Репликация вируса
Эта статья нуждается в дополнительных цитатах для проверки . ( апрель 2020 г. ) |
| гриппа Жизненный цикл вируса |
|---|
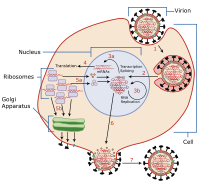 |
Репликация вируса — это образование биологических вирусов в процессе заражения в клетках-мишенях-хозяевах. Вирусы должны сначала проникнуть в клетку, прежде чем произойдет репликация вируса. Создавая большое количество копий своего генома и упаковывая эти копии, вирус продолжает заражать новых хозяев. Репликация между вирусами очень разнообразна и зависит от типа задействованных в них генов. Большинство ДНК-вирусов собираются в ядре, тогда как большинство РНК-вирусов развиваются исключительно в цитоплазме. [1]
Производство/репликация вируса
[ редактировать ]Вирусы размножаются только в живых клетках. Клетка - хозяин должнаобеспечить энергетическое и синтетическое оборудование, а также низко-молекулярные предшественники для синтеза вирусных белков и нуклеиновых кислот. [2]
Репликация вируса происходит в семь стадий:
- Вложение
- Вход (Проникновение)
- Удаление покрытия
- Репликация
- Сборка
- Созревание
- Освобождение (стадия освобождения).
Вложение
[ редактировать ]Это первый этап репликации вируса. Некоторые вирусы прикрепляются к клеточной мембране -хозяина клетки свою ДНК или РНК и вводят в хозяина , чтобы инициировать инфекцию. Прикрепление к клетке-хозяину часто достигается с помощью белка прикрепления вируса, который выходит из белковой оболочки , также известной как капсид вируса . Этот белок отвечает за связывание с поверхностным рецептором на плазматической мембране (или мембранными углеводами) клетки-хозяина. Вирусы могут использовать функции нормальных клеточных рецепторов, чтобы обеспечить прикрепление, имитируя молекулы, которые связываются с рецепторами клеток-хозяев. Например, риновирус использует свой белок прикрепления вируса для связывания с рецептором ICAM-1 на клетках-хозяевах, который обычно используется для облегчения адгезии между другими клетками-хозяевами. [3]
Вход
[ редактировать ]Вход, или проникновение, является вторым этапом репликации вируса. Этот этап характеризуется прохождением вируса через плазматическую мембрану клетки-хозяина. Наиболее распространенным способом проникновения вируса в клетку-хозяина является рецептор-опосредованный эндоцитоз , который не требует энергетических затрат для вируса, а только для клетки-хозяина. Рецепторно-опосредованный эндоцитоз возникает, когда молекула (в данном случае вирус) связывается с рецептором на мембране клетки. Ряд химических сигналов от этого связывания заставляет клетку обертывать прикрепленный вирус плазматической мембраной вокруг себя, образуя вируссодержащий везикулу внутри клетки. [3]
Вирусы проникают в клетки-хозяева, используя различные механизмы, включая эндоцитарные и неэндоцитарные пути. [4] Они также могут сливаться на плазматической мембране и распространяться внутри хозяина посредством слияния или слияния клеток. [5] Вирусы прикрепляются к белкам на поверхности клетки-хозяина, известным как клеточные рецепторы или факторы прикрепления, чтобы облегчить проникновение. [6] Имеющиеся данные показывают, что вирусы используют ионные каналы в клетках-хозяевах во время проникновения вируса. Слияние: внешние вирусные белки способствуют слиянию вириона с плазматической мембраной. [7] При этом в мембране хозяина образуется пора, и после проникновения вирион становится непокрытым, а его геномный материал затем переносится в цитоплазму. [8] Межклеточное слияние: некоторые вирусы вызывают экспрессию специфического белка на поверхности инфицированных клеток для привлечения неинфицированных клеток. [9] Это взаимодействие заставляет неинфицированную клетку сливаться с инфицированной клеткой при более низких уровнях pH с образованием многоядерной клетки, известной как синцитий. [10] Эндоцитарные пути: процесс, при котором внутриклеточный везикула образуется путем инвагинации мембраны, что приводит к поглощению внеклеточных и мембраносвязанных компонентов, в данном контексте вируса. [11] Неэндоцитарный путь: процесс, при котором вирусные частицы попадают в клетку путем слияния внеклеточной вирусной оболочки и мембраны клетки-хозяина. [4]
Удаление покрытия
[ редактировать ]Снятие покрытия — третий этап репликации вируса. Снятие покрытия определяется удалением белковой «оболочки» вириона и высвобождением его генетического материала. Этот этап происходит в той же области, где происходит транскрипция вируса. Различные вирусы имеют разные механизмы снятия покрытия. клетки-хозяина Некоторые РНК-вирусы, такие как риновирусы, используют низкий уровень pH в эндосомах для активации механизма снятия покрытия. Это включает в себя высвобождение риновирусом белка, который создает отверстия в эндосоме и позволяет вирусу высвободить свой геном через отверстия. Многие ДНК-вирусы перемещаются в ядро клетки-хозяина и высвобождают свой генетический материал через ядерные поры. [3]
Репликация
[ редактировать ]Четвертым этапом вирусного цикла является репликация, которая определяется быстрым производством вирусного генома. То, как вирус подвергается репликации, зависит от типа генетического материала, которым обладает вирус. Основываясь на своем генетическом материале, вирусы захватывают соответствующие клеточные механизмы для получения указанного генетического материала. Вирусы, содержащие двухцепочечную ДНК (дцДНК), имеют тот же генетический материал, что и все организмы, и поэтому могут использовать ферменты репликации в ядре клетки-хозяина для репликации вирусного генома. Многие РНК-вирусы обычно реплицируются в цитозоле клетки-хозяина и могут напрямую получать доступ к рибосомам для производства вирусных белков, когда РНК находится в репликативной форме.
Вирусы могут проходить два типа жизненных циклов: литический цикл и лизогенный цикл. В литическом цикле вирус внедряет свой геном в клетку-хозяина и инициирует репликацию, захватывая клеточный механизм хозяина для создания новых копий вируса. [12] В лизогенном жизненном цикле вирусный геном включается в геном хозяина. Геном хозяина будет проходить свой обычный жизненный цикл, реплицируя и делясь, реплицируя вирусный геном вместе со своим собственным. [13] Вирусный геном можно запустить для начала производства флаконов с помощью химических стимуляторов и стимуляторов окружающей среды. [14] Как только лизогенный вирус вступает в литический жизненный цикл, он продолжает действовать по путям продукции вируса и приступает к транскрипции/выработке мРНК. (например: герпес, вирус простого герпеса (ВПГ)-1, лизогенные бактериофаги и т. д.)
Сборка
[ редактировать ]Сборка — это когда вновь созданные вирусные белки и геномы собираются и соединяются вместе, образуя незрелые вирусы. Как и другие этапы, способ сборки конкретного вируса зависит от типа вируса. Сборка может происходить в плазматической мембране, цитозоле, ядре, аппарате Гольджи и других местах внутри клетки-хозяина. Некоторые вирусы вставляют свой геном в капсид только после того, как капсид завершен, в то время как у других вирусов волевой капсид окутывает геном во время его копирования. [2]
Созревание
[ редактировать ]Это последний шаг перед формированием компетентного вируса. Обычно это включает модификации капсида, которые обеспечиваются ферментами (закодированными хозяином или вирусом). [3]
Освобождение (стадия освобождения)
[ редактировать ]Последним этапом репликации вируса является высвобождение, когда вновь собранные и зрелые вирусы покидают клетку-хозяина. То, как вирус высвобождается из клетки-хозяина, зависит от типа вируса. Одним из распространенных типов выпуска является почкование. Это происходит, когда вирусы, образующие оболочку из плазматической мембраны хозяина, огибают мембрану капсида. Когда вирус изгибает плазматическую мембрану, он начинает охватывать весь капсид до тех пор, пока вирус больше не перестанет прикрепляться к клетке-хозяину. Другой распространенный способ, которым вирусы покидают клетку-хозяина, — это лизис клеток , при котором вирусы лизуют клетку, вызывая ее разрыв, в результате чего высвобождаются зрелые вирусы, находившиеся в клетке-хозяине. [3]
Балтиморская классификационная система
[ редактировать ]Вирусы подразделяются на 7 типов генов, каждый из которых имеет свои собственные семейства вирусов, которые, в свою очередь, сами имеют разные стратегии репликации. [15] Дэвид Балтимор , биолог, лауреат Нобелевской премии , разработал систему под названием Балтиморская классификационная система, позволяющая классифицировать различные вирусы на основе их уникальной стратегии репликации. На основе этой системы существует семь различных стратегий репликации (Балтиморский класс I, II, III, IV, V, VI, VII). Семь классов вирусов перечислены здесь кратко и в общих чертах. [16]
Класс 1: Вирусы с двухцепочечной ДНК.
[ редактировать ]Этот тип вируса обычно должен проникнуть в ядро хозяина , прежде чем он сможет размножаться. Некоторым из этих вирусов требуются полимеразы для репликации своего генома клеток-хозяев , в то время как другие, такие как аденовирусы или вирусы герпеса, кодируют свои собственные факторы репликации. Однако в любом случае репликация вирусного генома сильно зависит от клеточного состояния, допускающего репликацию ДНК, и, следовательно, от клеточного цикла . Вирус может заставить клетку подвергнуться принудительному делению , что может привести к трансформации клетки и, в конечном итоге, к раку . Примером семейства в этой классификации является Adenoviridae .
Есть только один хорошо изученный пример, когда семейство вирусов класса 1 не реплицируется внутри ядра. Это семейство поксвирусов , в которое входят высокопатогенные вирусы, поражающие позвоночных .
Класс 2: Вирусы с одноцепочечной ДНК.
[ редактировать ]Вирусы, подпадающие под эту категорию, включают вирусы, которые не так хорошо изучены, но все же имеют большое значение для позвоночных. Два примера включают Circoviridae и Parvoviridae . Они реплицируются внутри ядра и во время репликации образуют промежуточную двухцепочечную ДНК. Анелловирус человека, называемый TTV, включен в эту классификацию и обнаруживается почти у всех людей, бессимптомно поражая почти все основные органы .
РНК-вирусы:Полимераза РНК-вирусов лишена функций корректуры, присущих полимеразам ДНК-вирусов. Это способствовало тому, что РНК-вирусы имели более низкую точность репликации по сравнению с ДНК-вирусами, что привело к высокой мутагенности РНК-вирусов, что может увеличить их общую выживаемость. [17] РНК-вирусам не хватает способности идентифицировать и восстанавливать несовпадающие или поврежденные нуклеотиды, и, таким образом, РНК-геномы склонны к мутациям, вызванным механизмами, внутренними и внешними по отношению к репликации вируса. [18] РНК-вирусы представляют собой терапевтический обоюдоострый меч: РНК-вирусы могут противостоять воздействию противовирусных препаратов, вызывать эпидемии и заражать несколько видов хозяев из-за своей мутагенной природы, что затрудняет их лечение. Однако белок обратной транскриптазы, который часто поставляется с РНК-вирусом, может использоваться в качестве непрямой мишени для РНК-вирусов, предотвращая транскрипцию и синтез вирусных частиц. [19] (Это основа препаратов против СПИДа и ВИЧ. [20] )
Класс 3: Вирусы с двухцепочечной РНК.
[ редактировать ]Как и большинство вирусов с РНК- геномами, двухцепочечные РНК-вирусы не полагаются на полимеразы хозяина для репликации в такой степени, как вирусы с ДНК- геномами. Вирусы с двухцепочечной РНК изучены не так хорошо, как другие классы. Этот класс включает два основных семейства: Reoviridae и Birnaviridae . Репликация является моноцистронной и включает отдельные сегментированные геномы, а это означает, что каждый из генов кодирует только один белок, в отличие от других вирусов, которые демонстрируют более сложную трансляцию.
Классы 4 и 5: вирусы с одноцепочечной РНК.
[ редактировать ]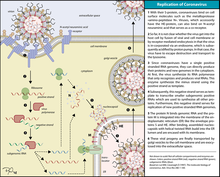
Эти вирусы состоят из двух типов, однако оба имеют тот факт, что репликация происходит преимущественно в цитоплазме и что репликация не так зависит от клеточного цикла, как репликация ДНК-вирусов. Этот класс вирусов также является одним из наиболее изученных типов вирусов наряду с вирусами с двухцепочечной ДНК.
Класс 4: Вирусы с одноцепочечной РНК – положительный смысл.
[ редактировать ]РНК-вирусы с положительным смыслом и, по сути, все гены, определенные как позитивные, могут быть напрямую доступны рибосомам хозяина для немедленного образования белков. Их можно разделить на две группы, обе из которых реплицируются в цитоплазме:
- Вирусы с полицистронной мРНК , где геномная РНК образует мРНК и транслируется в полипротеиновый продукт, который впоследствии расщепляется с образованием зрелых белков. Это означает, что ген может использовать несколько методов для производства белков из одной и той же цепи РНК, уменьшая размер своего генома.
- Вирусы со сложной транскрипцией, для которых могут быть использованы субгеномные мРНК, рибосомальный сдвиг рамки и протеолитический процессинг полипротеинов. Все это разные механизмы производства белков из одной и той же цепи РНК.
Примеры этого класса включают семейства Coronaviridae , Flaviviridae и Picornaviridae .
Класс 5: Вирусы с одноцепочечной РНК – отрицательный смысл.
[ редактировать ]РНК-вирусы с отрицательным смыслом, а также все гены, определенные как отрицательные, не могут быть напрямую доступны рибосомам хозяина для немедленного образования белков. Вместо этого они должны транскрибироваться вирусными полимеразами в «читаемый» дополнительный положительный смысл. Их также можно разделить на две группы:
- Вирусы, содержащие несегментированные геномы , для которых первым этапом репликации является транскрипция из генома с отрицательной цепью с помощью вирусной РНК-зависимой РНК-полимеразы с образованием моноцистронных мРНК, кодирующих различные вирусные белки. Затем создается копия генома с положительным смыслом, которая служит матрицей для производства генома с отрицательной цепью. Репликация происходит внутри цитоплазмы.
- Вирусы с сегментированными геномами, репликация которых происходит в цитоплазме и у которых вирусная РНК-зависимая РНК-полимераза продуцирует моноцистронные мРНК из каждого сегмента генома.
Примеры этого класса включают семейства Orthomyxoviridae , Paramyxoviridae , Bunyaviridae , Filoviridae и Rhabdoviridae (включая бешенство ).
Класс 6: Вирусы с одноцепочечной РНК с положительным смыслом, которые реплицируются через промежуточную ДНК.
[ редактировать ]Хорошо изученное семейство вирусов этого класса включает ретровирусы . Одной из определяющих особенностей является использование обратной транскриптазы для преобразования РНК с положительным смыслом в ДНК. Вместо того, чтобы использовать РНК в качестве шаблонов белков, они используют ДНК для создания шаблонов, которые встраиваются в геном хозяина с помощью интегразы . Затем репликация может начаться с помощью полимераз клетки-хозяина.
Класс 7: Вирусы с двухцепочечной ДНК, которые реплицируются через промежуточную одноцепочечную РНК.
[ редактировать ]Эта небольшая группа вирусов, примером которой является вирус гепатита B , имеет двухцепочечный геном с пробелами, который впоследствии заполняется с образованием ковалентно замкнутого круга ( cccDNA ), который служит матрицей для производства вирусных мРНК и субгеномной РНК. Прегеномная РНК служит матрицей для обратной транскриптазы вируса и для производства генома ДНК.
Ссылки
[ редактировать ]- ^ Робертс, Р.Дж. (2001). Патология рыб (3-е изд.). Elsevier Науки о здоровье.
- ↑ Перейти обратно: Перейти обратно: а б Брукс, Джорджия; Кэрролл, Карен С.; Бутель, Джанет; Морс, Стивен (21 декабря 2012 г.). Медицинская микробиология Джавеца Мельника и Адельбергса (26-е изд.). МакГроу Хилл Профессионал. ISBN 978-0-07-181578-9 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Лаутен, Дженнифер (2016). «Репликация вируса». Основная вирусология человека . Эльзевир. стр. 49–70. дои : 10.1016/b978-0-12-800947-5.00004-1 . ISBN 978-0-12-800947-5 . ПМК 7149683 .
- ↑ Перейти обратно: Перейти обратно: а б Димитров, Димитер С. (февраль 2004 г.). «Вхождение вируса: молекулярные механизмы и биомедицинские применения» . Обзоры природы Микробиология . 2 (2): 109–122. дои : 10.1038/nrmicro817 . ПМК 7097642 . ПМИД 15043007 .
- ^ Собхи, Хайтам (2017). «Сравнительный обзор проникновения и прикрепления вируса во время крупных и гигантских вирусных инфекций, вызываемых дцДНК» . Архив вирусологии . 162 (12): 3567–3585. дои : 10.1007/s00705-017-3497-8 . ПМК 5671522 . ПМИД 28866775 .
- ^ Марш, М. (2006). «Запись вируса: открой кунжут» . Клетка . 124 (4). Cell Press: 729–740. дои : 10.1016/j.cell.2006.02.007 . ПМК 7112260 . ПМИД 16497584 .
- ^ Гроув, Джо (2011). «Клеточная биология рецептор-опосредованного проникновения вируса» . Журнал клеточной биологии . 195 (7). Журнал клеточной биологии: 1071–1082. дои : 10.1083/jcb.201108131 . ПМК 3246895 . ПМИД 22123832 .
- ^ Килиан, Маргарет (2006). «Белки, слитые с мембраной вируса: несколько способов сделать шпильку» . Обзоры природы Микробиология . 4 (1). Природа: 67–76. дои : 10.1038/nrmicro1326 . ПМК 7097298 . ПМИД 16357862 .
- ^ Саттентау, Квентин (2008). «Избегая пустоты: распространение вирусов человека от клетки к клетке» . Обзоры природы Микробиология . 6 (11): 815–826. дои : 10.1038/nrmicro1972 . ПМИД 18923409 .
- ^ Чжун, Пэн (февраль 2013 г.). «Передача вирусов от клетки к клетке» . Современное мнение в вирусологии . Занесение вируса / Экологическая вирусология. 3 (1): 44–50. дои : 10.1016/j.coviro.2012.11.004 . ПМЦ 3587356 . ПМИД 23219376 .
- ^ Барроу, Эрик (2013). «Многомасштабные перспективы проникновения вируса посредством эндоцитоза» . Вирусологический журнал . 10 :177. дои : 10.1186/1743-422X-10-177 . ПМЦ 3679726 . ПМИД 23734580 .
- ^ Кэмпбелл, Аллан (2003). «Будущее биологии бактериофагов» . Обзоры природы Генетика . 4 (6): 471–477. дои : 10.1038/nrg1089 . ПМЦ 7097159 . ПМИД 12776216 .
- ^ Кристина, Ховард-Варона (2017). «Лизогения в природе: механизмы, влияние и экология умеренных фагов» . Журнал ISME . 11 (7): 1511–1520. Бибкод : 2017ISMEJ..11.1511H . дои : 10.1038/ismej.2017.16 . ПМК 5520141 . ПМИД 28291233 .
- ^ Чжан, Мэнхуэй (2022 г.). «Переходы жизненного цикла фагов умеренного климата: регулирующие факторы и потенциальные экологические последствия» . Вирусы . 14 (9). MDPI: 1904. doi : 10.3390/v14091904 . ПМЦ 9502458 . ПМИД 36146712 .
- ^ Гирдхар, Кхьяти; Поуис, Амайя; Райсингани, Амол; Хрудинова, Мартина; Хуан, Жуйсюй; Тран, Ту; Севги, Каан; Догус Догру, Юсуф; Алтиндис, Эмра (29 сентября 2021 г.). «Вирусы и метаболизм: влияние вирусных инфекций и вирусных инсулинов на метаболизм хозяина» . Ежегодный обзор вирусологии . 8 (1): 373–391. doi : 10.1146/annurev-virology-091919-102416 . ISSN 2327-056X . ПМЦ 9175272 . ПМИД 34586876 .
- ^ Диммок, Найджел; и др. (2007). Введение в современную вирусологию (6-е изд.). Издательство Блэквелл.
- ^ Хофер, Урсула (2013). «Обмануть машину корректуры коронавируса» . Обзоры природы Микробиология . 11 (10): 662–663. дои : 10.1038/nrmicro3125 . ПМК 7097040 . ПМИД 24018385 .
- ^ Эверетт, Клинтон Смит (2017). «Не такая уж и бесконечная податливость РНК-вирусов: вирусные и клеточные детерминанты скорости мутаций РНК-вирусов» . ПЛОС Патогены . 13 (4): e1006254. doi : 10.1371/journal.ppat.1006254 . ПМЦ 5407569 . ПМИД 28448634 .
- ^ Барр, Дж. Н. (2016). «Генетическая нестабильность РНК-вирусов» . Стабильность генома . Эльзевир: 21–35. дои : 10.1016/B978-0-12-803309-8.00002-1 . ISBN 978-0-12-803309-8 . ПМК 7149711 .
- ^ «Лечение ВИЧ: основы» . hivinfo.nih.gov . НАЦИОНАЛЬНЫЕ ИНСТИТУТЫ ЗДРАВООХРАНЕНИЯ США.

| Базы данных органов управления : Национальные |
|---|