USF включает две основные изоформы : USF1 и USF2 . Ген USF1 расположен в участке хромосомы 1q22-q23 как у человека, так и у мышей; Ген USF2 расположен на хромосоме 19q13 у человека и хромосоме 19q7 у мышей соответственно. [9] Транскрипты USF1 и USF2 содержат 10 экзонов и могут подвергаться удалению экзона 4 во время альтернативного сплайсинга . [7] [9] С точки зрения ауторегуляции эти продукты удаления экзона 4 действуют как доминантно-негативные регуляторы и, как обнаружено, подавляют USF-зависимую экспрессию генов. [7] [9]
Сравнение USF1 и USF2 по транскриптам и функции белка
Хотя USF1 и USF2 имеют 70% общей аминокислотной последовательности в области bHLH-LZ, в их полноразмерных белках обнаружено только 40% сходства. Кроме того, USF1 и USF2 демонстрируют различное содержание белка в зависимости от типа клеток. [7] Было обнаружено, что экспрессия USF1 и USF2 увеличивается во время дифференцировки эритроидных клеток. [10] Несмотря на повсеместную экспрессию обеих изоформ, USF1 и USF2 опосредуют различные биологические процессы и функции в клетках. В то время как USF1 модулирует метаболизм, иммунный ответ и защиту тканей, USF2 в первую очередь контролирует эмбриональное развитие , функцию мозга, метаболизм железа и фертильность . [7] Структурно высококонсервативная структура bHLH-LZ на C-конце USF обеспечивает высокую специфичность связывания и способствует образованию гомодимеров USF1 или гетеродимеров USF1-USF2 для связывания ДНК. [7] [9] [11] С другой стороны, USF-специфическая область (USR) в N-концевой области облегчает ядерную транслокацию и активацию USF1.
Симметричный мотив E-box является основной мишенью транскрипционных факторов bHLH-LZ, а USF1 обладает высокой аффинностью связывания с основной последовательностью CACGTG в мотиве. [9] Активность связывания USF1 с ДНК можно модулировать посредством специфичного для типа клеток метилирования и ацетилирования ДНК по мотиву E-box или посредством посттранскрипционных модификаций белка USF1. Например, метилирование CpG центрального мотива E-box ингибирует образование комплекса USF1 с его факторами котранскрипции и, следовательно, снижает экспрессию соответствующего гена в клетках лимфосаркомы мыши . [9] Напротив, фосфорилирование USF1 митоген-активируемыми протеинкиназами p38 , протеинкиназой A или протеинкиназой C увеличивает его связывание с мотивом E-box и активирует транскрипцию гена. [9]
Митоген-активируемая протеинкиназа (MAPK) фосфорилирует остатки серина и треонина белков-субстратов и преобразует внеклеточные сигналы, индуцированные факторами роста, митогенами или цитокинами, во внутриклеточные каскады фосфорилирования, которые регулируют пролиферацию, дифференцировку клеток, реакции на стресс и апоптоз (запрограммированную гибель клеток) . [7]
Сигнальные каскады ERK и JNK
Фосфорилирование с помощью МАРК вызывает конформационные изменения белка USF и открывает для взаимодействия его ДНК-связывающий домен. Это повышенное структурное воздействие усиливает связывание ДНК и, следовательно, транскрипционную активность USF. [13]
ERK1 (также известный как MAPK3 ) и ERK2 (также известный как MAPK1 ) фосфорилируют USF1 в ответ на передачу сигналов TFG-β в гладкомышечных клетках сосудов. [13] Передача сигналов SMAD2 и SMAD3 после активации рецептора TFG-β также может взаимодействовать с путями EGFR/ERK для активации USF1, который, в свою очередь, регулирует экспрессию гена ингибитора активатора плазминогена-1 (PAI-1), важного биомаркера и предсказателя сердечно-сосудистых заболеваний. -связанная смерть [13] и маркер плохого прогноза при раке молочной железы. [7]
Казеинкиназа 2 или CK-II (CK2) представляет собой тетрамерный фермент, состоящий из двух каталитических и двух регуляторных субъединиц. В клетках поджелудочной железы CK2 фосфорилирует USF1, PDX1 и MST1, подавляя экспрессию инсулина. [14]
Трансформирующий фактор роста β 1 ( TGF бета 1 ) кодируется геном TFGB 1, который содержит E-бокс в области промотора и участвует в избыточном накоплении внеклеточного матрикса в условиях высокого уровня глюкозы. [8] Обнаружено, что сверхэкспрессия USF1 или USF2 повышает активность промотора TFGB1 в клетках эмбриональных почек человека. Однако только сверхэкспрессия USF1 приводит к увеличению секреции TGF-β1.
Тромбоспондин 1 (TSP1) участвует в развитии диабетической нефропатии . USF1/2 связывается с мотивом E-box (CAGATG) на промоторе THBS1 человека и регулирует индуцированную высоким содержанием глюкозы экспрессию TSP1 в мезангиальных клетках . [8] Было обнаружено, что сверхэкспрессия USF2 увеличивает активность промотора THBS1 и экспрессию TSP1. Результирующее увеличение экспрессии TSP1 дополнительно способствует образованию активного TGF-β. [8]
Взаимодействие между USF1 и другими факторами транскрипции, включая SP1 , PEA3 (также известный как ETV4 ) и MTF1 , также приводит к совместной регуляции транскрипции. Например, мотив лейциновой молнии USF1 вовлекает PEA3 в тройной комплекс и совместно регулирует транскрипцию BAX , регулятора апоптоза. [9] Другой мишенью, регулируемой USF1, является топоизомераза III (hTOP3⍺), которая катализирует топологические изменения ДНК, модифицирует структуры суперспиралей ДНК и увеличивает доступность хроматина для экспрессии генов. [9] Подобные взаимодействия существуют между USF1 и JMJD1C или H3K9 деметилазой , в которых молекулярные взаимодействия изменяют доступность хроматина и повышают транскрипцию ряда липогенных генов, включая FASN , ACC , ACLY и SREBP1 . [15]
Общая модель дупликации гетерохроматина во время деления клеток
Хромосомы обычно подразделяют на эухроматин и гетерохроматин с различными модификациями гистонов, уровнями уплотнения и полученными в результате паттернами экспрессии генов. Гетерохроматин представляет собой сильно конденсированный и транскрипционно репрессированный домен хроматина, который характеризуется различными комбинациями посттрансляционных модификаций гистонов. [17] Гетерохроматин необходим для стабильности генома и регуляции экспрессии генов. Однако он может распространяться на соседние участки ДНК и инактивировать экспрессию генов. [17] [18] Таким образом, граничные элементы хромосом необходимы для блокирования такого стохастического распространения гетерохроматина и поддержания стабильной экспрессии генов. [19] Было обнаружено, что USF1 и USF2 рекрутируют различные комплексы, модифицирующие гистоны, включая комплекс гистон-метилтрансферазы H3 Set1 и аргинин-3-метилтрансферазу H4 PRMT1 , причем последняя, как известно, устанавливает активные домены хроматина. [19] Связывание USF1/USF2 обеспечивает высокий уровень активации модификаций гистонов на соседних нуклеосомах и, таким образом, предотвращает распространение модификаций, подавляющих хроматин, из гетерохроматина, таких как метилирование H3K9 и K27. [19]
Модификации гистонов ДНК
Другие модификации хроматина, связанные с USF1/USF2, включают привлечение E3 убиквитинлигазы, RNF20 , для мониубиквитинирования гистона H2B . [19] Обнаружено, что потеря RNF20 вызывает распространение модификаций молчания из гетерохроматического домена размером 16 т.п.н. в локус β-глобина . [19] Более того, USF1 и USF2 могут связываться с гиперчувствительным сайтом HS4 5'-ДНКазы I и рекрутировать ацетилтрансферазу H3, PCAF , которая блокирует распространение гетерохроматина в локус β-глобина. [18]
Известно, что USF связывает промотор пируваткиназы L-типа с ДНК при высоких уровнях глюкозы и инсулина. Избыточный инсулин активирует киназы и фосфатазы, которые посттрансляционно модифицируют USF, белок 1C, связывающий регуляторные элементы стерола (SREBP1C), белок, связывающий углевод-чувствительный элемент (ChREBP) и X-рецептор печени (LXR). [11] При стимуляции инсулином USF1 и USF2 связываются с E-боксами в местах -332 и -65 в промоторной области FASN , которая кодирует синтазу жирных кислот (FAS) для липогенеза. [11]
Различные посттрансляционные модификации USF1 определяют его активность и сигнальные пути, а также могут влиять на процесс липогенеза. , опосредованного USF, вызывает внутриклеточное накопление жирных кислот и нарушает регуляцию экспрессии генов и клеточных процессов, таких как выживание опухолевых клеток. de novo Обнаружено, что аномальное увеличение синтеза жирных кислот [20]
В ответ на повышение уровня инсулина ДНК-протеинкиназа (ДНК-ПК), участвующая в восстановлении повреждений ДНК, дефосфорилируется и активируется. [11] Активная форма DNA-PK опосредованно фосфорилирует USF1 по S262 через AMP-активируемую протеинкиназу (AMPK). Фосфорилирование S262 увеличивает взаимодействие USF1 с SREBP1C вблизи регуляторного элемента стерола (SRE) и облегчает синергетическую активацию SREBP1C и транскрипцию нижестоящих липогенных генов. [11]
USF1, модифицированный фосфорилированием S262 и ацетилированием K237, также рекрутирует связанный с BGR1 (также известный как SMARCA4 ) фактор 60c (BAF60c). [15] Затем BAF60c фосфорилируется атипичной протеинкиназой C (aPKC) по S257, что позволяет ему образовывать комплекс LipoBAF на промоторах липогенных генов для регулирования структуры хроматина и транскрипции генов. [15]
Напротив, HDAC9 деацетилирует USF1 во время клеточного голодания, предотвращает рекрутирование факторов, взаимодействующих с USF1, и подавляет активацию транскрипции липогенных генов. [11]
Транскрипция USF1 претерпевает активную динамику во время клеточного мейоза, при котором мРНК USF1 сначала значительно увеличивается в течение 2-8 клеток, а затем снижается до неопределяемого уровня на стадии бластоцисты, что указывает на ее роль в активации генома эмбриона. [22] Было показано, что нокаут миРНК USF1 ставит под угрозу уровень бластоцисты и нарушает регуляцию транскриптов твист-родственного белка 2 (увеличивается), фактора дифференцировки роста-9 и фоллистатина (снижается), воздействуя на область E-box их промотор-связывающего элемента во время созревания ооцита. [22]
USF1 как инсулин-чувствительный фактор транскрипции, который становится активным в ответ на высокий уровень глюкозы, способствует трансактивации генов, участвующих в метаболизме липидов, включая печеночную липазу ( LIPC ), ядерный фактор гепатоцитов 4 альфа ( HNF4A ), аполипопротеин AI ( APOA1) , Аполипопротеин L1 ( APOL1 ) и белок, родственный гаптоглобину ( HPR ) . [25] В частности, известно, что APOL1 образует комплекс с APOA-I и HDL, способствуя клеточной аутофагии в ответ на травмы и предотвращая гломерулярные заболевания; однако вариант риска APOL1 , специфичный для подоцитов, ингибирует клеточную аутофагию и может вызвать заболевание почек. [25]
Продолжительность: 3 секунды. 0:03 Липогенез de novo в живых клетках
Раковые клетки демонстрируют ряд фенотипов, в том числе заметное увеличение аэробного гликолиза , выработку молочной кислоты (известное как эффект Варбурга) , повышенный синтез белка и ДНК, а также повышенный de novo или эндогенных синтез жирных кислот синтазой жирных кислот (FAS) . [20] ФАС синтезирует в основном пальмитат из малонил-КоА, который далее этерифицируется до триглицеридов для хранения энергии. Обычно FASN активен во время эмбриогенеза и в легких плода для производства смазки; однако физиологически он слабо экспрессируется в нераковых взрослых клетках. Напротив, аномальная сверхэкспрессия FASN обнаруживается при нескольких типах рака, включая рак молочной железы , колоректальный рак , рак предстательной железы , рак поджелудочной железы и рак яичников. [26] FASN -опосредованный синтез липидов de novo составляет более 93% триглицеридов в опухолевых клетках. [20] В частности, опухолевые клетки предпочитают гликолиз окислению для потребления энергии и перенаправляют гликолитические продукты на de novo синтез жирных кислот , чтобы поставлять липиды для производства мембран и липидирование белков для быстрой пролиферации клеток. [20] Например, , что путь PI3K-AKT обнаружено усиливается в клетках рака простаты LNCaP, стимулируя сверхэкспрессию FASN . Одновременно сверхэкспрессия синтазы жирных кислот также посттрансляционно поддерживается за счет USP2α -опосредованного снижения убиквитинирования, стабилизируя FAS для конститутивной передачи сигнала. [20] Помимо липогенеза de novo , ФАС способствует локализации VEGFR-2 в липидном рафте мембраны эндотелиальных клеток и, таким образом, усиливает ангиогенез при развитии опухоли. [26] Между тем, взаимная активация передачи сигналов FAS и ERBB2 ( HER2 ) также усиливает онкогенез, при котором амплификация ERBB2 связана с повышенной выживаемостью и пролиферацией раковых клеток и плохим прогнозом при раке молочной железы и желудка; Увеличение ERBB2, в частности, способствует 18-25% случаев рака молочной железы. [27] В клетках рака простаты и клетках промиелоцитарного лейкоза активация USF1 также достигает высокого уровня экспрессии PAI-1 и ингибирует спонтанный или индуцированный камптотецином апоптоз. [13]
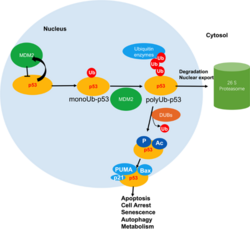
MDM2 опосредует полиубиквитинирование p53 и протеасомную деградацию.
Снижение взаимодействия USF1-p53 и повышение нестабильности p53.
Плохой прогноз рака желудка связан с низкой экспрессией USF1 и p53. [28] Среди больных раком желудка у 88% пациентов диагностируется инфекция H. pylor i, а у половины пациентов наблюдается снижение экспрессии USF1 в опухолевых тканях. Механически H. pylor i индуцирует гиперметилирование ДНК в промоторных областях USF1 и USF2 и ингибирует экспрессию. Снижение экспрессии снижает взаимодействие между USF1 и p53 при возникновении повреждения ДНК, заставляя p53 чаще связываться с E3-убиквитинлигазой HDM2 (также известной как MDM2 ) и увеличивая нестабильность p53 в раковых клетках. [28]
Семейная комбинированная гиперлипидемия (FCHL) была впервые использована для описания липидных нарушений у 47 членов родословной из Сиэтла с гиперхолестеринемией и гипертриглицеридемией. [29] Основные липидные профили FCHL характеризуются высокими уровнями холестерина/триглицеридов в сыворотке, аполипопротеина B (APOB) и ЛПНП. Генетические данные позволяют предположить, что локус, связанный с FCHL, на хромосоме человека 1q21-q23 связан с метаболическими синдромами. [30] Точное картирование этих связанных регионов идентифицирует USF1 как первый позиционно клонированный ген для FCHL и мишень для лечения FCHL. Кроме того, ядерный фактор гепатоцитов 4 альфа (HNF4A) также участвует в повышенных уровнях липидов и метаболических синдромах. Было показано, что совместные эффекты USF1 и HNF4A регулируют экспрессию аполипопротеина A-II ( APOA2 ) и аполипопротеина C-III ( APOC3 ). [30] Мутации в USF1 , HNF4A и аполипопротеинах также повышают восприимчивость пациентов к FCHL. [30] Дополнительные гены, подвергающиеся регуляции USF1 и участвующие в метаболизме глюкозы/липидов, включают аполипопротеин A5 ( APOA5 ), аполипопротеин E ( APOE ), гормон-чувствительную липазу ( LIPE ), печеночную липазу ( LIPC ), глюкокиназу ( GCK ), островково-специфическую глюкозо-специфическую липазу (LIPE). Белок, родственный каталитической субъединице 6-фосфатазы ( IGRP ), инсулин, рецептор глюкагона ( GCGR ) и АТФ-связывающий кассетный транспортер A1 ( ABCA1 ). [30]
Генрион А.А., Мартинес А., Маттеи М.Г., Кан А., Раймонжан М. (январь 1995 г.). «Структура, последовательность и хромосомное расположение гена факторов транскрипции USF2 у мышей». Геномика . 25 (1): 36–43. дои : 10.1016/0888-7543(95)80107-W . ПМИД 7774954 .
Arc.Ask3.Ru Номер скриншота №: 630c4e30ee74c4567d3dfef9127886a0__1716486960 URL1:https://arc.ask3.ru/arc/aa/63/a0/630c4e30ee74c4567d3dfef9127886a0.html Заголовок, (Title) документа по адресу, URL1: USF1 - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)