Синсюлун
| Синсюлун Временной диапазон:
| |
|---|---|

| |
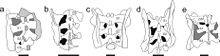
| Фотографии элементов скелета (ан) и реконструкция скелета (о) Синсюлуна. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | † Зауроподоморфа |
| Клэйд : | † Зауроподиформы |
| Род: | †Синсюлун Ван и др. , 2017 |
| Типовой вид | |
| † Ван и др. , 2017
| |
Синсюлун (что означает «дракон моста Синсю») — род двуногих из ранней юры Китая зауроподообразных . Он содержит единственный вид, X. chengi , описанный Wang et al. в 2017 году от трех экземпляров, двух взрослых особей и неполовозрелой особи, которые в совокупности составляют практически полный скелет. Взрослые особи этого рода имели длину 4–5 метров (13–16 футов) и высоту 1–1,5 метра (3 фута 3 дюйма – 4 фута 11 дюймов). Филогенетический анализ показывает, что Синсюлун наиболее тесно связан с современным ему Цзиншанозавром , хотя альтернативная позиция за пределами как Sauropodiformes, так и Massospondylidae также вероятна.
Несмотря на их близкое родство, Синсюлун заметно отличается от Цзиншанозавра - и от большинства базальных зауроподоморфов - наличием ряда черт, подобных зауроподам . К ним относятся крестец , содержащий четыре позвонка; лобок ; с исключительно длинной верхней частью и бедренная кость , первая и пятая плюсневые кости стопы, а также лопатка широкая и крепкая. Вероятно, это представляет собой адаптацию к поддержанию большого веса тела, в частности толстого кишечника. Однако, в отличие от зауроподов, Синсюлун по-прежнему был двуногим.
Описание
[ редактировать ]
Синсюлун среднего размера был зауроподиформом , с длиной взрослой особи 4–5 метров (13–16 футов) и высотой 1–1,5 метра (3 фута 3 дюйма – 4 фута 11 дюймов) в бедрах. Два более крупных экземпляра, LFGT-D0002 и LFGT-D0003, судя по полному сращению черепа, являются взрослыми особями; третий экземпляр, LFGT-D0001, на 14% меньше и, судя по неполному сращению позвонков, вероятно, представляет собой полувзрослую особь. [ 1 ]
Череп и челюсть
[ редактировать ]
В отличие от современного Люфенгозавра , [ 2 ] Синсюлун не имеет гребня на боковой стороне верхней челюсти . По всей длине верхней челюсти сохранилось одиннадцать зубных лунок. Дальше назад, перед глазницей, слезная железа имеет заметный выступ около вершины переднего конца, который также наблюдается у Lufengosaurus . [ 2 ] Адеопаппозавр , Массоспондил , [ 3 ] и Riojasaurus , но не Yunnanosaurus , Jingshanosaurus или последующие зауроподиформы. Контактная поверхность между скуловой костью и заглазничной костью довольно длинная, как у Lufengosaurus , но не у Yunnanosaurus . [ 4 ] Среди трех ветвей скуловой кости направленная назад ветвь образует угол 80° с выступающей вверх ветвью, которая похожа на Платеозавра и Текодонтозавра, но намного крупнее, чем у других зауроподоморфов . [ 1 ] [ 4 ] [ 5 ] [ 6 ]
У основания черепа квадратно-скуловая кость имеет две ветви: одну направленную вперед, а другую вверх; они примерно перпендикулярны друг другу, в отличие от Люфенгозавра (угол 45 °), Юннанозавра (угол 60 °) и Цзиншанозавра (угол 110 °). Над квадратноскуловой квадратом имеются два сочленяющихся мыщелка : подтреугольный, обращенный наружу, и более закругленный, обращенный внутрь; последний мыщелок расположен ближе к низу, как у Lufengosaurus и Yunnanosaurus , но не у Plateosaurus . В задней части черепа, между теменными и надзатылочными костями , имеется хорошо развитое посттеменное окно; сама надзатылочная часть наклонена вперед на нижнем конце, чтобы закруглить основание черепа. Базиптеригоидные отростки длинные, тонкие и выступают вниз и наружу, как у платеозавра , в отличие от люфенгозавра и джингшанозавра , образуя угол 80° друг с другом. [ 1 ]
По сравнению с Lufengosaurus , Yunnanosaurus и Jingshanosaurus , угловые и надугольные отверстия простираются намного дальше перед нижнечелюстным отверстием у Xingxiulong , который ближе к Adeopapposaurus и Plateosaurus . [ 7 ] Суставной сустав имеет выступающий внутрь пирамидальный отросток, являющийся продолжением челюстного сустава; на заднем конце он также имеет направленный вверх отросток в виде язычка, который также наблюдается у Coloradisaurus . [ 8 ] Цзиншанозавр и пока еще безымянный зауроподоморф. [ 1 ]
Позвонки
[ редактировать ]имеется десять шейных позвонков В шее Синсюлуна . Проатлан, атрофированный позвонок, расположенный впереди атланта , ограничен спереди верхними сторонами большого затылочного отверстия . Хотя о самом атласе сложно что-либо сказать из-за повреждений, следующая за ним ось имеет относительно короткий центр , слегка сжатый по бокам и снизу. В целом остальные шейные позвонки относительно короткие: их длина всего в 2,5–3 раза превышает их высоту; к задней части шеи они становятся все короче, как у Люфенгозавра . Другие базальные зауроподоморфы, [ 9 ] [ 10 ] включая Jingshanosaurus , шейные позвонки обычно в 3-4 раза длиннее их высоты. В нижней части центров 4-9 шейных позвонков имеется выступающий киль, как у Lufengosaurus , Jingshanosaurus и других базальных зауроподоморфов. В задней части шеи нервные отростки становятся похожими на стол. [ 1 ]
Четырнадцать спинных позвонков слабо амфицельные или вогнутые с обоих концов, что типично для базальных зауроподоморфов. Также характерно наличие лезвийных килей на нижней стороне первого-третьего спинных позвонков, которые больше нигде не встречаются; кроме того, спинные позвонки спереди имеют типичные низкие, короткие и несколько пластинчатые нервные отростки. Однако необычно то, что те же типы нервных отростков наблюдаются и в последних трех спинных позвонках, что иначе наблюдается только у базальных ящеротазовых, таких как Herrerasaurus и Eoraptor . В средних и задних спинных позвонках верхний задний угол нервных отростков выступает наружу, образуя вогнутый задний край, который наблюдается у некоторых других базальных зауроподоморфов, но не у Lufengosaurus , Jingshanosaurus , Yunnanosaurus и Riojasaurus . [ 1 ]

, что необычно для базальных зауроподоморфов Синсюлун имеет четыре крестцовых позвонка . Эта черта обычно наблюдается у более производных зауроподоморфов, таких как Melanorosaurus , Leonerasaurus , [ 11 ] Барапазавр , [ 12 ] и Шунозавр . Первый из этих крестцовых позвонков, спинно-крестцовый, не очень хорошо сращен со следующим крестцовым позвонком; он также не полностью сращен с соответствующим крестцовым ребром на поперечных отростках . Вместе дорсо-крестцовые и первая пара крестцовых ребер выдвигаются вперед и касаются подвздошной кости . Второй и третий крестцовые позвонки, два первичных крестца, примерно одинаковой длины и хорошо срослись друг с другом. Как и у других базальных зауроподоморфов, поперечные отростки третьего крестцового позвонка выступают назад и наружу. То же самое касается поперечного отростка и крестцовых ребер четвертого крестцового позвонка, которые сзади снова соприкасаются с подвздошной костью. Все нервные отростки пластинчатые, как и несколько последних спинных позвонков. [ 1 ]
Вполне вероятно, что Синсюлун имел более 35 сохранившихся хвостовых позвонков в хвосте. Эти позвонки в целом высокие и очень крепкие, с вогнутыми сторонами, как и у других базальных зауроподоморфов. Кроме того, все они кажутся амфицельными. Поперечные отростки первых нескольких хвостовых позвонков широкие, плоские, направлены вверх и наружу; поперечные отростки хвостовых позвонков дальше в хвосте более тонкие и горизонтальные. Что касается нервных отростков, то они высокие, тонкие и направлены несколько назад. [ 1 ]
Конечности
[ редактировать ]конца лопатки Синсюлуна Оба довольно расширены; ширина нижнего конца составляет 56% длины лопатки, а ширина верхнего конца составляет 49% длины лопатки. Различные другие базальные зауроподоморфы, в том числе Lufengosaurus и Jingshanosaurus , имеют более расширенный нижний конец, но их лопатки обычно более тонкие; и наоборот, антетонитрус [ 13 ] и Lessemsaurus имеют одинаково крепкие лопатки, но у этих двух таксонов верхний конец более расширен. Как и у Jingshanosaurus , максимальная ширина мощного стержня лопатки составляет 19-20% длины всей кости; Антетонитрус [ 13 ] и у лессемзавра стержень еще более крепкий, тогда как у большинства базальных зауроподоморфов стержень более узкий. [ 1 ]
Бугристость на внутренней поверхности верхнего конца плечевой кости развита довольно слабо у Синсюлуна , в отличие от большинства базальных зауроподоморфов (включая Lufengosaurus и Yunnanosaurus ). Как и у Юннанозавра и Цзиншанозавра , локтевая кость составляет около 61% длины плечевой кости; это соотношение составляет 68% у Lufengosaurus , что приводит к более длинной локтевой кости. Верхний конец локтевой кости довольно расширен, с выраженными переднемедиальным и переднелатеральным отростками; эти отростки вместе с неглубокой лучевой ямкой в совокупности образуют сочленение локтевой кости с лучевой костью . Последняя кость тонкая и составляет около 54% длины плечевой кости. Части руки, на которых было как минимум четыре пальца, известны, но недостаточно подробно описаны. [ 1 ]
В целом подвздошная кость похожа на другие базальные зауроподоморфы. Передняя часть подвздошной кости не превышает уровня лобковой ножки или ее сочленения с лобком . В отличие от других базальных зауроподоморфов, задний конец несколько квадратный, а не заостренный, а нижняя часть очень вогнутая, если смотреть сбоку (у других базальных зауроподоморфов она в основном прямая или даже выпуклая). Седалищная ножка, или часть, которая сочленяется с седалищной костью , имеет на нижнем конце небольшую выступающую пятку. Что касается самой лобковой кости, то ее верхний конец (известный как лобковая пластинка) относительно длинный, составляя 40% длины кости, а нижняя часть (известная как лобковый фартук), наоборот, относительно короткая. Это не похоже на других базальных зауроподоморфов, но похоже на некоторых базальных зауроподов. Наружная поверхность лобкового фартука несколько вогнута, а нижний конец расширен вперед и назад примерно на 16% длины всей кости. Наконец, расширенная запирательная пластинка седалищной кости имеет на своей стороне бороздку. [ 1 ]
На бедренной кости малый вертел простирается ниже уровня головки бедренной кости , в отличие от Юннанозавра и Цзиншанозавра . При взгляде спереди вертел расположен близко к средней линии кости, как и у других базальных зауроподоморфов, за исключением Antetonitrus и Melanorosaurus (у которых он ближе к внешнему краю). Точно так же четвертый вертел находится вблизи средней линии, в отличие от многих базальных зауроподоморфов. [ 8 ] включая Люфенгозавра . Из двух отростков нижней части большеберцовой кости задний тоньше и выступает больше наружу, чем вниз, чем передний; это отличается от большинства других зауроподоморфов, у которых они оба одинаково толстые, и либо они одинаково выступают наружу (как у Lufengosaurus и Jingshanosaurus ), либо передняя часть выступает дальше наружу (как у Yunnanosaurus ). [ 1 ]
Задняя часть таранной кости имеет выпуклость вблизи нижнего конца кости. Подобная выпуклость также наблюдается у Mussaurus . [ 6 ] хотя и лучше развит и расположен ближе к средней линии. Из плюсневых костей первая является самой широкой и прочной. Пятая плюсневая кость необычно расширена на верхнем конце, ширина которой составляет 85% длины всей кости. У других зауроподоморфов этот показатель обычно составляет 50-77%, [ 14 ] хотя Antetonitrus может иметь такую же расширенную плюсневую кость. Ниже плюсневых костей пять пальцев имеют фаланговую формулу 2-3-4-5-1. [ 1 ]
Открытие и присвоение имени
[ редактировать ]
LFGT-D0002, зрелый голотип Синсюлонга , состоит из частичного черепа с челюстями; с седьмого по девятый шейные позвонки , с восьмого по четырнадцатый спинные позвонки , весь крестец и 35 хвостовых позвонков; ребер и шевронов фрагменты ; левая подвздошная кость , а также части лобковой и седалищной костей ; обе бедренные и большеберцовые кости , части малоберцовой кости , левая лодыжка и обе ступни (почти целые). [ 1 ]
Существуют два упомянутых экземпляра. LFGT-D0003, также зрелый, состоит из частичного черепа с челюстями; с третьего по десятый шейные позвонки, а также все спинные позвонки и большая часть крестца; фрагменты ребер и шевронов; обе лопатки , плечевые кости , локтевые кости и лучевые кости , а также часть кисти; части как подвздошной кости (правая целая), так и лобковой кости (левая целая); левое бедро, части большеберцовой и малоберцовой костей (правые целые), а также правая лодыжка и части стоп. LFGT-D0001, который меньше по размеру и, вероятно, незрелый, состоит из оси , шейных позвонков с третьего по десятый, всех спинных позвонков и крестца, а также первых девятнадцатых хвостовых позвонков; фрагменты ребер и шевронов; правые лопатка и подвздошная кость, а также части правой лобковой и седалищной кости; части бедра и левой большеберцовой кости, а также левую лодыжку. [ 1 ]
Эти экземпляры были обнаружены и захоронены вместе недалеко от деревни Санкешу округа Луфэн , Юньнань , Китай , в 2013 году и хранятся в Бюро земель и ресурсов округа Луфэн. Породы, в которых сохранились эти образцы, состоящие из пурпурного алевритового аргиллита , принадлежат раннеюрской пачке Шаван формации Луфэн которой предварительно датируется геттангом . , возраст [ 1 ]
Синсюлун был описан в 2017 году Я-Минг Ван, Хай-Лу Ю и Тао Ван. Родовое название (буквально означающее «созвездие») относится к мосту Синсю (星宿), который был построен во времена династии Мин китайской . Между тем, видовое название «Чэньги» дано в честь профессора Чжэн-У Ченга, который внес большой вклад в биостратиграфию Китая, в том числе в районе Луфэн, и умер в 2015 году. [ 1 ]
Классификация
[ редактировать ]2017 года Филогенетический анализ , проведенный на основе набора данных McPhee et al. , опубликованный в 2015 году с описанием Пуланесауры , [ 15 ] обнаружили, что Синсюлун был базальным членом группы Sauropodiformes после того, как Blikanasaurus был удален из набора данных. Внутри этой группы он наиболее близок к современному Цзиншанозавру . Черты, общие для Синсюлуна и Цзиншанозавра, включают подвисочное окно , полностью расположенное за глазницей; ширина лопатки ; составляет не менее 20% ее длины лобковый передник, или нижняя часть лобка , имеющий вогнутую наружную поверхность; расширение нижней части фартука, составляющее не менее 15% длины всей кости; и угол между головкой бедренной кости и осью поперечного сечения диафиза бедренной кости составляет около 30°. Топология, восстановленная в результате этого анализа, воспроизведена ниже. [ 1 ]
Были протестированы различные альтернативные филогенетические размещения Синсюлонга ; из них наиболее правдоподобная альтернатива предполагает, что его поместят в качестве базального члена Massopoda , за пределами Massospondylidae или Sauropodiformes, что требует всего лишь двух дополнительных эволюционных шагов. Вероятно, это связано с изогнутым верхним краем заглазничной кости и наличием отростка позади челюстного сустава на суставной кости , которые являются признаками, которые по сравнению с Massospondylidae или Sauropodiformes можно рассматривать как относительно примитивные. Такой альтернативный сценарий предполагает значительную мозаичную эволюцию на этом эволюционном уровне. Размещения, которые требовали трех дополнительных эволюционных шагов, помещали Синсюлуна между Цзиншанозавром и Анчизавром или между Юннанозавром и Цзиншанозавром . [ 1 ]
Любопытно, что Синсюлун обладает многими характеристиками, которые обычно наблюдаются у зауроподов . из четырех позвонков К ним относятся крестец (который Мусзавр также конвергентно ); приобрел [ 6 ] [ 11 ] [ 16 ] длинная лобковая пластинка, или верхняя часть лобка, занимающая 40% длины кости (у большинства других базальных зауроподоморфов этот показатель составляет 25% , [ 8 ] 33% у большинства зауроподов, [ 17 ] и 45-50% у Camarasauromorpha. [ 18 ] ); и относительная прочность бедренной кости, [ 11 ] первая плюсневая кость , [ 19 ] верхний конец пятой плюсневой кости и лопатки. Эти уникальные характеристики, вероятно, являются продуктом конвергентной эволюции, а не общего происхождения. [ 1 ]
Палеобиология
[ редактировать ]Общая прочность скелета Синсюлуна , особенно бедра, бедренной кости и стопы, сходна с скелетом зауроподов и в совокупности позволяет предположить, что у него был относительно большой кишечник и общая высокая масса тела. Однако, в отличие от зауроподов, Синсюлун был двуногим ; у него отсутствуют приспособления зауроподов к четвероногому движению, включая относительно более длинные передние конечности, локтевую кость с выступающим отростком на передней стороне и бедренную кость , имеющую относительно прямой стержень. [ 19 ] Вместо этого его локтевая и бедренная кости в целом больше напоминают типичного базального зауроподоморфа . Большие и крепкие лопатки Xingxiulong Jingshanosaurus , , и Yunnanosaurus возможно, увеличили подвижность передних конечностей во время двуногого передвижения, но позже эта черта была принята как адаптация к четвероногому образу жизни у зауроподов. [ 1 ] [ 15 ]
Палеоэкология
[ редактировать ]Формация Луфэн содержит аргиллиты и алевролиты из озер, рек и прибрежных отложений. [ 20 ] Многие зауроподоморфы , помимо Синсюлонга , известны из формации Луфэн, в том числе Lufengosaurus huenei , L. magnus , Yunnanosaurus huangi , «Gyposaurus» sinensis , Jingshanosaurus xinwaensis , Chuxiongosaurus lufengensis , [ 1 ] Xixiposaurus suni , « Yizhousaurus sunae », [ 21 ] и Pachysurus несовершенный Также присутствуют тероподы Sinosaurus triassicus , Что такое Lukosaurus , [ 20 ] Шидаизавр джинаэ , [ 21 ] [ 22 ] и Eshanosaurus deguchiianus ; [ 23 ] птицетазовые Tatisaurus oehleri [ 20 ] и Bienosaurus lufengensis ; крокодиломорфы Microchampsa Dibothrosuchus elaphros , Platyognathus hsui , scutata , Dianosuchus changchiawaensis и Dianchungosaurus lufengensis ; индетерминантный архозавр Strigosuchus licinus ; сфенодонты и Clevosaurus petilus , C. wangi C. mcgilli ; тритилодонтидные Dianzhongia цинодонты Bienotherium yunnanense , B.minor , B.magnum , Lufengia deltcata , Yunnanodon brevirostre и longirostrata ; млекопитающие Sinoconodon rigneyi , Morganucodon oehleri , M. heikoupengensis и Kunminia minima ; проганохелиидные черепахи; и амфибия -лабиринтодонт . [ 20 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v Ван, Ю.-М.; Вы, Х.-Л.; Ван, Т. (2017). «Новый базальный зауроподиформный динозавр из нижней юры провинции Юньнань, Китай» . Научные отчеты . 73 : 41881. Бибкод : 2017NatSR...741881W . дои : 10.1038/srep41881 . ПМК 5312170 . ПМИД 28205592 .
- ^ Jump up to: а б Барретт, премьер-министр; Апчерч, П.; Ван, X.-L. (2005). «Краниальная остеология Lufengosaurus huenei Young (Dinosauria: Prosauropoda) из нижней юры Юньнани, Китайская Народная Республика». Журнал палеонтологии позвоночных . 25 (4): 806–822. doi : 10.1671/0272-4634(2005)025[0806:COOLHY]2.0.CO;2 . JSTOR 4524508 . S2CID 86257677 .
- ^ Сьюс, Х.-Д.; Рейс, РР; Хинич, С.; Раат, Массачусетс (2004). «На черепе Massospondylus carinatus Owen, 1854 (Dinosauria: Sauropodomorpha) из формаций Эллиот и Кларенс (нижняя юра) Южной Африки» . Анналы музея Карнеги . 73 (4): 239–257. дои : 10.5962/стр.316084 . S2CID 251542198 .
- ^ Jump up to: а б Барретт, премьер-министр; Апчерч, П.; Чжоу, X.-D.; Ван, X.-L. (2007). «Череп Yunnanosaurus huangi Young, 1942 (Dinosauria: Prosauropoda) из формации Нижний Луфэн (нижняя юра) в Юньнани, Китай» (PDF) . Зоологический журнал Линнеевского общества . 150 (2): 319–341. дои : 10.1111/j.1096-3642.2007.00290.x .
- ^ Лу, Дж.; Кобаяши, Ю.; Ли, Т.; Чжун, С. (2010). «Новый базальный динозавр-зауропод из бассейна Луфэн, провинция Юньнань, Юго-Западный Китай». Акта Геологика Синика . 84 (6): 1336–1342. дои : 10.1111/j.1755-6724.2010.00332.x . S2CID 130303928 .
- ^ Jump up to: а б с Отеро, А.; Пол, Д. (2013). «Посткраниальная анатомия и филогенетические взаимоотношения Mussaurus patagonicus (Dinosauria, Sauropodomorpha)». Журнал палеонтологии позвоночных . 33 (5): 1138–1168. дои : 10.1080/02724634.2013.769444 . hdl : 11336/21805 . S2CID 86110822 .
- ^ Прието-Маркес, А.; Норелл, М. (2011). «Переописание почти полного черепа платеозавра (Dinosauria, Sauropodomorpha) из позднего триаса Троссингена (Германия)» . Новитаты Американского музея (3727): 1–58. дои : 10.1206/3727.2 . hdl : 2246/6138 . S2CID 55272856 .
- ^ Jump up to: а б с Апалдетти, К.; Мартинес, Р.Н.; Пол, Д.; Саутер, Т. (2014). «Переописание черепа Coloradisaurus brevis (Dinosauria, Sauropodomorpha) из позднетриасовой формации Лос-Колорадос в бассейне Союза Исчигуаласто-Вилла, северо-запад Аргентины». Журнал палеонтологии позвоночных . 34 (5): 1113–1132. дои : 10.1080/02724634.2014.859147 . hdl : 11336/36518 . S2CID 86158311 .
- ^ Мартинес, Р.Н. (2009). « Adeopapposaurus mognai , gen. et sp. nov. (Dinosauria: Sauropodomorpha), с комментариями об адаптациях базальных Sauropodomorpha». Журнал палеонтологии позвоночных . 29 (1): 142–164. дои : 10.1671/039.029.0102 . S2CID 85074392 .
- ^ Апалдетти, К.; Мартинес, Р.Н.; Алькобер, ОА; Пол, Д. (2011). «Новый базальный зауроподоморф (Dinosauria: Saurischia) из формации Кебрада-дель-Барро (бассейн Марайес-Эль-Каррисаль), северо-запад Аргентины» . ПЛОС ОДИН . 6 (11): e26964. Бибкод : 2011PLoSO...626964A . дои : 10.1371/journal.pone.0026964 . ПМК 3212523 . ПМИД 22096511 .
- ^ Jump up to: а б с Пол, Д.; Гарридо, А.; Серда, Айова (2011). «Новый динозавр-зауроподоморф из ранней юры Патагонии, а также происхождение и эволюция крестца типа зауропод» . ПЛОС ОДИН . 6 (1): e14572. Бибкод : 2011PLoSO...614572P . дои : 10.1371/journal.pone.0014572 . ПМК 3027623 . ПМИД 21298087 .
- ^ Джайн, СЛ; Катти, Т.С.; Рой-Чоудхури, Т.; Чаттерджи, С. (1975). «Динозавр зауропод из нижнеюрской формации Кота в Индии». Труды Королевского общества B: Биологические науки . 188 (1091): 221–228. Бибкод : 1975РСПСБ.188..221J . дои : 10.1098/rspb.1975.0014 . S2CID 84957608 .
- ^ Jump up to: а б Макфи, BW; Йейтс, AM; Шуаньер, Ж.Н.; Абдала, Ф. (2014). «Полная анатомия и филогенетические взаимоотношения Antetonitrus ingenipes (Sauropodiformes, Dinosauria): значение для происхождения Sauropoda». Зоологический журнал Линнеевского общества . 171 (1): 151–205. дои : 10.1111/zoj.12127 . S2CID 82631097 .
- ^ Макфи, BW; Шуаньер, Ж.Н.; Йейтс, AM; Вильетти, Пенсильвания (2015). «Второй вид Eucnemesaurus Van Hoepen, 1920 (Dinosauria, Sauropodomorpha): новая информация о разнообразии и эволюции фауны зауроподоморфов нижней формации Эллиот в Южной Африке (последний триас)». Журнал палеонтологии позвоночных . 35 (5): e980504. дои : 10.1080/02724634.2015.980504 . S2CID 86805607 .
- ^ Jump up to: а б Макфи, BW; Боннан, МФ; Йейтс, AM; Невелинг, Дж.; Шуаньер, Ж.Н. (2015). «Новый базальный зауропод из доарской юры Южной Африки: свидетельства разделения ниш на границе зауроподоморф-зауропод?» . Научные отчеты . 5 : 13224. дои : 10.1038/srep13224 . ПМК 4541066 . ПМИД 26288028 .
- ^ Апчерч, П. (1998). «Филогенетические взаимоотношения динозавров-зауроподов» . Зоологический журнал Линнеевского общества . 124 (1): 43–103. дои : 10.1006/zjls.1997.0138 .
- ^ Уилсон, Дж.А.; Серено, ПК (1998). «Ранняя эволюция и филогения высшего уровня динозавров-зауроподов» (PDF) . Журнал палеонтологии позвоночных . 18 (sup002): 1–79. дои : 10.1080/02724634.1998.10011115 .
- ^ Сальгадо, Л.; Кориа, РА; Кальво, ДЖО (1997). «Эволюция зауроподов-титанозаврид. I: Филогенетический анализ, основанный на посткраниальных данных» . Амегиниана . 34 (1): 3–32. ISSN 0002-7014 .
- ^ Jump up to: а б Йейтс, AM; Боннан, МФ; Невелинг, Дж.; Чинсами, А.; Черная Борода, МГ (2009). «Новый переходный динозавр-зауроподоморф из ранней юры Южной Африки и эволюция питания и четвероногости зауроподов» . Труды Королевского общества B: Биологические науки . 277 (1682): 787–794. дои : 10.1098/rspb.2009.1440 . ПМЦ 2842739 . ПМИД 19906674 .
- ^ Jump up to: а б с д Луо, З.; У, Х.-С. (1997). «Маленькие четвероногие формации Нижний Луфэн, Юньнань, Китай» . Во Фрейзере, Северная Каролина; Сьюс, Х.-Д. (ред.). В тени динозавров: раннемезозойские четвероногие . Кембридж: Издательство Кембриджского университета. стр. 251–270. ISBN 9780521458993 .
- ^ Jump up to: а б Син, Л.; Ротшильд, Б.М.; Ран, Х.; Мияшита, Т.; Лица, WS; Секия, Т.; Чжан, Дж.; Ван, Т.; Донг, З. (2015). «Слияние позвонков у двух раннеюрских динозавров-зауроподоморфов из формации Луфэн в Юньнани, Китай» . Acta Palaeontologica Polonica . 60 (3): 643–649. дои : 10.4202/app.00001.2013 .
- ^ У, Х.-С.; Карри, Пи Джей; Донг, З.; Пан, С.; Ван, Т. (2009). «Новый динозавр-теропод из средней юры Луфэна, Юньнань, Китай». Акта Геологика Синика . 83 (1): 9–24. дои : 10.1111/j.1755-6724.2009.00002.x . S2CID 140695367 .
- ^ Сюй, Х.; Чжао, X.; Кларк, Дж. М. (2001). «Новый теризинозавр из нижнеюрской формации Нижний Луфэн в Юньнани, Китай». Журнал палеонтологии позвоночных . 21 (3): 477–483. doi : 10.1671/0272-4634(2001)021[0477:ANTFTL]2.0.CO;2 . JSTOR 20061976 . S2CID 131298010 .











