Селективный модулятор рецепторов эстрогена
| Селективный модулятор рецепторов эстрогена | |
|---|---|
| Класс препарата | |
 Тамоксифен – нестероидный трифенилэтиленовый антиэстроген, широко используемый препарат при лечении рака молочной железы . | |
| Идентификаторы классов | |
| Синонимы | СЭРМ; агонист/антагонист эстрогеновых рецепторов; ЭРАА |
| Использовать | Рак молочной железы , бесплодие , остеопороз , атрофия влагалища , диспареуния , контрацепция , мужской гипогонадизм , гинекомастия , боль в груди и др. |
| код АТС | G03XC |
| Биологическая цель | Рецептор эстрогена |
| Юридический статус | |
| В Викиданных | |
Селективные модуляторы рецепторов эстрогена ( SERM ), также известные как агонисты/антагонисты рецепторов эстрогена ( ERAA ), [1] [2] представляют собой класс препаратов , действующих на рецепторы эстрогена (ЭР). [3] Характерной чертой, отличающей эти вещества от чистых агонистов - антагонистов ЭР (то есть полных агонистов и молчащих антагонистов ), является то, что их действие различно в различных тканях, что позволяет им избирательно ингибировать или стимулировать эстрогеноподобное действие в различных тканях.
Медицинское использование
[ редактировать ]СЭРМ используются при различных заболеваниях, связанных с эстрогеном, включая лечение овуляторной дисфункции при лечении бесплодия , лечении и профилактике постменопаузального остеопороза , лечении и снижении риска рака молочной железы. [4] и лечение диспареунии вследствие менопаузы. СЭРМ также используется в сочетании с конъюгированными эстрогенами, показанными для лечения симптомов дефицита эстрогенов и вазомоторных симптомов, связанных с менопаузой. [5]

Примеры
[ редактировать ]Тамоксифен является гормональным препаратом первой линии при ER-положительном метастатическом раке молочной железы. Он используется для снижения риска рака молочной железы у женщин из группы высокого риска, а также в качестве адъювантного лечения протоковой узлам с отрицательным и положительным по подмышечным карциномы in situ . [5] [6] Лечение тамоксифеном также полезно для снижения плотности костей и уровня липидов в крови у женщин в постменопаузе. Побочные эффекты включают приливы жара и увеличение относительного риска развития рака эндометрия по сравнению с женщинами соответствующего возраста. [6] [4]
Торемифен , хлорированное производное тамоксифена, вызывает меньшее количество аддуктов ДНК в печени, чем тамоксифен в доклинических исследованиях , и был разработан для предотвращения рака печени. Он используется в качестве эндокринной терапии для женщин с ER/PR-позитивной стадией 4 или рецидивирующим метастатическим раком молочной железы. [7] и продемонстрировал аналогичную эффективность по сравнению с тамоксифеном в качестве адъювантного лечения рака молочной железы и лечения метастатического рака молочной железы. [6]
Ралоксифен используется для профилактики и лечения постменопаузального остеопороза и профилактики рака молочной железы у женщин в постменопаузе высокого риска с остеопорозом. [5] Доклинические и клинические отчеты показывают, что он значительно менее эффективен, чем эстроген, при лечении остеопороза. Он связан с приемлемым профилем эндометрия и не оказывает тамоксифеноподобного воздействия на матку, но связан с такими побочными эффектами, как венозная тромбоэмболия и вазомоторные симптомы, включая приливы. [4]
Оспемифен является аналогичным метаболитом торемифена. В отличие от тамоксифена, торемифен не является гепатоканцерогеном для крыс , и поэтому оспемифен также может быть более безопасным СЭРМ, чем тамоксифен. [4] Он используется для лечения умеренной и тяжелой диспареунии, симптома атрофии вульвы и влагалища, связанного с менопаузой. Клинические данные о раке молочной железы отсутствуют, но данные как in vitro , так и in vivo позволяют предположить, что оспемифен может обладать химиопрофилактической активностью в тканях молочной железы. [6]
Базедоксифен используется для лечения остеопороза у женщин в постменопаузе с повышенным риском переломов. Было показано, что он относительно безопасен и хорошо переносится. Он не показывает стимуляции молочной железы или эндометрия, и в первые два года небольшое увеличение лучше при венозной тромбоэмболии и в долгосрочной перспективе аналогично другим СЭРМ. Преимущество базедоксифена перед ралоксифеном заключается в том, что он повышает активность эндотелиальной синтазы оксида азота и не противодействует влиянию 17β-эстрадиола на вазомоторные симптомы. [5]
Первый тканеселективный эстрогеновый комплекс (TSEC) объединяет конъюгированные эстрогены и базедоксифен SERM для смешивания их активности. Комбинированная терапия используется для лечения умеренных и тяжелых вазомоторных симптомов, связанных с менопаузой, профилактики постменопаузального остеопороза, а также лечения симптомов дефицита эстрогенов у женщин в постменопаузе, не подвергшихся матке. Комбинация позволяет использовать преимущества эстрогена в отношении облегчения вазомоторных симптомов без эстрогенной стимуляции эндометрия. [5] [6]
СЭРМ также использовались некоторыми трансгендерами в качестве варианта гормонального лечения. [8]
Доступные формы
[ редактировать ]| Имя | Название бренда | Разрешенное использование | Запуск | Примечания |
|---|---|---|---|---|
| Анордрин | Цзы Юн | Экстренная контрацепция | 1970-е годы | Только в Китае в сочетании с мифепристоном. |
| Базедоксифен | Дуави | остеопороза Профилактика | 2013 | В сочетании с конъюгированными эстрогенами |
| Бропарэстрол | Акнестрол | Дерматология ; рака молочной железы Лечение | 1970-е годы | Снято с производства |
| Кломифен | Кломид | Женское бесплодие | 1967 | |
| Циклофенил | Сексовид | Женское бесплодие; Симптомы менопаузы | 1970 | В основном снято с производства |
| Лазофоксифен | Фаблин | Профилактика, лечение остеопороза; Вагинальная атрофия | 2009 | Только в Литве и Португалии. |
| Ормелоксифен | Сахели | Гормональная контрацепция | 1991 | Только в Индии |
| Оспемифен | Осфена | Диспареуния вследствие атрофии влагалища | 2013 | |
| Ралоксифен | Эвиста | Профилактика, лечение остеопороза; Профилактика рака молочной железы | 1997 | |
| Тамоксифен | Нольвадекс | Лечение рака молочной железы | 1978 | |
| Торремифен | Фарестон | Лечение рака молочной железы | 1997 | |
| Источники: см. отдельные статьи. | ||||
Фармакология
[ редактировать ]Фармакодинамика
[ редактировать ]SERM являются конкурентными частичными агонистами ER. [9] Различные ткани имеют разную степень чувствительности к активности эндогенных эстрогенов, поэтому СЭРМ оказывают эстрогенное или антиэстрогенное действие в зависимости от конкретной ткани, о которой идет речь, а также от процента внутренней активности (IA) СЭРМ. [10] Примером СЭРМ с высоким IA и, следовательно, главным образом с эстрогенным действием, является хлортрианизен , тогда как примером СЭРМ с низким IA и, следовательно, с преимущественно антиэстрогенным действием, является этамокситрифетол . СЭРМ, такие как кломифен и тамоксифен, находятся сравнительно ближе к середине по своей IA и балансу эстрогенной и антиэстрогенной активности. Ралоксифен – это СЭРМ, обладающий более сильным антиэстрогенным действием, чем тамоксифен; оба являются эстрогенными для костей, но ралоксифен является антиэстрогенным в матке , а тамоксифен - эстрогенным в этой части тела. [10]
Связывающий сайт
[ редактировать ]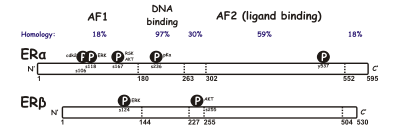
СЭРМ действуют на рецептор эстрогена (ER), который является внутриклеточным лиганд-зависимым активатором транскрипции и принадлежит к семейству ядерных рецепторов . [11] Были идентифицированы два разных подтипа ER: ERα и ERβ . ERα считается основной средой, в которой сигналы эстрогена передаются на уровне транскрипции, и является преобладающим ER в женских репродуктивных путях и молочных железах, тогда как ERβ находится преимущественно в сосудистых эндотелиальных клетках , костях и ткани мужской простаты. [12] Известно, что концентрации ERα и ERβ различаются в тканях в процессе развития, старения или болезненного состояния. [13] Многие характеристики этих двух типов схожи, например, размер (~ 600 и 530 аминокислот) и структура. ERα и ERβ имеют примерно 97% идентичности аминокислотных последовательностей в ДНК-связывающем домене и около 56% в лигандсвязывающем домене . [11] [13] Основное отличие лиганд-связывающих доменов определяется Leu -384 и Met -421 в ERα, которые в ERβ заменены на Met-336 и Ile -373 соответственно. [14] Вариация больше на N-конце между ERα и ERβ. [15]
ДНК-связывающий домен состоит из двух субдоменов . Один имеет проксимальный блок, который участвует в распознавании ДНК, а другой содержит дистальный блок, отвечающий за ДНК-зависимую димеризацию ДНК-связывающего домена . Последовательность проксимального бокса идентична у ERα и ERβ, что указывает на сходную специфичность и близость между двумя подгруппами. Глобулярные белки ДНК-связывающего домена содержат восемь цистеинов и обеспечивают тетраэдрическую координацию двух ионов цинка . Эта координация делает возможным связывание ER с элементами ответа на эстроген. [12] Лигандсвязывающий домен представляет собой глобулярную трехслойную структуру, состоящую из 11 спиралей и содержащую карман для природного или синтетического лиганда. [12] [11] Факторами, влияющими на аффинность связывания, являются главным образом наличие фенольного фрагмента, размер и форма молекулы, двойные связи и гидрофобность . [16]
Дифференциальное расположение спирали 12 активирующей функции 2 (AF-2) в лигандсвязывающем домене связанным лигандом определяет, оказывает ли лиганд агонистическое или антагонистическое действие. В рецепторах, связанных с агонистом, спираль 12 расположена рядом со спиралями 3 и 5. Спирали 3, 5 и 12 вместе образуют поверхность связывания для мотива NR-бокса, содержащегося в коактиваторах с канонической последовательностью LXXLL (где L представляет собой лейцин или изолейцин и X представляет собой любую аминокислоту). Несвязанные (апо) рецепторы или рецепторы, связанные с лигандами-антагонистами, отворачивают спираль 12 от поверхности связывания LXXLL, что приводит к преимущественному связыванию более длинного, богатого лейцином мотива, LXXXIXXX(I/L), присутствующего на корепрессорах NCoR1 или SMRT. Кроме того, некоторые кофакторы связываются с ER через окончания, сайт связывания ДНК или другие сайты связывания. Таким образом, одно соединение может быть агонистом ER в ткани, богатой коактиваторами , но антагонистом ER в тканях, богатых корепрессорами . [11]
Механизм действия
[ редактировать ]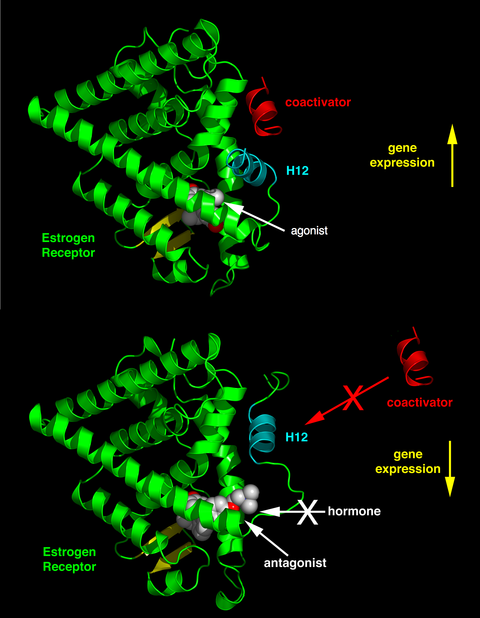
Эстрогенные соединения охватывают спектр активности:
- Полные агонисты (агонисты всех тканей), такие как природный эндогенный гормон эстрадиол.
- Смешанные агонисты/антагонисты (агонисты в одних тканях и антагонисты в других), такие как тамоксифен (СЭРМ).
- Чистые антагонисты (антагонисты во всех тканях), такие как фулвестрант .
Известно, что SERM стимулируют эстрогенное действие в таких тканях, как печень, кости и сердечно-сосудистая система, но известно, что они блокируют действие эстрогена там, где стимуляция нежелательна, например, в груди и матке. [19] Эта агонистическая или антагонистическая активность вызывает различные структурные изменения рецепторов, что приводит к активации или репрессии генов-мишеней эстрогена. [3] [19] [4] [20] SERM взаимодействуют с рецепторами путем диффузии в клетки и их связывания с субъединицами ERα или ERβ, что приводит к димеризации и структурным изменениям рецепторов. Это облегчает взаимодействие SERM с элементами ответа на эстроген, что приводит к активации эстроген-индуцируемых генов и опосредованию эффектов эстрогена. [19]
Уникальной особенностью SERM является их ткане- и клеточно-селективная активность. Появляется все больше доказательств того, что активность SERM в основном определяется избирательным привлечением корепрессоров и коактиваторов к генам-мишеням ER в определенных типах тканей и клеток. [4] [20] [21] SERM могут влиять на стабильность коактиваторного белка, а также регулировать активность коактиватора посредством посттрансляционных модификаций, таких как фосфорилирование . Множественные сигнальные пути роста, такие как HER2 , PKC , PI3K и другие, подавляются в ответ на лечение антиэстрогенами. Коактиватор стероидных рецепторов 3 (SRC-3) фосфорилируется активированными киназами , которые также усиливают его коактиваторную активность, влияют на рост клеток и в конечном итоге способствуют устойчивости к лекарствам. [21]
Соотношение ERα и ERβ в целевом сайте может быть еще одним способом определения активности SERM. Высокие уровни клеточной пролиферации хорошо коррелируют с высоким соотношением ERα:ERβ, но репрессия клеточной пролиферации коррелирует с доминированием ERβ над ERα. Соотношение ЭР в неопластической и нормальной тканях молочной железы может иметь важное значение при рассмотрении химиопрофилактики с помощью СЭРМ. [3] [19] [4] [20]
При рассмотрении различий между ERα и ERβ важны активирующая функция 1 (AF-1) и AF-2. Вместе они играют важную роль во взаимодействии с другими корегуляторными белками, которые контролируют транскрипцию генов . [19] [4] AF-1 расположен на аминоконце ER и только на 20% гомологичен ERα и ERβ. С другой стороны, AF-2 очень похож в ERα и ERβ, и отличается только одна аминокислота. [4] Исследования показали, что при переключении областей AF-1 в ERα и ERβ возникают специфические различия в транскрипционной активности. Как правило, SERM могут частично активировать сконструированные гены через ERα с помощью рецепторного элемента эстрогена, но не через ERβ. [19] [4] [20] Хотя ралоксифен и активная форма тамоксифена могут стимулировать репортерные гены, регулируемые AF-1, как в ERα, так и в ERβ. [4]
Открытие двух подтипов ER привело к синтезу ряда рецептор-специфичных лигандов, которые могут включать или выключать определенный рецептор. [4] Однако внешняя форма образующегося комплекса становится катализатором изменения ответа тканевой мишени на СЭРМ. [3] [19] [4] [20]
Рентгеновская кристаллография эстрогенов или антиэстрогенов показала, как лиганды программируют рецепторный комплекс на взаимодействие с другими белками. Лигандсвязывающий домен ER демонстрирует, как лиганды способствуют и предотвращают связывание коактиватора в зависимости от формы эстрогенового или антиэстрогенного комплекса. Широкий спектр лигандов, которые связываются с ER, может создавать спектр комплексов ER, которые являются полностью эстрогенными или антиэстрогенными в определенном сайте-мишени. [3] [4] [20] Основным результатом связывания лиганда с ЭР является структурная перестройка лигандсвязывающего кармана, прежде всего в AF-2 С-концевой области. Связывание лигандов с ЭР приводит к образованию гидрофобного кармана, который регулирует кофакторы и фармакологию рецептора. Правильное сворачивание лиганд-связывающего домена необходимо для активации транскрипции и для взаимодействия ER с рядом коактиваторов. [4]
Коактиваторы — это не просто белки-партнеры, соединяющие сайты в комплекс. Коактиваторы играют активную роль в модификации активности комплекса. Посттрансляционная модификация коактиваторов может привести к созданию динамической модели действия стероидных гормонов посредством множественных киназных путей, инициируемых рецепторами фактора роста клеточной поверхности . Под руководством множества специалистов по ремоделированию белков для формирования мультибелкового коактиваторного комплекса, который может взаимодействовать с фосфорилированным ER в определенном сайте промотора гена, основной коактиватор сначала должен рекрутировать определенный набор коактиваторов. Белки, которые коактиватор ядра собирает в коактивированный комплекс ядра, обладают индивидуальной ферментативной активностью по метилированию или ацетилированию соседних белков. Субстраты ER или кофермент A могут быть полиубиквитинированы в результате нескольких циклов реакции или, в зависимости от связывающих белков, они могут либо дополнительно активироваться, либо разрушаться под действием 26S протеасомы . [4]
Следовательно, чтобы иметь эффективную транскрипцию гена, которая запрограммирована и нацелена на структуру и статус фосфорилирования ER и коактиваторов, необходим динамический и циклический процесс ремоделирования способности к транскрипционной сборке, после чего транскрипционный комплекс мгновенно разрушается. обычно разрушается протеасомой. [4]
Структура и функции
[ редактировать ]Отношения структура-деятельность
[ редактировать ]Основная структура SERM имитирует матрицу 17β-эстрадиола . Они имеют два ароматических кольца, разделенных 1-3 атомами (часто расположение стильбенового типа). Между двумя фенилами ядра СЭРМ обычно имеют 4-замещенную фенильную группу, которая при связывании с ЭР выступает из положения эстратриенового ядра, так что спираль 12 перемещается из отверстия рецептора и блокирует пространство, где обычно должны располагаться белки-коактиваторы. связываться и вызывать активность агониста ER. В основной части SERM было много вариаций, в то время как гибкость в отношении того, что допускается в боковой цепи, была меньшей. [7] SERM можно классифицировать по их базовой структуре.
Трифенилэтилены первого поколения
[ редактировать ]
Первым основным структурным классом молекул типа SERM являются трифенилэтилены. Стильбеновое ядро (похожее на нестероидный эстроген, диэтилстильбестрол) по существу имитирует стероидные эстрогены, такие как 17β-эстрадиол, тогда как боковая цепь перекрывает 11-е положение стероидного ядра. [7] Производные трифенилэтилена имеют дополнительную фенильную группу, присоединенную к этиленовой мостиковой группе. Способность фенолов образовывать 3-положение Н-связей является важным требованием для связывания ER. [22]

Первый препарат – кломифен. [23] имеет хлорзаместитель в этиленовой боковой цепи, который обеспечивает такое же сродство связывания, как и обнаруженный позже препарат тамоксифен. Кломифен представляет собой смесь эстрогенных ( цис-форма ) и антиэстрогенных изомеров ( транс-форма ). [22] Цис и транс определяются с точки зрения геометрического соотношения двух незамещенных фенильных колец. [23] Два изомера кломифена имеют разные профили: транс-форма имеет активность, более похожую на активность тамоксифена, тогда как цис-форма ведет себя больше как 17β-эстрадиол. [7] Цис примерно в десять раз более эффективен, чем транс. Однако транс-изомер является наиболее мощным стимулятором гипертрофии эпителиальных клеток, поскольку кломифен является антагонистом в низких дозах и агонистом в высоких дозах. [23] Изомеры-антагонисты могут вызывать ингибирующие эстрогенные эффекты при раке матки и молочной железы, но эстрогенный изомер может сочетаться с новыми рецепторами, вызывая эстрогеноподобные эффекты в костях. [24]

Тамоксифен стал методом выбора для женщин с диагнозом всех стадий гормонозависимого рака молочной железы, то есть рака молочной железы, который является как ER, так и/или прогестерон-положительным. В США его также назначают для профилактической химиопрофилактики у женщин с высоким риском развития рака молочной железы. [25] Тамоксифен представляет собой чистый антиэстрогенный транс-изомер и оказывает различное действие на ткани-мишени эстрогена по всему организму. Тамоксифен оказывает селективное антиэстрогенное действие на молочные железы, но эстрогеноподобен при раке костей и эндометрия. [24] Тамоксифен подвергается I фазе метаболизма в печени под действием микросомальных ферментов цитохрома P450 (CYP) . Основными метаболитами тамоксифена являются N -десметилтамоксифен и 4-гидрокситамоксифен .
Кристаллографическая структура 4-гидрокситамоксифена [26] взаимодействует с аминокислотами ЭР внутри лигандсвязывающего домена. [27] Контакт между фенольной группой, молекулой воды, глутаматом и аргинином в рецепторе (ERα; Glu 353/Arg 394) разрешается в результате высокоаффинного связывания, так что образуется 4-гидрокситамоксифен с фенольным кольцом, напоминающим кольцо A 17β-. эстрадиол имеет относительное сродство связывания более чем в 100 раз выше, чем тамоксифен, который не содержит фенола. Если его ОН-группа удаляется или изменяется ее положение, аффинность связывания снижается. [7] [22]
Трифенилэтиленовая группа и боковая цепь необходимы для связывания тамоксифена с ЭР, тогда как для 4-гидрокситамоксифена боковая цепь и фенилпропен не являются решающими структурными элементами для связывания с ЭР. Основность и длина боковой цепи, по-видимому, не играют решающей роли для сродства связывания тамоксифена с ER или β-кольца тамоксифена, но стильбеновый фрагмент тамоксифена необходим для связывания с ER. Гидроксильная группа имеет особое значение для связывания ER 4-гидрокситамоксифена, а этильная боковая цепь тамоксифена выступает из лигандсвязывающего домена ER. [27]
У немногих пользователей тамоксифена наблюдался повышенный уровень рака матки, приливов и тромбоэмболий. Препарат также может вызывать гепатокарциному у крыс. Вероятно, это связано с этильной группой стильбенового ядра тамоксифена, которая подвержена аллильной окислительной активации, вызывающей алкилирование ДНК и разрыв цепи. Эта проблема позже исправляется с помощью торемифена. [7] Тамоксифен более беспорядочен, чем ралоксифен, в целевых сайтах из-за связи между аминокислотой ER в Asp-351 и антиэстрогенной боковой цепью SERM. Боковая цепь тамоксифена не может нейтрализовать Asp-351, поэтому этот сайт аллостерически влияет на AF-1 на проксимальном конце ER. Эту проблему решает препарат ралоксифен второго поколения. [24]

Торемифен — хлорированное производное нестероидного трифенилэтиленового антиэстрогена тамоксифена. [7] с хлорным заместителем в этиленовой боковой цепи, обеспечивающим сродство связывания, аналогичное сродству тамоксифена. [22] Структура и взаимосвязь активности торемифена аналогичны структуре тамоксифена, но он значительно лучше предыдущего препарата в отношении алкилирования ДНК. Присутствие добавленного атома хлора снижает стабильность катионов, образующихся из активированных аллильных метаболитов, и, таким образом, снижает потенциал алкилирования, и действительно, торемифен не вызывает образования аддукта ДНК в гепатоцитах грызунов . Торемифен защищает от потери костной массы у крыс с удаленными яичниками и клинически влияет на маркеры резорбции кости аналогично тамоксифену. [7] Торемифен подвергается I фазе метаболизма с помощью микросомальных ферментов цитохрома P450, как и тамоксифен, но преимущественно с помощью изоформы CYP3A4. Торемифен образует два основных метаболита: N-десметилторемифен и деаминогидрокситоремифен (оспемифен), подвергаясь N-деметилированию и дезаминированию-гидроксилированию. N-десметилторемифен обладает такой же эффективностью, как и торемифен, тогда как 4-гидрокситоремифен имеет более высокую аффинность связывания с ЭР, чем торемифен. [26] Роль 4-гидрокситоремифена аналогична роли 4-гидрокситамоксифена. [28]
Бензотиофены второго поколения
[ редактировать ]
препаратам SERM второго поколения Ралоксифен относится к бензотиофеновым . Он имеет высокое сродство к ЭР с мощной антиэстрогенной активностью и тканеспецифическими эффектами, отличными от эстрадиола. [19] Ралоксифен является агонистом ЭР в костях и сердечно-сосудистой системе, но в тканях молочной железы и эндометрии он действует как антагонист ЭР. Он интенсивно метаболизируется путем конъюгации глюкуронида в кишечнике, поэтому его биодоступность составляет всего 2%, в то время как биодоступность тамоксифена и торемифена составляет примерно 100%. [26]
Преимущество ралоксифена перед трифенилэтилентамоксифеном заключается в снижении влияния на матку. Гибкая шарнирная группа, а также антиэстрогенная боковая цепь фенил-4-пиперидиноэтокси важны для минимизации эффектов на матку. Благодаря своей гибкости боковая цепь может иметь ортогональное расположение относительно ядра. [7] так что амин боковой цепи ралоксифена находится на 1 Å ближе, чем тамоксифен, к аминокислоте Asp-351 в лигандсвязывающем домене ERα. [24] [29]
Критическая роль тесной связи между гидрофобной боковой цепью ралоксифена и гидрофобным остатком рецептора для изменения как формы, так и заряда внешней поверхности комплекса SERM-ER была подтверждена на примере производных ралоксифена. Увеличение расстояния взаимодействия между ралоксифеном и Asp-351 с 2,7 Å до 3,5-5 Å вызывает усиление эстрогеноподобного действия комплекса ралоксифен-ERα. При замене пиперидинового кольца ралоксифена на циклогексан лиганд теряет антиэстрогенные свойства и становится полным агонистом. Взаимодействие между антиэстрогенной боковой цепью SERM и аминокислотой Asp-351 является важным первым шагом в подавлении AF-2. Он перемещает спираль 12 подальше от лиганд-связывающего кармана, тем самым предотвращая связывание коактиваторов с комплексом SERM-ER. [24] [29]
Третье поколение
[ редактировать ]
Соединения третьего поколения либо не стимулируют матку, не улучшают потенцию, не вызывают значительного увеличения приливов жара или даже сочетают в себе эти положительные свойства. [7]
Первый дигидронафталин-СЭРМ, нафоксидин , был клиническим кандидатом для лечения рака молочной железы, но имел побочные эффекты, включая тяжелую фототоксичность. В нафоксидине все три фенила расположены в копланарном порядке, как тамоксифен. Но при гидрировании двойная связь нафоксидена восстановилась, и оба фенила оказались цис-ориентированными. Боковая цепь, содержащая амины, может затем принять аксиальную конформацию и расположить эту группу ортогонально плоскости ядра, как ралофоксифен и другие менее утеротропные SERM.

Модификации нафоксидина привели к созданию лазофоксифена. Лазофоксифен является одним из наиболее эффективных СЭРМ, защищающих от потери костной массы и снижения уровня холестерина. Превосходная эффективность лазофоксифена при пероральном приеме объясняется снижением кишечной глюкуронизации фенола. [7] В отличие от ралоксифена, лазофоксифен удовлетворяет требованиям фармакофорной модели, прогнозирующей устойчивость к глюкуронидации стенки кишечника. Структурное требование — неплоская топология со стерическим объемом, близким к плоскости конденсированной бициклической ароматической системы. [30] Взаимодействия между ЭР и лазофоксифеном согласуются с общими особенностями распознавания SERM-ER. Большая гибкая боковая цепь лазофоксифена заканчивается пирролидиновой головной группой и направляется к поверхности белка, где она напрямую мешает позиционированию спирали AF-2. Между лазофоксифеном и Asp-351 образуется солевой мостик. Нейтрализацией заряда в этой области ЭР можно объяснить некоторые антиэстрогенные эффекты лазофоксифена. [12]

Индольная система служила основной единицей SERM, и когда амин присоединяется к индолу с помощью бензилоксиэтила, полученные соединения не обладают доклинической активностью в матке, сохраняя при этом крысиную кость с полной эффективностью в низких дозах. Базедоксифен является одним из таких соединений. Основной связывающий домен состоит из 2-фенил-3-метилиндола и гексаметиленаминового кольца в аффектной области боковой цепи. Метаболизируется путем глюкуронидации, при этом абсолютная биодоступность составляет 6,2%, что в 3 раза выше, чем у ралоксифена. Он оказывает агонистическое действие на костный и липидный обмен, но не на эндометрий молочной железы и матки. [31] Он хорошо переносится и не вызывает увеличения частоты приливов. [ написание? ] гипертрофия матки или болезненность молочных желез. [7]

Оспемифен представляет собой трифенилэтилен и известный метаболит торемифена. Структурно он очень похож на тамоксифен и торемифен. Оспемифен не имеет 2-(диметиламино)этоксигруппы, как тамоксифен. Исследования взаимосвязи структура-активность показали, что при удалении этой группы агонистической активности тамоксифена в матке значительно снижается, но не в костях и сердечно-сосудистой системе. Доклинические и клинические данные показывают, что оспемифен хорошо переносится и не имеет серьезных побочных эффектов. Преимущества, которые оспемифен может иметь перед другими СЭРМ, заключаются в его нейтральном воздействии на приливы и эффекте агониста ER на влагалище, улучшающем симптомы сухости влагалища. [32]
Режимы привязки
[ редактировать ]
Известно, что SERM имеют четыре различных режима связывания с ER. Одной из этих особенностей являются прочные водородные связи между лигандом и Arg-394 и Glu-353 ERα, которые выстилают «карман А-кольца» и помогают лиганду оставаться в связывающем кармане ER. Это отличается от 17β-эстрадиола, который связан водородной связью с His-524 в «кармане D-кольца». [13] Другие отличительные особенности связывания с лиганд-связывающим карманом заключаются в почти плоской «ядерной» структуре, обычно состоящей из биарильного гетероцикла , эквивалентного A-кольцу и B-кольцу 17β-эстрадиола, с соответствующим сайтом связывания; объемистая боковая цепь биарильной структуры, аналогичная B-кольцу 17β-эстрадиола, и, наконец, вторая боковая группа, эквивалентная C- и D-кольцу и обычно ароматическая, заполняет оставшийся объем лигандсвязывающего кармана. [30]
Небольшие различия между двумя подтипами ER были использованы для разработки подтип-селективных модуляторов ER, но большое сходство между двумя рецепторами делает разработку очень сложной. Аминокислоты в лиганд-связывающих доменах различаются по двум положениям: Leu-384 и Met-421 в ERα и Met-336 и Ile-373 в ERβ, но они имеют одинаковую гидрофобность и занимаемые объемы. Однако форма и вращательный барьер аминокислотных остатков не одинаковы, что позволяет различать α- и β-грани полости связывания между ERα и ERβ. Это вызывает преимущественное ERα-связывание заместителей лиганда , которые ориентированы вниз по направлению к Met-336, тогда как заместители лиганда, ориентированные вверх по направлению к Met-336, с большей вероятностью связываются с ERβ. Другое отличие заключается в Val-392 в ERα, который заменен Met-344 в ERβ. Объем связывающего кармана ERβ немного меньше, а форма немного отличается от кармана ERα. Многие ERβ-селективные лиганды имеют в основном плоское расположение, поскольку полость связывания ERβ немного уже, чем у ERα, однако это само по себе приводит к умеренной селективности. Чтобы достичь высокой селективности, лиганд должен разместить заместители очень близко к одному или нескольким аминокислотным различиям между ERα и ERβ, чтобы создать сильную силу отталкивания по отношению к рецептору другого подтипа. Кроме того, структура лиганда должна быть жесткой. В противном случае отталкивающие взаимодействия могут привести к конформационным изменениям лиганда и, следовательно, к созданию альтернативных способов связывания. [13]
Трифенилэтилены первого поколения
[ редактировать ]Тамоксифен преобразуется цитохромом Р450 печени в 4-гидрокситамоксифен. [12] и является более селективным антагонистом подтипа ERα, чем ERβ. [33] 4-гидрокситамоксифен связывается с ЭР в том же кармане связывания, который распознает 17β-эстрадиол. Распознавание 4-гидрокситамоксифена рецептором, по-видимому, контролируется двумя структурными особенностями 4-гидрокситамоксифена: фенольным кольцом А и объемной боковой цепью. Фенольное кольцо А образует водородные связи с боковыми группами Arg-394, Glu-354 ER и структурно консервативной воды. Объемная боковая цепь, выступающая из полости связывания, вытесняет спираль 12 из кармана связывания лиганда и перекрывает часть кармана связывания коактиватора. В образовании комплекса ER-4-гидрокситамоксифен участвуют белки-корепрессоры. Это приводит к снижению синтеза ДНК и ингибированию активности эстрогенов. [12] Кломифен и торимефен обладают сродством связывания, аналогичным сродству тамоксифена. [22] Таким образом, эти два препарата являются более селективными антагонистами подтипа ERα, чем ERβ. [33]
Бензотиофены второго поколения
[ редактировать ]
Ралоксифен, как и 4-гидрокситамоксифен, связывается с ERα с помощью гидроксильной группы его фенольного «кольца А» посредством водородных связей с Arg-394 и Glu-353. В дополнение к этим связям ралоксифен образует вторую водородную связь с ER через боковую группу His-524 из-за присутствия второй гидроксильной группы в «D-кольце». Эта водородная связь также отличается от связи между 17β-эстрадиолом и His-524, поскольку имидазольное кольцо His-524 повернуто, чтобы компенсировать разницу в положении кислорода в ралоксифене и в 17β-эстрадиоле. Как и в 4-гидрокситамоксифене, объемная боковая цепь ралоксифена замещает спираль 12. [12]
Третье поколение
[ редактировать ]Взаимодействие лазофоксифена с ERα типично для взаимодействий между SERM-ERα, например, почти плоская топология (карбоцикл тетрагидронафталина), водородные связи с Arg-394 и Glu-353 и фенильные боковые цепи лазофоксифена, заполняющие C-кольцо и D-кольцо. объем лигандсвязывающего кармана. Лазофоксифен отклоняет спираль 12 и предотвращает связывание белков-коактиваторов с мотивами LXXLL. Это достигается за счет того, что лазофоксифен занимает пространство, обычно заполняемое боковой группой Leu-540, и модулирует конформацию остатков спирали 11 (His-524, Leu-525). препарата Кроме того, лазофоксифен также напрямую препятствует расположению спирали 12 со стороны этилпирролидиновой группы . [12] Исследования in vitro показывают, что базедоксифен конкурентно блокирует 17β-эстрадиол за счет высокого и одинакового связывания как с ERα, так и с ERβ. [34] Основной связывающий домен базедоксифена состоит из 2-фенил-3-метилиндола и гексаметиленаминового кольца в затронутой области боковой цепи. [31]
Оспемифен представляет собой окислительно-дезаминированный метаболит торемифена, который связывается с ЭР так же, как торемифен и тамоксифен. Конкурентное связывание с ERα и ERβ трех метаболитов: 4-гидрокси оспемифена, 4'-гидрокси оспемифена и 4-гидрокси-карбоновой кислоты с боковой цепью оспемифена, по крайней мере, столь же велико, как и у исходного соединения. [35]
История
[ редактировать ]Открытие СЭРМ стало результатом попыток разработать новые противозачаточные средства. Кломифен и тамоксифен предотвращали зачатие у крыс, но действовали наоборот у людей. Кломифен успешно вызывал овуляцию у бесплодных женщин , и 1 февраля 1967 года он был одобрен в США для лечения дисфункции овуляции у женщин, пытающихся зачать ребенка. [5] Токсикологические проблемы препятствовали долгосрочному использованию кломифена и дальнейшей разработке лекарств для других потенциальных применений, таких как лечение и профилактика рака молочной железы . [6]
Прошло еще десять лет, прежде чем тамоксифен был одобрен в декабре 1977 года не как противозачаточное средство, а как гормональное средство для лечения и профилактики рака молочной железы. [6] Открытие в 1987 году того, что SERM тамоксифен и ралоксифен , которые тогда считались антиэстрогенами из-за антагонистического воздействия на ткани молочной железы, продемонстрировали эстрогенные эффекты в предотвращении потери костной массы у крыс с удаленными яичниками, оказало большое влияние на наше понимание функции рецепторов эстрогена и ядерных рецепторов. в общем. [7] Термин SERM был введен для описания этих соединений, которые обладают комбинацией активности агониста эстрогена , частичного агониста или антагониста в зависимости от ткани. [5] Было показано, что торемифен совместим с тамоксифеном, и в 1996 году он был одобрен для использования при лечении рака молочной железы у женщин в постменопаузе. [36]
Ралоксифен изначально не смог стать лекарством от рака молочной железы из-за его плохих результатов по сравнению с тамоксифеном в лабораторных условиях. [19] но эстрогенное воздействие ралоксифена на кости привело к его повторному открытию и одобрению в 1997 году. [6] Он был одобрен для профилактики и лечения остеопороза и стал первым клинически доступным СЭРМ для предотвращения как остеопороза, так и рака молочной железы. [7] Оспемифен был одобрен 26 февраля 2013 года для лечения диспареунии средней и тяжелой степени , которая является симптомом обусловленной менопаузой вульвы и влагалища, атрофии . Комбинированная терапия конъюгированными эстрогенами SERM и базедоксифеном была одобрена 3 октября 2013 года для лечения вазомоторных симптомов, связанных с менопаузой. Базедоксифен также используется для профилактики постменопаузального остеопороза. [6] Поиск мощного СЭРМ с эффективностью для костей и лучшей биодоступностью, чем ралоксифен, привел к открытию лазофоксифена. [12] Хотя лазофоксифен был одобрен в 2009 году, он не продавался в течение трех лет после одобрения, поэтому срок действия его регистрационного удостоверения истек. [37] В Европе базедоксифен показан для лечения остеопороза у женщин в постменопаузе с повышенным риском переломов, тогда как в Индии или мелоксифен используется для лечения дисфункциональных маточных кровотечений и контроля над рождаемостью. [6]
См. также
[ редактировать ]- Эстрогендепривационная терапия
- Список селективных модуляторов рецепторов эстрогена
- Селективный модулятор андрогенных рецепторов
- Селективный деградатор рецепторов эстрогена
- Селективный модулятор рецепторов
- Хронология развития лечения рака
Ссылки
[ редактировать ]- ^ Хирш Х.Д., Ши Э., Такер Х.Л. (июнь 2017 г.). «ERAA для лечения менопаузы: приветствуем «дизайнерские эстрогены» » . Клив Клин, Джей Мед . 84 (6): 463–470. дои : 10.3949/ccjm.84a.15140 . ПМИД 28628428 .
- ^ Арчер Д.Ф. (август 2020 г.). «Оспемифен: меньший венозный тромбоз, чем другие селективные модуляторы эстрогеновых рецепторов у женщин в постменопаузе с атрофией вульвовагини». Менопауза . 27 (8): 846–847. дои : 10.1097/GME.0000000000001600 . ПМИД 32576803 . S2CID 220045301 .
- ^ Перейти обратно: а б с д и Риггс Б.Л., Хартманн Л.С. (февраль 2003 г.). «Селективные модуляторы рецепторов эстрогена - механизмы действия и применение в клинической практике». Медицинский журнал Новой Англии . 348 (7): 618–29. дои : 10.1056/NEJMra022219 . ПМИД 12584371 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д Максимов П.Ю., Ли Т.М., Джордан В.К. (май 2013 г.). «Открытие и разработка селективных модуляторов эстрогеновых рецепторов (SERM) для клинической практики» . Современная клиническая фармакология . 8 (2): 135–55. дои : 10.2174/1574884711308020006 . ПМЦ 3624793 . ПМИД 23062036 .
- ^ Перейти обратно: а б с д и ж г Пикар Дж. Х., Комм Б.С. (сентябрь 2015 г.). «Селективные модуляторы рецепторов эстрогена и комбинированная терапия конъюгированными эстрогенами/базедоксифеном: обзор воздействия на грудь». Пострепродуктивное здоровье . 21 (3): 112–21. дои : 10.1177/2053369115599090 . ПМИД 26289836 . S2CID 206825977 .
- ^ Перейти обратно: а б с д и ж г час я дж Миркин С., Пикар Дж.Х. (январь 2015 г.). «Селективные модуляторы рецепторов эстрогена (SERM): обзор клинических данных». Матуритас . 80 (1): 52–7. дои : 10.1016/j.maturitas.2014.10.010 . ПМИД 25466304 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Миллер КП (2002). «SERM: эволюционная химия, революционная биология». Текущий фармацевтический дизайн . 8 (23): 2089–111. дои : 10.2174/1381612023393404 . ПМИД 12171520 .
- ^ Сюй, Джейн Ю.; О'Коннелл, Мишель А.; Нотини, Лорен; Чунг, Ада С.; Цвикль, Сав; Панг, Кен С. (18 июня 2021 г.). «Селективные модуляторы рецепторов эстрогена: потенциальный вариант небинарной гормональной помощи, подтверждающей пол?» . Границы эндокринологии . 12 : 701364. дои : 10.3389/fendo.2021.701364 . ISSN 1664-2392 . ПМЦ 8253879 . ПМИД 34226826 .
- ^ Кэмерон Дж.Л., Кэмерон А.М. (20 ноября 2013 г.). Современная хирургическая терапия . Elsevier Науки о здоровье. стр. 582–. ISBN 978-0-323-22511-3 .
- ^ Перейти обратно: а б Хуан X, Асланян Р.Г. (19 апреля 2012 г.). Тематические исследования в области открытия и разработки современных лекарств . Джон Уайли и сыновья. стр. 392–394. ISBN 978-1-118-21967-6 .
- ^ Перейти обратно: а б с д Кремозер С., Альберс М., Беррис Т.П., Дойшле У., Коегль М. (октябрь 2007 г.). «Панирование SNuRM: использование профилирования кофакторов для рационального открытия селективных модуляторов ядерных рецепторов». Открытие наркотиков сегодня . 12 (19–20): 860–9. дои : 10.1016/j.drudis.2007.07.025 . ПМИД 17933688 .
- ^ Перейти обратно: а б с д и ж г час я Розано С., Стек-Мартина Э., Лаппано Р., Маджолини М. (2011). «Структурный подход к открытию новых селективных модуляторов рецепторов эстрогена». Современная медицинская химия . 18 (8): 1188–94. дои : 10.2174/092986711795029645 . ПМИД 21291367 .
- ^ Перейти обратно: а б с д Нильссон С., Келер К.Ф., Густавссон Йо (октябрь 2011 г.). «Разработка подтип-селективных препаратов на основе рецепторов эстрогена». Обзоры природы. Открытие наркотиков . 10 (10): 778–92. дои : 10.1038/nrd3551 . ПМИД 21921919 . S2CID 23043739 .
- ^ Келер К.Ф., Хельгеро Л.А., Халдосен Л.А., Уорнер М., Густафссон Дж.А. (май 2005 г.). «Размышления об открытии и значении бета-рецептора эстрогена» . Эндокринные обзоры . 26 (3): 465–78. дои : 10.1210/er.2004-0027 . ПМИД 15857973 .
- ^ Дутертр М., Смит К.Л. (ноябрь 2000 г.). «Молекулярные механизмы действия селективного модулятора эстрогеновых рецепторов (СЭРМ)» . Журнал фармакологии и экспериментальной терапии . 295 (2): 431–7. ПМИД 11046073 .
- ^ Сюй С, Ян В, Ли Ю, Ван Ю (январь 2010 г.). «Открытие модуляторов рецепторов эстрогена: обзор виртуального скрининга и усилий SAR». Мнение экспертов об открытии лекарств . 5 (1): 21–31. дои : 10.1517/17460440903490395 . ПМИД 22823969 . S2CID 207492889 .
- ^ Бжозовский А.М., Пайк А.К., Даутер З., Хаббард Р.Э., Бонн Т., Энгстрем О., Оман Л., Грин Г.Л., Густафссон Йо, Карлквист М. (1997). «Молекулярные основы агонизма и антагонизма рецептора эстрогена». Природа . 389 (6652): 753–8. Бибкод : 1997Natur.389..753B . дои : 10.1038/39645 . ПМИД 9338790 . S2CID 4430999 .
- ^ Шиау А.К., Барстад Д., Лория П.М., Ченг Л., Кушнер П.Дж., Агард Д.А., Грин Г.Л. (1998). «Структурная основа распознавания рецептора/коактиватора эстрогена и антагонизм этого взаимодействия тамоксифена» . Клетка . 95 (7): 927–37. дои : 10.1016/S0092-8674(00)81717-1 . ПМИД 9875847 . S2CID 10265320 .
- ^ Перейти обратно: а б с д и ж г час я Муса М.А., Хан М.О., Купервуд Дж.С. (2007). «Медицинская химия и новые стратегии, применяемые к разработке селективных модуляторов рецепторов эстрогена (SERM)». Современная медицинская химия . 14 (11): 1249–61. дои : 10.2174/092986707780598023 . ПМИД 17504144 .
- ^ Перейти обратно: а б с д и ж Льюис Дж.С., Джордан ВК (декабрь 2005 г.). «Селективные модуляторы рецепторов эстрогена (СЭРМ): механизмы антиканцерогенеза и лекарственной устойчивости». Мутационные исследования . 591 (1–2): 247–63. дои : 10.1016/j.mrfmmm.2005.02.028 . ПМИД 16083919 .
- ^ Перейти обратно: а б Фэн Кью, О'Мэлли BW (ноябрь 2014 г.). «Модуляция ядерных рецепторов - роль корегуляторов в действиях селективных модуляторов эстрогеновых рецепторов (SERM)» . Стероиды . 90 : 39–43. doi : 10.1016/j.steroids.2014.06.008 . ПМК 4192004 . ПМИД 24945111 .
- ^ Перейти обратно: а б с д и Фанг Х., Тонг В., Ши Л.М., Блер Р., Перкинс Р., Бранхам В., Хасс Б.С., Се Кью, Дайал С.Л., Моланд С.Л., Шихан Д.М. (март 2001 г.). «Взаимосвязь структура-активность для большого разнообразного набора природных, синтетических и экологических эстрогенов». Химические исследования в токсикологии . 14 (3): 280–94. CiteSeerX 10.1.1.460.20 . дои : 10.1021/tx000208y . ПМИД 11258977 .
- ^ Перейти обратно: а б с Кларк Дж. Х., Маркаверич Б. М. (1981). «Агонистически-антагонистические свойства кломифена: обзор». Фармакология и терапия . 15 (3): 467–519. дои : 10.1016/0163-7258(81)90055-3 . ПМИД 7048350 .
- ^ Перейти обратно: а б с д и Дженсен Э.В., Джордан ВК (июнь 2003 г.). «Рецептор эстрогена: модель молекулярной медицины». Клинические исследования рака . 9 (6): 1980–9. ПМИД 12796359 .
- ^ Сингх М.Н., Стрингфеллоу Х.Ф., Параскеваидис Э., Мартин-Хирш П.Л., Мартин Флорида (апрель 2007 г.). «Тамоксифен: важные аспекты многофункционального соединения с органоспецифическими свойствами». Обзоры лечения рака . 33 (2): 91–100. дои : 10.1016/j.ctrv.2006.09.008 . ПМИД 17178195 . ]
- ^ Перейти обратно: а б с Морелло К.К., Вурц Г.Т., ДеГрегорио М.В. (30 сентября 2012 г.). «Фармакокинетика селективных модуляторов рецепторов эстрогена». Клиническая фармакокинетика . 42 (4): 361–72. дои : 10.2165/00003088-200342040-00004 . ПМИД 12648026 . S2CID 13003168 .
- ^ Перейти обратно: а б де Медина П., Фавр Г., Пуаро М. (ноябрь 2004 г.). «Множественное нацеливание противоопухолевого препарата тамоксифена: исследование структуры и активности». Современная медицинская химия. Противораковые агенты . 4 (6): 491–508. дои : 10.2174/1568011043352696 . ПМИД 15579015 .
- ^ Готье С., Мэйлхот Дж., Лабри Ф. (май 1996 г.). «Новый высокостереоселективный синтез (Z)-4-гидрокситамоксифена и (Z)-4-гидрокситоремифена посредством реакции Макмерри». Журнал органической химии . 61 (11): 3890–3893. дои : 10.1021/jo952279l . ПМИД 11667248 .
- ^ Перейти обратно: а б Джордан ВК (март 2003 г.). «Антиэстрогены и селективные модуляторы эстрогеновых рецепторов как многофункциональные лекарства. 2. Клинические аспекты и новые агенты». Журнал медицинской химии . 46 (7): 1081–111. дои : 10.1021/jm020450x . ПМИД 12646017 .
- ^ Перейти обратно: а б Вайдос Ф.Ф., Хот Л.Р., Геохеган К.Ф., Саймонс С.П., ЛеМотт П.К., Дэнли Д.Е., Аммирати М.Дж., Пандит Дж. (май 2007 г.). «Кристаллическая структура 2,0 А лигандсвязывающего домена ERalpha в комплексе с лазофоксифеном» . Белковая наука . 16 (5): 897–905. дои : 10.1110/ps.062729207 . ПМК 2206632 . ПМИД 17456742 .
- ^ Перейти обратно: а б Кунг А.В., Чу Э.Ю., Сюй Л. (июнь 2009 г.). «Базедоксифен: новый селективный модулятор рецепторов эстрогена для лечения постменопаузального остеопороза». Экспертное заключение по фармакотерапии . 10 (8): 1377–85. дои : 10.1517/14656560902980228 . ПМИД 19445558 . S2CID 20781017 .
- ^ Дженнари Л., Мерлотти Д., Валлегги Ф., Нути Р. (июнь 2009 г.). «Использование оспемифена у женщин в постменопаузе». Экспертное заключение об исследуемых препаратах . 18 (6): 839–49. дои : 10.1517/13543780902953715 . ПМИД 19466874 . S2CID 21537130 .
- ^ Перейти обратно: а б Танеджа С.С., Смит М.Р., Далтон Дж.Т., Рэгоу С., Барнетт Г., Штайнер М., Веверка К.А. (март 2006 г.). «Торемифен - многообещающая терапия для профилактики рака простаты и осложнений андрогенной депривационной терапии». Экспертное заключение об исследуемых препаратах . 15 (3): 293–305. дои : 10.1517/13543784.15.3.293 . ПМИД 16503765 . S2CID 29510508 .
- ^ Паласиос С (2006). «Эффекты СЭРМ на эндометрий» . В Санчес AC, Калаф и Альсина Х, Дуэньяс-Диес Х.Л. (ред.). Селективные модуляторы рецепторов эстрогена - новая марка многоцелевых препаратов (1-е изд.). Берлин: Шпрингер. стр. 282–3. дои : 10.1007/3-540-34742-9_11 . ISBN 978-3-540-24227-7 .
- ^ «Сэншио (оспемифен)» . Европейское агентство лекарственных средств (EMA). Архивировано из оригинала 20 июня 2018 г. Проверено 2 ноября 2015 г.
- ^ «Фарестон» . Европейское агентство лекарственных средств (EMA). Архивировано из оригинала 20 июня 2018 г. Проверено 2 ноября 2015 г.
- ^ «Фаблин» . Европейское агентство лекарственных средств (EMA). Архивировано из оригинала 12 февраля 2018 г. Проверено 2 ноября 2015 г.