Манизавр
| Манизавр Временной диапазон:
Поздняя юра , | |
|---|---|

| |
| С 3-го по 9-й спинные позвонки | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | † Зауроподоморфа |
| Клэйд : | † Зауропода |
| Суперсемейство: | † Диплодокоидея |
| Семья: | † Диплодокиды |
| Род: | † Манизавр Бонапарт и Матеус , 1999 г. |
| Разновидность: | † D. lourinhanensis
|
| Биномиальное имя | |
| † Динхейрозавр lourinhanensis Бонапарт и Матеус, 1999 г.
| |
Dinheirosaurus — род диплодоцидных динозавров - зауроподов , известный по окаменелостям, обнаруженным на территории современной Португалии . Он может представлять собой разновидность суперзавра . [ 1 ] Единственный вид — Dinheirosaurus lourinhanensis , впервые описанный Хосе Бонапартом и Октавио Матеусом в 1999 году по позвонкам и некоторым другим материалам из формации Лориньян . Хотя точный возраст формации неизвестен, ее можно датировать примерно ранним титоном поздней юры .
Известный материал включает два шейных позвонка , девять спинных позвонков , несколько ребер , фрагмент лобковой кости и множество гастролитов . Из материала диагностическими являются только позвонки, причем ребра и лобковая кость слишком фрагментарны или общие, чтобы отличить Dinheirosaurus . Этот материал впервые был описан как относящийся к роду Lourinhasaurus , но были замечены различия, и в 1999 году Бонапарт и Матеус переописали материал под новым биномом Dinheirosaurus lourinhanensis . Другой экземпляр, ML 418, предположительно динхейрозавр , как теперь известно, принадлежит другому португальскому диплодоциду. Это означает, что динхейрозавр жил рядом со многими тероподами , зауроподами, тиреофорами и орнитоподами , а также по крайней мере еще с одним диплодоцидом.
Динхейрозавр — диплодоцид, родственник апатозавра , диплодока , барозавра , суперзавра и торниерии . Среди них ближайшим родственником Динхейрозавра является Суперзавр .
Открытие и присвоение имени
[ редактировать ]ML 414 был впервые обнаружен в 1987 году г-ном Карлосом Анунсиасао. Он был связан с Музеем Лориньяна , и после раскопок, продолжавшихся с момента открытия до 1992 года, [ 2 ] Затем образец был перенесен в музей и занесен в каталог под номером 414. [ 3 ] Дантас и др. предварительно объявил ML 414, как только раскопки были завершены. Чтобы удалить окаменелости из окружающей породы, потребовались бульдозер и наклонный молот. Окаменелости находились на вершине прибрежного утеса и после извлечения были отправлены в Лориньян двумя блоками с помощью крана. За год до того, как его описали как новый таксон, Dantas et al. присвоил ML 414 Lourinhasaurus alenquerensis , ранее сгруппированному в группу Apatosaurus . Хосе Бонапарт и Октавио Матеус изучили материал Lourinhasaurus и пришли к выводу, что один экземпляр под названием ML 414 более тесно связан с диплодоцидами формации Моррисон и, таким образом, оправдывает новое биномиальное название. Этот новый вид был описан как Dinheirosaurus lourinhanensis , что в полном значении означает « ящерица Порту-Динхейру из Лориньяна ». [ 2 ] [ 3 ]
динхейрозавра Материал включал позвонки , ребра , части таза и гастролиты . Позвонки определенно происходили из шейного и спинного отделов и сочленялись. Две шейные кости сохранились не очень хорошо, хотя двенадцать спинных костей сочленены и находятся в хорошем состоянии. Другой материал позвонка включает семь фрагментарных центров и несколько нескрепленных нервных дуг. Сохранилось 12 спинных ребер, а также некоторые аппендикулярные элементы. [ 2 ] Дэвид Вейшампель и др. не признал весь материал принадлежащим Dinheirosaurus , и обнаружил в голотипе только 9 дорсальных костей, при этом неверно истолковав лобковую кость как фрагмент конечности. Они также неверно заявили, что он был найден в формации Камадас-де-Алькобаса . [ 4 ] Другая пара позвонков, под номером коллекции ML 418, первоначально была отнесена к Dinheirosaurus Бонапартом и Матеусом, но теперь считается отдельным новым безымянным родом диплодоцидов. [ 3 ] [ 5 ]
Описание
[ редактировать ]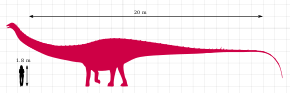
Динхейрозавр был диплодоцидом среднего размера, с удлиненной шеей и хвостом. [ 4 ] Основные особенности этого рода основаны на анатомии его позвонков: было обнаружено несколько позвонков по всему позвоночнику. [ 2 ] В общей сложности динхейрозавр имел приблизительную длину 20–25 метров (66–82 футов) и весил 8,8 метрических тонны (9,7 коротких тонны). [ 6 ] [ 7 ] [ 8 ]
Животное малоизвестно по непозвоночному материалу, в настоящее время состоящему только из частичных ребер и фрагмента таза. Одно из ребер прикреплено к шейным отделам и весьма фрагментарно. Он удлиненный, хотя это может быть особенностью искажения. Также Бонапартом и Матеусом не описан ряд грудных ребер. Два ребра находятся с левой стороны животного. Они имеют Т-образную форму в поперечном сечении и имеют плезиоморфные черты, хотя их незавершенное состояние делает их идентификацию сомнительной. Сохранилось множество правых ребер, включая стержни и головки. Они аналогичны левым ребрам, что также указывает на отсутствие пневматизации. [ 3 ] Другой аппендикулярный (непозвоночный) материал включает очень неполный и фрагментарный стержень лобковой кости и более ста гастролитов . Лобковая кость практически не имеет анатомических особенностей, а гастролиты не были подробно описаны Mannion et al. в 2012 году. [ 3 ]
Позвонки
[ редактировать ]Самый отличительный материал динхейрозавра происходит из позвонков, которые хорошо представлены и описаны. Из шейных позвонков сохранились только две из предполагаемых пятнадцати. По данным Бонапарта и Матеуса (1999), шейные шейки будут иметь номера 13 и 14. Судя по всему, 15-я шейка была утеряна во время раскопок и удаления голотипа и единственного экземпляра Dinheirosaurus . Согласно первоначальному описанию, тринадцатая шейная часть была препарирована только в латеро-вентральной части. Длина центра составляет 71 см (28 дюймов), а четырнадцатый шейный отдел в целом очень похож. 63 см (25 дюймов) — это общий размер 14-го центра шейного отдела, который хорошо сохранился, целый и вогнутый по нижнему краю. Нервный отросток , сжатый сверху по сравнению с шейными отделами диплодока , массивен и выступает вверх к заднему концу. [ 2 ]
Известен относительно полный ряд спинных позвонков, насчитывающий от одного до семи. Однако все спинные части деформированы вверх из-за степени сохранности. Бонапарт и Матеус (1999) отметили, что положение спинных костей не было точным и что на самом деле первый спинной плавник мог быть последним шейным или даже вторым спинным. Похожая нумерация была обнаружена у Diplodocus : первая и вторая спинные кости по анатомии аналогичны последней и предпоследней шейным отделам. Длина спинной кости варьируется от 58 см (23 дюйма) первой спинной кости до 25 см (9,8 дюйма) седьмой, восьмой и девятой спинной кости. Высота позвонков также весьма варьируется: наименьшая высота составляет от 51 см (20 дюймов) до 76 см (30 дюймов), увеличиваясь от первого спинного позвонка. [ 2 ]
Классификация
[ редактировать ]Динхейрозавр не очень хорошо известен, и, как следствие, его филогенетическое положение не определено. В 2012 г. во время переописания таксона Филипом Мэннионом и др. было обнаружено На обеих кладограммах , что он является сестринским видом Supersaurus vivianae и вместе образует наиболее базальные диплодоцины . Кладограмма 2012 года, опубликованная Mannion et al. и используя модифицированную матрицу Уитлока (2011) обнаружили, что Dinheirosaurus был более примитивным, чем Torneria , и более продвинутым, чем Apatosaurus . [ 3 ] Однако кладограмма 2014 года обнаружила, что их группа поддерживалась, но на самом деле была более примитивной, чем Apatosaurus , и, следовательно, находилась за пределами Diplodocinae . [ 9 ] В 2015 году Dinheirosaurus lourinhanensis был признан видом суперзавра в новой комбинации S. lourinhanensis ; их результаты показаны ниже. [ 1 ] Однако, чтобы подтвердить или опровергнуть эту гипотезу, необходимо подготовить несколько остатков голотипа. [ 10 ]
| Диплодокиды |
| ||||||||||||||||||||||||||||||||||||
Ранее Dinheirosaurus был отнесен к Diplodocidae, за исключением Apatosaurus , поскольку анатомические различия довольно велики. Бонапарт и Матеус обнаружили, что некоторые присутствующие особенности позволяют предположить, что Динхейрозавр был более производным, чем Диплодок , но плезиоморфные присутствующие особенности позволяют сделать вывод, что они разветвлялись отдельно, и Динхейрозавр не является потомком Диплодока . [ 2 ] Исследование 2004 года, проведенное Upchurch et al. обнаружили, что Dinheirosaurus был промежуточным диплодокоидом, наряду с Cetiosauriscus , Amphicoelias и Losillasaurus . [ 4 ]
Палеобиология
[ редактировать ]Будучи диплодоцидом, вполне вероятно, что динхейрозавр обладал хлыстовым хвостом. Если бы это было так, предполагалось, что его хвост можно было бы использовать как кнут со сверхзвуковой скоростью. [ 11 ] или, в последнее время, как орган осязания, позволяющий поддерживать связь с другими членами группы. [ 12 ] Будучи родственником апатозавра и диплодока , динхейрозавр, вероятно, обладал квадратной мордой. Это означает, что это, вероятно, был неизбирательный зауропод, питавшийся наземными водами. [ 13 ]
Пищеварение
[ редактировать ]Динхейрозавр — один из относительно немногих зауроподов, у которого гастролиты были обнаружены явно рядом с типовым экземпляром. В 2007 году эксперимент с использованием Dinheirosaurus , Diplodocus (= Seismosaurus ) и Cedarosaurus проверил, использовали ли зауроподы свои гастролиты в птичьего типа желудочной мельнице . При анализе учитывалось, что среди сотен найденных зауроподов гастролиты известны лишь у нескольких ассоциированных экземпляров. Авторы решили использовать трех зауроподов с наиболее ассоциированными гастролитами: Dinheirosaurus , Diplodocus и Cedarosaurus , из-за большого количества гастролитов, обнаруженных у птиц. В то время как у птиц обычно было обнаружено 1,05% гастролитов от массы тела, зауропод Diplodocus , у которого было наибольшее количество гастролитов, накопил только 0,03% массы тела. Это означает, что, поскольку у других зауроподов Dinheirosaurus и Cedarosaurus было меньше гастролитов по сравнению с массой тела, у зауроподов вряд ли развилась желудочная мельница птичьего типа, и вместо этого они могли использовать гастролиты для поглощения минералов. [ 14 ]
Палеоэкология
[ редактировать ]Динхейрозавр был одним из многих динозавров, живших в формации Лориньян в позднеюрском периоде. [ 2 ] Многие тероподы , зауроподы и особенно птицетазовые также происходят из формации Лориньян, которая содержит фауну, аналогичную североамериканской формации Моррисон. [ 4 ] Известны многие тероподы, включая безымянный род абелизаврид ; [ 15 ] аллозавриды europaeus Allosaurus ; цератозавр dentisculatus Ceratosaurus ; [ 16 ] целурозавры , jurassica Aviatyrannis [ 17 ] и ср. Ричардоэстезия ; промежуточный теропод; [ 15 ] и мегалозаврид Torvosaurus gurneyi . [ 17 ] Зауроподы менее распространены, есть только промежуточные диплодоциды , а также динхейрозавр ; [ 3 ] камаразаврид ; alenquerensis Lourinhasaurus туриазавр Zby atlanticus ; [ 18 ] и известен брахиозаврид Lusotitan . [ 19 ] Широко представлены птицетазовые животные, идентифицированные останки которых сохранились до Trimucrodon cuneatus ; [ 20 ] Алокодон куэней ; [ 21 ] стегозавры longicollum Dacentrurus Armatus , Miragaia , [ 22 ] и Stegosaurus ungulatus ; [ 23 ] анкилозаврида ; zbyszewski Dracopelta [ 24 ] орнитоподы , loureiroi Draconyx [ 25 ] Камптозавр сп., [ 18 ] Филлодон Хенкелли , [ 21 ] и ср. Дриозавр сп. [ 26 ]
Биогеография
[ редактировать ]Многие эузавроподы , в том числе динхейрозавры, были обнаружены в поздней юре Европы. Зауроподы происходят примерно из основания титона , судя по присутствию Anchispirocyclina lusitanica . Один зауропод, диплодоцид, в настоящее время основанный на безымянном образце, включающем позвонки и некоторые кости, явно отличается от Dinheirosaurus и Losillasaurus , что подтверждает наличие как минимум двух, а возможно, и большего количества диплодоцидов в поздней юре Испании и Португалии. Это уникальное разнообразие диплодокоидов во всей Европе, причем единственные другие роды, возможно, недиплодокоидные ( Cetiosauriscus ) или классифицируются как Rebbachisauridae . Это говорит о том, что биогеография примитивных зауроподов неполна, и, возможно, примитивные эузавроподы и диплодоциды сохранились в поздней юре, возможно, до берриаса . [ 5 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б ТСЧопп, Э.; МАТЕУС, О.; Бенсон, RBJ (2015). «Филогенетический анализ на уровне конкретного уровня и таксономический обзор Diplodocidae (Dinosauria, Sauropoda)» . ПерДж . 3 : е857. дои : 10.7717/peerj.857 . ПМЦ 4393826 . ПМИД 25870766 .
- ^ Jump up to: а б с д и ж г час Бонапарт Дж.; Мэтью, О. (1999). «Новый диплодоцид, Dinheirosaurus lourinhanensis gen. et sp. nov., из позднеюрских отложений Португалии» (PDF) . Журнал Аргентинского музея естественных наук . 5 (2): 13–29. ISSN 0524-9511 . Архивировано из оригинала (PDF) 28 сентября 2007 г.
- ^ Jump up to: а б с д и ж г Мэннион, Вашингтон; Апчерч, Пол; Мэтью, О.; Барнс, Р.Н.; Джонс, MEH (2012). «Новая информация об анатомии и систематическом положении Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) из поздней юры Португалии с обзором европейских диплодокоидов» (PDF) . Журнал систематической палеонтологии . 10 (3): 521–551. дои : 10.1080/14772019.2011.595432 . ISSN 1478-0941 . S2CID 56468989 .
- ^ Jump up to: а б с д Вейшампель, Д.Б.; Додсон, П.; Осмольска, Х. (2004). Динозаврия (Второе изд.). Беркли: Издательство Калифорнийского университета. стр. 259–549 . ISBN 978-0-520-94143-4 . OCLC 801843269 .
- ^ Jump up to: а б Ройо-Торрес, Р.; Кобос, А.; Аберастури, А.; Эспилес, Э.; Фиерро, И.; Гонсалес А.; Люке, Л.; Мампель, Л.; Алькала, Л. (2007). «Стоянки Риодевы (Теруэль, Испания), проливающие свет на филогению европейских зауроподов» (PDF) . Геогасета . 41 : 183–186. ISSN 0213-683X . Архивировано из оригинала (PDF) 24 сентября 2015 года . Проверено 24 августа 2014 г.
- ^ Матеус, О. (2010). «Палеонтологические коллекции музея Лориньян (Португалия)» (PDF) . В Брандао, JM; Каллапез, премьер-министр; МАТЕУС, О.; и др. (ред.). Геологические коллекции и музеи: миссия и управление . Университет Коимбры и Центр исследований и философии истории науки Коимбры. стр. 121–126.
- ^ Матеус, О. (2009). «ЗАВРОПОД ДИНОЗАВР TURIASAURUS RIODEVENSIS В ПОЗДНЕЙ ЮРЕ ПОРТУГАЛИИ» (PDF) . Журнал палеонтологии позвоночных.
- ^ Молина-Перес и Ларраменди (2020). Факты и цифры о динозаврах: зауроподы и другие зауроподоморфы . Нью-Джерси: Издательство Принстонского университета. п. 256. Бибкод : 2020dffs.book.....М .
- ^ Галлина, Пенсильвания; Апестегия, С.Н.; Халуза, А.; Канале, Джи (2014). «Выживший диплодоцид-зауропод из раннего мела Южной Америки» . ПЛОС ОДИН . 9 (5): е97128. Бибкод : 2014PLoSO...997128G . дои : 10.1371/journal.pone.0097128 . ПМК 4020797 . ПМИД 24828328 .
- ^ МОЧО, П.; Ройо-Торрес, Р.; Эскасо, Ф.; МАЛАФАЙЯ, Э.; Мигель Чавес, К.; НАРВЕЗ, И.; Перес-Гарсия, А.; ПИМЕНТЕЛЬ, Н.; Сильва, Британская Колумбия, и Ортега, Ф. (2017). «Находка позднеюрских зауроподов в бассейне Луситар (Португалия): географическое и литостратиграфическое распространение» (PDF) . Электронная палеонтология . 20 (2). дои : 10.26879/662 . S2CID 54817543 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мирвольд, Н.П.; Карри, Пи Джей (1997). «Сверхзвуковые зауроподы? Динамика хвоста у диплодоцидов» (PDF) . Палеобиология . 23 (4): 393–409. Бибкод : 1997Pbio...23..393M . дои : 10.1017/S0094837300019801 . JSTOR 2401127 . S2CID 83696153 .
- ^ Барон, Мэтью Г. (3 октября 2021 г.). «Тактильные хвосты: новая гипотеза функции удлиненных хвостов диплодоцидных зауроподов» . Историческая биология . 33 (10): 2057–2066. дои : 10.1080/08912963.2020.1769092 . ISSN 0891-2963 . S2CID 219762797 .
- ^ Уитлок, Дж. А. (2011). Фарке, А.А. (ред.). «Выводы о пищевом поведении диплодокоидов (Sauropoda: Dinosauria) на основании формы морды и анализа микроизноса» . ПЛОС ОДИН . 6 (4): e18304. Бибкод : 2011PLoSO...618304W . дои : 10.1371/journal.pone.0018304 . ПМК 3071828 . ПМИД 21494685 .
- ^ Уингз, О.; Сандер, член парламента (2007). «У динозавров-зауроподов нет желудочной мельницы: новые данные анализа массы и функции гастролита у страусов» . Труды Королевского общества Б. 274 (1610): 635–640. дои : 10.1098/rspb.2006.3763 . ПМК 2197205 . ПМИД 17254987 .
- ^ Jump up to: а б Хендрикс, К.; Матеус, О. (2014). «Зубы Abelisauridae (Dinosauria: Theropoda) из поздней юры Португалии и филогения на основе зубочелюстной системы как вклад в идентификацию изолированных зубов теропод» (PDF) . Зоотакса . 3759 (1): 1–74. дои : 10.11646/zootaxa.3759.1.1 . ПМИД 24869965 .
- ^ Матеус, О.; Вален, А.; Антунес, Монтана (2006). «Крупная фауна теропод формации Лориньян (Португалия) и ее сходство с формацией Моррисон с описанием нового вида аллозавра » (PDF) . Бюллетень Музея естественной истории и науки Нью-Мексико . 36 : 123–129.
- ^ Jump up to: а б Хендрикс, К.; Матеус, О.В. (2014). Эванс, Алистер Роберт (ред.). « Torvosaurus gurneyi n. sp., крупнейший наземный хищник из Европы, и предлагаемая терминология анатомии верхней челюсти у нонавианских теропод» . ПЛОС ОДИН . 9 (3): е88905. Бибкод : 2014PLoSO...988905H . дои : 10.1371/journal.pone.0088905 . ПМЦ 3943790 . ПМИД 24598585 .
- ^ Jump up to: а б Матеус, О; Мэннион, Вашингтон; Апчерч, П. (2014). « Zby atlanticus , новый туриазавровый зауропод (Dinosauria, Eusauropoda) из поздней юры Португалии» (PDF) . Журнал палеонтологии позвоночных . 34 (3): 618–634. Бибкод : 2014JVPal..34..618M . дои : 10.1080/02724634.2013.822875 . S2CID 59387149 .
- ^ Мэннион, Вашингтон; Апчерч, П.; Барнс, Р.; Матеус, О. (2013). «Остеология позднеюрского португальского динозавра-зауропода Lusotitan atalaiensis (Macronaria) и история эволюции базальных титанозавроформ» . Зоологический журнал Линнеевского общества . 168 : 98–206. дои : 10.1111/zoj.12029 .
- ^ Руис-Оменьака, Дж.И.; Канудо, Джи (2004). «Орнитоподовые динозавры нижнего мела Пиренейского полуострова». Гео-темы . 6 (5): 63–65.
- ^ Jump up to: а б Талборн, Р.А. (1973). «Зубы птицетазовых динозавров поздней юры Португалии с описанием гипилофодонтида ( Phylodon henkeli gen. et sp. nov.) из бурого угля Гимарота». Вклад в знание кимерриджской фауны буроугольного рудника Гимарота (Лейрия, Португалия) . Том. 22. Геологические службы Португалии, Память (новая серия). стр. 89–134.
- ^ Матеус, О.; Мейдмент, С.; Кристиансен, Н. (2009). «Новый стегозавр с длинной шеей, имитирующий зауроподов, и эволюция пластинчатых динозавров» (PDF) . Труды Лондонского королевского общества Б. 276 (1663): 1815–1821. дои : 10.1098/рспб.2008.1909 . ПМЦ 2674496 . ПМИД 19324778 .
- ^ Эскасо, Ф.; Ортега, Ф.; Дантас, П.; Малафайя, Э.; Пиментел, Нидерланды; Переда-Субервиола, X.; Санс, Дж.Л.; Куллберг, Дж. К.; Куллберг, MAC; Баррига, Ф. (2006). «Новые свидетельства существования общего динозавра в верхней юре прото-Северной Атлантики: стегозавр из Португалии» (PDF) . Естественные науки . 94 (5): 367–74. Бибкод : 2007NW.....94..367E . дои : 10.1007/s00114-006-0209-8 . ПМИД 17187254 . S2CID 10930309 .
- ^ Гальтон, премьер-министр (1980). «Частичный скелет Dracopelta zbyszewski n. gen. и n. sp., анкилозаврового динозавра из верхней юры Португалии». Геобиос . 13 (3): 451–457. Бибкод : 1980Geobi..13..451G . дои : 10.1016/s0016-6995(80)80081-7 .
- ^ МАТЕУС, О.; Антунес, Монтана (2001). « Draconyx loureiroi , новый камптозавриды (Dinosauria, Ornithopoda) из поздней юры Лориньяна, Португалия» (PDF) . Анналы палеонтологии . 87 (1): 61–73. Бибкод : 2001AnPal..87...61M . дои : 10.1016/S0753-3969(01)88003-4 .
- ^ Антунес, Монтана; Матеус, О. (2003). «Динозавры Португалии» (PDF) . Аккаунты Палевол . 2 (1): 77–95. Бибкод : 2003CRPal...2...77A . дои : 10.1016/S1631-0683(03)00003-4 .











