Белковый домен WRKY
| ВРКИ | |||
|---|---|---|---|
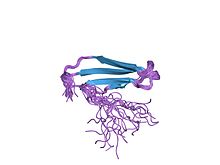 структура решения c-терминального домена wrky atwrky4 | |||
| Идентификаторы | |||
| Символ | ВРКИ | ||
| Пфам | PF03106 | ||
| Пфам Клан | CL0274 | ||
| ИнтерПро | ИПР003657 | ||
| |||
Домен WRKY находится в семействе транскрипционных факторов WRKY , классе транскрипционных факторов. [1] Домен WRKY встречается почти исключительно у растений, хотя гены WRKY, по-видимому, присутствуют у некоторых дипломонад , социальных амеб и других амебозой , а также грибов incertae sedis . Они отсутствуют у других нерастительных видов. Факторы транскрипции WRKY были важной областью исследований растений в течение последних 20 лет. [2] ДНК-связывающий домен WRKY распознает W-box (T)TGAC(C/T) (и варианты этой последовательности) цис-регуляторный элемент .
Структура
[ редактировать ]Транскрипционные факторы WRKY содержат один или два белковых домена WRKY. Белковый домен WRKY представляет собой ДНК-связывающий домен длиной от 60 до 70 аминокислот. Домен характеризуется высококонсервативным основным мотивом WRKYGQK и областью цинковых пальцев. Домен цинкового пальца цистеина 22-23 и гистидина встречается как тип CX 4-5 CX любой HXH или CX 7 CX 23 HXC, где X может быть аминокислотой . [3] Цинковый палец связывает Zn +2 ион, который необходим для функции белка. [4] Хотя WRKYGQK высококонсервативен в большинстве доменов WRKY, зарегистрированы вариации основной последовательности. [2] [5] Часто встречающимся вариантом основной последовательности является WRKYGKK, который присутствует у большинства видов растений. [2] [3] [5] [6] [7]
Структура домена белка WRKY была впервые определена в 2005 году с помощью ядерного магнитного резонанса (ЯМР), а затем с помощью кристаллографии . [4] [8] Белковый домен WRKY имеет глобулярную форму, состоящую из пяти антипараллельных β-цепей . Основной мотив WRKYGQK находится на второй β-цепи. [8] Восемнадцать аминокислот высококонсервативны в белковом домене WRKY, включая основной мотив, цистеины и гистидины, связывающие цинковые пальцы, а также триаду, образующую солевой мостик DWK. [8] Триада состоит из консервативного триптофана (W) основного мотива, а также аспарагиновой кислоты (D) на четыре аминокислоты выше и лизина (K) на 29 аминокислот ниже него, стабилизируя весь домен. [8] Пять аминокислот третьей β-цепи (PRSYY) также хорошо консервативны в домене WRKY. [8] Важно отметить, что гены WRKY содержат консервативный интрон в домене WRKY, который находится в месте, кодирующем PR аминокислотной последовательности PRSYY. [3] тем самым объясняя сохранение этого мотива.
Взаимодействие WRKY-ДНК
[ редактировать ]Домен WRKY образует уникальную клиновидную структуру, которая перпендикулярно входит в большую бороздку цепи ДНК. [9] Белковые домены WRKY взаимодействуют с цис-элементом (T/A)TGAC(T/A), также называемым W-боксом. [1] [10] [11] Недавние данные свидетельствуют о том, что ядро GAC W-бокса является основной мишенью домена WRKY, а фланкирующие последовательности помогают диктовать взаимодействие ДНК с очень специфическими белками WRKY. [12] Остатки RKYGQK корового мотива и дополнительные остатки аргинина и лизина домена WRKY отвечают за взаимодействие с фосфатным остовом семи последовательных пар оснований ДНК, включая ядро GAC. [9] [12] Замена триптофана, тирозина или лизина мотива WRKYGQK на аланин полностью устраняет связывание ДНК. [8] [13] что указывает на то, что эти аминокислоты необходимы для распознавания элемента W-box. Хотя это и не обязательно, замена аргинина, глицина или глутамина WRKYGQK на аланин снижает связывание ДНК с W-боксом. [8] [13] В целом, эти сложные взаимодействия белкового домена WRKY и ДНК приводят к активации генов, необходимых для многочисленных аспектов развития и защиты растений.
Внешние ссылки
[ редактировать ]- Семейство WRKY в PlantTFDB: база данных факторов транскрипции растений
- Семейство транскрипционных факторов WRKY на информационном ресурсе Arabidopsis
- Лаборатория Раштона
- Лаборатория Сомссих
- Лаборатория Шэнь
- Список публикаций Сомссича, связанных с WRKY
- Юлгем Лаборатория
Ссылки
[ редактировать ]- ^ Jump up to: а б Раштон П.Дж., Торрес Дж.Т., Парниске М., Вернерт П., Халброк К., Сомссич И.Е. (октябрь 1996 г.). «Взаимодействие элиситор-индуцированных ДНК-связывающих белков с элиситорными элементами ответа в промоторах генов PR1 петрушки» . Журнал ЭМБО . 15 (20): 5690–700. дои : 10.1002/j.1460-2075.1996.tb00953.x . ПМЦ 452313 . ПМИД 8896462 .
- ^ Jump up to: а б с Шлуттенхофер С., Юань Л. (февраль 2015 г.). «Регуляция специализированного метаболизма факторами транскрипции WRKY» . Физиология растений . 167 (2): 295–306. дои : 10.1104/стр.114.251769 . ПМЦ 4326757 . ПМИД 25501946 .
- ^ Jump up to: а б с Юлгем Т., Раштон П.Дж., Робацек С., Сомссич И.Е. (май 2000 г.). «Суперсемейство растительных транскрипционных факторов WRKY». Тенденции в науке о растениях . 5 (5): 199–206. дои : 10.1016/s1360-1385(00)01600-9 . ПМИД 10785665 .
- ^ Jump up to: а б Ямасаки К., Кигава Т., Иноуэ М., Татено М., Ямасаки Т., Ябуки Т. и др. (март 2005 г.). «Структура раствора ДНК-связывающего домена Arabidopsis WRKY» . Растительная клетка . 17 (3): 944–56. дои : 10.1105/tpc.104.026435 . ПМЦ 1069710 . ПМИД 15705956 .
- ^ Jump up to: а б Чжан Ю, Ван Л (январь 2005 г.). «Суперсемейство транскрипционных факторов WRKY: его происхождение у эукариот и распространение у растений» . Эволюционная биология BMC . 5 :1. дои : 10.1186/1471-2148-5-1 . ПМК 544883 . ПМИД 15629062 .
- ^ Сонг Х, Ван П, Нан З, Ван Икс (2014). «Гены транскрипционного фактора WRKY у Lotus japonicus» . Международный журнал геномики . 2014 : 420128. дои : 10.1155/2014/420128 . ПМЦ 3976811 . ПМИД 24745006 .
- ^ Сюн В, Сюй Икс, Чжан Л, Ву П, Чэнь Ю, Ли М, Цзян Х, Ву Г (июль 2013 г.). «Полногеномный анализ семейства генов WRKY физического ореха (Jaтрофа curcas L.)». Джин . 524 (2): 124–32. дои : 10.1016/j.gene.2013.04.047 . ПМИД 23644253 .
- ^ Jump up to: а б с д и ж г Дуань М.Р., Нань Дж., Лян Ю.Х., Мао П., Лу Л., Ли Л., Вэй С., Лай Л., Ли Ю., Су XD (2007). «Механизм связывания ДНК, выявленный с помощью кристаллической структуры белка WRKY1 Arabidopsis thaliana с высоким разрешением» . Исследования нуклеиновых кислот . 35 (4): 1145–54. дои : 10.1093/нар/gkm001 . ПМК 1851648 . ПМИД 17264121 .
- ^ Jump up to: а б Ямасаки К., Кигава Т., Ватанабэ С., Иноуэ М., Ямасаки Т., Секи М., Шинозаки К., Ёкояма С. (март 2012 г.). «Структурная основа распознавания последовательности ДНК транскрипционным фактором Arabidopsis WRKY» . Журнал биологической химии . 287 (10): 7683–91. дои : 10.1074/jbc.M111.279844 . ПМЦ 3293589 . ПМИД 22219184 .
- ^ Юлгем Т., Раштон П.Дж., Шмельцер Э., Халброк К., Сомссич И.Е. (сентябрь 1999 г.). «Ранние ядерные события в передаче сигналов защиты растений: быстрая активация генов факторами транскрипции WRKY» . Журнал ЭМБО . 18 (17): 4689–99. дои : 10.1093/emboj/18.17.4689 . ПМЦ 1171542 . ПМИД 10469648 .
- ^ де Патер С., Греко В., Фам К., Мемилинк Дж., Кийне Дж. (декабрь 1996 г.). «Характеристика цинк-зависимого активатора транскрипции Arabidopsis» . Исследования нуклеиновых кислот . 24 (23): 4624–31. дои : 10.1093/нар/24.23.4624 . ПМК 146317 . ПМИД 8972846 .
- ^ Jump up to: а б Брэнд Л.Х., Фишер Н.М., Хартер К., Кольбахер О., Ванке Д. (ноябрь 2013 г.). «Выяснение эволюционно консервативных особенностей связывания ДНК факторов транскрипции WRKY с помощью молекулярной динамики и анализов связывания in vitro» . Исследования нуклеиновых кислот . 41 (21): 9764–78. дои : 10.1093/nar/gkt732 . ПМЦ 3834811 . ПМИД 23975197 .
- ^ Jump up to: а б Маео К., Хаяси С., Кодзима-Сузуки Х., Мориками А., Накамура К. (ноябрь 2001 г.). «Роль консервативных остатков домена WRKY в ДНК-связывании белков семейства WRKY табака» . Бионауки, биотехнологии и биохимия . 65 (11): 2428–36. дои : 10.1271/bbb.65.2428 . ПМИД 11791715 . S2CID 22671192 .