диджей (2000). «Верхнемеловые флоры острова Кинг-Джордж, Западная Антарктида, и их палеоэкологические и фитогеографические последствия». Меловые исследования . 21 (2–3): 181–209. дои : 10.1006/cres.2000.0221 .
- Филкорн, Х.Ф. (1994). «Ископаемые склерактиновые кораллы из бассейна Джеймса Росс, Антарктида». Серия антарктических исследований . 65 : 1–96.
- Харрис, AC; Рейн, Джи (2002). «Склерит позднемеловой бабочки (Insecta: Lepidoptera) из ущелья Ракайя, Кентербери, Новая Зеландия» . Журнал Королевского общества Новой Зеландии . 32 (3): 457–462. дои : 10.1080/03014223.2002.9517704 .
- Пул, И.; Меннега, AMW; Кантрилл, диджей (2003). «Вальдивские экосистемы в позднем меловом и раннем третичном периоде Антарктиды: дополнительные свидетельства миртовой и эвкрифиевой ископаемой древесины». Обзор палеоботаники и палинологии . 124 (1–2): 9–27. дои : 10.1016/S0034-6667(02)00244-0 . hdl : 1874/31608 . S2CID 129281012 .
- Рич, THV (2007). Полярные динозавры Австралии . Серия Музея природы Виктории. Музей Виктории. ISBN 978-0-9758370-2-3 .
- Спеден, Айова (1973). «Распределение, стратиграфия и стратиграфические взаимоотношения меловых отложений, западная часть полуострова Раукумара, Новая Зеландия» . Новозеландский журнал геологии и геофизики . 16 (2): 243–268. дои : 10.1080/00288306.1973.10431456 .
- Стилвелл, доктор медицинских наук; Витакка, Джей Джей; Мэйс, К. (2016). «Южнополярные парниковые насекомые (Arthropoda: Insecta: Coleoptera) из среднемеловой формации Тупуанги, острова Чатем, восточная Зеландия». Алчеринга . 40 (4): 502–508. дои : 10.1080/03115518.2016.1144385 . S2CID 130271909 .