Клеточная культура

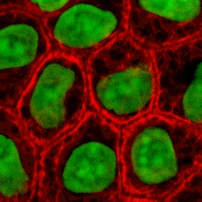
Культура клеток или культура тканей — это процесс, с помощью которого клетки выращиваются в контролируемых условиях, как правило, за пределами их естественной среды. После того как интересующие клетки были выделены из живой ткани , их впоследствии можно хранить в тщательно контролируемых условиях. Их необходимо хранить при температуре тела (37°C) в инкубаторе. [1] Эти условия различаются для каждого типа клеток, но обычно состоят из подходящего сосуда с субстратом или богатой средой , которая поставляет необходимые питательные вещества ( аминокислоты , углеводы , витамины , минералы ), факторы роста , гормоны и газы ( CO 2 , O 2 ), а также регулирует физико-химическую среду ( рН-буфер , осмотическое давление , температуру ). Большинству клеток требуется поверхность или искусственный субстрат для формирования прикрепившейся культуры в виде монослоя (толщиной в одну клетку), тогда как другие можно выращивать в свободном плавании в среде в виде суспензионной культуры . [2] Этому обычно способствуют использование жидкой, полутвердой или твердой питательной среды , такой как бульон или агар . Культура тканей обычно относится к культуре клеток и тканей животных, при этом более конкретный термин «культура тканей растений» для растений используется . Продолжительность жизни большинства клеток детерминирована генетически, но некоторые клетки, культивируемые в клетках, были «трансформированы» в иммортализованные клетки, которые будут воспроизводиться бесконечно, если будут обеспечены оптимальные условия.
На практике термин «культура клеток» теперь относится к культивированию клеток, полученных из многоклеточных эукариот , особенно клеток животных , в отличие от других типов культур, которые также выращивают клетки, таких как культура растительных тканей , культура грибов и микробиологическая культура . микробов ) . Историческое развитие и методы культивирования клеток тесно взаимосвязаны с культурой тканей и культурой органов . Вирусная культура также связана с клетками, являющимися хозяевами вирусов.
Лабораторный . метод поддержания линий живых клеток (популяция клеток, произошедших от одной клетки и содержащих один и тот же генетический состав), отделенных от исходного источника ткани, стал более надежным в середине 20 века [3] [4]
История [ править ]
Английский физиолог XIX века Сидней Рингер разработал солевые растворы, содержащие хлориды натрия, калия, кальция и магния, пригодные для поддержания биения изолированного сердца животного вне тела. [5] В 1885 году Вильгельм Ру удалил часть мозговой пластинки куриного эмбриона в течение нескольких дней , и подержал ее в теплом солевом растворе установив основной принцип культуры тканей. В 1907 году зоолог Росс Грэнвилл Харрисон продемонстрировал рост эмбриональных клеток лягушки, которые дали начало нервным клеткам в среде свернувшейся лимфы . В 1913 Э. Стейнхардт, К. Израэль и Р. А. Ламберт вырастили коровьей оспы вирус во фрагментах ткани роговицы морской свинки . [6] В 1996 году впервые была использована регенеративная ткань для замены небольшой длины уретры, что привело к пониманию того, что техника получения образцов ткани, выращивания их вне тела без каркаса и повторного применения может быть использована для только небольшие расстояния менее 1 см. [7] [8] [9] Росс Грэнвилл Харрисон , работавший в Медицинской школе Джонса Хопкинса , а затем в Йельском университете , опубликовал результаты своих экспериментов с 1907 по 1910 год, установивших методологию культуры тканей . [10]
Готлиб Хаберландт первым указал на возможности культуры изолированных тканей, культуры тканей растений . [11] Он предположил, что с помощью этого метода можно определить потенциальные возможности отдельных клеток с помощью культуры тканей, а также взаимное влияние тканей друг на друга. Со времени первоначальных утверждений Хаберландта были реализованы методы культивирования тканей и клеток, что привело к значительным открытиям в биологии и медицине. Свою первоначальную идею тотипотенциальности он представил в 1902 году, заявив, что «теоретически все растительные клетки способны дать начало полноценному растению». [12] [13] [14] Термин «культура ткани» был придуман американским патологом Монтроузом Томасом Берроузом . [15]
Методы культивирования клеток значительно продвинулись в 1940-х и 1950-х годах для поддержки исследований в области вирусологии . Выращивание вирусов в культурах клеток позволило получить очищенные вирусы для изготовления вакцин . Инъекционная вакцина против полиомиелита, разработанная Джонасом Солком, была одним из первых продуктов, массово производимых с использованием методов культивирования клеток. Эта вакцина стала возможной благодаря исследованиям клеточных культур Джона Франклина Эндерса , Томаса Хакла Веллера и Фредерика Чепмена Роббинса , которые были удостоены Нобелевской премии за открытие метода выращивания вируса в культурах клеток почек обезьян . Культура клеток способствовала разработке вакцин от многих заболеваний. [1]
Современное использование [ править ]

В современном использовании термин «культура ткани» обычно относится к росту клеток из ткани многоклеточного организма in vitro . Эти клетки могут представлять собой клетки, выделенные из организма-донора ( первичные клетки ) или иммортализованную клеточную линию . Клетки погружают в культуральную среду, которая содержит необходимые питательные вещества и источники энергии, необходимые для выживания клеток. [16] Таким образом, в более широком смысле «культура ткани» часто используется как синоним «культуры клеток». С другой стороны, строгое значение термина «культура ткани» относится к культивированию кусочков ткани, т.е. культуре эксплантатов .
Культура тканей — важный инструмент для изучения биологии клеток многоклеточных организмов. Он обеспечивает in vitro модель ткани в четко определенной среде, которой можно легко манипулировать и анализировать. В культуре тканей животных клетки можно выращивать в виде двумерных монослоев (традиционная культура) или внутри волокнистых каркасов или гелей для получения более натуралистичных трехмерных тканеподобных структур (3D-культура). Эрик Саймон в отчете о гранте NIH SBIR 1988 года показал, что электропрядение можно использовать для производства полимерных волокнистых каркасов нано- и субмикронного размера, специально предназначенных для использования в качестве клеточных и тканевых субстратов in vitro . Это раннее использование электроформованных волокнистых решеток для клеточной культуры и тканевой инженерии показало, что различные типы клеток будут прикрепляться к поликарбонатным волокнам и размножаться на них. Было отмечено, что в отличие от уплощенной морфологии, обычно наблюдаемой в 2D-культуре, клетки, выращенные на электропряденых волокнах, демонстрировали более округлую трехмерную морфологию, обычно наблюдаемую в тканях. живой . [17]
Культура тканей растений , в частности, связана с выращиванием целых растений из небольших кусочков растительной ткани, культивируемых в среде. [18]
культуры Концепции млекопитающих клеток
Выделение клеток [ править ]
Клетки можно выделить из тканей для культуры ex vivo несколькими способами. Клетки легко очищаются от крови; однако только белые клетки способны расти в культуре. Клетки можно выделить из твердых тканей путем расщепления внеклеточного матрикса с помощью таких ферментов , как коллагеназа , трипсин или проназа , перед перемешиванием ткани для высвобождения клеток в суспензию. [19] [20] Альтернативно, кусочки ткани можно поместить в питательную среду , а вырастающие клетки можно будет использовать для культивирования. Этот метод известен как культура эксплантатов .
Клетки, культивированные непосредственно у субъекта, известны как первичные клетки. За исключением некоторых, полученных из опухолей, большинство первичных культур клеток имеют ограниченную продолжительность жизни.
Установленная или иммортализованная клеточная линия приобрела способность бесконечно размножаться либо посредством случайной мутации, либо преднамеренной модификации, такой как экспрессия гена теломеразы искусственная .Многочисленные клеточные линии хорошо известны как представители определенных типов клеток .
Поддержание клеток в культуре [ править ]
Большинство изолированных первичных клеток подвергаются процессу старения и перестают делиться после определенного количества удвоений популяции, в целом сохраняя при этом свою жизнеспособность (описанную как предел Хейфлика ).

Помимо температуры и газовой смеси, наиболее часто меняющимся фактором в культуральных системах является среда для роста клеток . Рецепты питательных сред могут различаться по pH , концентрации глюкозы, факторам роста и наличию других питательных веществ. Факторы роста, используемые в качестве дополнения к среде, часто получают из сыворотки крови животных, например, фетальной бычьей сыворотки (FBS), бычьей телячьей сыворотки, лошадиной сыворотки и свиной сыворотки. Одним из осложнений использования этих ингредиентов, полученных из крови, является возможность загрязнения культуры вирусами или прионами , особенно в медицинских биотехнологических приложениях. Текущая практика заключается в том, чтобы свести к минимуму или исключить использование этих ингредиентов, где это возможно, и использовать лизат тромбоцитов человека (hPL). [21] Это позволяет избежать беспокойства о межвидовом загрязнении при использовании FBS с клетками человека. hPL стал безопасной и надежной альтернативой прямой замене FBS или другой сыворотки животных. Кроме того, химически определенные среды для устранения любых следов сыворотки крови (человеческой или животной) можно использовать , но это не всегда может быть достигнуто с использованием разных типов клеток. Альтернативные стратегии включают получение крови животных из стран с минимальным риском ГЭКРС / ТГЭ , таких как США, Австралия и Новая Зеландия. [22] и использование очищенных питательных концентратов, полученных из сыворотки, вместо сыворотки цельного животного для клеточной культуры. [23]
Плотность посева (количество клеток на объем культуральной среды) играет решающую роль для некоторых типов клеток. Например, более низкая плотность посева заставляет гранулезные клетки проявлять выработку эстрогена, тогда как более высокая плотность посева заставляет их выглядеть как прогестерон продуцирующие клетки текалютеина, . [24]
Клетки можно выращивать либо в суспензии , либо в прикрепившихся культурах . [25] Некоторые клетки естественным образом живут во взвешенном состоянии, не прикрепляясь к поверхности, например клетки, существующие в кровотоке. Существуют также клеточные линии, которые были модифицированы для выживания в суспензионных культурах, поэтому их можно выращивать до более высокой плотности, чем позволяют условия адгезии. Адгезивным клеткам необходима поверхность, такая как пластик или микроноситель для тканевой культуры , которая может быть покрыта компонентами внеклеточного матрикса (такими как коллаген и ламинин) для увеличения адгезионных свойств и предоставления других сигналов, необходимых для роста и дифференцировки. Большинство клеток, происходящих из твердых тканей, являются адгезивными. Другим типом адгезивной культуры является органотипическая культура , которая предполагает выращивание клеток в трехмерной (3-D) среде, в отличие от двумерных культуральных чашек. Эта система 3D-культуры биохимически и физиологически более похожа на ткань in vivo , но ее технически сложно поддерживать из-за многих факторов (например, диффузии). [26]
Базальные среды клеточных для культур
Существуют различные виды сред для культивирования клеток, которые обычно используются в науках о жизни, включая следующие:
сред для Компоненты культивирования клеток
| Компонент | Функция |
|---|---|
| Источник углерода ( глюкоза / глютамин ) | Источник энергии |
| Аминокислота | Строительные блоки белка |
| Витамины | Способствовать выживанию и росту клеток |
| Сбалансированный солевой раствор | Изотоническая кофакторов смесь ионов для поддержания оптимального осмотического давления внутри клеток и обеспечения необходимых ионов металлов, которые действуют в качестве для ферментативных реакций, клеточной адгезии и т. д. |
| Краситель феноловый красный | Индикатор pH . Цвет фенолового красного меняется от оранжево-красного при pH 7–7,4 до желтого при кислом (более низком) pH и фиолетового при основном (более высоком) pH. |
| Бикарбонатный/HEPES буфер | Он используется для поддержания сбалансированного pH в среде. |
роста Типичные условия
| Параметр | |
|---|---|
| Температура | 37 °С |
| СО2 | 5% |
| Относительная влажность | 95% |
клеточных Перекрестное загрязнение линий
Перекрестное загрязнение клеточных линий может стать проблемой для ученых, работающих с культивируемыми клетками. [27] Исследования показывают, что в 15–20% случаев клетки, использованные в экспериментах, были ошибочно идентифицированы или загрязнены другой клеточной линией. [28] [29] [30] Проблемы с перекрестным загрязнением клеточных линий были обнаружены даже в линиях из панели NCI-60 , которые регулярно используются для исследований по скринингу лекарств. [31] [32] Основные репозитории клеточных линий, в том числе Американская коллекция типовых культур (ATCC), Европейская коллекция клеточных культур (ECACC) и Немецкая коллекция микроорганизмов и клеточных культур (DSMZ), получили от исследователей материалы о линиях клеток, которые были ими ошибочно идентифицированы. [31] [33] Такое заражение создает проблему для качества исследований, проводимых с использованием линий клеточных культур, и основные репозитории в настоящее время проверяют подлинность всех представленных линий клеток. [34] ATCC использует с короткими тандемными повторами (STR) дактилоскопию ДНК для аутентификации своих клеточных линий. [35]
Чтобы решить эту проблему перекрестного загрязнения клеточных линий, исследователям рекомендуется аутентифицировать свои клеточные линии на ранних стадиях, чтобы установить идентичность клеточной линии. Аутентификацию следует повторять перед замораживанием запасов клеточных линий, каждые два месяца во время активного культивирования и перед любой публикацией данных исследований, полученных с использованием клеточных линий. Для идентификации клеточных линий используются многие методы, включая изоферментный анализ, типирование антигена лимфоцитов человека (HLA), хромосомный анализ, кариотипирование, морфологический и STR-анализ . [35]
Одним из значительных перекрестных загрязнителей клеточных линий является иммортализованная клеточная линия HeLa . Заражение HeLa было впервые отмечено в начале 1960-х годов в нечеловеческой культуре в США. Внутривидовое заражение было обнаружено в девятнадцати клеточных линиях в семидесятые годы. В 1974 году было обнаружено, что пять клеточных линий человека из Советского Союза относятся к HeLa. Последующее исследование, анализировавшее более 50 клеточных линий, показало, что половина из них имела маркеры HeLa, но примесная HeLa гибридизировалась с исходными клеточными линиями. о заражении клеток HeLa из капель воздуха Сообщалось . HeLa даже неосознанно вводился людям Джонасом Солком во время испытаний вакцины в 1978 году. [36]
Прочие технические проблемы [ править ]
Поскольку клетки обычно продолжают делиться в культуре, они обычно растут, заполняя доступную площадь или объем. Это может вызвать несколько проблем:
- Истощение питательных веществ в питательной среде
- Изменения pH питательной среды
- Накопление апоптотических / некротических (мертвых) клеток
- Контакт между клетками может стимулировать остановку клеточного цикла, вызывая остановку деления клеток, что называется контактным ингибированием .
- Межклеточный контакт может стимулировать клеточную дифференцировку .
- Генетические и эпигенетические изменения, при которых естественный отбор измененных клеток потенциально приводит к чрезмерному росту аномальных, адаптированных к культуре клеток со сниженной дифференцировкой и повышенной пролиферативной способностью. [37]
Выбор культуральной среды может повлиять на физиологическую значимость результатов экспериментов с культурами клеток из-за различий в составе и концентрации питательных веществ. [38] Недавно была продемонстрирована систематическая погрешность в сгенерированных наборах данных для CRISPR и RNAi скринингов подавления генов . [39] и для метаболического профиля линий раковых клеток . [38] Использование питательной среды , которая лучше отражает физиологические уровни питательных веществ, может повысить физиологическую значимость исследований in vitro , а в последнее время и таких типов сред, как Plasmax. [40] и плазмоподобная среда человека (HPLM), [41] были разработаны.
Манипуляции с культивируемыми клетками [ править ]
Среди распространенных манипуляций, проводимых с культуральными клетками, - смена среды, пассирование клеток и трансфекция клеток.Обычно они выполняются с использованием методов культуры тканей, основанных на асептике . Асептическая техника направлена на предотвращение загрязнения бактериями, дрожжами или другими клеточными линиями. Манипуляции обычно проводятся в боксе биобезопасности или в боксе с ламинарным потоком, чтобы исключить попадание микроорганизмов. антибиотики (например, пенициллин и стрептомицин ) и противогрибковые средства (например, амфотерицин B и раствор антибиотика-антимикотика В питательную среду также можно добавлять ).
Когда клетки подвергаются метаболическим процессам, вырабатывается кислота и уровень pH снижается. Часто индикатор pH в среду добавляют для измерения истощения питательных веществ.
Изменения в СМИ [ править ]
В случае приверженных культур среду можно удалить непосредственно аспирацией, а затем заменить. Замена среды в неприкрепившихся культурах включает центрифугирование культуры и ресуспендирование клеток в свежей среде.
Проходящие клетки [ править ]
Пассирование (также известное как субкультивирование или расщепление клеток) включает перенос небольшого количества клеток в новый сосуд. Клетки можно культивировать в течение более длительного времени, если их регулярно расщеплять, поскольку это позволяет избежать старения, связанного с длительной высокой плотностью клеток. Суспензионные культуры легко пассируются небольшим количеством культуры, содержащей несколько клеток, разведенных в большем объеме свежей среды. Для адгезивных культур сначала необходимо отделить клетки; обычно это делается со смесью трипсин - ЭДТА ; однако теперь для этой цели доступны другие смеси ферментов. Затем небольшое количество отдельных клеток можно использовать для посева новой культуры. Некоторые клеточные культуры, такие как клетки RAW, механически соскребают с поверхности сосуда резиновыми скребками.
Трансфекция и трансдукция [ править ]
Другой распространенный метод манипулирования клетками включает введение чужеродной ДНК путем трансфекции . Это часто выполняется, чтобы заставить клетки ген экспрессировать интересующий . Совсем недавно трансфекция конструкций РНКи была реализована как удобный механизм подавления экспрессии определенного гена/белка. ДНК также можно вставлять в клетки с помощью вирусов методами, называемыми трансдукцией , инфицированием или трансформацией . Вирусы как паразитические агенты хорошо подходят для внедрения ДНК в клетки, поскольку это часть их нормального процесса размножения.
клеток Установленные линии человека
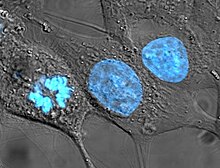
Клеточные линии, происходящие от человека, вызывают некоторые споры в биоэтике , поскольку они могут пережить свой родительский организм и позже использоваться для открытия прибыльных методов лечения. В своем новаторском решении в этой области Верховный суд Калифорнии постановил в деле Мур против Регентов Калифорнийского университета , что пациенты-люди не имеют прав собственности на клеточные линии, полученные из органов, удаленных с их согласия. [42]
Можно слить нормальные клетки с иммортализованной клеточной линией . Этот метод используется для получения моноклональных антител . Короче говоря, лимфоциты, выделенные из селезенки (или, возможно, крови) иммунизированного животного, объединяют с бессмертной клеточной линией миеломы (линия В-клеток) с получением гибридомы , которая обладает антителоспецифичностью первичного лимфоцита и бессмертием миеломы. Селективную питательную среду (HA или HAT) используют для отбора неслитых клеток миеломы; первичные лимфоктии быстро погибают в культуре, и выживают только слитые клетки. Их проверяют на выработку необходимых антител, обычно в пулах вначале, а затем после однократного клонирования.
Клеточные штаммы [ править ]
Штамм клеток получают либо из первичной культуры, либо из клеточной линии путем отбора или клонирования клеток, обладающих специфическими свойствами или характеристиками, которые необходимо определить. Клеточные штаммы — это клетки, адаптированные к культуре, но, в отличие от клеточных линий, имеющие ограниченный потенциал деления. Неиммортализованные клетки перестают делиться после 40–60 удвоений популяции. [43] и после этого они теряют способность к размножению (генетически детерминированное событие, известное как старение). [44]
Применение клеточных культур [ править ]
Массовое культивирование линий животных клеток имеет основополагающее значение для производства вирусных вакцин и других продуктов биотехнологии. Культура стволовых клеток человека используется для увеличения количества клеток и дифференциации клеток в различные типы соматических клеток для трансплантации. [45] Культуру стволовых клеток также используют для сбора молекул и экзосом, которые выделяют стволовые клетки, в целях разработки терапевтических средств. [46]
Биологические продукты, полученные с помощью технологии рекомбинантной ДНК (рДНК) в культурах клеток животных, включают ферменты , синтетические гормоны , иммунобиологические препараты ( моноклональные антитела , интерлейкины , лимфокины ) и противораковые средства . Хотя многие более простые белки можно получить с использованием рДНК в бактериальных культурах, более сложные гликозилированные ( модифицированные углеводами) белки в настоящее время необходимо производить в клетках животных. Важным примером такого сложного белка является гормон эритропоэтин . Стоимость выращивания культур клеток млекопитающих высока, поэтому проводятся исследования по получению таких сложных белков в клетках насекомых или высших растений, использованию одиночных эмбриональных клеток и соматических эмбрионов в качестве источника для прямого переноса генов посредством бомбардировки частицами, экспрессии транзитных генов и наблюдение конфокальной микроскопии является одним из его применений. Предлагается также подтвердить одноклеточное происхождение соматических эмбрионов и асимметрию первого клеточного деления, запускающего этот процесс.
Культура клеток также является ключевым методом клеточного сельского хозяйства , целью которого является создание как новых продуктов, так и новых способов производства существующих сельскохозяйственных продуктов, таких как молоко, (выращенное) мясо , ароматизаторы и рог носорога из клеток и микроорганизмов. Поэтому это считается одним из средств достижения сельского хозяйства без животных . Это также центральный инструмент для обучения клеточной биологии. [47]
Клеточная культура измерениях в двух
Исследования в области тканевой инженерии , стволовых клеток и молекулярной биологии в основном включают культивирование клеток на плоских пластиковых чашках. Этот метод известен как двумерная (2D) клеточная культура, и был впервые разработан Вильгельмом Ру, который в 1885 году удалил часть мозговой пластинки эмбрионального цыпленка и выдержал ее в теплом физиологическом растворе в течение нескольких дней на плоском стекле. тарелка. Благодаря развитию полимерных технологий возникла сегодняшняя стандартная пластиковая чашка для двумерной культуры клеток, широко известная как чашка Петри . Юлиусу Рихарду Петри , немецкому бактериологу , приписывают это изобретение, когда он работал помощником Роберта Коха . Различные исследователи сегодня также используют лабораторные колбы для культивирования , конические контейнеры и даже одноразовые пакеты, подобные тем, которые используются в одноразовых биореакторах .
Помимо чашек Петри, ученые уже давно выращивают клетки в матрицах биологического происхождения, таких как коллаген или фибрин, а в последнее время и в синтетических гидрогелях, таких как полиакриламид или ПЭГ. Они делают это, чтобы вызвать фенотипы, которые не экспрессируются на традиционно жестких субстратах. Растет интерес к контролю жесткости матрицы . [48] концепция, которая привела к открытиям в таких областях, как:
- Самообновление стволовых клеток [49] [50]
- Спецификация происхождения [51]
- Фенотип раковых клеток [52] [53] [54]
- Фиброз [55] [56]
- Функция гепатоцитов [57] [58] [59]
- механоощущение [60] [61] [62]
Клеточная культура в трех измерениях [ править ]
Клеточная культура в трех измерениях рекламировалась как «Новое измерение биологии». [63] В настоящее время практика культивирования клеток по-прежнему основана на различных комбинациях одиночных или множественных клеточных структур в 2D. [64] В настоящее время наблюдается рост использования 3D-культур клеток в исследовательских областях, включая разработку лекарств , биологию рака, регенеративную медицину , наноматериалов оценку и фундаментальные исследования в области медико-биологических наук . [65] [66] [67] 3D-культуры клеток можно выращивать с использованием каркаса или матрицы или без каркаса. Культуры на основе каркаса используют бесклеточную трехмерную матрицу или жидкую матрицу. Безкаркасные методы обычно создаются в суспензиях. [68] Существует множество платформ, используемых для облегчения роста трехмерных клеточных структур, включая каркасные системы, такие как гидрогелевые матрицы. [69] и твердые каркасы, а также системы без каркасов, такие как пластины с низкой адгезией, магнитная левитация с помощью наночастиц , [70] подвесные тарелки, [71] [72] и вращающаяся клеточная культура. Культивирование клеток в 3D приводит к широким различиям в характеристиках экспрессии генов и частично имитирует ткани в физиологических состояниях. [73] 3D-модель клеточной культуры продемонстрировала рост клеток, аналогичный росту клеток in vivo, чем монослойная культура, и все три культуры были способны поддерживать рост клеток. [74] Поскольку 3D-культуры были разработаны, оказалось, что они имеют большой потенциал для разработки моделей опухолей и исследования злокачественной трансформации и метастазирования. 3D-культуры могут предоставить расширенный инструмент для понимания изменений, взаимодействий и клеточной передачи сигналов. [75]
культура клеток в каркасах - 3D
Эрик Саймон в отчете о гранте NIH SBIR 1988 года показал, что электропрядение можно использовать для производства нано- и субмикронных волокнистых каркасов из полистирола и поликарбоната, специально предназначенных для использования в качестве клеточных субстратов in vitro . Это раннее использование электропряденых волокнистых решеток для клеточной культуры и тканевой инженерии показало, что различные типы клеток, включая фибробласты крайней плоти человека (HFF), трансформированную карциному человека (HEp-2) и эпителий легких норки (MLE), будут прикрепляться к поликарбонатным волокнам и размножаться на них. . Было отмечено, что в отличие от уплощенной морфологии, обычно наблюдаемой в 2D-культуре, клетки, выращенные на волокнах электропрядения, демонстрировали более гистотипическую округлую трехмерную морфологию, обычно наблюдаемую in vivo . [17]
3D-культура клеток в гидрогелях [ править ]
Поскольку природный внеклеточный матрикс (ЕСМ) важен для выживания, пролиферации, дифференциации и миграции клеток, различные гидрогелевые культуральные матрицы, имитирующие естественную структуру ЕСМ, рассматриваются как потенциальные подходы к культивированию клеток, подобному in vivo. [76] Гидрогели состоят из взаимосвязанных пор с высокой степенью удержания воды, что обеспечивает эффективный транспорт таких веществ, как питательные вещества и газы. Для 3D-культуры клеток доступно несколько различных типов гидрогелей из натуральных и синтетических материалов, включая гидрогели экстракта ЕСМ животных, белковые гидрогели, пептидные гидрогели, полимерные гидрогели и гидрогель наноцеллюлозы на основе древесины .
культивирование клеток с помощью магнитной 3D - левитации
Метод 3D-клеточного культивирования методом магнитной левитации (MLM) представляет собой применение выращивания 3D-тканей путем воздействия на клетки, обработанные сборками магнитных наночастиц, в пространственно изменяющихся магнитных полях с использованием неодимовых магнитных драйверов и стимулирования межклеточных взаимодействий путем подъема клеток в воздух. Жидкостная граница стандартной чашки Петри. Сборки магнитных наночастиц состоят из магнитных наночастиц оксида железа, наночастиц золота и полимера полилизина. Трехмерное культивирование клеток масштабируется: можно культивировать от 500 клеток до миллионов клеток или от одной чашки до высокопроизводительных систем малого объема.
и Тканевая инженерия культура
Культура клеток является фундаментальным компонентом культуры тканей и тканевой инженерии , поскольку она закладывает основы выращивания и поддержания клеток in vitro .Основное применение культуры клеток человека находится в производстве стволовых клеток, где мезенхимальные стволовые клетки можно культивировать и криоконсервировать для будущего использования. Тканевая инженерия потенциально предлагает значительные улучшения в недорогом медицинском обслуживании сотен тысяч пациентов ежегодно.
Вакцины [ править ]
Вакцины от полиомиелита , кори , эпидемического паротита , краснухи и ветряной оспы в настоящее время производятся на клеточных культурах. Из-за H5N1 угрозы пандемии исследования по использованию клеточной культуры для вакцин против гриппа финансируются правительством США . Новаторские идеи в этой области включают вакцины на основе рекомбинантной ДНК , например вакцину, созданную с использованием аденовируса человека (вируса простуды) в качестве вектора, [77] [78] и новые адъюванты. [79]
Совместное культивирование клеток [ править ]
Метод совместного культивирования используется для изучения перекрестных помех между двумя или более типами клеток на планшете или в трехмерной матрице. Культивирование различных стволовых клеток и взаимодействие иммунных клеток можно исследовать на модели in vitro, аналогичной биологической ткани. Поскольку большинство тканей содержат более одного типа клеток, важно оценить их взаимодействие в среде трехмерной культуры, чтобы лучше понять их взаимодействие и ввести миметические ткани. Существует два типа совместного культивирования: прямое и косвенное. В то время как прямое взаимодействие предполагает, что клетки находятся в прямом контакте друг с другом в одной и той же культуральной среде или матрице, непрямое взаимодействие включает в себя разные среды, что позволяет участвовать сигнальным и растворимым факторам. [15] [80]
Дифференцировку клеток в тканевых моделях во время взаимодействия между клетками можно изучать с помощью системы совместного культивирования для моделирования раковых опухолей, для оценки влияния лекарств на терапевтические испытания и для изучения влияния лекарств на терапевтические испытания. Система совместного культивирования в 3D-моделях может предсказать реакцию на химиотерапию и эндокринную терапию, если микроокружение определяет биологическую ткань для клеток.
Метод совместного культивирования используется в тканевой инженерии для создания тканей при непосредственном взаимодействии нескольких клеток. [81]

клеток в микрофлюидном Культура устройстве
Техника микрофлюидики представляет собой разработанные системы, которые могут выполнять процессы в потоке, размер которого обычно составляет микроны. Микрофлюидные чипы также известны как «лаборатория на чипе», и они могут проводить непрерывные процедуры и этапы реакции с запасным количеством реагентов и пространством. Такие системы позволяют идентифицировать и изолировать отдельные клетки и молекулы в сочетании с соответствующими биологическими анализами и методами высокочувствительного обнаружения. [82] [83]
Орган-на-чипе [ править ]
Системы OoC имитируют и контролируют микроокружение клеток путем выращивания тканей в микрофлюидике. Сочетая тканевую инженерию, производство биоматериалов и клеточную биологию, он дает возможность создать биомиметическую модель для изучения заболеваний человека в лаборатории. В последние годы наука о 3D-культуре клеток добилась значительного прогресса, что привело к развитию OoC. ОоС рассматривается как доклинический этап, который приносит пользу фармацевтическим исследованиям, разработке лекарств и моделированию заболеваний. [84] [85] OoC — это важная технология, которая может преодолеть разрыв между тестированием на животных и клиническими исследованиями, а также благодаря достижениям науки может заменить исследования in vivo по доставке лекарств и патофизиологические исследования. [86]
немлекопитающих Культура клеток
Помимо культуры хорошо зарекомендовавших себя иммортализованных клеточных линий, клетки из первичных эксплантов множества организмов можно культивировать в течение ограниченного периода времени, прежде чем наступит старение (см. Предел Хейфлика). Культивированные первичные клетки широко использовались в исследованиях, как и кератоциты рыб в исследованиях миграции клеток. [87] [47] [88]
культивирования Методы клеток растительных
Культуры растительных клеток обычно выращивают в виде суспензионных культур клеток в жидкой среде или в виде каллусных культур на твердой среде. Культивирование недифференцированных растительных клеток и каллусов требует надлежащего баланса гормонов роста растений ауксина и цитокинина . [ нужна ссылка ]
Культура клеток насекомых [ править ]
Клетки, полученные из Drosophila melanogaster (в первую очередь, клетки Шнайдера 2 ), можно использовать для экспериментов, которые может быть трудно провести на живых мухах или личинках, таких как биохимические исследования или исследования с использованием миРНК . Клеточные линии, полученные из армейского червя Spodoptera frugiperda , включая Sf9 и Sf21 , и из капустной петельки Trichoplusia ni , клетки High Five , обычно используются для экспрессии рекомбинантных белков с использованием бакуловируса . [89]
Методы культивирования бактерий и дрожжей [ править ]
Для бактерий и дрожжей небольшие количества клеток обычно выращивают на твердой подложке, содержащей внедренные в нее питательные вещества, обычно это гель, такой как агар, тогда как крупномасштабные культуры выращиваются на клетках, суспендированных в питательном бульоне. [ нужна ссылка ]
культуры вирусной Методы
Культура вирусов требует культуры клеток млекопитающих, растений, грибкового или бактериального происхождения в качестве хозяев для роста и репликации вируса. Целые вирусы дикого типа , рекомбинантные вирусы или вирусные продукты могут быть получены в типах клеток, отличных от их естественных хозяев, при правильных условиях. В зависимости от вида вируса инфекция и репликация вируса могут привести к лизису клетки-хозяина и образованию вирусной бляшки . [ нужна ссылка ]
клеточные Общие линии
- Клеточные линии человека
- DU145 ( рак простаты )
- H295R ( адренокортикальный рак )
- HeLa ( рак шейки матки )
- КБМ-7 ( хронический миелогенный лейкоз )
- LNCaP (рак простаты)
- MCF-7 ( рак молочной железы )
- MDA-MB-468 (рак молочной железы)
- PC3 (рак простаты)
- SaOS-2 ( рак кости )
- SH-SY5Y ( нейробластома , клонированная из миеломы )
- Т-47Д (рак молочной железы)
- THP-1 острый миелолейкоз ( )
- У-87 МГ ( глиобластома )
- Национального института рака Панель из 60 линий раковых клеток ( NCI60 )
- животных Линии клеток
- Веро африканской зеленой мартышки Chlorocebus ) клеток почек эпителиальных ( линия
- Клетка BHK21 (почка детеныша хомячка)
- Клетка MDBK (бычья почка Мадина-Дарби)
- DF-1клетка (куриный фибробласт)
- Мышиные клеточные линии
- крысы Линии опухолевых клеток
- GH3 ( опухоль гипофиза )
- PC12 ( феохромоцитома )
- Линии растительных клеток
- Клетки табака BY-2 (хранятся в виде суспензионной культуры клеток , представляют собой модельную систему растительных клеток)
- Клеточные линии других видов
Список клеточных линий [ править ]
| Клеточная линия | Значение | Организм | Ткань происхождения | Морфология | Ссылки |
|---|---|---|---|---|---|
| 3Т3-Л1 | «3-дневный трансфер, инокулят 3 х 10^5 клеток» | Мышь | Эмбрион | Фибробласт | ECACC Целлозавр |
| 4Т1 | Мышь | Молочная железа | ATCC Целлозавр | ||
| 1321Н1 | Человек | Мозг | Астроцитома | ECACC Целлозавр | |
| 9л | Крыса | Мозг | Глиобластома | ECACC Целлозавр | |
| А172 | Человек | Мозг | Глиобластома | ECACC Целлозавр | |
| А20 | Мышь | В- лимфома | В- лимфоцит | Целлозавр | |
| А253 | Человек | Подчелюстной проток | Рак головы и шеи | ATCC Целлозавр | |
| А2780 | Человек | яичник | Карцинома яичника | ECACC Целлозавр | |
| А2780АДР | Человек | яичник | Устойчивое к адриамицину производное A2780 | ECACC Целлозавр | |
| A2780cis | Человек | яичник | Устойчивое к цисплатину производное A2780 | ECACC Целлозавр | |
| А431 | Человек | Эпителий кожи | Плоскоклеточная карцинома | ECACC Целлозавр | |
| А549 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| АБ9 | данио | КОНЕЦ | Фибробласт | ATCC Целлозавр | |
| АХЛ-1 | Армянский хомяк Лунг-1 | Хомяк | Легкое | ECACC Целлозавр | |
| ОДО | Мышь | Костный мозг | Строма | ПМИД 2435412 [90] Целлозавр | |
| Б16 | Мышь | Меланома | ECACC Целлозавр | ||
| Б35 | Крыса | Нейробластома | ATCC Целлозавр | ||
| БКП-1 | Человек | КПМК | ВИЧ+ первичная выпотная лимфома | ATCC Целлозавр | |
| БЕАС-2Б | Эпителий бронхов + гибрид вируса аденовируса 12-SV40 (Ad12SV40) | Человек | Легкое | Эпителиальный | ECACC Целлозавр |
| изгиб.3 | Эндотелий головного мозга 3 | Мышь | Мозг/ кора головного мозга | Эндотелий | Целлозавр |
| БХК-21 | Детеныш хомячка Почка-21 | Хомяк | Почка | Фибробласт | ECACC Целлозавр |
| БОСК23 | Упаковочная клеточная линия, полученная из HEK 293 | Человек | Почка (эмбриональная) | Эпителий | Целлозавр |
| БТ-20 | Опухоль молочной железы-20 | Человек | Эпителий молочной железы | Карцинома молочной железы | ATCC Целлозавр |
| БхПК-3 | Биопсия ксенотрансплантата рака поджелудочной железы линии 3 | Человек | Аденокарцинома поджелудочной железы | Эпителиальный | ECACC Целлозавр |
| C2C12 | Мышь | миобласт | ECACC Целлозавр | ||
| C3H-10T1/2 | Мышь | Эмбриональная мезенхимальная клеточная линия | ECACC Целлозавр | ||
| С6 | Крыса | Мозговой астроцит | Глиома | ECACC Целлозавр | |
| С6/36 | Насекомое — азиатский тигровый комар | Личиночная ткань | ECACC Целлозавр | ||
| Како-2 | Человек | Двоеточие | Колоректальный рак | ECACC Целлозавр | |
| Кал-27 | Человек | Язык | Плоскоклеточная карцинома | ATCC Целлозавр | |
| Калу-3 | Человек | Легкое | Аденокарцинома | ATCC Целлозавр | |
| CGR8 | Мышь | Эмбриональные стволовые клетки | ECACC Целлозавр | ||
| ДАВАТЬ | Яичник китайского хомяка | Хомяк | яичник | Эпителий | ECACC Целлозавр |
| ХМЛ Т1 | Хронический миелолейкоз Т-лимфоцит 1 | Человек | Острая фаза ХМЛ | Т-клеточный лейкоз | ДСМЗ Целлозавр |
| СМТ12 | Опухоль молочной железы собаки 12 | Собака | Молочная железа | Эпителий | Целлозавр |
| КОР-L23 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| КОР-L23/5010 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| COR-L23/СЛР | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| КОР-L23/R23- | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| КОС-7 | Cercopithecus aethiops , дефектный по происхождению СВ-40 | Обезьяна Старого Света — Cercopithecus aethiops ( Chlorocebus ) | Почка | Фибробласт | ECACC Целлозавр |
| COV-434 | Человек | яичник | Гранулезно-клеточная карцинома яичника | ПМИД 8436435 [91] ECACC Целлозавр | |
| CT26 | Мышь | Двоеточие | Колоректальный рак | Целлозавр | |
| Д17 | Собака | Метастазы в легкие | Остеосаркома | ATCC Целлозавр | |
| ДАОЙ | Человек | Мозг | Медуллобластома | ATCC Целлозавр | |
| ДХ82 | Собака | Гистиоцитоз | Моноциты / макрофаги | ECACC Целлозавр | |
| ДУ145 | Человек | Андрогенонечувствительная карцинома простаты | ATCC Целлозавр | ||
| ДуКаП | Рак твердой мозговой оболочки простаты | Человек | Метастатический рак простаты | Эпителиальный | PMID 11317521 [92] Целлозавр |
| E14Tg2a | Мышь | Эмбриональные стволовые клетки | ECACC Целлозавр | ||
| EL4 | Мышь | Т-клеточный лейкоз | ECACC Целлозавр | ||
| ЭМ-2 | Человек | Взрывной кризис ХМЛ | Линия Ph+ ХМЛ | ДСМЗ Целлозавр | |
| ЭМ-3 | Человек | Взрывной кризис ХМЛ | Линия Ph+ ХМЛ | ДСМЗ Целлозавр | |
| ЕМТ6/АР1 | Мышь | Молочная железа | Эпителиальноподобный | ECACC Целлозавр | |
| ЕМТ6/АР10.0 | Мышь | Молочная железа | Эпителиальноподобный | ECACC Целлозавр | |
| FM3 | Человек | Метастазирование в лимфатические узлы | Меланома | ECACC Целлозавр | |
| GL261 | Глиома 261 | Мышь | Мозг | Глиома | Целлозавр |
| H1299 | Человек | Легкое | Карцинома легких | ATCC Целлозавр | |
| ХаКаТ | Человек | Кожа | Кератиноциты | CLS Целлозавр | |
| ГКА2 | Человек | Двоеточие | Аденокарцинома | ECACC Целлозавр | |
| 293 крон | Эмбриональная почка человека 293 | Человек | Почка (эмбриональная) | Эпителий | ECACC Целлозавр |
| ХЕК 293Т | HEK 293 производная | Человек | Почка (эмбриональная) | Эпителий | ECACC Целлозавр |
| Весь | «Генриетта Лакс» | Человек | Эпителий шейки матки | Рак шейки матки | ECACC Целлозавр |
| Гепа1c1c7 | Клон 7 клона 1 гепатомы линии 1 | Мышь | Гепатома | Эпителиальный | ECACC Целлозавр |
| Всегда G2 | Человек | Печень | Гепатобластома | ECACC Целлозавр | |
| Дай пять | Насекомое (моль) — Trichoplusia ni | яичник | Целлозавр | ||
| ХЛ-60 | Лейкемия человека-60 | Человек | Кровь | Миелобласт | ECACC Целлозавр |
| ХТ-1080 | Человек | Фибросаркома | ECACC Целлозавр | ||
| ХТ-29 | Человек | Эпителий толстой кишки | Аденокарцинома | ECACC Целлозавр | |
| J558L | Мышь | Миелома | В-лимфоцитная клетка | ECACC Целлозавр | |
| Юркат | Человек | Белые кровяные тельца | Т-клеточный лейкоз | ECACC Целлозавр | |
| ТЫ | Человек | лимфобластоид | В-клетка, трансформированная EBV | ECACC Целлозавр | |
| К562 | Человек | лимфобластоид | Взрывной кризис ХМЛ | ECACC Целлозавр | |
| КБМ-7 | Человек | лимфобластоид | Взрывной кризис ХМЛ | Целлозавр | |
| КСЛ-22 | Человек | лимфобластоид | ХМЛ | ДСМЗ Целлозавр | |
| КГ1 | Человек | лимфобластоид | ПОД | ECACC Целлозавр | |
| Ку812 | Человек | лимфобластоид | Эритролейкемия | ECACC Целлозавр | |
| КЁ-1 | Киото-1 | Человек | лимфобластоид | ХМЛ | ДСМЗ Целлозавр |
| Л1210 | Мышь | Лимфоцитарный лейкоз | Асцитическая жидкость | ECACC Целлозавр | |
| Л243 | Мышь | Гибридома | Секретирует mAb L243 (против HLA-DR) | ATCC Целлозавр | |
| ЛНКаП | Рак лимфатического узла простаты | Человек | Аденокарцинома предстательной железы | Эпителиальный | ECACC Целлозавр |
| МА-104 | Микробиологические ассоциации-104 | Африканская зеленая обезьяна | Почка | Эпителиальный | Целлозавр |
| МА2.1 | Мышь | Гибридома | Секретирует mAb MA2.1 (против HLA-A2 и HLA-B17). | ATCC Целлозавр | |
| Ма-Мел 1, 2, 3....48 | Человек | Кожа | Ряд меланомы клеточных линий | ECACC Целлозавр | |
| МС-38 | Мышь Колон-38 | Мышь | Двоеточие | Аденокарцинома | Целлозавр |
| МЦФ-7 | Мичиганский онкологический фонд-7 | Человек | Грудь | Инвазивная протоковая карцинома молочной железы ER+, PR+ | ECACC Целлозавр |
| МЦФ-10А | Мичиганский онкологический фонд-10A | Человек | Эпителий молочной железы | ATCC Целлозавр | |
| МДА-МБ-157 | Доктор медицины Андерсон - Метастатическая грудь-157 | Человек | Метастазирование плеврального выпота | Карцинома молочной железы | ECACC Целлозавр |
| МДА-МБ-231 | Доктор медицины Андерсон - Метастатическая грудь-231 | Человек | Метастазирование плеврального выпота | Карцинома молочной железы | ECACC Целлозавр |
| МДА-МБ-361 | Доктор медицины Андерсон - Метастатическая грудь-361 | Человек | Меланома (загрязненная М14) | ECACC Целлозавр | |
| МДА-МБ-468 | Доктор медицины Андерсон - Метастатическая грудь-468 | Человек | Метастазирование плеврального выпота | Карцинома молочной железы | ATCC Целлозавр |
| МДСК II | Мадин Дарби Canine Kidney II | Собака | Почка | Эпителий | ECACC Целлозавр |
| МГ63 | Человек | Кость | Остеосаркома | ECACC Целлозавр | |
| МИА ПаКа-2 | Человек | Простата | Карцинома поджелудочной железы | ATCC Целлозавр | |
| МАТЬ/0.2R | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| Моно-Мак-6 | Человек | Белые кровяные тельца | Миелоидно-метапластический ОМЛ | ДСМЗ Целлозавр | |
| МРК-5 | Клеточные штаммы 5 Совета медицинских исследований | Человек | Легкие (фетальные) | Фибробласт | ECACC Целлозавр |
| МТД-1А | Мышь | Эпителий | Целлозавр | ||
| МойКонец | Эндотелиальный миокард | Мышь | Эндотелий | Целлозавр | |
| НЦИ-H69 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| НЦИ-H69/СЛР | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| НЦИ-H69/LX10 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| НЦИ-H69/LX20 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| НЦИ-H69/LX4 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| Нейро-2а | Мышь | Нерв/ нейробластома | Нейрональные стволовые клетки | ECACC Целлозавр | |
| НИЗ-3Т3 | НИЗ , 3-дневный перенос, инокулят 3 х 10 5 клетки | Мышь | Эмбрион | Фибробласт | ECACC Целлозавр |
| НАЛМ-1 | Человек | Периферическая кровь | Взрывной кризис ХМЛ | ATCC Целлозавр | |
| НК-92 | Человек | Лейкемия/лимфома | ATCC Целлозавр | ||
| ШАГ-2 | Человек | Метастазы в легкие | Эмбриональный рак | ECACC Целлозавр | |
| СЗ-145 | Человек | Кожа | Меланома | ESTDAB. Архивировано 16 ноября 2011 г. в Wayback Machine Cellosaurus. | |
| ХОРОШО | Опоссум Почка | Виргинский опоссум — Didelphis Virginiana | Почка | ECACC Целлозавр | |
| Клеточные линии OPCN/OPCT | Человек | Простата | Диапазон линий опухоли простаты | Целлозавр | |
| P3X63Ag8 | Мышь | Миелома | ECACC Целлозавр | ||
| ПАНК-1 | Человек | воздуховод | Эпителиоидная карцинома | ATCC Целлозавр | |
| ПК12 | Крыса | Мозговое вещество надпочечников | Феохромоцитома | ECACC Целлозавр | |
| ПК-3 | Рак простаты-3 | Человек | Костные метастазы | Карцинома простаты | ECACC Целлозавр |
| Вглядеться | Человек | Т-клеточный лейкоз | ДСМЗ Целлозавр | ||
| ПНТ1А | Человек | Простата | SV40-трансформированная опухолевая линия | ECACC Целлозавр | |
| ПНТ2 | Человек | Простата | SV40-трансформированная опухолевая линия | ECACC Целлозавр | |
| Пт К2 | Вторая клеточная линия, полученная из Potorous tridactylis. | Long-nosed potoroo - Potorous tridactylus | Почка | Эпителиальный | ECACC Целлозавр |
| Раджи | Человек | В- лимфома | лимфобластоподобный | ECACC Целлозавр | |
| РБЛ-1 | Крысиный базофильный лейкоз-1 | Крыса | Лейкемия | Базофильная клетка | ECACC Целлозавр |
| Ренка | Карцинома почки | Мышь | Почка | Рак почки | ATCC Целлозавр |
| РИН-5Ф | Мышь | поджелудочная железа | ECACC Целлозавр | ||
| РМА-С | Мышь | Т-клеточная опухоль | Целлозавр | ||
| С2 | Шнайдер 2 | Насекомое — Drosophila melanogaster | Эмбрионы поздней стадии (20–24 часа) | ATCC Целлозавр | |
| СаОС-2 | Саркома ОСтеогенная-2 | Человек | Кость | Остеосаркома | ECACC Целлозавр |
| Sf21 | Spodoptera frugiperda 21 | Насекомое (моль) — Spodoptera frugiperda | яичник | ECACC Целлозавр | |
| Сф9 | Spodoptera frugiperda 9 | Насекомое (моль) — Spodoptera frugiperda | яичник | ECACC Целлозавр | |
| SH-SY5Y | Человек | Метастазы в костный мозг | Нейробластома | ECACC Целлозавр | |
| Нет | Человек | Эпителий шейки матки | Рак шейки матки | ATCC Целлозавр | |
| СК-БР-3 | Слоан-Кеттеринг Рак молочной железы 3 | Человек | Грудь | Карцинома молочной железы | ДСМЗ Целлозавр |
| СК-ОВ-3 | Слоана-Кеттеринга 3 Рак яичников | Человек | яичник | Карцинома яичника | ECACC Целлозавр |
| СК-Н-Ш | Человек | Мозг | Эпителиальный | ATCC Целлозавр | |
| Т2 | Человек | Т-клеточный лейкоз/ гибридома В-клеточной линии | ATCC Целлозавр | ||
| Т-47Д | Человек | Грудь | Протоковая карцинома молочной железы | ECACC Целлозавр | |
| Т84 | Человек | Метастазы в легкие | Колоректальный рак | ECACC Целлозавр | |
| Т98Г | Человек | Глиобластома-астроцитома | Эпителий | ECACC Целлозавр | |
| ТЭЦ-1 | Человек | моноцит | Острый моноцитарный лейкоз | ECACC Целлозавр | |
| U2OS | Человек | Остеосаркома | Эпителиальный | ECACC Целлозавр | |
| U373 | Человек | Глиобластома-астроцитома | Эпителий | ECACC Целлозавр | |
| U87 | Человек | Глиобластома-астроцитома | Эпителиальноподобный | ECACC Целлозавр | |
| U937 | Человек | Лейкемическая моноцитарная лимфома | ECACC Целлозавр | ||
| ВКаП | Позвоночный рак простаты | Человек | Метастазы в позвонки | Карцинома простаты | ECACC Целлозавр |
| Истинный | На эсперанто: verda (зеленый, зеленая обезьяна), reno (почка). | Африканская зеленая мартышка — Chlorocebus sabaeus. | Эпителий почек | ECACC Целлозавр | |
| ВГ-1 | Человек | Первичная выпотная лимфома | Целлозавр | ||
| WM39 | Человек | Кожа | Меланома | ESTDAB Целлозавр | |
| ВТ-49 | Человек | лимфобластоид | ECACC Целлозавр | ||
| ЯЦ-1 | Мышь | Лимфома | ECACC Целлозавр | ||
| МАЛЕНЬКИЙ | Человек | лимфобластоид | В-клетка, трансформированная EBV | Иммунология человека [93] ECACC Целлозавр |
См. также [ править ]
- Биологическое бессмертие
- Анализы клеточных культур
- Измерение импеданса электрического элемента-подложки
- Список зараженных клеточных линий
- Список NCI-60 клеточных линий
- Список панели LL-100 клеточных линий
- Список клеточных линий рака молочной железы
- Микрофизиометрия
Ссылки и примечания [ править ]
- ↑ Перейти обратно: Перейти обратно: а б Тейлор М.В. (2014). «История клеточной культуры». Вирусы и человек: история взаимодействия . Чам: Международное издательство Springer. стр. 41–52. дои : 10.1007/978-3-319-07758-1_3 . ISBN 978-3-319-07757-4 .
- ^ Харрис А.Р., Питер Л., Беллис Дж., Баум Б., Кабла А.Дж., Чаррас Г.Т. (октябрь 2012 г.). «Характеристика механики культивируемых клеточных монослоев» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (41): 16449–16454. Бибкод : 2012PNAS..10916449H . дои : 10.1073/pnas.1213301109 . ПМЦ 3478631 . ПМИД 22991459 .
- ^ «Некоторые вехи развития культуры тканей и клеток» . Проверено 19 апреля 2006 г.
- ^ «Клеточная культура» . Проверено 19 апреля 2006 г.
- ^ «Whonamedit — решение Рингера» . whonamedit.com . Проверено 9 июня 2014 г.
- ^ Стейнхардт Э., Израильтянин С., Ламберт Р.А. (1913). «Исследования по культивированию вируса коровьей оспы». Журнал инфекционных болезней . 13 (2): 294–300. дои : 10.1093/infdis/13.2.294 . ISSN 0022-1899 . JSTOR 30073371 .
- ^ Атала А (2009). «Выращивание новых органов» . ТЕДМЕД . Проверено 23 августа 2021 г.
- ^ «Животные и альтернативы в тестировании» . Архивировано из оригинала 25 февраля 2006 года . Проверено 19 апреля 2006 г.
- ^ Фентем Дж.Х. (февраль 2006 г.). «Совместная работа над ответом на вызовы политики ЕС по замене испытаний на животных» . Альтернативы лабораторным животным . 34 (1): 11–18. дои : 10.1177/026119290603400116 . ПМИД 16522146 . S2CID 10339716 .
- ^ Шифф Дж.А. (февраль 2002 г.). «Невоспетый герой медицинских исследований» . Журнал выпускников Йельского университета . Архивировано из оригинала 14 ноября 2012 года . Проверено 19 апреля 2006 г.
- ^ Боннер Дж. (июнь 1936 г.). «Культуры растительных тканей с точки зрения гормонов» . Труды Национальной академии наук Соединенных Штатов Америки . 22 (6): 426–430. Бибкод : 1936PNAS...22..426B . дои : 10.1073/pnas.22.6.426 . JSTOR 86579 . ПМЦ 1076796 . ПМИД 16588100 .
- ^ Хаберландт, Г. (1902) Эксперименты с культурой изолированных растительных клеток. Зона встреч Академическая наука Вена. Математика-Естествознание кл., отд. J. 111, 69–92.
- ^ Ноэ AC (октябрь 1934 г.). «Готлиб Хаберландт» . Физиология растений . 9 (4): 850–855. дои : 10.1104/стр.9.4.850 . ПМК 439112 . ПМИД 16652925 .
- ^ Культура тканей растений . 100 лет со дня Готлиба Хаберландта. Лаймер, Маргит; Рюкер, Вальтрауд (ред.), 2003. Springer. ISBN 978-3-211-83839-6
- ↑ Перейти обратно: Перейти обратно: а б Каррел А., Берроуз М.Т. (март 1911 г.). «Культивирование тканей in vitro и метод ITS» . Журнал экспериментальной медицины . 13 (3): 387–396. дои : 10.1084/jem.13.3.387 . ПМК 2125263 . ПМИД 19867420 .
- ^ Мартин Б.М. (1 декабря 2013 г.). Методы культуры тканей: введение . Springer Science & Business Media. стр. 29–30. ISBN 978-1-4612-0247-9 .
- ↑ Перейти обратно: Перейти обратно: а б Саймон Э.М. (1988). «Заключительный отчет фазы I: Волокнистые субстраты для клеточных культур (R3RR03544A)» . Исследовательские ворота . Проверено 22 мая 2017 г.
- ^ Урри, Л.А., Кэмпбелл, Н.А., Каин, М.Л., Рис, Дж.Б., Вассерман, С. (2007). Биология. Соединенное Королевство: Издательская компания Бенджамина-Каммингса. п. 860
- ^ Фойгт Н., Пирман К.М., Добрев Д., Дибб К.М. (сентябрь 2015 г.). «Методы выделения предсердных клеток крупных млекопитающих и человека» . Журнал молекулярной и клеточной кардиологии . 86 : 187–198. дои : 10.1016/j.yjmcc.2015.07.006 . ПМИД 26186893 .
- ^ Луч В.Е., Шихан К.А., Вольска Б.М. (сентябрь 2011 г.). «Методы выделения, культивирования и переноса генов кардиомиоцитов» . Журнал молекулярной и клеточной кардиологии . 51 (3): 288–298. дои : 10.1016/j.yjmcc.2011.06.012 . ПМК 3164875 . ПМИД 21723873 .
- ^ Хемеда Х., Гибель Б., Вагнер В. (16 февраля 2014 г.) Оценка лизата тромбоцитов человека по сравнению с фетальной бычьей сывороткой для культуры мезенхимальных стромальных клеток Цитотерапия p170-180, выпуск 2 doi.10.1016
- ^ «Пост-блог | Бовал БиоСолюшнс, ООО» . bovalco.com. Архивировано из оригинала 10 сентября 2014 года . Проверено 2 декабря 2014 г.
- ^ «LipiMAX очищенный раствор липопротеинов из бычьей сыворотки» . Селборнские биологические службы . 2006. Архивировано из оригинала 19 июля 2012 года . Проверено 2 февраля 2010 г.
- ^ Портела В.М., Замберлам Г., Прайс, Калифорния (апрель 2010 г.). «Плотность клеточного покрытия изменяет соотношение экспрессии генов эстрогенных и прогестагенных ферментов в культивируемых гранулезных клетках» . Фертильность и бесплодие . 93 (6): 2050–2055. doi : 10.1016/j.fertnstert.2009.01.151 . ПМИД 19324349 .
- ^ Жаккар Н., Макаун Р.Дж., Супер А, Гриффин Л.Д., Верайтч Ф.С., Сита Н. (октябрь 2014 г.). «Автоматизированная и онлайн-характеристика роста культуры прикрепленных клеток в микрофабрикатном биореакторе» . Журнал автоматизации лабораторий . 19 (5): 437–443. дои : 10.1177/2211068214529288 . ПМК 4230958 . ПМИД 24692228 .
- ^ Хампель С. (октябрь 2015 г.). «Органотипические культуры срезов мозга: обзор» . Нейронаука . 305 : 86–98. doi : 10.1016/j.neuroscience.2015.07.086 . ПМК 4699268 . ПМИД 26254240 .
- ^ Неймарк Дж. (февраль 2015 г.). «Линия атаки» . Наука . 347 (6225): 938–940. Бибкод : 2015Sci...347..938N . дои : 10.1126/science.347.6225.938 . ПМИД 25722392 .
- ^ Дрекслер Х.Г., Диркс В.Г., Маклауд Р.А. (октябрь 1999 г.). «Ложные линии гемопоэтических клеток человека: перекрестное загрязнение и неверные интерпретации» . Лейкемия . 13 (10): 1601–1607. дои : 10.1038/sj.leu.2401510 . ПМИД 10516762 .
- ^ Дрекслер Х.Г., Маклауд Р.А., Диркс В.Г. (декабрь 2001 г.). «Перекрестное загрязнение: HS-Султан — это не миелома, а клеточная линия лимфомы Беркитта» . Кровь . 98 (12): 3495–3496. дои : 10.1182/blood.V98.12.3495 . ПМИД 11732505 .
- ^ Кабрера С.М., Кобо Ф., Ньето А., Кортес Х.Л., Монтес Р.М., Каталина П., Конча А. (июнь 2006 г.). «Тесты идентичности: определение перекрестного загрязнения клеточных линий» . Цитотехнология . 51 (2): 45–50. дои : 10.1007/s10616-006-9013-8 . ПМЦ 3449683 . ПМИД 19002894 .
- ↑ Перейти обратно: Перейти обратно: а б Чаттерджи Р. (февраль 2007 г.). «Клеточная биология. Случаи ошибочной идентификации». Наука . 315 (5814): 928–931. дои : 10.1126/science.315.5814.928 . ПМИД 17303729 . S2CID 13255156 .
- ^ Лискович М., Равид Д. (январь 2007 г.). «Пример ошибочной идентификации линий раковых клеток: клетки MCF-7/AdrR (переименованные в NCI/ADR-RES) получены из клеток карциномы яичника человека OVCAR-8». Письма о раке . 245 (1–2): 350–352. дои : 10.1016/j.canlet.2006.01.013 . ПМИД 16504380 .
- ^ Маклауд Р.А., Диркс В.Г., Мацуо Ю., Кауфманн М., Милч Х., Дрекслер Х.Г. (ноябрь 1999 г.). «Широко распространенное внутривидовое перекрестное заражение линий опухолевых клеток человека, возникающее в источнике» . Международный журнал рака . 83 (4): 555–563. doi : 10.1002/(SICI)1097-0215(19991112)83:4<555::AID-IJC19>3.0.CO;2-2 . ПМИД 10508494 .
- ^ Мастерс-младший (апрель 2002 г.). «Клетки HeLa 50 лет спустя: хорошие, плохие и ужасные». Обзоры природы. Рак . 2 (4): 315–319. дои : 10.1038/nrc775 . ПМИД 12001993 . S2CID 991019 .
- ↑ Перейти обратно: Перейти обратно: а б Данэм Дж. Х., Гутмиллер П. (2008). «Занимаемся хорошей наукой: проверка подлинности клеточной линии» (PDF) . Примечания к ячейке . 22 :15–17. Архивировано из оригинала (PDF) 28 октября 2008 года . Проверено 28 октября 2008 г.
- ^ Брендан П. Люси, Уолтер А. Нельсон-Рис, Гровер М. Хатчинс; Генриетта Лакс, клетки HeLa и загрязнение клеточной культуры. Arch Pathol Lab Med, 1 сентября 2009 г.; 133 (9): 1463–1467. дои: https://doi.org/10.5858/133.9.1463
- ^ Нгуен Х.Т., Гинс М., Спитс С. (2012). «Генетическая и эпигенетическая нестабильность плюрипотентных стволовых клеток человека» . Обновление репродукции человека . 19 (2): 187–205. дои : 10.1093/humupd/dms048 . ПМИД 23223511 .
- ↑ Перейти обратно: Перейти обратно: а б Лагзиэль С., Готлиб Э., Шломи Т. (декабрь 2020 г.). «Заботьтесь о своих СМИ». Природный метаболизм . 2 (12): 1369–1372. дои : 10.1038/s42255-020-00299-y . ПМИД 33046912 . S2CID 222319735 .
- ^ Лагзиел С., Ли В.Д., Шломи Т. (апрель 2019 г.). «Вывод о зависимости рака от метаболических генов на основе крупномасштабного генетического скрининга» . БМК Биология . 17 (1): 37. дои : 10.1186/s12915-019-0654-4 . ПМК 6489231 . ПМИД 31039782 .
- ^ Ванде Вурде Дж., Акерманн Т., Пфетцер Н., Самптон Д., Маккей Г., Кална Г. и др. (январь 2019 г.). «Улучшение метаболической точности моделей рака с помощью физиологической среды для культивирования клеток» . Достижения науки . 5 (1): eaau7314. Бибкод : 2019SciA....5.7314V . дои : 10.1126/sciadv.aau7314 . ПМК 6314821 . ПМИД 30613774 .
- ^ Кантор Дж.Р., Абу-Ремале М., Канарек Н., Фрейнкман Э., Гао Х., Луисен А. и др. (апрель 2017 г.). «Физиологическая среда изменяет клеточный метаболизм и обнаруживает мочевую кислоту как эндогенный ингибитор UMP-синтазы» . Клетка . 169 (2): 258–272.e17. дои : 10.1016/j.cell.2017.03.023 . ПМЦ 5421364 . ПМИД 28388410 .
- ^ «Мур против Регентов Калифорнийского университета (1990) 51 C3d 120» . Online.ceb.com . Проверено 27 января 2012 г.
- ^ Хейфлик Л. (сентябрь 1998 г.). «Краткая история смертности и бессмертия культивируемых клеток» . Медицинский журнал Кейо . 3. 47 (3): 174–182. дои : 10.2302/kjm.47.174 . ПМИД 9785764 .
- ^ «Справочник по тканям Уортингтона» . Проверено 30 апреля 2013 г.
- ^ Цянь Л., Зальцман В.М. (2004). «Улучшение экспансии и нейрональной дифференцировки мезенхимальных стволовых клеток посредством модификации поверхности культуры». Биоматериалы . 25 (7–8): 1331–1337. doi : 10.1016/j.bimaterials.2003.08.013 . ПМИД 14643607 .
- ^ Магуайр Дж. (май 2016 г.). «Терапия на основе взрослых стволовых клеток и кривая хайпа» . Письма ACS по медицинской химии . 7 (5): 441–443. doi : 10.1021/acsmedchemlett.6b00125 . ПМЦ 4867479 . ПМИД 27190588 .
- ↑ Перейти обратно: Перейти обратно: а б Прието Д., Апарисио Дж., Сотело-Сильвейра-младший (ноябрь 2017 г.). «Анализ клеточной миграции: недорогой лабораторный эксперимент для курсов клеточной биологии и биологии развития с использованием кератоцитов из чешуи рыбы» . Образование в области биохимии и молекулярной биологии . 45 (6): 475–482. дои : 10.1002/bmb.21071 . ПМИД 28627731 .
- ^ Дишер Д.Э., Джанми П., Ван Ю.Л. (ноябрь 2005 г.). «Тканевые клетки чувствуют жесткость своего субстрата и реагируют на него». Наука . 310 (5751): 1139–1143. Бибкод : 2005Sci...310.1139D . CiteSeerX 10.1.1.318.690 . дои : 10.1126/science.1116995 . ПМИД 16293750 . S2CID 9036803 .
- ^ Гилберт П.М., Хавенстрайт К.Л., Магнуссон К.Е., Сакко А., Леонарди Н.А., Крафт П. и др. (август 2010 г.). «Эластичность субстрата регулирует самообновление стволовых клеток скелетных мышц в культуре» . Наука . 329 (5995): 1078–1081. Бибкод : 2010Sci...329.1078G . дои : 10.1126/science.1191035 . ПМЦ 2929271 . ПМИД 20647425 .
- ^ Чоудхури Ф., Ли Ю., По Ю.К., Йокогама-Тамаки Т., Ван Н., Танака Т.С. (декабрь 2010 г.). Чжоу Цзы (ред.). «Мягкие субстраты способствуют гомогенному самообновлению эмбриональных стволовых клеток посредством подавления тяги клеточного матрикса» . ПЛОС ОДИН . 5 (12): е15655. Бибкод : 2010PLoSO...515655C . дои : 10.1371/journal.pone.0015655 . ПМК 3001487 . ПМИД 21179449 .
- ^ Энглер А.Дж., Сен С., Суини Х.Л., Дишер Д.Е. (август 2006 г.). «Эластичность матрицы определяет спецификацию происхождения стволовых клеток» . Клетка . 126 (4): 677–689. дои : 10.1016/j.cell.2006.06.044 . ПМИД 16923388 .
- ^ Пашек М.Дж., Захир Н., Джонсон К.Р., Лакинс Дж.Н., Розенберг Г.И., Гефен А. и др. (сентябрь 2005 г.). «Напряженный гомеостаз и злокачественный фенотип» . Раковая клетка . 8 (3): 241–254. дои : 10.1016/j.ccr.2005.08.010 . ПМИД 16169468 .
- ^ Левенталь К.Р., Ю Х., Касс Л., Лакинс Дж.Н., Эгеблад М., Эрлер Дж.Т. и др. (ноябрь 2009 г.). «Сшивание матрикса ускоряет прогрессирование опухоли за счет усиления передачи сигналов интегрина» . Клетка . 139 (5): 891–906. дои : 10.1016/j.cell.2009.10.027 . ПМК 2788004 . ПМИД 19931152 .
- ^ Тилгман Р.В., Коуэн Ч.Р., Мих Дж.Д., Корякина Ю., Джиоэли Д., Слэк-Дэвис Дж.К. и др. (сентябрь 2010 г.). Хотчин Н.А. (ред.). «Жесткость матрицы регулирует рост раковых клеток и клеточный фенотип» . ПЛОС ОДИН . 5 (9): е12905. Бибкод : 2010PLoSO...512905T . дои : 10.1371/journal.pone.0012905 . ПМЦ 2944843 . ПМИД 20886123 .
- ^ Лю Ф, Мих Дж.Д., Ши Б.С., Хо А.Т., Шариф А.С., Тагер А.М., Чумперлин DJ (август 2010 г.). «Усиление фиброза по обратной связи за счет повышения жесткости матрикса и подавления ЦОГ-2» . Журнал клеточной биологии . 190 (4): 693–706. дои : 10.1083/jcb.201004082 . ПМК 2928007 . ПМИД 20733059 .
- ^ Випфф П.Дж., Рифкин Д.Б., Мейстер Дж.Дж., Хинц Б. (декабрь 2007 г.). «Сокращение миофибробластов активирует латентный TGF-бета1 из внеклеточного матрикса» . Журнал клеточной биологии . 179 (6): 1311–1323. дои : 10.1083/jcb.200704042 . ПМК 2140013 . ПМИД 18086923 .
- ^ Джордж П.С., Хуэй Дж.Дж., Гомбос З., Маккормик М.Э., Ван А.Ю., Уэмура М. и др. (декабрь 2007 г.). «Повышенная жесткость печени крысы предшествует отложению матрикса: последствия фиброза». Американский журнал физиологии. Физиология желудочно-кишечного тракта и печени . 293 (6): G1147–G1154. дои : 10.1152/ajpgi.00032.2007 . ПМИД 17932231 . S2CID 201357 .
- ^ Ли Л., Шарма Н., Чиппада У., Цзян Х., Шлосс Р., Ярмуш М.Л., Ланграна Н.А. (май 2008 г.). «Функциональная модуляция клеток гепатоцитов, происходящих из ES, посредством изменения податливости субстрата». Анналы биомедицинской инженерии . 36 (5): 865–876. дои : 10.1007/s10439-008-9458-3 . ПМИД 18266108 . S2CID 21773886 .
- ^ Семлер Э.Дж., Ланчин П.А., Дасгупта А., Моге П.В. (февраль 2005 г.). «Инженерия гепатоцеллюлярного морфогенеза и функции с помощью лиганд-презентирующих гидрогелей с градуированной механической податливостью». Биотехнология и биоинженерия . 89 (3): 296–307. дои : 10.1002/бит.20328 . ПМИД 15744840 .
- ^ Фридланд Дж. К., Ли М. Х., Беттигер Д. (январь 2009 г.). «Механически активируемый переключатель интегрина контролирует функцию альфа5бета1». Наука . 323 (5914): 642–644. Бибкод : 2009Sci...323..642F . дои : 10.1126/science.1168441 . ПМИД 19179533 . S2CID 206517419 .
- ^ Чан CE, Odde DJ (декабрь 2008 г.). «Динамика тяги филоподий на податливых субстратах». Наука . 322 (5908): 1687–1691. Бибкод : 2008Sci...322.1687C . дои : 10.1126/science.1163595 . ПМИД 19074349 . S2CID 28568350 .
- ^ Дюпон С., Морсут Л., Арагона М., Энцо Э., Джулитти С., Корденонси М. и др. (июнь 2011 г.). «Роль ЯП/ТАЗ в механотрансдукции». Природа . 474 (7350): 179–183. дои : 10.1038/nature10137 . hdl : 11380/673649 . ПМИД 21654799 . S2CID 205225137 .
- ^ «наркотик [адрес электронной почты защищен] » . Nature.com . Проверено 26 марта 2013 г.
- ^ Дуэлл Б.Л., Криппс А.В., Шембри М.А., Улетт Г.К. (2011). «Модели кокультуры эпителиальных клеток для изучения инфекционных заболеваний: преимущества и ограничения» . Журнал биомедицины и биотехнологии . 2011 : 852419. doi : 10.1155/2011/852419 . ПМЦ 3189631 . ПМИД 22007147 .
- ^ Баррила Дж., Радтке А.Л., Краббе А., Саркер С.Ф., Хербст-Краловец М.М., Отт К.М., Никерсон Калифорния (ноябрь 2010 г.). «Органотипические трехмерные модели клеточных культур: использование сосуда с вращающимися стенками для изучения взаимодействия хозяина и патогена» . Обзоры природы. Микробиология . 8 (11): 791–801. дои : 10.1038/nrmicro2423 . ПМИД 20948552 . S2CID 6925183 .
- ^ Мапанао АК, Волиани В (июнь 2020 г.). «Трехмерные модели опухолей: содействие прорывам в трансляционных исследованиях нанотераностики». Прикладные материалы сегодня . 19 : 100552. doi : 10.1016/j.apmt.2019.100552 . S2CID 213634060 .
- ^ Кассано Д., Санти М., Д'Аутилиа Ф., Мапанао А.К., Луин С., Волиани В. (2019). «Фототермический эффект с помощью выделяемых сверхмалых в наноархитектурах, реагирующих на БИК-диапазон» . Горизонты материалов . 6 (3): 531–537. дои : 10.1039/C9MH00096H . hdl : 11384/77439 . ISSN 2051-6347 .
- ^ Эдмондсон Р., Бройль Дж.Дж., Адкок А.Ф., Ян Л. (май 2014 г.). «Трехмерные системы клеточных культур и их применение в разработке лекарств и клеточных биосенсорах» . Технологии анализа и разработки лекарств . 12 (4): 207–218. дои : 10.1089/adt.2014.573 . ПМК 4026212 . ПМИД 24831787 .
- ^ Бхаттачарья М., Малинен М.М., Лорен П., Лу Ю.Р., Куисма С.В., Каннинен Л. и др. (декабрь 2012 г.). «Гидрогель нанофибриллярной целлюлозы способствует созданию трехмерной культуры клеток печени» . Журнал контролируемого выпуска . 164 (3): 291–298. дои : 10.1016/j.jconrel.2012.06.039 . ПМИД 22776290 .
- ^ ДеРоса MC, Монреаль С, Шнитцер М, Уолш Р, Султан Ю (февраль 2010 г.). «Нанотехнологии в удобрениях» . Природные нанотехнологии . 5 (2): 91. Бибкод : 2010НатНа...5...91Д . дои : 10.1038/nnano.2010.2 . ПМИД 20130583 .
- ^ Сяо А.Ю., Тунг Ю.К., Цюй X, Патель Л.Р., Пиента К.Дж., Такаяма С. (май 2012 г.). «384 подвесных капельных массива обеспечивают превосходные Z-факторы и позволяют универсально формировать сфероиды для совместной культуры» . Биотехнология и биоинженерия . 109 (5): 1293–1304. дои : 10.1002/бит.24399 . ПМК 3306496 . ПМИД 22161651 .
- ^ Мапанао А.К., Санти М., Фарачи П., Каппелло В., Кассано Д., Волиани В. (сентябрь 2018 г.). «Эндогенно запускаемые сверхмалые в наноархитектуре: оценка нацеливания на трехмерные сфероиды карциномы поджелудочной железы» . АСУ Омега . 3 (9): 11796–11801. дои : 10.1021/acsomega.8b01719 . ПМК 6173554 . ПМИД 30320273 .
- ^ Гош С., Борш А., Гош С., Заволан М. (апрель 2021 г.). «Транскрипционный ландшафт линии клеток гепатомы, выращенной на каркасах белков внеклеточного матрикса» . БМК Геномика . 22 (1): 238. дои : 10.1186/s12864-021-07532-2 . ПМК 8025518 . ПМИД 33823809 .
- ^ Фонтура Х.К., Виззер К., Дос Сантос Ф.Г., Лигабуэ Р.А., Вайнлих Р., Пуга Р.Д. и др. (февраль 2020 г.). «Сравнение 2D и 3D моделей клеточных культур на предмет роста клеток, экспрессии генов и устойчивости к лекарствам». Материаловедение и инженерия. C. Материалы для биологических применений . 107 : 110264. doi : 10.1016/j.msec.2019.110264 . hdl : 10923/20413 . ПМИД 31761183 . S2CID 208277016 .
- ^ Хабанджар О, Диаб-Ассаф М, Калдефи-Чезет Ф, Делорт Л (ноябрь 2021 г.). «3D-системы культивирования клеток: применение в опухолях, преимущества и недостатки» . Международный журнал молекулярных наук . 22 (22): 12200. дои : 10.3390/ijms222212200 . ПМЦ 8618305 . ПМИД 34830082 .
- ^ Тиббит М.В., Ансет К.С. (июль 2009 г.). «Гидрогели как имитаторы внеклеточного матрикса для 3D-культуры клеток» . Биотехнология и биоинженерия . 103 (4): 655–663. дои : 10.1002/бит.22361 . ПМЦ 2997742 . ПМИД 19472329 .
- ^ «Создана вакцина от птичьего гриппа по-быстрому» . Проводной . 26 января 2006 г. Проверено 31 января 2010 г.
{{cite magazine}}: Неизвестный параметр|agency=игнорируется ( помогите ) - ^ Гао В., Солофф А.С., Лу Х, Монтекальво А., Нгуен Д.С., Мацуока Ю. и др. (февраль 2006 г.). «Защита мышей и домашней птицы от летального вируса птичьего гриппа H5N1 посредством иммунизации на основе аденовируса» . Журнал вирусологии . 80 (4): 1959–1964. doi : 10.1128/JVI.80.4.1959-1964.2006 . ПМЦ 1367171 . ПМИД 16439551 .
- ^ «NIAID использует Chiron для разработки вакцины против птичьего гриппа H9N2» . Национальный институт аллергии и инфекционных заболеваний (НИАИД) . 17 августа 2004 года . Проверено 31 января 2010 г.
- ^ Мики, Ясухиро; Оно, Кацухико; Хата, Сюко; Сузуки, Такаши; Кумамото, Хироюки; Сасано, Хиронобу (сентябрь 2012 г.). «Преимущества совместного культивирования перед культурой моноклеток при моделировании среды in vivo» . Журнал биохимии стероидов и молекулярной биологии . 131 (3–5): 68–75. дои : 10.1016/j.jsbmb.2011.12.004 . ISSN 0960-0760 . ПМИД 22265957 . S2CID 19646957 .
- ^ Пашос, Николаос К.; Браун, Венди Э.; Эшварамурти, Раджалакшманан; Ху, Джерри С.; Афанасиу, Кириакос А. (3 февраля 2014 г.). «Достижения в тканевой инженерии посредством совместной культуры стволовых клеток» . Журнал тканевой инженерии и регенеративной медицины . 9 (5): 488–503. дои : 10.1002/терм.1870 . ISSN 1932-6254 . ПМИД 24493315 . S2CID 1991776 .
- ^ Диттрих, Петра С.; Манц, Андреас (март 2006 г.). «Лаборатория на чипе: микрофлюидика в открытии лекарств» . Nature Reviews Открытие лекарств . 5 (3): 210–218. дои : 10.1038/nrd1985 . ISSN 1474-1784 . ПМИД 16518374 . S2CID 35904402 .
- ^ Террелл, Джон А.; Джонс, Кертис Г.; Кабандана, Гирасо Кеза Мония; Чен, Чэнпэн (2020). «От клеток на чипе к органам на чипе: каркасные материалы для трехмерной культуры клеток в микрофлюидике» . Журнал химии материалов Б. 8 (31): 6667–6685. дои : 10.1039/D0TB00718H . hdl : 11603/21825 . ПМИД 32567628 . S2CID 219972841 .
- ^ У, Цируй; Ван, Сяохун; У, Цзиньбо, Сяоли; Вэнь, Сюцин (12 февраля 2020 г.) . перспективы »
- ^ Люнг, Чак Мин; де Хаан, Пим; Рональдсон-Бушар, Кейси; Ким, Ге-А; Ко, Джихун; Ро, Хун Сок; Чен, Чжу; Хабибович, Памела; Чон, Ну Ли; Такаяма, Шуичи; Шулер, Майкл Л.; Вуньяк-Новакович, Гордана; Фрей, Оливье; Верпорте, Элизабет; То, И-Чин (12 мая 2022 г.). «Путеводитель по органу-на-чипе» . Учебники по методам Nature Reviews . 2 (1): 1–29. дои : 10.1038/s43586-022-00118-6 . ISSN 2662-8449 . S2CID 248756548 .
- ^ Ма, Чао; Пэн, Яньсун; Ли, Хунтун; Чен, Вэйцян (февраль 2021 г.). «Орган на чипе: новая парадигма разработки лекарств» . Тенденции в фармакологических науках . 42 (2): 119–133. дои : 10.1016/j.tips.2020.11.009 . ПМК 7990030 . ПМИД 33341248 .
- ^ Рапанан Дж.Л., Купер К.Е., Лейва К.Дж., Халл Э.Э. (август 2014 г.). «Коллективная миграция клеток первичных кератоцитов рыбок данио». Экспериментальные исследования клеток . 326 (1): 155–165. дои : 10.1016/j.yexcr.2014.06.011 . ПМИД 24973510 .
- ^ Ли Дж., Джейкобсон К. (ноябрь 1997 г.). «Состав и динамика адгезии клеток с субстратом в движущихся кератоцитах рыб» . Журнал клеточной науки . 110 (22): 2833–2844. дои : 10.1242/jcs.110.22.2833 . ПМИД 9427291 .
- ^ Другманд Дж.К., Шнайдер Ю.Дж., Агатос С.Н. (2012). «Клетки насекомых как фабрики по биопроизводству» . Достижения биотехнологии . 30 (5): 1140–1157. doi : 10.1016/j.biotechadv.2011.09.014 . ПМИД 21983546 .
- ^ Хант П., Робертсон Д., Вайс Д., Ренник Д., Ли Ф., Витте О.Н. (март 1987 г.). «Один тип стромальных клеток, полученных из костного мозга, поддерживает рост ранних лимфоидных и миелоидных клеток in vitro». Клетка . 48 (6): 997–1007. дои : 10.1016/0092-8674(87)90708-2 . ПМИД 2435412 . S2CID 31499611 .
- ^ ван ден Берг-Баккер К.А., Хагемейер А., Франкен-Постма Э.М., Смит В.Т., Куппен П.Дж., ван Равенсвай Клаасен Х.Х. и др. (февраль 1993 г.). «Создание и характеристика 7 клеточных линий карциномы яичника и одной клеточной линии гранулезной опухоли: особенности роста и цитогенетика». Международный журнал рака . 53 (4): 613–620. дои : 10.1002/ijc.2910530415 . ПМИД 8436435 . S2CID 6182244 .
- ^ Ли Ю.Г., Коренчук С., Лер Дж., Уитни С., Вессела Р., Пиента К.Дж. (2001). «Создание и характеристика новой линии клеток рака простаты человека: DuCaP». В Виво . 15 (2): 157–162. ПМИД 11317521 .
- ^ Оу Д., Митчелл Л.А., Декари Д., Тингл Эй.Дж., Непом Г.Т. (март 1998 г.). «Беспорядочное распознавание Т-клетками эпитопа капсидного белка краснухи, ограниченного молекулами DRB1 * 0403 и DRB1 * 0901, имеющими супертип HLA DR». Иммунология человека . 59 (3): 149–157. дои : 10.1016/S0198-8859(98)00006-8 . ПМИД 9548074 .
Дальнейшее чтение [ править ]
- Пейси Л., Стед С., Глив Дж., Томчик К., Деринг Л. (2006). «Культура нервных стволовых клеток: создание нейросфер, микроскопический анализ и криоконсервация» . Протокол обмена . дои : 10.1038/nprot.2006.215 .
- Гилаберт Х.А., Монтальво ГБ, Арталехо А.Р. (2006). «Первичные культуры хромаффинных клеток крысы: стандартизация и оценка качества анализов отдельных клеток» . Протокол обмена . дои : 10.1038/nprot.2006.294 .
- Лосардо Р.Х., Гутьеррес Р.К., Пратес Х.К., Московичи М., Торрес А.Р., Мартинес М.А. (2015). «Сергей Федоров: пионер регенерации нейронов. Дань уважения Панамериканской ассоциации анатомии» . Международный журнал морфологии . 33 (2): 794–800. дои : 10.4067/S0717-95022015000200059 .
- Маклауд Р.А., Диркс В.Г., Мацуо Ю., Кауфманн М., Милч Х., Дрекслер Х.Г. (ноябрь 1999 г.). «Широко распространенное внутривидовое перекрестное заражение линий опухолевых клеток человека, возникающее в источнике» . Международный журнал рака . 83 (4): 555–563. doi : 10.1002/(SICI)1097-0215(19991112)83:4<555::AID-IJC19>3.0.CO;2-2 . ПМИД 10508494 .
- Мастерс-младший (апрель 2002 г.). «Клетки HeLa 50 лет спустя: хорошие, плохие и ужасные». Обзоры природы. Рак . 2 (4): 315–319. дои : 10.1038/nrc775 . ПМИД 12001993 . S2CID 991019 .
- Витковский Ю.А. (июль 1983 г.). «Экспериментальная патология и истоки культуры тканей: вклад Лео Леба» . Медицинская история . 27 (3): 269–288. дои : 10.1017/S0025727300042964 . ПМЦ 1139336 . ПМИД 6353093 .
Внешние ссылки [ править ]
- Таблица распространенных клеточных линий из 4-го изд. Альберта.
- Раковые клетки в культуре
- Эволюция поверхностей клеточных культур
- Гипертекстовая версия базы данных клеточных линий
- Приложения для культивирования клеток — ресурсы, включая рекомендации по применению и протоколы для создания идеальной среды для выращивания клеток с самого начала.
- Основы клеточной культуры - Введение в клеточную культуру, охватывающее такие темы, как лабораторное оборудование, безопасность и асептические методы, включая основные протоколы клеточных культур и видеообучение.
- База данных «Кто есть кто в области клеточных культур и связанных с ними исследований»
- Хранилища ячеек Coriell
- Введение в клеточную культуру. Этот вебинар знакомит с историей, теорией, основными методами и потенциальными подводными камнями культуры клеток млекопитающих.
- Национальный центр клеточных наук (NCCS), Пуна, Индия; национальный репозиторий клеточных линий/гибридом и т. д.
- Общественное здравоохранение Англии , Коллекции культур общественного здравоохранения Англии (ECACC)

| Базы данных органов управления : Национальные |
|---|