Восточная Гондвана

Южный полярный регион мелового периода состоял в континенте Восточной Гондвана -современный день Австралии, Зеландии и Антарктиды-продукт распада Гондваны в меловой периоде . Южный регион в течение этого времени был намного теплее, чем сегодня, от 4 до 8 ° C (39–46 ° F) в последнем меловом маастрихте на юго -восточной Австралии. Это помешало постоянным ледяным щитам развивать и поощрять полярные леса , в которых в значительной степени доминировали хвойные деревья , циады и папоротники , и полагались на умеренный климат и сильные осадки. Основные геологические геологические образования , которые регистрируют эту область, являются: Санта-Марта и Sobral Formations острова Сеймур у Антарктического полуострова ; Остров Снежного холма , Лопес де Бертодано и Основные озера на острове Джеймс Росс также у Антарктического полуострова; и формации Eumeralla и Wonthaggi в Австралии.
В южном полярном регионе размещались много эндемичных видов, в том числе несколько форм реликторов , которые вымерли в другом месте меловым. Из сборки динозавров наиболее разнообразными были маленькие гипсилофодонты , подобные динозаврам. Южный полярный регион также был домом для последней амфибии из лабиринтодонта , Куласучу . Выделение Антарктиды вызвало особую экосистему морской жизни, называемую провинцией Веделлианской провинции . [ 1 ]
Пейзаж
[ редактировать ]Геология
[ редактировать ]
Бассейн Ладади в южном Антарктическом полуострове - Пальмера -земля - содержит вулканическую породу , подтверждая большое магматическое событие в среднем мелах, называемом событием Палмер Лэнд. Эта область дала множество как макро- , так и микро- установки, так и окаменелостей Mollusk, представляющих раннего мела , а также в середине до позднего юрского периода . [ 3 ] [ 4 ] Площадь, которая сейчас является Антарктическим полуостровами по всему мезозойскому, субдуктировала прото -тихоокеанскую плиту , вызывая вулканическую активность. [ 5 ]
ископаемого Важной формированием является формация Санта-Марта на острове Джеймс Росс у Антарктического полуострова , одной из наиболее значимых формирования южного полушария , представляющих высокую наземную флору и фауну поздней меловой полярной области. Остров Сеймур в этом отношении похож на область обнаружения для позвоночных и беспозвоночных существ, таких как плезиооры - позднего мелового, простирающегося в эоценом . [ 4 ] Другими меловыми ископаемыми образованием в Антарктиде являются формация острова Снежного холма , формация Лопес де Бертодано и Формирование Скрытого Озера , также на острове Джеймса Росс. [ 6 ] [ 7 ]
Cove Формирование Eumeralla Cove и формация Wonthaggi в Виктории, Австралия, содержат ряд ископаемых динозавров и треков от раннего мелового. представляет Формирование Тахоры собой сборку рептилий меловой Новой Зеландии . [ 7 ]
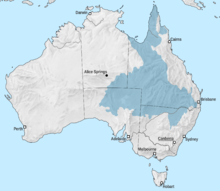
Море Эроманга было внутренним морем по всему тому, что было бы Австралией, которая была образована в раннемеловом. Море достигло бассейна Эроманга с севера через Карпентарный бассейн . Южный конец моря включала лагуны и реки, а также к востоку в заливе Сурат . Когда Индия ушла от Австралии в раннем меловом месте, бассейн Перт также заполнен морской водой. Область, которая будет коралловым морем, была рифтовой долиной . Когда Австралия и Антарктида развалились по всему меловая часть, между ними образовалось море. [ 8 ]
Растительность
[ редактировать ]Бриценица углекислого характеризуется теплыми глобальными температурами, вызванными высокими количествами газа и, возможно, метановыми парниковыми газами в атмосфере. Это вызвало отсутствие постоянного покрытия льда в полярных регионах, хотя уровень углекислого газа упал с 115 до 66 миллионов лет назад (MYA), возможно, позволив некоторое постоянное ледяное покрытие. Вполне возможно, что на протяжении всего мелового, разработанных нескольких небольших ледяных простоя. [ 9 ] Повышенная тектоническая активность , вызывая большее геотермальное тепло , могло предотвратить оледенение и повышение глобальной температуры. Температура могла быть до 15 ° C (27 ° F) теплее, чем в современном эпохе. [ 10 ] [ 11 ] [ 12 ]
Юрский период
[ редактировать ]Ландшафт полярного региона Средней Юрского периода был реконструирован из остатков лесов, обнаруженных в Новой Зеландии, которые были от 70 до 80 ° С во время юры. Растительность была в основном состоит из хвойных, циад и других спортивных полей , а также папоротников; На лесном дне выросли ликоподы , бриофиты , грибы и водоросли. У него был умеренный климат с большим количеством осадков. [ 13 ] [ 14 ]
Ранний меловой
[ редактировать ]В раннем мелах , Восточная Гондвана (Австралия, Антарктида и Зеландия ) начали отделяться от Южной Америки, а Индия и Мадагаскар также начали отделяться примерно в то же время. [ 9 ] Тропическая зона , возможно, расширилась во время меловых, до 32 ° ю.ш. , что позволило бы круглый год рост деревьев в Антарктике в полярных лесах . Обнаружение нескольких зрелых вечнозеленого и лиственных деревьев указывают на тепловую температуру с умеренными сезонами, в которых отсутствует широко распространенное замораживание, по крайней мере, между широты 70 и 85 ° с . [ 15 ] [ 16 ] Тем не менее, также возможно, что жизнь растений может быть представителем только теплых летних месяцев. [ 17 ] Многое из того, что известно о жизни растений Восточной Гондваны во время меловой, состоит из пыльцевых остатков и сжатия листьев с северного Антарктического полуострова . [ 18 ] В зависимости от широты, полярные зимы длились от шести недель до четырех с половиной месяцев. [ 19 ]

Тем не менее, скалы от ранней меловой формирования Вонтхаджи в юго -восточной Австралии свидетельствуют о сезонно замороженной земле. [ 21 ] Его географическое расположение в раннем меловом фоне примерно на 78 ° С указывает на эту область от одного до трех месяцев темноты в зим, и эта область является репрезентативной поймой в ледниковый питание . [ 22 ] Свидетельство раннего мела оледенения и холодного климата было обнаружено в отложениях в бассейне Эроманга в современной Центральной Австралии или от 60 до 80 ° С в раннем меловом периоде. Этот бассейн был большим внутренним морем в меловом виде. Тем не менее, аналогичные формирования могли быть созданы простой потоком мусора, и поэтому возможно, что оледенение никогда не было там. [ 23 ] [ 24 ] Общее покрытие полярного льда во время мезозоя, возможно, составляло треть размера, как это в наше время, хотя холодные снимки температуры подзыскания, возможно, произошли на протяжении ранних меловых. [ 25 ]
Поздний меловой
[ редактировать ]Навес в современного Александр который был около 75 ° , вокруг острова полярных лесах С. [ 4 ] [ 26 ] [ 16 ] Считается, что эти деревья оставались бездействующими полярными зимами до лета под полуночным солнцем . [ 27 ]
Свидетельство о цветущих растениях, датируемых примерно 80 млн. Лет в позднемеловом, предполагает существование умеренных лесов , в том числе в современной Австралии, Новой Зеландии и Южной Южной Америке. [ 20 ] Некоторые цветочные останки были обнаружены около 60 ° с , и в такой низкой широте возможно, что эта область была подвержена полярной зимы и сезонной погоде, хотя цветы предполагают годовой температурный диапазон 8–15 ° C (46–59 ° Е) и дождливый климат. [ 16 ] [ 18 ]
Остатки пыльцы из юго -восточной Австралии идентичны живым видам растений Австралии: хвойными деревьями, цветущими растениями, которые населяют районы с большим количеством осадков и средиземноморским климатом , а также склерофильные кустарники , что указывает на уникальный ландшафт тропических лесов и открытых кустарников. [ 28 ] Указано размером кольца деревьев на ископаемых деревьях, антарктические полярные леса имели тенденцию к охлаждению на протяжении всего маастрихтского возраста от 72 до 66 млн лет, от средней годовой температуры 7 ° C (45 ° F) до более сезонно экстремальных 4– 8 ° C (39–46 ° F). [ 29 ] Эти растения, скорее всего, пережили событие склеогенового вымирания мелового палеогена , которое убило большую часть жизни 66 млн. Лет на вулканическом Антарктическом полуострове ; Окаменелости растений, датируемые 60 млн. Лет в палеоцене с острова Сеймур у этого полуострова, признаны предками умеренных растений, населяющих современную Австралию и Южную Америку. [ 27 ]
The Late Cretaceous (Campanian) Zamek and Half Three Point Formations of King George Island were located at 60°S and display a rich assemblage of fossil flora, such as Podocarpus; Araucaria; the leptosporangiate ferns Cladophlebis and Clavifera; and a variety of Magnoliopsida flowering plants, Dicotylophyllum, Myrciophyllum santacruzensis, Nothofagus, Sterculiaephyllum australis, Monimiophyllum, and so forth.[30][31][32] The Sobral Formation of Seymour Island spanning the Cretaceous–Paleogene boundary at a paleolatitude of 63°S provided a new genus of fossil flower in the family Cunoniaceae, Eucryphiaceoxylon eucryphioides.[33]
Ecology
[edit]Much as in Australia today, East Gondwana played host to many endemic animals, which included many relict species of families that had gone extinct in the rest of the Cretaceous world. It is possible the polar regions of the Late Cretaceous had been inhabited by groups of plants and animals whose ancestry can be traced back to the Ordovician.[16][34] The gradual isolation of Antarctica in the Late Cretaceous created a distinct group of aquatic creatures called the Weddellian Province.[35]
Dinosaurs
[edit]Birds
[edit]
The remains of the ancestor of modern birds, the Neornithes, are uncommon in the Mesozoic, with a large radiation occurring in the Neogene of Antarctica. However, the discovery of the Late Cretaceous Vegavis, a goose-like bird, on Vega Island indicates that the major modern bird groups were already common in the Cretaceous. A femur belonging to an unidentified seriema-like bird was also discovered on Vega Island. Bird footprints were preserved in Dinosaur Cove, and, being larger than most Cretaceous bird species, indicate an abundance of larger enantiornithe or ornithurine birds during the Early Cretaceous.[36]
Two diving birds, possible primitive divers, were discovered in Late Cretaceous Chile and Antarctica: Neogaeornis and Polarornis. Polarornis may have been capable of both diving and flight.[36] The earliest penguins, Crossvallia and Waimanu, are known from 61–62 Ma in the Paleocene, however molecular data suggests penguins first evolved in the Late Cretaceous. Given that these penguins were dated so close to the Cretaceous–Paleogene extinction event, the group either evolved before the event or very rapidly afterwards.[36][37]
Non-avian
[edit]
Dinosaur fossils are rare from the South Polar region, and major fossil-bearing locations are the James Ross Island group; Beardmore glacier in Antarctica; Roma, Queensland; Mangahouanga stream in New Zealand; and Dinosaur Cove in Victoria, Australia.[39] The dinosaur remains of this region, such as those found in Victoria, consist only fragmentary pieces, making identification controversial. For example, disputed identifications of an allosaurid which may represent an abelisaurid, the ceratopsian Serendipaceratops which could be an ankylosaur, and the difficult-to-classify theropod Timimus have consequently been made.[38]
The supercontinent Pangaea of the Jurassic allowed major dinosaur clades to achieve a global distribution before breaking up, and several closely related cognates existed between South Polar forms and forms found elsewhere despite separation by the Tethys Ocean. However, dinosaur groups that achieved pan-Gondwanan distribution over the course of the Cretaceous would have had to have used the land bridge connecting Australia to South America via Antarctica in the South Polar region.[34] The South Polar iguanodontian Muttaburrasaurus is most closely related to European rhabdodontids, which were the dominant group in Europe during the Late Cretaceous. The Cretaceous South Polar Kunbarrasaurus is identified as being part of a unique lineage of Gondwanan ankylosaurs, which suggests homogeneity in the Gondwanan fauna.[40][41] The Dromaeosauridae are known from Antarctica, and represent a relict population from a previously world-wide distribution.[42] Despite these apparent cross-continental migrations, it is unlikely that South Polar dinosaurs migrated out of the polar forests during the winter, as they were either too massive–such as ankylosaurs–or too small–such as troodontids–to travel long distances, and a large sea between East Gondwana and other continents impeded any such migrations in the Late Cretaceous.[43][44] It is possible, to cope with the winter conditions, some dinosaurs hibernated, such as the theropod Timimus.[45]

The most common and diverse group found so far are the hypsilophodont-like dinosaurs, making up half of the dinosaur taxa found in southeastern Australia, which is unseen in more tropical regions, perhaps indicating some kind of advantage over other dinosaurs in the poles.[16] Being small with grinding dentition, they likely fed on low-lying vegetation such as lycopods and podocarp seed pods.[46] The hypsilophodont-like Leaellynasaura had large eye sockets, larger than more tropical hypsilophodont-like dinosaurs, and may have had acute night vision, suggesting that Leaellynasaura, and perhaps other hypsilophodont-like dinosaurs, lived in the polar areas for year-round or most of the year, including polar winters. Bonegrowth was continuous throughout its life, indicating it did not hibernate, being possible by perhaps being endothermic or poikilothermic,[39][47] or by digging burrows.[48] However, it is possible that the large eyes are merely due to ontogenesis, that is, relatively large eye-sockets may have been a feature only seen in juveniles or perhaps was a birth-defect, since there is only one specimen known.[43][19]
Though remains are scant and, consequently, taxonomic descriptions can be dubious,[49] the Victorian theropod remains have been assigned to seven different clades: Ceratosauria, Megaraptora, Tyrannosauroidea, and Maniraptora. However, tyrannosauroids are not known from other Gondwanan continents, and are more known from northern Laurasia.[50] Unlike the other Gondwanan continents whose apex predators where abelisaurids and carcharodontosaurids, the discovery of Australovenator, Rapator and an unnamed species in Australia suggests the megaraptorans were the top predators of East Gondwana. The tail vertebrae of an unknown theropod, dubbed "Joan Wiffen's theropod", were discovered in the Late Jurassic/Early Cretaceous rocks of New Zealand.[51]

Four titanosaurs–Australotitan, Savannasaurus, Diamantinasaurus, and Wintonotitan–discovered in the Winton Formation make up the sauropod assemblage of Cretaceous Australia, though, these creatures probably avoided the polar regions as their remains are completely absent in Southeast Australia which was within the South Polar region in the Cretaceous. However, it is likely that at least the titanosaurs migrated to Australia from South America, which would have required them to pass through Antarctica, since titanosaurs evolved in the Cretaceous after the break-up of Pangaea. It is possible the Bonarelli Event in the Middle Cretaceous may have made Antarctica warmer and therefore more hospitable to sauropods.[50] These dinosaurs probably fed on the fleshy seeds of podocarp and yew trees, as well as the commonplace forked ferns of the time.[46]
Paleocene
[edit]After an asteroid impact, the ensuing impact winter is thought to have killed off the dinosaurs along with much of Mesozoic life in the Cretaceous–Paleogene extinction event. However, the lack of an abrupt extinction horizon in Antarctic or Australian sediments for plant and bivalve fossils during this time period indicates a less powerful impact in the South Polar region.[16] Given that the dinosaurs and other fauna of the polar regions of the Cretaceous were well adapted for living in long periods of dark and cold weather, it has been postulated that this community might have survived the event.[19]
Rivers and lakes
[edit]The last temnospondyls–a group of giant amphibians which mainly died out after the Triassic–inhabited the South Polar region into the Early Cretaceous. Koolasuchus, perhaps the last of the temnospondyls, is thought to have survived in regions where it was too cold for their competitors, the neosuchians–a group of reptiles containing modern crocodilians–which are inactive in water below 10 °C (50 °F), to survive.[52] Though neosuchians are known from Cretaceous Australia, it is thought that they stayed away from the polar region, arriving to Australia over-seas rather than over-land.[34]

It is likely the temnospondyls inhabited the freshwater systems of polar Australia until the Bonarelli Event in the Middle Cretaceous around 100 mya increased temperatures and allowed neosuchians to inhabit Antarctica. These neosuchians, at an adult size of no more than 2.5 metres (8.2 ft) in length, likely led to the extinction of the temnospondyls in tandem with more-developed ray-finned fish which perhaps targeted their larvae. The migration of neosuchians into the region suggests that average winter temperatures were greater than 5.5 °C (41.9 °F), with an average annual temperature of more than 14.2 °C (57.6 °F). However, polar neosuchians are only known from an almost complete skeleton of Isisfordia, and other neosuchian remains are of undetermined species.[52]
Plesiosaurs inhabited freshwater river and estuary systems, given the locations of their remains, probably colonizing Australia in the Early to Middle Jurassic. Their remains, mainly teeth, have been documented from southeastern Australia deriving from the Late Cretaceous, though they were never described as the remains are too sparse to do so. The teeth share some affinity with pliosaurs, notably the rhomaleosaurids and Leptocleidus, which died out in the Early Cretaceous, indicating the polar freshwater systems may have been a refuge for the pliosaurs of the Cretaceous. In contrast to modern marine reptiles, these South Polar plesiosaurs probably had a better tolerance of colder waters.[53][24]
Oceans
[edit]
Early to Middle Cretaceous marine reptile remains of South Australia include five families of plesiosaurs–Cryptoclididae, Elasmosauridae, Polycotylidae, Rhomaleosauridae, and Pliosauridae–and the ichthyosaur family Ophthalmosauridae. The discovery of several juvenile plesiosaur remains suggest they used the nutrient-rich waters of the coast as sheltered calving grounds, the cold deterring predators such as sharks. Most of the plesiosaurs discovered had a cosmopolitan distribution, however endemic forms existed there such as Opallionectes and a possible new species of cryptoclidid. A dubious species of elasmosaurid Woolungasaurus, was named in 1928, one of the earliest description of an Australian marine reptile. Several mollusc, gastropod, ammonite, bony fish, chimaerid, and squid-like belemnite remains have been recovered as well.[53] The coastal area may have experienced winter freezing, and these reptiles, in response, may have migrated north during the winter, had a more active metabolism than tropical reptiles, have hibernated in freshwater areas much like the modern day American alligator (Alligator mississippiensis), or have been endothermic similar to modern day leatherback sea turtles (Dermochelys coriacea). The lower number of plesiosaurs and higher number of ichthyosaurs and sea turtles in more northerly areas of Australia indicates a preference for colder areas in plesiosaurs.[53]

Several Late Cretaceous oceanic plesiosaurs and mosasaurs have been discovered in New Zealand and Antarctica, with some, such as Mauisaurus, being endemic, while others, such as Prognathodon, having a cosmopolitan distribution.[54][55] Elasmosaurs and pliosaurs are known from one to three species from this area. The discovery of three cryptoclidids in the Southern Hemisphere–Morturneria from Antarctica, Aristonectes from South America, and Kaiwhekea from New Zealand–indicates a diversification of the family in the Late Cretaceous of this region and perhaps an increasing productivity of the early Southern Ocean.[56]
Pterosaurs
[edit]
Two clades of pterosaurs are represented in Early Cretaceous Australia, Pteranodontoidea and Ctenochasmatoidea, remains mainly deriving from the Toolebuc Formation and areas of Queensland and New South Wales. It is thought that at least six pterosaur taxa existed in Cretaceous Australia, however, given the fragmentary nature of the remains, many fossils recovered originate from undetermined pterosaur. Fossils were found in shallow-water environments and lagoons, indicating a diet of fish and other aquatic life. The ctenochasmatids were the only archaeopterodactyloids to survive into the Cretaceous. The only pterosaur tooth remains discovered in Australia deriving from the Early Cretaceous belong to Mythunga and a possible Late Cretaceous anhanguerid. Mythunga is estimated to have had a 4.5-meter (15 ft) wingspan, much larger than any other archaeopterodactyloid discovered, though it is possible the pterosaur is more related to the Anhangueridae or Ornithocheiridae.[57][58][59] However, pterosaur remains existing in what were the non-polar regions of Australia, given their ability to migrate by air, perhaps did not need to cross a land bridge through the polar regions to arrive there, meaning they did not ever inhabit the South Polar region.[16][34]
Of the Late Cretaceous pterosaurs, only the remains belonging to the family Azhdarchidae–found in the Carnarvon and Perth basins in Western Australia–were assigned to a taxon. A possible representative of Ornithocheiridae was found in Late Cretaceous Western Australia, though the family was previously thought to have gone extinct in the Early Cretaceous.[57][58][59]
Mammals
[edit]
Seven mammals have been discovered from Early Cretaceous Australia: an undescribed ornithorhynchid, Kryoryctes, Kollikodon, Ausktribosphenos, Bishops, Steropodon, and Corriebaatar; all of which were endemic to Australia during this time. It is likely mammals crossed the Antarctic land bridge between Australia and South America in the Early Cretaceous, and likely the ancestors of the endemic mammals of Australia arrived during the Jurassic across the supercontinent Pangaea.[60][61]
Invertebrates
[edit]Several fossils of insects and crustaceans are known from South Polar Cretaceous sediments of New Zealand. The Late Cretaceous Mangaotanean Monro Conglomerate was situated at 68°S and provided fossils of Helastia sp.,[62] and the crab Hemioon novozelandicum was found in the Swale Siltstone, located at 76°S during the late Albian.[63] Several specimens of insects were also found in the Tupuangi Formation of the Chatham Islands at a latitude of 79°S during the Cenomanian to Turonian.[64]
See also
[edit]- Dinosaur Cove
- Dinosaur Dreaming
- Geology of Antarctica, including paleontology
- List of Australian and Antarctic dinosaurs
- Gondwana Rainforests
- Polar forests of the Cretaceous
- "Spirits of the Ice Forest", episode 5 of Walking with Dinosaurs
References
[edit]- ^ Otero, Rodrigo A.; Soto-Acuña, Sergio; S, Christian Salazar; Oyarzún, José Luis (2015-03-27). "New elasmosaurids (Sauropterygia, Plesiosauria) from the Late Cretaceous of the Magallanes Basin, Chilean Patagonia: Evidence of a faunal turnover during the Maastrichtian along the Weddellian Biogeographic Province". Andean Geology. 42 (2): 237–267. doi:10.5027/andgeoV42n2-a05. ISSN 0718-7106.
- ^ "Dinosaur Cove, Australia". This Dynamic Earth. USGS. Retrieved 14 March 2015.
- ^ Vaughan, A. P. M.; Eagles, G.; Flowerdew, M. J. (2012). "Evidence for a two-phase Palmer Land event from crosscutting structural relationships and emplacement timing of the Lassiter Coast Intrusive Suite, Antarctic Peninsula: Implications for mid-Cretaceous Southern Ocean plate configuration" (PDF). Tectonics. 3 (1): TC1010. Bibcode:2012Tecto..31.1010V. doi:10.1029/2011TC003006. S2CID 129316703.
- ^ Jump up to: a b c Riffenburgh 2007, p. 72.
- ^ Riffenburgh 2007, p. 551.
- ^ Pirrie, D.; Crame, J. A.; Lomas, S. A.; Riding, J. B. (1997). "Late Cretaceous stratigraphy of the Admiralty Sound region, James Ross Basin, Antarctica". Cretaceous Research. 18 (1): 109–137. doi:10.1006/cres.1996.0052.
- ^ Jump up to: a b Weishampel, David B. (2004). "Dinosaur distribution (Early Cretaceous, Australasia)". In Weishampel, D. B.; Dodson, P.; Osmólska, H. (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 573–574. ISBN 978-0-520-24209-8.
- ^ Dettmann, M. E.; Molnar, R. E.; Douglas, J. G. (1992). "Australian cretaceous terrestrial faunas and floras: biostratigraphic and biogeographic implications". Cretaceous Research. 13 (3): 207–262. doi:10.1016/0195-6671(92)90001-7.
- ^ Jump up to: a b Hay, W. W. (2008). "Evolving ideas about the Cretaceous climate and ocean circulation". Cretaceous Research. 29 (5): 725–753. doi:10.1016/j.cretres.2008.05.025.
- ^ Ladant, J.; Donnadieu, Y. (2016). "Palaeogeographic regulation of glacial events during the Cretaceous supergreenhouse". Nature Communications. 7: 12771. Bibcode:2016NatCo...712771L. doi:10.1038/ncomms12771. PMC 5036002. PMID 27650167.
- ^ Beerling, D. J.; Osborne, C. P. (2002). "Physiological Ecology of Mesozoic Polar Forests in a High CO2 Environment". Annals of Botany. 89 (3): 329–339. doi:10.1093/aob/mcf045. PMC 4233824. PMID 12096745.
- ^ Peralta-Medina, E.; Falcon-Lang, H. J. (2012). "Cretaceous forest composition and productivity inferred from a global fossil wood database". Geology. 40 (3): 219–222. Bibcode:2012Geo....40..219P. doi:10.1130/G32733.1.
- ^ Thorn, V. (2001). "Vegetation communities of a high palaeolatitude Middle Jurassic forest in New Zealand". Palaeogeography, Palaeoclimatology, Palaeoecology. 168 (3–4): 273–289. Bibcode:2001PPP...168..273T. doi:10.1016/S0031-0182(01)00203-6.
- ^ Pole, M. (1999). "Structure of a near-polar latitude forest from the New Zealand Jurassic". Palaeogeography, Palaeoclimatology, Palaeoecology. 147 (1–2): 121–139. Bibcode:1999PPP...147..121P. doi:10.1016/S0031-0182(98)00151-5.
- ^ Douglas, J. G.; Williams, G. E. (1982). "Southern polar forests: The Early Cretaceous floras of Victoria and their palaeoclimatic significance". Palaeogeography, Palaeoclimatology, Palaeoecology. 39 (3–4): 171–185. Bibcode:1982PPP....39..171D. doi:10.1016/0031-0182(82)90021-9.
- ^ Jump up to: a b c d e f g Crame, J. A. (1992). "Late Cretaceous palaeoenvironments and biotas: an Antarctic perspective". Antarctic Science. 4 (4): 371–382. Bibcode:1992AntSc...4..371C. doi:10.1017/S0954102092000555. S2CID 140637028.
- ^ Pires, E. F.; Guerra-Sommer, M. (2011). "Growth ring analysis of fossil coniferous woods from Early Cretaceous of Araripe Basin (Brazil)" (PDF). Anais da Academia Brasileira de Ciências. 83 (2): 409–423. doi:10.1590/S0001-37652011005000005. PMID 21625801.
- ^ Jump up to: a b Poole, I.; Gottwald, H.; Francis, J. E. (2000). "Illicioxylon, an element of Gondwanan polar forests? Late Cretaceous and Early Tertiary woods of Antarctica". Annals of Botany. 86 (2): 421–432. doi:10.1006/anbo.2000.1208.
- ^ Jump up to: a b c Vickers-Rich, P.; Rich, T. H. (1993). "Australia's Polar Dinosaurs". Scientific American. 269 (1): 50–55. Bibcode:1993SciAm.269a..50V. doi:10.1038/scientificamerican0793-50. JSTOR 24941547.
- ^ Jump up to: a b Gottwald, H. (2001). "Monimiaceae sensu lato, an element of Gondwanan polar forests: Evidence from the late Cretaceous-early tertiary wood flora of Antarctica". Australian Systematic Botany. 14 (2): 207–230. doi:10.1071/SB00022.
- ^ Constantine, A.; Chinsamy-Turan, A.; Rich, P. V.; Rich, T. (1998). "Periglacial environments and polar dinosaurs". South African Journal of Science. 94 (3): 137–141.
- ^ Close, R. A.; Vicker-Rich, P.; Trusler, P.; Chiappe, L. M.; O'Connor, Jingmai; Rich, T. H.; Kool, L.; Komarower, P. (2008). "Earliest Gondwanan bird from the Cretaceous of southeastern Australia". Journal of Vertebrate Paleontology. 29 (2): 616–619. doi:10.1671/039.029.0214. S2CID 130306474.
- ^ Alley, N. F.; Frakes, L. A. (2003). "First known Cretaceous glaciation: Livingston Tillite Member of the Cadna-owie Formation, South Australia". Australian Journal of Earth Sciences. 50 (2): 139–144. Bibcode:2003AuJES..50..139A. doi:10.1046/j.1440-0952.2003.00984.x. S2CID 128739024.
- ^ Jump up to: a b Kear, B. P. (2004). "Plesiosaur remains from Cretaceous high-latitude non-marine deposits in Southeastern Australia". Journal of Vertebrate Paleontology. 26 (1): 196–199. doi:10.1671/0272-4634(2006)26[196:PRFCHN]2.0.CO;2. S2CID 130794388.
- ^ Price, G. D. (1999). "The evidence and implications of polar ice during the Mesozoic" (PDF). Earth-Science Reviews. 48 (3): 183–210. Bibcode:1999ESRv...48..183P. doi:10.1016/S0012-8252(99)00048-3.
- ^ Falcon-Lang, H. J.; Cantrill, D. J. (2001). "Leaf phenology of some mid-Cretaceous polar forests, Alexander Island, Antarctica". Geological Magazine. 138 (1): 39–52. Bibcode:2001GeoM..138...39F. doi:10.1017/S0016756801004927. S2CID 129461955.
- ^ Jump up to: a b Riffenburgh 2007, p. 413.
- ^ Деттманн, я; Джарзен Д.М. (1991). «Свидетельство пыльцы для поздней меловой дифференциации протеас в южных полярных лесах». Канадский журнал ботаники . 69 (4): 901–906. doi : 10.1139/b91-116 .
- ^ Фрэнсис, JE; Пул, И. (2002). «Меловой и ранний третичный климат Антарктиды: доказательства ископаемого дерева» (PDF) . Палеогеография, палеоклиматология, палеоэкология . 182 (1–2): 47–64. Bibcode : 2002ppp ... 182 ... 47f . doi : 10.1016/s0031-0182 (01) 00452-7 .
- ^ Хилл Сборка в FossilWorks .org Castle
- ^ Скуа .org Ассамблея Бэй
- ^ Половина трех очков сборки на FossilWorks .org
- ^ Sobral Formation at Fossilworks .org
- ^ Jump up to: а беременный в дюймовый Molnar, Re (1989). «Земные тетраподы в меловой Антарктиде». В Крейне, JA (ред.). Происхождение и эволюция антарктической биоты . Тол. 47. Специальные публикации геологического общества. С. 131–140. Bibcode : 1989gslsp..47..131m . Citeseerx 10.1.1.1010.6989 . doi : 10.1144/gsl.sp.1989.047.01.11 . S2CID 140579588 .
{{cite book}}:|journal=игнорируется ( помощь ) - ^ Riffenburgh 2007 , p. 146
- ^ Jump up to: а беременный в дюймовый Тамбусси, C.; Hospitalleche, CA (2007). PDF во время PDF . Revista Associacón Geológca 62 : 604–617.
- ^ Хакетт, SJ; Кимбалл, RT; Reddy, S.; Боуи, РКК; Браун, Эль; Браун, MJ (2008). «Филогеномное исследование птиц показывает их эволюционную историю». Наука . 320 (5884): 1763–1768. Bibcode : 2008Sci ... 320.1763H . doi : 10.1126/science.1157704 . PMID 18583609 . S2CID 6472805 .
- ^ Jump up to: а беременный Бенсон, RBJ; Рич, Т.Г.; Викерс-Рич, П.; Холл, М. (2012). «Фауна Theropod из южной Австралии указывает на высокое полярное разнообразие и провинция динозавров, основанный на климате» . Plos один . 7 (5): E37122. BIBCODE : 2012PLOSO ... 737122B . doi : 10.1371/journal.pone.0037122 . PMC 3353904 . PMID 22615916 .
- ^ Jump up to: а беременный Рич, Т.Г.; Викерс-Рич, П.; Gangloff, RA (2002). «Полярные динозавры» (PDF) . Наука . 295 (5557): 979–980. doi : 10.1126/science.1068920 . PMID 11834803 . S2CID 28065814 .
- ^ Сото-Акунья, Серхио; Варгас, Александр О.; Калуза, Джонатан; Леппе, Марсело А.; Botelho, Joao F.; Palma-Liberona, Хосе; Саймон-Гутсштейн, Каролина; Фернандес, Рой А.; Ортис, Эктор; Милла, Вероника; Аравена, Барбара (декабрь 2021 г.). «Причудливое хвостовое оружие в переходном анкилозавре из Субарктического Чили» . Природа . 600 (7888): 259–263. Bibcode : 2021natur.600..259s . Doi : 10.1038/s41586-021-04147-1 . ISSN 1476-4687 . PMID 34853468 . S2CID 244799975 .
- ^ Frauenfelder, Timothy G.; Белл, Фил Р.; Броум, Том; Бевитт, Джозеф Дж.; Бикнелл, Рассел, округ Колумбия; Кир, Бенджамин П.; WROE, Стивен; Кампионе, Николас Э. (2022-03-28). «Новые анокилозаврские черепные остатки из нижнего мелового (верхнего альбианского) образования Toolebuc of Queensland, Австралия» . Границы в науке Земли . 10 doi : 10.3389/feart.2022.803505 . ISSN 2296-6463 .
- ^ Дело, JA; Мартин, JE; Reguero, M. (2007). «Дромаозавр из маастрихтского острова Джеймс Росс и покойной меловой фауны Антарктики динозавров». Геологическая служба США и Национальные академии . Citeseerx 10.1.1.546.3890 .
- ^ Jump up to: а беременный Ричардс, ТВ; Ричардс; Викерс-Рич, П. (2000). "Где мы знаем; куда мы идем?" Полем Динозавры тьмы . Издательство Университета Индианы. ISBN 978-0-253-33773-3 .
- ^ Белл, PR; Snivy, E. (2007). «Полярные динозавры на параде: обзор миграции динозавров» . Алхринге: австралийский журнал палеонтологии . 32 (3): 271–284. doi : 10.1080/03115510802096101 .
- ^ Чинсми, а.; Rich, T.; Викерс-Рич, П. (1998). «Полярная гистология кости динозавров». Журнал палеонтологии позвоночных . 18 (2): 385–390. doi : 10.1080/02724634.1998.10011066 .
- ^ Jump up to: а беременный Деттманн, я; Молнар, Re; Дуглас, JG; Burger, D.; Fielding, C. (1992). «Австралийские меловые земные фауны и флора: биостратиграфические и биогеографические последствия». Мерашные исследования . 13 (3): 207–262. doi : 10.1016/0195-6671 (92) 90001-7 .
- ^ Вудворд, HN; Рич, Т.Г.; Чинсми, а.; Викерс-Рич, П. (2011). «Динамика роста полярных динозавров Австралии» . Plos один . 6 (8): E23339. BIBCODE : 2011PLOSO ... 6233339W . doi : 10.1371/journal.pone.00233339 . PMC 3149654 . PMID 21826250 .
- ^ Мартин, AJ (2009). «Динозавр зарывается в группе Otway (Albian) из Виктории, Австралии, и их отношение к меловой полярной среде». Мерашные исследования . 30 (5): 1223–1237. doi : 10.1016/j.cretres.2009.06.003 .
- ^ Белый, Массачусетс; Белл, PR; Кук, Ag; Poropat, SF; Elliott, DA (2015). «Целью австралийского завода Wintonensis (Theropoda, Megaraptoridae); последствия для зубного зуба Megaraptorid» . ПЕРЕЙ . 3 : E1512. doi : 10.7717/peerj.1512 . PMC 4690360 . PMID 26713256 .
- ^ Jump up to: а беременный в Poropat, SF; Mannion, PD; Upchurch, P.; Хокналл, СА; Кир, BP (2016). «Новые австралийские сауропод проливают свет на меловые палеобиогеографии динозавров» . Научные отчеты . 6 (34467): 34467. Bibcode : 2016natsr ... 634467p . doi : 10.1038/srep34467 . PMC 5072287 . PMID 27763598 .
- ^ Молнар, Re; Wiffen, J.; Хейс, Б. (1998). «Возможная тероподная кость из последней юры Новой Зеландии» (PDF) . Новозеландский журнал геологии и геофизики . 41 (2): 145–148. doi : 10.1080/00288306.1998.9514798 .
- ^ Jump up to: а беременный в Уоррен, А.; Kool, L.; Cleeland, M.; Викерс-Рич, П. (1991). «Ранний меловой лабиринтодонт» . Алхринге . 15 (3–4): 327–332. doi : 10.1080/03115519108619027 .
- ^ Jump up to: а беременный в Кир, BP (2006). «Морские рептилии из нижнего мела Южной Австралии: элементы высокоширотной сборки холодной воды» . Палеонтология . 49 (4): 837–856. doi : 10.1111/j.1475-4983.2006.00569.x .
- ^ Мартин, JE; Фернандес, М. (2007). «Синонимия рода позднего мела -мозазавра (Squamata) Lakumasaarus из Антарктиды с Taniwhasaurus из Новой Зеландии и его применением при сходстве с Фуналом с доказом Wedellian» Геологический журнал 42 (2): 203–2 Doi : 10.1002/ gj.1 S2CID 128429649
- ^ Wiffen, J. (1989). Полем А 33 (1): 67–8 doi : 10.1080/002888306.10290 .
- ^ Круикшанк, Ари; Fordyce, RW (2003). «Новая морская рептилия (Sauropterygia) из Новой Зеландии: дальнейшие доказательства позднего мела австралийского излучения криптокладидных плезиоооров» . Палеонтология . 45 (3): 557–575. doi : 10.1111/1475-4983.00249 . S2CID 129320404 .
- ^ Jump up to: а беременный Broughman, T.; Смит, et; Bell, PR (2017). «Изолированные зубы анхангвинии (Pterosauria: pterodactyloidea) из нижнего мела молнии, Новый Южный Уэльс, Австралия» . ПЕРЕЙ . 3 (5): E3256. doi : 10.7717/peerj.3256 . PMC 5419211 . PMID 28480142 .
- ^ Jump up to: а беременный Келлер, Ава; Рич, Т.Г.; Коста, FR; Викерс-Рич, П.; Kear, BP; Уолтерс, М. (2009). «Новые изолированные птеродактилоидные кости из формации Toolebuc Albian (Западный Квинсленд, Австралия) с комментариями к австралийской фауне птерозавра» . Алхринге . 34 (3): 219–230. doi : 10.1080/0311555110036565552 . S2CID 129915551 .
- ^ Jump up to: а беременный Kear, BP; Дьякон, GL; Сиверсон М. (2009). «Остатки позднего мелового птерозавра из молекапа зелени и западной Австралии». Алхринге . 34 (3): 273–279. doi : 10.1080/031155511003661651 . S2CID 129729142 .
- ^ Рич, Т.Г.; Викерс-Рич, П.; Flannery, TF; Кир, BP (2009). «Австралийские многотуркуляции и его палеобиогеографические последствия» . Acta Palaeontologica Polonica . 54 (1): 1–6. doi : 10.4202/app.2009.0101 .
- ^ Арчер, М.; Flannery, TF; Ritchie, A.; Molnar, Re (1985). «Первое мезозойское млекопитающее из Австралии - ранняя меловая монотрима». Природа . 318 (6044): 363–366. Bibcode : 1985natur.318..363a . doi : 10.1038/318363a0 . S2CID 4342084 .
- ^ Монро конгломерат на исчезающих заводах .org
- ^ Swale Stream на FossilWorks .org
- ^ Waihere Bay, Pitt Island at FossilWorks .org
Дальнейшее чтение
[ редактировать ]- Riffenburgh, B. (2007). Энциклопедия Антарктики . Тол. 1. Тейлор и Фрэнсис. ISBN 978-0-415-97024-2 .
- Dutra, TL; Баттен, DJ (2000). «Верхняя меловая флора острова короля Джорджа, Западная Антарктида, и их палеоэкологические и фитогеографические последствия». Мерашные исследования . 21 (2–3): 181–209. doi : 10.1006/cres.2000.0221 .
- Filkorn, HF (1994). «Окаменечные склерактинские кораллы из бассейна Джеймса Росса, Антарктида». Антарктическая серия исследований . 65 : 1–96.
- Харрис, AC; Рейн, Джи (2002). «Склерит от поздней меловой мотыльки (Insecta: Lepidoptera) из ущелья Ракайя, Кентербери, Новая Зеландия» . Журнал Королевского общества Новой Зеландии . 32 (3): 457–462. doi : 10.1080/03014223.2002.9517704 .
- Пул, я; Меннега, AMW; Cantrill, DJ (2003). «Валдивианские экосистемы в позднем меловом и раннем третичном в Антарктиде: дальнейшие доказательства из бессмысленного и евцифического ископаемого дерева». Обзор палеоботании и палинологии . 124 (1–2): 9–27. doi : 10.1016/s0034-6667 (02) 00244-0 . HDL : 1874/31608 . S2CID 129281012 .
- Rich, THV (2007). Полярные динозавры Австралии . Музей Victoria Nature Series. Музей Виктория. ISBN 978-0-9758370-2-3 .
- Speden, IA (1973). «Распространение, стратиграфия и стратиграфические отношения мела -отложений, полуостров Западного Раукумара, Новая Зеландия» . Новозеландский журнал геологии и геофизики . 16 (2): 243–268. doi : 10.1080/00288306.1973.10431456 .
- Стилуэлл, JD; Vitacca, JJ; Mays, C. (2016). «Южные полярные теплицы насекомые (членистоногие: инсекта: Coleoptera) из среднего мозговой формы Тупуанги, Чатем-острова, Восточная Зеландия». Алхринге . 40 (4): 502–508. doi : 10.1080/031155518.2016.1144385 . S2CID 130271909 .