Семейство TCF/LEF
Семейство TCF/LEF ( семейство факторов Т-клеток/факторов лимфоидного энхансера ) представляет собой группу генов, которые кодируют факторы транскрипции , которые связываются с ДНК через SOX-подобный групповой домен с высокой подвижностью . Они участвуют в сигнальном пути Wnt , особенно во время эмбрионального развития. [2] и развитие стволовых клеток, [3] но также было обнаружено, что он играет роль в развитии рака [4] и диабет. [5] Факторы TCF/LEF привлекают коактиватор бета-катенин для усиления элементов генов, на которые они нацелены. Они также могут вербовать членов семейства корепрессоров Граучо . [6]
История
[ редактировать ]Открытие генов TCF/LEF как компонентов ядерного пути Wnt в 90-х годах. [7] [8] стал ключевым прорывом в области исследований передачи сигналов Wnt, заполнив важный пробел в знаниях и позволив последующее понимание регуляции транскрипции генов-мишеней Wnt, особенно при эмбриональном развитии и раке.
До этого открытия было известно только, что вышестоящие механизмы передачи сигналов Wnt регулируют цитоплазматическое содержание белка бета-катенина, который, как следствие, перемещается в ядро клетки. Однако, поскольку в белковой структуре бета-катенина не обнаружено какого-либо ДНК-связывающего домена, все еще было неясно, как ядерный бета-катенин может регулировать гены-мишени Wnt. После открытия была создана модель, согласно которой бета-катенин, регулируемый передачей сигналов Wnt, в ядре прикрепляется к ДНК-связывающим белкам TCF/LEF, которые распознают консенсусную последовательность ДНК вокруг ядра «CTTTG», называемую элементом ответа Wnt (WRE). [9]
Это правило, согласно которому взаимодействие бета-катенин-TCF на ДНК регулирует экспрессию гена-мишени Wnt, тем не менее, было нарушено примерами Wnt- и бета-катенин-независимых функций белков TCF/LEF (например, при развитии ЦНС рыбок данио). [10] ) и функциональная ассоциация Wnt-регулируемого бета-катенина с другими ДНК-связывающими транскрипционными факторами, такими как SOX, [11] ФОКСО , [12] TBXТВХ [13] С другой стороны, это взаимодействие бета-катенин-TCF на ДНК теперь оказывается всего лишь ядром гораздо более крупных белковых комплексов, регулирующих транскрипцию, называемых энхансосомами Wnt. [14] И наоборот, были обнаружены дополнительные механизмы, регулирующие функцию белка TCF/LEF, такие как фосфорилирование. [15] и сумойлирование . [16]
Структура
[ редактировать ]Структура и функция белков TCF/LEF объясняют эту бимодальную функцию. Гены TCF/LEF кодируют белки со сложной структурой, которую, однако, можно обобщить, рассмотрев четыре основных домена:
- N-концевой домен : опосредует взаимодействие с бета-катенином , который является высококонсервативным и опосредует функцию активатора транскрипции .
- Контрольная область : включает последовательности, регулирующие и опосредующие функцию репрессора транскрипции и кодирующие домен, связывающий репрессор транскрипции для семейства Groucho.
- ДНК-связывающий домен : включает очень высококонсервативный ДНК-связывающий домен HMG ( группа высокой мобильности ) и NLS ( последовательность ядерной локализации ).
- С-концевой хвост : может содержать дополнительный ДНК-связывающий домен и дополнительный домен, связывающий репрессор транскрипции. [17]
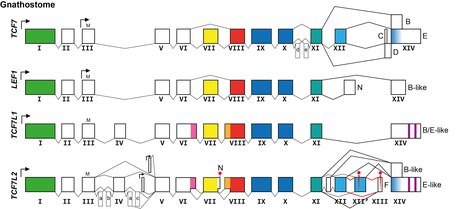
Разнообразие в структуре и функциях белков TCF/LEF обусловлено наличием разных генов. У людей и челюстных позвоночных обычно есть четыре гена, кодирующие белки TCF/LEF:
- TCF7 (также называемый TCF1 )
- LEF1 (также называемый TCF1α )
- TCF7L1 (также называемый TCF3 )
- TCF7L2 (также называемый TCF4 )
Дальнейшее разнообразие происходит за счет экспрессии одним и тем же геном альтернативных транскриптов, кодирующих разные изоформы белка, особенно генов TCF7 и TCF7L2 :
- Существуют изоформы, экспрессируемые с помощью вторичных промоторов, которые кодируют белки, у которых отсутствует обычный N-конец и, следовательно, в частности, домен, связывающий бета-катенин (см. Выше). Эти изоформы белка функционируют не как бимодальные факторы транскрипции, а как конститутивные репрессоры, и они невосприимчивы к регуляции передачи сигналов Wnt, расположенной выше. [18] [19]
- В части транскрипта, кодирующей контрольную область, присутствуют изоформы альтернативного сплайсинга, которые влияют на склонность кодируемых изоформ белка действовать как репрессоры транскрипции (без бета-катенина) или активаторы транскрипции (с бета-катенином). [20]
- Существуют изоформы альтернативного сплайсинга в части транскрипта, кодирующей С-концевой хвост, что приводит к образованию изоформ белка с дополнительным ДНК-связывающим доменом и без него и приводит к изменению рамки считывания, в которой последний экзон транслируется с дополнительным ДНК-связывающим доменом или без него. домен, связывающий транскрипционный корепрессор. [21] [22]
Функция
[ редактировать ]Белки TCF/LEF действуют как бимодальные факторы транскрипции:
- Как описано, белки TCF/LEF действуют как активаторы транскрипции в сочетании с ядерным бета-катенином (и коактиваторами транскрипции, прикрепленными к бета-катенину);
- но без бета-катенина белки TCF/LEF функционируют как репрессоры транскрипции (прикрепленные к корепрессорам транскрипции, членам семейства Groucho).
Т.о., как следствие, гены-мишени Wnt активно репрессируются в отсутствие активности передачи сигналов Wnt, а затем активируются, когда передача сигналов Wnt активно направляет бета-катенин в ядро. [23]
Гены TCF/LEF поддерживают разнообразные функции в эмбриональном развитии, биологии стволовых клеток и при заболеваниях. [24] [25] Учитывая консервативность структуры, функции различных генов и белков TCF/LEF часто дублируются во многих органах и тканях, где важна передача сигналов Wnt, однако генетический анализ с самого начала предполагал, что эта избыточность является лишь частичной, предполагая, что гены и белки TCF/LEF являются избыточными. Функции, специфичные для изоформ TCF, многие из которых только сейчас начинают обнаруживаться.
Выдающиеся функции генов TCF/LEF в эмбриональном развитии включают индукцию дорсальной оси позвоночных, формирование передне-заднего паттерна развивающейся центральной нервной системы, развитие нервного гребня и многие функции в развитии органов. Выдающиеся функции генов TCF/LEF в развитии стволовых клеток особенно хорошо изучены во время цикла волосяного фолликула . [26] [27] Гены TCF/LEF играют роль во многих видах рака, причем их роль при колоректальном раке, возможно, наиболее изучена. [28] Однако с генами TCF/LEF также связаны и другие заболевания человека, особенно диабет 2 типа . [29] [30]
Ссылки
[ редактировать ]- ^ Торрес-Агила, Нурия П.; Салонна, Марика; Хопплер, Стефан; Ферье, Дэвид ЭК (3 февраля 2022 г.). «Эволюционная диверсификация канонического эффектора передачи сигналов Wnt TCF/LEF у хордовых» . Развитие, рост и дифференциация . 64 (3): 120–137. дои : 10.1111/dgd.12771 . hdl : 2164/18617 . ISSN 0012-1592 . ПМК 9303524 . ПМИД 35048372 . S2CID 246161317 .
- ^ Логан, Катриона Ю.; Нуссе, Роэл (ноябрь 2004 г.). «Сигнальный путь WNT в развитии и заболеваниях». Ежегодный обзор клеточной биологии и биологии развития . 20 (1): 781–810. doi : 10.1146/annurev.cellbio.20.010403.113126 . ПМИД 15473860 .
- ^ Нусс, Р. (май 2008 г.). «Передача сигналов Wnt и контроль стволовых клеток» . Клеточные исследования . 18 (5): 523–7. дои : 10.1038/cr.2008.47 . ПМИД 18392048 . S2CID 2503910 .
- ^ Жан, Т; Риндторф, Н; Бутрос, М. (март 2017 г.). «Передача сигналов Wnt при раке» . Онкоген . 36 (11): 1461–1473. дои : 10.1038/onc.2016.304 . ПМЦ 5357762 . ПМИД 27617575 .
- ^ Лаудес, М. (апрель 2011 г.). «Роль передачи сигналов WNT в превращении мезенхимальных стволовых клеток человека в преадипоциты» . Журнал молекулярной эндокринологии . 46 (2): Р65-72. дои : 10.1530/JME-10-0169 . ПМИД 21247979 .
- ^ Брантьес Х., Баркер Н., ван Эс Дж., Клеверс Х. (февраль 2002 г.). «TCF: Леди Джастис выносит окончательный вердикт по результатам передачи сигналов Wnt». Биол. Хим . 383 (2): 255–61. дои : 10.1515/BC.2002.027 . ПМИД 11934263 . S2CID 25665021 .
- ^ Беренс, Дж; фон Крис, JP; Кюль, М; Брюн, Л; Ведлих, Д; Гроссшедль, Р; Бирчмайер, W (15 августа 1996 г.). «Функциональное взаимодействие бета-катенина с фактором транскрипции LEF-1». Природа . 382 (6592): 638–42. Бибкод : 1996Natur.382..638B . дои : 10.1038/382638a0 . ПМИД 8757136 . S2CID 4369341 .
- ^ Хубер, О; Корн, Р; Маклафлин, Дж; Осуги, М; Херрманн, Б.Г.; Кемлер, Р. (сентябрь 1996 г.). «Ядерная локализация бета-катенина путем взаимодействия с фактором транскрипции LEF-1» . Механизмы развития . 59 (1): 3–10. дои : 10.1016/0925-4773(96)00597-7 . ПМИД 8892228 . S2CID 14324471 .
- ^ Кадиган, КМ; Уотерман, М.Л. (1 ноября 2012 г.). «TCF/LEFs и передача сигналов Wnt в ядре» . Перспективы Колд-Спринг-Харбор в биологии . 4 (11): а007906. doi : 10.1101/cshperspect.a007906 . ПМЦ 3536346 . ПМИД 23024173 .
- ^ Дункан, Р.Н.; Панахи, С; Пиотровский, Т; Дорский, Р.И. (2015). «Идентификация генов Wnt, экспрессируемых в зонах нейронных предшественников во время развития мозга рыбок данио» . ПЛОС ОДИН . 10 (12): e0145810. Бибкод : 2015PLoSO..1045810D . дои : 10.1371/journal.pone.0145810 . ПМК 4699909 . ПМИД 26713625 .
- ^ Кормиш, доктор медицинских наук; Грешник, Д; Зорн, AM (январь 2010 г.). «Взаимодействие между факторами SOX и передачей сигналов Wnt/бета-катенин в развитии и заболевании» . Динамика развития . 239 (1): 56–68. дои : 10.1002/dvdy.22046 . ПМЦ 3269784 . ПМИД 19655378 .
- ^ Эссерс, Массачусетс; де Врис-Смитс, LM; Баркер, Н.; Полдерман, ЧП; Бургеринг, БМ; Корсваген, ХК (20 мая 2005 г.). «Функциональное взаимодействие между бета-катенином и FOXO в передаче сигналов окислительного стресса». Наука . 308 (5725): 1181–4. Бибкод : 2005Sci...308.1181E . дои : 10.1126/science.1109083 . ПМИД 15905404 . S2CID 24572861 .
- ^ Циммерли, Дарио; Боррелли, Костанца; Хауреги-Мигель, Амайя; Седерхольм, Саймон; Брюч, Саломея; Думпас, Николаос; Райхмут, Ян; Мерфи-Зейлер, Фабьен; Аге, Мишель; Баслер, Конрад; Мур, Андреас Э; Канту, Клаудио (18 августа 2020 г.). «TBX3 действует как тканеспецифичный компонент транскрипционного комплекса Wnt/β-катенин» . электронная жизнь . 9 . doi : 10.7554/eLife.58123 . ПМЦ 7434441 . ПМИД 32808927 .
- ^ Гаммонс, М; Биенц, М. (апрель 2018 г.). «Мультипротеиновые комплексы, управляющие передачей сигнала Wnt». Современное мнение в области клеточной биологии . 51 : 42–49. дои : 10.1016/j.ceb.2017.10.008 . ПМИД 29153704 .
- ^ Сокол, С.Ю. (июль 2011 г.). «Передача сигналов Wnt посредством фосфорилирования фактора Т-клеток» . Клеточные исследования . 21 (7): 1002–12. дои : 10.1038/cr.2011.86 . ПМК 3193496 . ПМИД 21606952 .
- ^ Ямамото, Х; Ихара, М; Мацуура, Ю; Кикучи, А. (1 мая 2003 г.). «Сумойлирование участвует в бета-катенин-зависимой активации Tcf-4» . Журнал ЭМБО . 22 (9): 2047–59. дои : 10.1093/emboj/cdg204 . ПМК 156076 . ПМИД 12727872 .
- ^ Хопплер, Стефан; Уотерман, Мэриан Л. (2014). «Эволюционное разнообразие структуры, функций и регуляции TCF/LEF позвоночных». Передача сигналов WNT в развитии и заболеваниях : 225–237. дои : 10.1002/9781118444122.ch17 . ISBN 9781118444122 .
- ^ Ван де Ветеринг, М; Кастроп, Дж; Коринек, В; Клеверс, Х. (март 1996 г.). «Обширный альтернативный сплайсинг и использование двойного промотора позволяют генерировать изоформы белка Tcf-1 со свойствами дифференциального контроля транскрипции» . Молекулярная и клеточная биология . 16 (3): 745–52. дои : 10.1128/MCB.16.3.745 . ПМК 231054 . ПМИД 8622675 .
- ^ Ованес, К; Ли, ТВ; Мунгия, Дж. Э.; Труонг, Т; Милованович, Т; Лоуренс Марш, Дж; Холкомб, РФ; Уотерман, М.Л. (май 2001 г.). «Бета-катенин-чувствительные изоформы лимфоидного энхансерного фактора-1 избирательно экспрессируются при раке толстой кишки». Природная генетика . 28 (1): 53–7. дои : 10.1038/ng0501-53 . ПМИД 11326276 . S2CID 28974522 .
- ^ Лю, Ф; ван ден Брук, О; Дестре, О; Хопплер, С. (декабрь 2005 г.). «Особая роль генов Xenopus Tcf/Lef в опосредовании специфических ответов на передачу сигналов Wnt/бета-катенина в развитии мезодермы» . Разработка . 132 (24): 5375–85. дои : 10.1242/dev.02152 . ПМИД 16291789 . S2CID 20515922 .
- ^ Атча, ФА; Сайед, А; Ву, Б; Ховертер, Северная Каролина; Ёкояма, штат Нью-Йорк; Тинг, Дж. Х.; Мунгия, Дж. Э.; Мангалам, HJ; Марш, Дж. Л.; Уотерман, ML (декабрь 2007 г.). «Уникальный ДНК-связывающий домен превращает факторы Т-клеток в сильные эффекторы Wnt» . Молекулярная и клеточная биология . 27 (23): 8352–63. дои : 10.1128/MCB.02132-06 . ПМК 2169181 . ПМИД 17893322 .
- ^ Равиндранат, Эй Джей; Кадиган, КМ (3 августа 2016 г.). «Роль C-зажима при колоректальном раке, связанном с Wnt» . Раки . 8 (8): 74. doi : 10.3390/cancers8080074 . ПМЦ 4999783 . ПМИД 27527215 .
- ^ Рамакришнан, AB; Кэдиган, КМ (2017). «Гены-мишени Wnt и где их найти» . F1000Исследования . 6 : 746. дои : 10.12688/f1000research.11034.1 . ПМЦ 5464219 . ПМИД 28649368 .
- ^ Хопплер, С; Кавана, CL (1 февраля 2007 г.). «Сигнализация Wnt: в основе разнообразие» . Журнал клеточной науки . 120 (Часть 3): 385–93. дои : 10.1242/jcs.03363 . ПМИД 17251379 . S2CID 30976795 .
- ^ Хопплер, Стефан; Мун, Рэндалл Т. (2014). Передача сигналов Wnt в развитии и заболеваниях: молекулярные механизмы и биологические функции . Хобокен, Нью-Джерси. ISBN 9781118444122 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ ДасГупта, Р; Фукс, Э. (октябрь 1999 г.). «Множественные роли активированных транскрипционных комплексов LEF/TCF во время развития и дифференцировки волосяных фолликулов». Разработка . 126 (20): 4557–68. дои : 10.1242/dev.126.20.4557 . ПМИД 10498690 .
- ^ Меррилл, Би Джей; Гат, У; ДасГупта, Р; Фукс, Э. (1 июля 2001 г.). «Tcf3 и Lef1 регулируют дифференцировку мультипотентных стволовых клеток кожи» . Гены и развитие . 15 (13): 1688–705. дои : 10.1101/gad.891401 . ПМК 312726 . ПМИД 11445543 .
- ^ Майер, Клаус-Дитер; Магон де ла Жикле, Суизик; Кроме того, Фозан; Хопплер, Стефан (май 2020 г.). «Различная экспрессия LEF/TCF при колоректальном раке человека коррелирует с измененным Wnt-регулируемым транскриптомом в метаанализе биопсий пациентов» . Гены . 11 (5): 538. doi : 10.3390/genes11050538 . ПМЦ 7288467 . ПМИД 32403323 .
- ^ Джин, Т; Лю, Л. (ноябрь 2008 г.). «Эффект сигнального пути Wnt TCF7L2 и сахарный диабет 2 типа» . Молекулярная эндокринология . 22 (11): 2383–92. дои : 10.1210/me.2008-0135 . ПМИД 18599616 .
- ^ Чен, X; Аяла, я; Шеннон, К; Фуркодо, М; Ачарья, Северная Каролина; Дженкинсон, CP; Хейккинен, С; Нортон, Л. (апрель 2018 г.). «Ген диабета и эффектор пути Wnt TCF7L2 регулирует развитие и функцию адипоцитов» . Диабет . 67 (4): 554–568. дои : 10.2337/db17-0318 . ПМК 5860863 . ПМИД 29317436 .
- CDD NCBI: cd01388 (SOX-TCF_HMG-box); человеческие белки