МиоД
| МИОД1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | MYOD1 , MYF3, MYOD, PUM, bHLHc1, миогенная дифференцировка 1, MYODRIF | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 159970 ; МГИ : 97275 ; Гомологен : 7857 ; Генные карты : MYOD1 ; ОМА : MYOD1 – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





MyoD , также известный как белок 1, определяющий миобласты . [5] представляет собой белок животных, который играет важную роль в регуляции мышечной дифференцировки . MyoD, обнаруженный в лаборатории Гарольда М. Вайнтрауба , [6] принадлежит к семейству белков, известных как миогенные регуляторные факторы (MRF). [7] Эти bHLH (основная спираль-петля-спираль) факторы транскрипции действуют последовательно при миогенной дифференцировке. Члены семейства MRF позвоночных включают MyoD1, Myf5 , миогенин и MRF4 (Myf6). У беспозвоночных животных обычно обнаруживается один белок MyoD.
MyoD является одним из самых ранних маркеров миогенной активности. MyoD экспрессируется на чрезвычайно низких и практически необнаружимых уровнях в покоящихся сателлитных клетках , но экспрессия MyoD активируется в ответ на физическую нагрузку или повреждение мышечной ткани. Эффект MyoD на сателлитные клетки зависит от дозы; Высокая экспрессия MyoD подавляет обновление клеток, способствует терминальной дифференцировке и может индуцировать апоптоз. Хотя MyoD отмечает детерминацию миобластов, развитие мышц не прекращается резко у мутантов мышей, лишенных гена MyoD. Вероятно, это связано с функциональной избыточностью Myf5 и/или Mrf4. Тем не менее, комбинация MyoD и Myf5 жизненно важна для успеха миогенеза . [8] [9]
История
[ редактировать ]MyoD был клонирован с помощью функционального анализа формирования мышц, о котором Дэвис, Вайнтрауб и Лассар опубликовали в журнале Cell в 1987 году. Впервые он был описан как ядерный фосфопротеин в 1988 году Тапскоттом, Дэвисом, Тайером, Ченгом, Вайнтраубом и Лассаром в журнале Science . Исследователи экспрессировали комплементарную ДНК (кДНК) мышиного белка MyoD в различных клеточных линиях ( фибробластах и адипобластах ) и обнаружили, что MyoD превращает их в миогенные клетки. [6] [10] В следующем году та же исследовательская группа провела несколько тестов, чтобы определить как структуру, так и функцию белка, подтвердив свое первоначальное предположение о том, что активный центр белка состоит из спирали-петли-спирали (теперь называемой основной спиралью-петлей-спиралью ) для димеризация и основной сайт выше этой области bHLH облегчают связывание ДНК только после того, как она становится димером белка . [11] С тех пор MyoD стала активной областью исследований, поскольку о многих аспектах ее функции до сих пор известно относительно мало.
Функция
[ редактировать ]Функция MyoD в развитии состоит в том, чтобы присоединить клетки мезодермы к линии скелетных миобластов, а затем регулировать это продолжающееся состояние. MyoD также может регулировать восстановление мышц. MyoD мРНК Сообщается также, что уровни повышаются при старении скелетных мышц.
Одним из основных действий MyoD является удаление клеток из клеточного цикла (остановка пролиферации для остановки терминального клеточного цикла в дифференцированных миоцитах) путем усиления транскрипции р21 и миогенина . MyoD ингибируется циклинзависимыми киназами ( CDK ). CDK, в свою очередь, ингибируются p21. Таким образом, MyoD усиливает свою собственную активность в клетке по принципу прямой связи.
Устойчивая экспрессия MyoD необходима для сохранения экспрессии генов, связанных с мышцами. [12]
MyoD также является важным эффектором фенотипа быстрых мышечных волокон (типы IIA, IIX и IIB). [13] [14]
Механизмы
[ редактировать ]MyoD является фактором транскрипции и может также управлять ремоделированием хроматина посредством связывания с мотивом ДНК, известным как E-box . Известно, что MyoD взаимодействует с сотнями промоторов мышечных генов и обеспечивает пролиферацию миобластов . Хотя это и не до конца понятно, в настоящее время считается, что MyoD функционирует как основной контроллер миогенеза в ассоциации включения/выключения, опосредованной фосфорилированием KAP1 (KRAB [Krüppel-like Associated Box]-ассоциированный белок 1) . [15] KAP1 локализован в генах, связанных с мышцами, в миобластах вместе с MyoD и Mef2 (фактором усиления транскрипции миоцитов). Здесь он служит каркасом и рекрутирует коактиваторы p300 и LSD1 , в дополнение к нескольким корепрессорам, которые включают G9a и гистондеацетилазу HDAC1. Последствием этого рекрутирования коактиватора/корепрессора является подавление промоции областей мышечных генов. Когда киназа MSK1 фосфорилирует KAP1, корепрессоры, ранее связанные с каркасом, высвобождаются, позволяя MyoD и Mef2 активировать транскрипцию. [16]
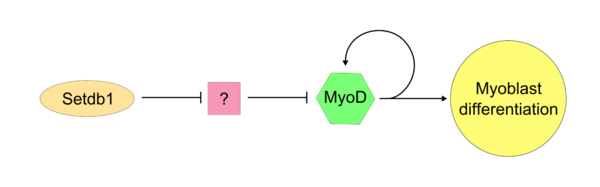
Как только «главный контроллер» MyoD становится активным, SETDB1 необходим для поддержания экспрессии MyoD внутри клетки. Setdb1, по-видимому, необходим для поддержания как экспрессии MyoD, так и генов, специфичных для мышечных тканей, поскольку снижение экспрессии Setdb1 приводит к серьезной задержке дифференцировки и детерминации миобластов. [17] В миобластах с истощением Setdb1, обработанных экзогенным MyoD, дифференцировка миобластов успешно восстанавливается. В одной модели действия Setdb1 на MyoD Setdb1 подавляет ингибитор MyoD. Этот неопознанный ингибитор, вероятно, действует конкурентно против MyoD во время типичной клеточной пролиферации. Доказательством этой модели является то, что снижение Setdb1 приводит к прямому ингибированию дифференцировки миобластов, что может быть вызвано высвобождением неизвестного ингибитора MyoD.
Также было показано, что MyoD действует совместно с геном-супрессором опухоли , ретинобластомой (pRb), вызывая остановку клеточного цикла в терминально дифференцированных миобластах. [18] Это осуществляется посредством регуляции циклина , циклина D1 . Арест клеточного цикла (при котором миобласты указывают на завершение миогенеза) зависит от непрерывной и стабильной репрессии циклина D1. И MyoD, и pRb необходимы для репрессии циклина D1, но вместо того, чтобы действовать непосредственно на циклин D1, они действуют на Fra-1, который находится непосредственно перед циклином D1. MyoD и pRb необходимы для репрессии Fra-1 (и, следовательно, циклина D1), поскольку ни MyoD, ни pRb сами по себе недостаточны для индукции репрессии циклина D1 и, следовательно, остановки клеточного цикла. В интронном энхансере Fra-1 обнаружено два консервативных сайта связывания MyoD. Существует совместное действие MyoD и pRb на интронный энхансер Fra-1, который подавляет энхансер, тем самым подавляя циклин D1 и в конечном итоге приводя к остановке клеточного цикла терминально дифференцированных миобластов. [19]
Передача сигналов Wnt может влиять на MyoD
[ редактировать ]Было показано, что передача сигналов Wnt из соседних тканей индуцирует клетки в сомитах, которые получают эти сигналы Wnt, экспрессировать Pax3 и Pax7 в дополнение к миогенным регуляторным факторам , включая Myf5 и MyoD. В частности, Wnt3a может напрямую индуцировать экспрессию MyoD посредством взаимодействий цис-элемента с дистальным энхансером и ответным элементом Wnt . [20] Wnt1 из дорсальной части нервной трубки и Wnt6/ Wnt7a из поверхностной эктодермы также участвуют в стимулировании миогенеза в сомитах; последние сигналы могут действовать преимущественно через Миод.
В типичных мышцах взрослого человека в состоянии покоя (отсутствие физиологического стресса) экспрессируются специфические белки семейства Wnt: Wnt5a , Wnt5b, Wnt7a и Wnt4 . Когда мышца повреждается (что требует регенерации), экспрессия Wnt5a, Wnt5b и Wnt7a увеличивается. Когда мышца завершает восстановление, количество Wnt7b и Wnt3a также увеличивается. Такое формирование паттерна экспрессии сигналов Wnt при восстановлении мышечных клеток индуцирует дифференцировку клеток-предшественников, что уменьшает количество доступных сателлитных клеток. Wnt играет решающую роль в регуляции сателлитных клеток, старении и регенерации скелетных мышц. Известно, что Wnts активируют экспрессию Myf5 и MyoD с помощью Wnt1 и Wnt7a. Wnt4, Wnt5 и Wnt6 повышают экспрессию обоих регуляторных факторов, но на более тонком уровне. Кроме того, MyoD увеличивает уровень Wnt3a, когда миобласты подвергаются дифференцировке. Активируется ли MyoD с помощью Wnt посредством прямого воздействия на цис-регуляцию или посредством непрямых физиологических путей, еще предстоит выяснить. [21]
Коактиваторы и репрессоры
[ редактировать ]IFRD1 является положительным кофактором MyoD, поскольку он сотрудничает с MyoD при индукции транскрипционной активности MEF2C (путем вытеснения HDAC4 из MEF2C); более того, IFRD1 также подавляет транскрипционную активность NF-κB , который, как известно, ингибирует накопление мРНК MyoD. [22] [23]
NFATc1 является фактором транскрипции, который регулирует состав типов волокон, и переход от быстрого сокращения к медленному в результате аэробных упражнений требует экспрессии NFATc1. Экспрессия MyoD является ключевым фактором транскрипции в быстросокращающихся волокнах, который ингибируется NFATc1 в окислительных типах волокон. NFATc1 ингибирует MyoD посредством физического взаимодействия с N-концевым доменом активации MyoD, что приводит к ингибированию рекрутирования необходимого коактиватора транскрипции p300 . NFATc1 физически нарушает взаимодействие между MyoD и p300. Это устанавливает молекулярный механизм, с помощью которого типы волокон изменяются in vivo в результате физических упражнений с противоположными ролями для NFATc1 и MyoD. NFATc1 контролирует этот баланс путем физического ингибирования MyoD в медленносокращающихся мышечных волокнах. [24]
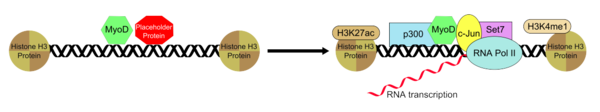
Гистондеацетилтрансфераза p300 взаимодействует с MyoD во взаимодействии, которое важно для образования мышечных трубочек из фибробластов, опосредованного MyoD. Рекрутирование р300 является лимитирующим процессом превращения фибробластов в мышечные трубочки. [25] Известно также, что помимо p300 MyoD рекрутирует Set7, H3K4me1 , H3K27ac и RNAP II к энхансеру, который связан с ним, и это позволяет активировать мышечный ген, специфичный для состояния и установленный рекрутированием MyoD. Однако эндогенный p300 необходим для функционирования MyoD, действуя как важный коактиватор.MyoD ассоциативно связывается с областью энхансера вместе с «предполагаемым пионерским фактором», который помогает установить и поддерживать их обоих в специфической и неактивной конформации. После удаления или инактивации белка-заполнителя, связанного с энхансером, разрешается привлечение дополнительной группы транскрипционных факторов, которые помогают положительно регулировать активность энхансера, и это приводит к тому, что комплекс MyoD-фактор транскрипции-энхансер переходит в транскрипционно активное состояние. .
Взаимодействия
[ редактировать ]Было показано, что MyoD взаимодействует с:
- Си-джун , [26]
- CREB-связывающий белок , [27] [28]
- CSRP3 , [29]
- Циклинзависимая киназа 4 , [30] [31]
- Ингибитор циклинзависимой киназы 1С , [32]
- ЭП300 , [28] [33]
- HDAC1 , [34] [35]
- ИД1 , [36] [37] [38] [39] [40] [41]
- ИД2 , [37]
- МДФИ , [42]
- НЕ [43]
- белок ретинобластомы , [35] [44]
- Ретиноид X-рецептор альфа [45]
- СТАТ3 , [46] и
- ТКФ3 . [37] [47]
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Ensembl выпуск 89: ENSG00000129152 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000009471 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «P15172 (MYOD1_ЧЕЛОВЕК)» . ЮниПротКБ . Проверено 17 июля 2019 г.
- ^ Jump up to: а б Дэвис Р.Л., Вайнтрауб Х., Лассар А.Б. (декабрь 1987 г.). «Экспрессия одной трансфицированной кДНК превращает фибробласты в миобласты». Клетка . 51 (6): 987–1000. дои : 10.1016/0092-8674(87)90585-X . ПМИД 3690668 . S2CID 37741454 .
- ^ «Ген Энтрез: миогенная дифференцировка 1 MYOD1» .
- ^ Рудницкий М.А., Шнегельсберг П.Н., Стед Р.Х., Браун Т., Арнольд Х.Х., Джениш Р. (декабрь 1993 г.). «MyoD или Myf-5 необходим для формирования скелетных мышц». Клетка . 75 (7): 1351–1359. дои : 10.1016/0092-8674(93)90621-В . ПМИД 8269513 . S2CID 27322641 .
- ^ Хинитс Ю., Уильямс В.К., Свитман Д., Донн Т.М., Ма Т.П., Моенс С.Б. и др. (октябрь 2011 г.). «Дефектное развитие черепного скелета, личиночная летальность и гаплонедостаточность у рыбок данио-мутанта Myod» . Дев. Биол . 358 (1): 102–112. дои : 10.1016/j.ydbio.2011.07.015 . ПМК 3360969 . ПМИД 21798255 .
- ^ Тапскотт С.Дж., Дэвис Р.Л., Тайер М.Дж., Ченг П.Ф., Вайнтрауб Х., Лассар А.Б. (октябрь 1988 г.). «MyoD1: ядерный фосфопротеин, требующий области гомологии Myc для преобразования фибробластов в миобласты». Наука . 242 (4877): 405–511. Бибкод : 1988Sci...242..405T . дои : 10.1126/science.3175662 . ПМИД 3175662 .
- ^ Дэвис Р.Л., Ченг П.Ф., Лассар А.Б., Тайер М., Тапскотт С., Вайнтрауб Х. (1989). «MyoD и ахета-щиток: 4-5 аминокислот отличают миогенез от нейрогенеза». Симпозиум принцессы Такамацу . 20 : 267–278. ПМИД 2562185 .
- ^ Фонг А.П., Тапскотт С.Дж. (октябрь 2013 г.). «Программирование и перепрограммирование скелетных мышц» . Текущее мнение в области генетики и развития . 23 (5): 568–573. дои : 10.1016/j.где.2013.05.002 . ПМЦ 3775946 . ПМИД 23756045 .
- ^ Хьюз С.М., Койши К., Рудницки М., Мэггс А.М. (январь 1997 г.). «Белок MyoD по-разному накапливается в быстрых и медленных волокнах скелетных мышц и необходим для нормального баланса типов волокон у грызунов» . Мех Дев . 61 (1–2): 151–163. дои : 10.1016/S0925-4773(96)00631-4 . ПМИД 9076685 . S2CID 17769090 .
- ^ Элерс ML, Celona B, Black BL (сентябрь 2014 г.). «NFATc1 контролирует тип волокон скелетных мышц и является негативным регулятором активности MyoD» . Отчеты по ячейкам . 8 (6): 1639–1648. дои : 10.1016/j.celrep.2014.08.035 . ПМК 4180018 . ПМИД 25242327 .
- ^ Сингх К., Кассано М., Планета Е, Себастьян С., Джанг С.М., Сохи Дж. и др. (март 2015 г.). «Переключатель фосфорилирования KAP1 контролирует функцию MyoD во время дифференцировки скелетных мышц» . Гены и развитие . 29 (5): 513–525. дои : 10.1101/gad.254532.114 . ПМК 4358404 . ПМИД 25737281 .
- ^ Бэкингем М., Ригби П.В. (февраль 2014 г.). «Регуляторные сети генов и механизмы транскрипции, контролирующие миогенез» . Развивающая клетка . 28 (3): 225–238. дои : 10.1016/j.devcel.2013.12.020 . ПМИД 24525185 .
- ^ Сон YJ, Чхве Дж.Х., Ли Х (февраль 2015 г.). «Setdb1 необходим для миогенной дифференцировки клеток миобластов C2C12 посредством поддержания экспрессии MyoD» . Молекулы и клетки . 38 (4): 362–372. doi : 10.14348/molcells.2015.2291 . ПМК 4400312 . ПМИД 25715926 .
- ^ Раджаби Х.Н., Такахаши С., Юэн М.Е. (август 2014 г.). «Белок ретинобластомы и MyoD действуют вместе, вызывая репрессию Fra-1 и, в свою очередь, циклина D1 во время остановки терминального клеточного цикла, связанной с миогенезом» . Журнал биологической химии . 289 (34): 23417–23427. дои : 10.1074/jbc.M113.532572 . ПМК 4156083 . ПМИД 25006242 .
- ^ Милевска М., Грабец К., Гжельковска-Ковальчик К. (май 2014 г.). «[Взаимодействие сигнальных путей пролиферации и дифференцировки в миогенезе]» . Достижения гигиены и экспериментальной медицины . 68 : 516–526. дои : 10.5604/17322693.1101617 . ПМИД 24864103 .
- ^ Пан Ю.К., Ван XW, Тенг Х.Ф., Ву Ю.Дж., Чанг Х.К., Чен С.Л. (февраль 2015 г.). «Сигнальные пути Wnt3a активируют экспрессию MyoD, нацеливаясь на цис-элементы внутри и снаружи его дистального энхансера» . Отчеты по биологическим наукам . 35 (2): 1–12. дои : 10.1042/BSR20140177 . ПМК 4370097 . ПМИД 25651906 .
- ^ Мотохаси Н., Асакура А. (январь 2014 г.). «Неоднородность и самообновление мышечных сателлитных клеток» . Границы клеточной биологии и биологии развития . 2 (1): 1. doi : 10.3389/fcell.2014.00001 . ПМК 4206996 . ПМИД 25364710 .
- ^ Микели Л., Леонарди Л., Конти Ф., Буанне П., Кану Н., Карузо М. и др. (март 2005 г.). «PC4 коактивирует MyoD, снимая опосредованное гистондеацетилазой 4 ингибирование фактора 2C энхансера миоцитов» . Мол. Клетка. Биол . 25 (6): 2242–59. дои : 10.1128/MCB.25.6.2242-2259.2005 . ПМЦ 1061592 . ПМИД 15743821 .
- ^ Микели Л., Леонарди Л., Конти Ф., Мареска Г., Колазингари С., Маттеи Е. и др. (февраль 2011 г.). «PC4/Tis7/IFRD1 стимулирует регенерацию скелетных мышц и участвует в дифференцировке миобластов в качестве регулятора MyoD и NF-kappaB» . Ж. Биол. Хим . 286 (7): 5691–707. дои : 10.1074/jbc.M110.162842 . ПМК 3037682 . ПМИД 21127072 .
- ^ Элерс ML, Celona B, Black BL (сентябрь 2014 г.). «NFATc1 контролирует тип волокон скелетных мышц и является негативным регулятором активности MyoD» . Отчеты по ячейкам . 8 (6): 1639–1648. дои : 10.1016/j.celrep.2014.08.035 . ПМК 4180018 . ПМИД 25242327 .
- ^ Сарторелли В., Хуан Дж., Хамамори Ю., Кедес Л. (февраль 1997 г.). «Молекулярные механизмы миогенной коактивации p300: прямое взаимодействие с доменом активации MyoD и с MADS-боксом MEF2C» . Молекулярная и клеточная биология . 17 (2): 1010–1026. дои : 10.1128/mcb.17.2.1010 . ПМК 231826 . ПМИД 9001254 .
- ^ Бенгал Э., Рэнсоне Л., Шарфманн Р., Дварки В.Дж., Тапскотт С.Дж., Вайнтрауб Х. и др. (февраль 1992 г.). «Функциональный антагонизм между белками c-Jun и MyoD: прямая физическая ассоциация». Клетка . 68 (3): 507–19. дои : 10.1016/0092-8674(92)90187-h . ПМИД 1310896 . S2CID 44966899 .
- ^ Полесская А, Нагубнева И, Дюке А, Бенгал Э, Робин П, Харель-Беллан А (август 2001 г.). «Взаимодействие между ацетилированным MyoD и бромодоменом CBP и/или p300» . Мол. Клетка. Биол . 21 (16): 5312–20. дои : 10.1128/MCB.21.16.5312-5320.2001 . ПМЦ 87255 . ПМИД 11463815 .
- ^ Jump up to: а б Сарторелли В., Хуан Дж., Хамамори Ю., Кедес Л. (февраль 1997 г.). «Молекулярные механизмы миогенной коактивации p300: прямое взаимодействие с доменом активации MyoD и с MADS-боксом MEF2C» . Мол. Клетка. Биол . 17 (2): 1010–26. дои : 10.1128/mcb.17.2.1010 . ПМК 231826 . ПМИД 9001254 .
- ^ Конг Ю, Флик М.Дж., Кудла А.Дж., Конечный С.Ф. (август 1997 г.). «Мышечный белок LIM способствует миогенезу за счет усиления активности MyoD» . Мол. Клетка. Биол . 17 (8): 4750–60. дои : 10.1128/mcb.17.8.4750 . ПМК 232327 . ПМИД 9234731 .
- ^ Чжан Дж. М., Чжао X, Вэй Ц, Патерсон Б. М. (декабрь 1999 г.). «Прямое ингибирование активности киназы G (1) cdk с помощью MyoD способствует прекращению клеточного цикла миобластов и терминальной дифференцировке» . ЭМБО Дж . 18 (24): 6983–93. дои : 10.1093/emboj/18.24.6983 . ПМЦ 1171761 . ПМИД 10601020 .
- ^ Чжан Дж. М., Вэй Ц, Чжао X, Патерсон Б. М. (февраль 1999 г.). «Связь клеточного цикла и миогенеза посредством циклин-D1-зависимого взаимодействия MyoD с cdk4» . ЭМБО Дж . 18 (4): 926–33. дои : 10.1093/emboj/18.4.926 . ПМЦ 1171185 . ПМИД 10022835 .
- ^ Рейно Э.Г., Лейбович М.П., Тинтиньяк Л.А., Пелпель К., Гийе М., Лейбович С.А. (июнь 2000 г.). «Стабилизация MyoD путем прямого связывания с p57 (Kip2)» . Ж. Биол. Хим . 275 (25): 18767–76. дои : 10.1074/jbc.M907412199 . ПМИД 10764802 .
- ^ Лау П., Бейли П., Дохан Д.Х., Маскат Дж.Е. (январь 1999 г.). «Экзогенная экспрессия доминантно-негативного вектора RORalpha1 в мышечных клетках нарушает дифференцировку: RORalpha1 напрямую взаимодействует с p300 и myoD» . Нуклеиновые кислоты Рез . 27 (2): 411–20. дои : 10.1093/нар/27.2.411 . ПМК 148194 . ПМИД 9862959 .
- ^ Пури П.Л., Иецци С., Стиглер П., Чен Т.Т., Шильц Р.Л., Мускат Г.Е. и др. (октябрь 2001 г.). «Деацетилазы гистонов класса I последовательно взаимодействуют с MyoD и pRb во время скелетного миогенеза» . Мол. Клетка . 8 (4): 885–97. дои : 10.1016/s1097-2765(01)00373-2 . ПМИД 11684023 .
- ^ Jump up to: а б Мал А., Стурниоло М., Шильц Р.Л., Гош М.К., Хартер М.Л. (апрель 2001 г.). «Роль гистондеацетилазы HDAC1 в модуляции транскрипционной активности MyoD: ингибирование миогенной программы» . ЭМБО Дж . 20 (7): 1739–53. дои : 10.1093/emboj/20.7.1739 . ПМК 145490 . ПМИД 11285237 .
- ^ Гаркавцев И., Козин С.В., Чернова О., Сюй Л., Винклер Ф., Браун Е. и др. (март 2004 г.). «Белок-кандидат-супрессор опухоли ING4 регулирует рост опухоли головного мозга и ангиогенез». Природа . 428 (6980): 328–32. Бибкод : 2004Natur.428..328G . дои : 10.1038/nature02329 . ПМИД 15029197 . S2CID 4427531 .
- ^ Jump up to: а б с Ленглендс К., Инь Х, Ананд Г., Прочовник Е.В. (август 1997 г.). «Дифференциальные взаимодействия белков Id с факторами транскрипции основная спираль-петля-спираль» . Ж. Биол. Хим . 272 (32): 19785–93. дои : 10.1074/jbc.272.32.19785 . ПМИД 9242638 .
- ^ Финкель Т., Дюк Дж., Фирон Э.Р., Данг К.В., Томаселли Г.Ф. (январь 1993 г.). «Обнаружение и модуляция in vivo белок-белковых взаимодействий спираль-петля-спираль» . Ж. Биол. Хим . 268 (1): 5–8. дои : 10.1016/S0021-9258(18)54105-3 . ПМИД 8380166 .
- ^ Гупта К., Ананд Г., Инь Х, Гроув Л., Проховник Е.В. (март 1998 г.). «Mmip1: новый белок лейциновой молнии, который обращает вспять подавляющее воздействие членов семьи Mad на c-myc» . Онкоген . 16 (9): 1149–59. дои : 10.1038/sj.onc.1201634 . ПМИД 9528857 .
- ^ Маклафлин П., Элер Э., Карлайл Г., Лихт Дж.Д., Шефер Б.В. (октябрь 2002 г.). «Белок DRAL/FHL2, содержащий только LIM, взаимодействует с белком цинковых пальцев промиелоцитарного лейкоза и является его корепрессором» . Ж. Биол. Хим . 277 (40): 37045–53. дои : 10.1074/jbc.M203336200 . ПМИД 12145280 .
- ^ Линг М.Т., Чиу Ю.Т., Ли Т.К., Люнг С.К., Фунг М.К., Ван Х и др. (сентябрь 2008 г.). «Id-1 индуцирует протеасомно-зависимую деградацию белка HBX». Дж. Мол. Биол . 382 (1): 34–43. дои : 10.1016/j.jmb.2007.06.020 . ПМИД 18674781 .
- ^ Чен С.М., Краут Н., Грудин М., Вайнтрауб Х. (сентябрь 1996 г.). «I-mf, новый миогенный репрессор, взаимодействует с членами семейства MyoD» . Клетка . 86 (5): 731–41. дои : 10.1016/s0092-8674(00)80148-8 . ПМИД 8797820 . S2CID 16252710 .
- ^ Ленорман Дж.Л., Бенаюн Б., Гийе М., Вандромм М., Лейбович М.П., Лейбович С.А. (февраль 1997 г.). «Mos активирует миогенную дифференцировку, способствуя гетеродимеризации белков MyoD и E12» . Мол. Клетка. Биол . 17 (2): 584–93. дои : 10.1128/mcb.17.2.584 . ПМК 231783 . ПМИД 9001211 .
- ^ Гу В., Шнайдер Дж. В., Кондорелли Г., Каушал С., Махдави В., Надаль-Жинард Б. (февраль 1993 г.). «Взаимодействие миогенных факторов и белка ретинобластомы опосредует детерминацию и дифференцировку мышечных клеток». Клетка . 72 (3): 309–24. дои : 10.1016/0092-8674(93)90110-c . ПМИД 8381715 . S2CID 21581966 .
- ^ Фрошле А., Алрик С., Китцманн М., Карнак Г., Ораде Ф., Рошетт-Эгли С. и др. (июль 1998 г.). «Рецепторы ретиноевой кислоты и мышечные белки b-HLH: партнеры в миогенезе, индуцированном ретиноидами» . Онкоген . 16 (26): 3369–78. дои : 10.1038/sj.onc.1201894 . ПМИД 9692544 .
- ^ Катаока Ю., Мацумура И., Эзоэ С., Наката С., Такигава Э., Сато Ю. и др. (ноябрь 2003 г.). «Взаимное ингибирование между MyoD и STAT3 в регуляции роста и дифференцировки миобластов» . Ж. Биол. Хим . 278 (45): 44178–87. дои : 10.1074/jbc.M304884200 . ПМИД 12947115 .
- ^ Малеки С.Дж., Ройер К.А., Херлбурт Б.К. (июнь 1997 г.). «Гетеродимеры MyoD-E12 и гомодимеры MyoD-MyoD одинаково стабильны». Биохимия . 36 (22): 6762–7. дои : 10.1021/bi970262m . ПМИД 9184158 .
Внешние ссылки
[ редактировать ]- MyoD+Protein в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Обзор всей структурной информации, доступной в PDB для UniProt : P10085 (белок 1 для определения миобластов мыши) на PDBe-KB .
галерея PDB |
|---|
