Передача белковых сигналов в развитии сердца
Сердце — первый функциональный орган эмбриона позвоночного. Существует 5 стадий развития сердца .
Стадии развития сердца
[ редактировать ]Инициация
[ редактировать ]
Спецификация сердечных клеток-предшественников: Мезодерма латеральной пластинки расслаивается с образованием двух слоев: дорсальной соматической (париетальной) мезодермы и вентральной висцеральной мезодермы. Клетки-предшественники сердца происходят из двух областей висцеральной мезодермы, называемых кардиогенной мезодермой . Эти клетки могут дифференцироваться в эндокард , выстилающий камеры сердца и клапаны, и миокард , формирующий мускулатуру желудочков и предсердий.
Клетки сердца специфицированы в передней мезодерме такими белками, как Dickkopf-родственный белок 1 , гомолог Nodal и Cerberus , секретируемый передней энтодермой . Действуют ли Dickkopf-1 и Nodal непосредственно на сердечную мезодерму, является предметом исследования, но кажется, что они действуют, по крайней мере, опосредованно, стимулируя продукцию дополнительных факторов из передней энтодермы. Эти ранние сигналы необходимы для формирования сердца, так что удаление передней эндодермы блокирует формирование сердца. Передней энтодермы также достаточно для стимуляции дифференцировки сердца, поскольку она может индуцировать некардиогенную мезодерму из более задних положений эмбриона с образованием сердца.
Секреция ингибиторов Wnt (таких как Cerberus, Dickkopf и Crescent) передней энтодермой также предотвращает ингибирование формирования сердца Wnt3a и Wnt8, секретируемых нервной трубкой. Хорда секретирует антагонисты BMP (Chordin и Noggin), чтобы предотвратить образование кардиальной мезодермы в неподходящих местах.
Другие кардиогенные сигналы, такие как BMP и FGF, активируют экспрессию сердечных специфических транскрипционных факторов, таких как гомеодоменный белок Nkx2.5 . Nkx2.5 активирует ряд нижестоящих факторов транскрипции (таких как MEF2 и GATA), которые активируют экспрессию специфических белков сердечной мышцы. Мутации в Nkx2.5 приводят к дефектам развития сердца и врожденным порокам сердца.
Шаг 1: Формирование трубочек
[ редактировать ]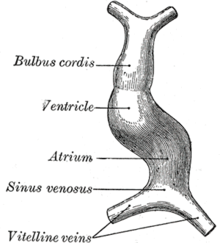
Миграция сердечных клеток-предшественников и слияние зачатков: сердечные клетки-предшественники мигрируют вперед по направлению к средней линии и сливаются в единую сердечную трубку. Фибронектин во внеклеточном матриксе направляет эту миграцию. Если это событие миграции блокируется, возникает расщелина кардии , при которой два зачатка сердца остаются разделенными. Во время слияния сердечная трубка располагается вдоль передней/задней оси для различных областей и камер сердца.
Окружающий мезокард дегенерирует, оставляя примитивное сердце прикрепленным только артериальными и венозными концами, которые анатомически прикреплены к глоточным дугам и поперечной перегородке соответственно. Развивающееся трубчатое сердце затем складывается вентрально и выпячивается в пяти областях по своей длине: первой, ближайшей к артериальному концу, является артериальный ствол, за ней следуют луковица сердца, примитивный желудочек, примитивное предсердие и венозный синус. Все пять эмбриональных расширений примитивного сердца развиваются во взрослые структуры сердца.
Шаг 2: Цикл
[ редактировать ]
Сердечная трубка изгибается вправо, меняя полярность с передней/задней на левую/правую. Подробный механизм неизвестен, однако для формирования петли необходим асимметрично локализованный транскрипционный фактор Pitx2 . Направление асимметрии устанавливается гораздо раньше во время эмбрионального развития, возможно, за счет вращения ресничек по часовой стрелке , и приводит к односторонней экспрессии Pitx2. Зацикливание также зависит от специфичных для сердца белков, активируемых Nkx2.5, таких как Hand1 , Hand2 и Xin.

Формирование камер сердца. Судьба клеток камер сердца охарактеризована до формирования петли сердца, но не может быть различена до момента образования петли. Hand1 локализован в левом желудочке, тогда как Hand2 локализован в правом желудочке.
Шаг 3: Формирование перегородки
[ редактировать ]Правильное расположение и функция клапанов имеют решающее значение для формирования камер и правильного кровотока. До этого времени эндокардиальная подушка служит импровизированным клапаном.
Шаг 3(а): Межпредсердная перегородка
[ редактировать ]Примитивный атриум разделяется на две части путем слияния нескольких структур. От крыши первичного предсердия спускается первичная перегородка , которая растет к эндокардиальным подушкам внутри предсердного канала . Прямо перед слиянием первичной перегородки с эндокардиальными подушками имеется временное пространство, называемое первичным отверстием. Как только они сливаются, в середине первичной перегородки образуется новое отверстие, называемое вторым отверстием или вторичным отверстием . Справа от первичной перегородки, также спускаясь с крыши первичного предсердия , спускается перегородка полулунной формы, называемая перегородкой вторичной . Свободные края вторичной перегородки образуют отверстие, называемое овальным окном , которое закрывается после рождения, когда первичная и вторичная перегородки сливаются друг с другом, завершая формирование межпредсердной перегородки .
Предсердный канал в свою очередь разделен на правую и левую стороны предсердно-желудочковой перегородкой, которая берет начало от слияния дорсальной и вентральной эндокардиальной подушки . Правая сторона предсердного канала станет трехстворчатым клапаном , а левая — двустворчатым клапаном .
Дефекты формирования АВ-перегородки приводят к дефектам атриовентрикулярной перегородки , включая персистирующий АВ-канал и атрезию трикуспидального клапана .
Шаг 3(б): Межжелудочковая перегородка
[ редактировать ]Дно на средней линии примитивного желудочка образует межжелудочковую перегородку , разделяющую камеру на две части. IV перегородка растет вверх по направлению к эндокардиальной подушке. По мере его роста появляется отверстие — межжелудочковое отверстие , которое в дальнейшем закрывается немышечной IV перегородкой.
Дефекты формирования IV перегородки вызывают дефекты межжелудочковой перегородки , которые сообщают оба желудочка.
Шаг 4: Перегородка выводного тракта
[ редактировать ]Артериальный ствол и прилежащая луковица сердца разделяются клетками нервного гребня . [1] Как только клетки туловищного гребня встречаются с клетками бульбарного гребня, они скручиваются друг вокруг друга в спиральной ориентации, сливаются и образуют аорто-легочную перегородку . [2] На этом закончится отделение аорты от легочного ствола . [3]
Дефекты этого процесса известны как дефект аортолегочной перегородки и вызывают стойкий артериальный ствол , неравномерное деление артериального ствола, транспозицию магистральных артерий , стеноз аортального и легочного клапанов или тетраду фалло .
Шаг 5: Формирование сердечного клапана
[ редактировать ]сердечные клапаны Формируются .
Нарушения этого процесса известны как пороки клапанов сердца .
Ссылки
[ редактировать ]- ^ Машхофф К.Л., Болдуин Х.С. (2000). «Молекулярные детерминанты миграции нервного гребня». Являюсь. Дж. Мед. Жене . 97 (4): 280–8. doi : 10.1002/1096-8628(200024)97:4<280::AID-AJMG1278>3.0.CO;2-N . ПМИД 11376439 .
- ^ Кирби М.Л., Гейл Т.Ф., Стюарт Д.Е. (1983). «Клетки нервного гребня способствуют нормальной аорто-легочной перегородке». Наука . 220 (4061): 1059–61. дои : 10.1126/science.6844926 . ПМИД 6844926 .
- ^ Цзян X, Рович Д.Х., Сориано П., МакМахон А.П., Суков Х.М. (2000). «Судьба сердечного нервного гребня млекопитающих...». Разработка . 127 (8). Кембридж, Англия: 1607–1616 гг. ПМИД 10725237 .