Селективный модулятор рецепторов прогестерона
| Селективный модулятор рецепторов прогестерона | |
|---|---|
| Класс препарата | |
 Улипристала ацетат , СПРМ , который используется в качестве средства экстренной контрацепции и при лечении миомы матки . | |
| Идентификаторы классов | |
| Синонимы | МАКК |
| Использовать | Экстренная контрацепция , миома матки |
| код АТС | G03XB |
| Биологическая цель | Рецептор прогестерона |
| Химический класс | Стероидный |
| Юридический статус | |
| В Викиданных | |
Селективный модулятор рецептора прогестерона ( SPRM ) — это агент , который действует на рецептор прогестерона (PR), биологическую мишень прогестагенов , таких как прогестерон . Характеристика, которая отличает такие вещества от полных агонистов рецепторов (например, прогестерона, прогестинов ) и полных антагонистов (например, аглепристона ), заключается в том, что их действие различается в разных тканях , т.е. агонистическое действие в одних тканях и антагонистическое в других. Этот смешанный профиль действия приводит к стимуляции или ингибированию тканеспецифическим образом, что еще больше увеличивает возможность отделения нежелательных побочных эффектов от разработки синтетических кандидатов на PR-модуляторы . [ 1 ]
История
[ редактировать ]С момента открытия гормона прогестерона в середине 1930-х годов. [ 2 ] [ 3 ] и особенно после открытия его рецептора в 1970 г. [ 4 ] [ 5 ] существует значительный интерес к разработке антагонистического агента для терапевтического применения. Были синтезированы различные аналоги прогестерона , известные как прогестины , и в 1981 году был представлен первый антагонист рецепторов прогестерона под названием RU 38486 (RU 486, мифепристон ). [ 6 ] [ 7 ] Однако клинические ограничения мифепристона из-за его относительно высокого сродства к глюкокортикоидным рецепторам по сравнению с рецептором прогестерона вызвали потребность в более селективных антагонистах прогестерона для минимизации риска побочных эффектов. [ 7 ] [ 8 ] [ 9 ] В качестве вклада были разработаны так называемые селективные модуляторы рецепторов прогестерона (SPRM). Они были описаны как агенты со смешанным антагонистическим и агонистическим действием на рецепторы прогестерона тканеспецифичным образом, при этом минимизируя взаимодействие с другими стероидными рецепторами . [ 10 ] [ 11 ] В отличие от антагонистов прогестерона, смешанные агонисты-антагонисты СПРМ, благодаря присущей им агонистической активности прогестерона, оказывают отсутствующее или лишь минимальное влияние на прерывание беременности и, таким образом, идеально подходят для лечения гинекологических заболеваний без исключения возможности беременности. [ 12 ] Оба стероидные [ 13 ] и нестероидные СПРМ [ 14 ] были описаны, и наиболее яркими примерами являются асоприснил , [ 15 ] который провалил клиническое испытание фазы 3 в 2008 году, [ 16 ] и улипристала ацетат , [ 17 ] первый СПРМ на рынке (2009 г. в Европе) [ 18 ] ).
Рецептор прогестерона
[ редактировать ]Рецептор
[ редактировать ]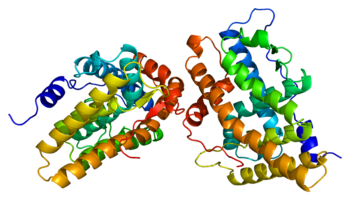
Как белок ( рецептор прогестерона рис. 1) является членом семейства лиганд-зависимых ядерных рецепторов гормонов . [ 19 ] рецептора прогестерона две основные изоформы , А и В, а также некоторые другие менее распространенные варианты сплайсинга Были идентифицированы , и все они кодируются одним и тем же геном из 8 экзонов . [ 20 ] [ 21 ] [ 22 ] [ 23 ] Как и другие стероидные ядерные рецепторы , полноразмерный белок, изоформа B, можно разделить на 4 функциональные области, а именно вариабельную N-концевую область, за которой следует высококонсервативный ДНК-связывающий домен , вариабельную шарнирную область и умеренно консервативный лигандсвязывающий домен. [ 20 ] [ 21 ] Сайт связывания лиганда, известный как домен AF2 , экспрессируется экзонами 4-8, что соответствует 253 аминокислотам , и его структура представляет большой интерес для разработки SPRM. [ 24 ] Он состоит из 10 α-спиралей (H1, H3-H12), образующих 3-слойный пучок, переплетенный 4 β-листами . H12 представляет собой конденсированную смежную единицу, состоящую из спиралей 10 и 11, которая, как предполагается, участвует в процессе связывания коактиватора . [ 25 ] Лигандсвязывающий домен рецептора находится в равновесии между двумя различными конформациями. Первая представляет собой конформацию агониста, которая способствует связыванию белков- коактиваторов , что, в свою очередь, способствует усилению регуляции транскрипции генов. [ 25 ] Вторая представляет собой антагонистическую конформацию, которая, напротив, способствует связыванию корепрессоров и, как следствие, снижению регуляции экспрессии генов. Полные агонисты, такие как прогестерон, которые проявляют агонистические свойства во всех тканях, сильно смещают конформационное равновесие в сторону агонистов. [ 25 ] И наоборот, полные антагонисты, такие как аглепристон, сильно смещают равновесие в сторону антагонистов. Наконец, общее соотношение концентраций коактиватора и корепрессора может различаться в разных типах клеток. [ 25 ]
Рецептор, связанный с G-белком
[ редактировать ]На рубеже тысячелетий стало очевидно, что активность прогестерона опосредуется не только транскрипционным фактором, но и мембраносвязанным рецептором, связанным с G-белком, обозначенным как 7TMPR . Когда рецептор активирован, он блокирует аденилатциклазу , что приводит к снижению биосинтеза внутриклеточного второго мессенджера цАМФ . [ 24 ]
Последующие механизмы
[ редактировать ]С 1990-х годов стало очевидно, что два основных изомера рецепторов, А и В, функционально различны в женской репродуктивной системе . Исследования, направленные на изучение профиля экспрессии изомеров, позволяют предположить, что изомеры экспрессируются в разных тканях в разное время на протяжении менструального цикла . [ 12 ] Было обнаружено, что PR-B активируется в строме и железистом эпителии во время фолликулярной фазы , но снижается в обеих тканях во время лютеиновой фазы . Напротив, PR-A активируется в обоих типах тканей в фолликулярной фазе и сохраняется в стромальной ткани во время поздней лютеиновой фазы. [ 12 ] Исследования показали, что активация PR-B важна для роста и развития молочной железы , тогда как PR-A играет значительную роль в нормальной репродуктивной функции и овуляции . Кроме того, исследования in vitro показали, что в идентичных условиях PR-B действует как более сильный трансактиватор репортерных генов , тогда как PR-A способен трансрепрессировать PR-B и другие стероидные рецепторы. [ 24 ] Были найдены различные причины такого разнообразия функций между изоформами. [ 26 ] Прежде всего следует упомянуть, что изоформа A рецептора прогестерона не имеет 164 N-концевых аминокислот по сравнению с изомером B, что лишает его функции активации AF-3 из-за потери вышестоящего сегмента B, в результате чего у него остаются только 2 функции активации. [ 27 ] Кроме того, исследования механизма показали разницу в рекрутировании кофакторов между изоформами. Благодаря этим функциональным различиям становится понятно, почему существует интерес к разработке препарата, который мог бы избирательно воздействовать на изоформы рецептора. Разработка SPRM в некоторых случаях была сосредоточена на этих двух разных изоформах. [ 24 ] [ 26 ] [ 27 ]
Взаимодействие SPRM с карманами связывания рецепторов
[ редактировать ]определенные взаимодействия между лигандом Было описано, что и рецептором прогестерона важны для связывания лиганда (рис. 2). Кристаллографические исследования прогестерона, связанного с его рецептором, выявили важное взаимодействие водородной связи между электроноакцепторной 3-кетогруппой прогестерона и остатками Gln 725 спирали-3 и Arg 766 спирали-5, которые удерживаются в положении структурной молекула воды. [ 26 ] Было показано, что это взаимодействие присутствует при взаимодействии с различными другими лигандами, например, мифепристоном, танапрогетом и асоприснилом, и, таким образом, его можно рассматривать как жизненно важное взаимодействие для функционирования как агонистов, так и антагонистов. [ 28 ] прогестерон и танапрогет Кроме того, было обнаружено, что образуют водородную связь с Asn 719 в спирали-3, обеспечивая возможность более высокой селективности и сродства, однако было обнаружено, что азоприснил SPRM не взаимодействует с этим остатком. [ 26 ] Несмотря на то, что полярный остаток Thr 894 находится в непосредственной близости от карбонильной группы С 20 прогестерона, между этими химическими группами не образуется никакой водородной связи. Важно отметить, что Thr 894 взаимодействует с другими лигандами. [ 26 ] [ 28 ]
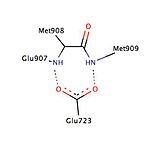

В различных исследованиях описано наличие гидрофобного кармана, называемого карманом 17α, который состоит из Leu 715 , Leu 718 , Phe 794 , Leu 797 , Met 801 и Tyr 890 и, по-видимому, обеспечивает дополнительное пространство для расширения лиганда независимо от агонизма или антагонизма. . Карман 17α, наряду с Met 756 и Met 759 в спирали-5, а также Met 909 , демонстрирует удивительную гибкость в аккомодации различных лигандов, что делает рецептор прогестерона очень адаптивным, когда дело доходит до связывания. [ 26 ] Исследования, сравнивающие конформационные изменения спирали-12, способствующие агонистическим и антагонистическим эффектам, показали важное взаимодействие водорода с остатком Glu 723 спирали-3. В неактивном состоянии Glu 723 стабилизирует конформацию спирали-12 путем образования водородной связи с аминами основной цепи в Met 908 и Met 909 . [ 26 ] [ 28 ] Когда лиганд оказывает агонистическое действие, например, оксимная группа азоприснила взаимодействует с карманом связывания агониста, тогда взаимодействие водородных связей между ранее упомянутыми остатками в спирали-12 и спирали-3 усиливается, что приводит к стыковке и привлечению коактиваторов. Однако когда антагонист, например, мифепристон, взаимодействует с этой системой водородных связей, тогда его диметиламинная группа сталкивается с Met 909 и дестабилизирует спираль-12, вызывая конформационные изменения, которые способствуют рекрутированию корепрессоров. [ 26 ] [ 28 ]
Механизм действия
[ редактировать ]Когда SPRM связываются с рецептором прогестерона, равновесие между двумя конформационными состояниями более сбалансировано и, следовательно, легче нарушается различиями в клеточной среде. В тканях, где концентрация коактиваторов выше, чем корепрессоров, избыток коактиваторов сдвигает равновесие в сторону агонистов. И наоборот, в тканях, где концентрация корепрессора выше, равновесие смещается в направлении антагониста. [ 29 ] [ 30 ] Следовательно, SPRM проявляют агонистическую активность в тканях, где преобладают коактиваторы, и антагонистическую активность, где корепрессоры находятся в избытке.
В неактивном состоянии рецептор прогестерона, как и другие стероидные рецепторы, образует комплекс, состоящий из самого себя, белков теплового шока (hsp70, hsp90) и иммунофилинов . [ 31 ] [ 32 ] Было показано, что после активации из-за связывания гормона с лиганд-связывающим карманом рецепторный комплекс диссоциирует, запуская ядерный импорт и придавая рецептору свойство димеризации (рис. 3). В ядре димер взаимодействует с элементом ответа на гормон прогестерон в ДНК, вызывая активацию или подавление активности гена. [ 33 ] [ 34 ] [ 35 ] [ 36 ] Различные исследования показали, что он влияет на экспрессию до 100 различных генов, в зависимости от изомера рецептора. [ 26 ] Под действием агонизма происходят конформационные изменения, при которых альфа-спирали 3, 4 и 12 создают поверхность стыковки для белков-коактиваторов, которые действуют как связующие факторы между рецептором и общим механизмом транскрипции. [ 37 ] [ 38 ] Однако антагонист предотвращает правильную упаковку альфа-спирали 12 против спиралей 3 и 4, нарушая способность рецептора взаимодействовать с коактиваторами, что позволяет рекрутировать корепрессоры , такие как SMRT и NCoR . [ 39 ] Из-за минимального рекрутирования корепрессоров во время связывания агониста Liu et al., 2002 постулировали, что соотношение между рекрутированием коактиваторов и корепрессоров может быть определяющим фактором, считается ли соединение агонистом, антагонистом или смешанным агонистом. антагонист. [ 40 ] Селективные модуляторы рецепторов прогестерона описаны как агенты со смешанной агонист-антагонистической активностью, и, следовательно, механизм действия должен быть обусловлен балансом этих функций.
Отношения структура-деятельность
[ редактировать ]Стероидные СПРМ
[ редактировать ]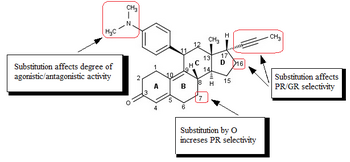
Исследования аналогов мифепристона , в основном направленные на улучшение соотношения антипрогестантной/антиглюкокортикоидной активности, [ 1 ] [ 41 ] привело к открытию SPRM. [ 42 ] Модификации 17-альфа-пропинильной группы или рядом с ней (рис. 4) в кольце D играют ключевую роль в связывании с рецептором прогестерона и/или рецептором глюкокортикоида. [ 41 ] [ 42 ] [ 43 ] Незначительные изменения в области 17-альфа приводят к образованию антипрогестинов со сниженной антиглюкокортикоидной активностью, где альфа относится к абсолютному стероидному стереодескриптору. [ 41 ] [ 42 ] [ 43 ] [ 44 ] [ 45 ] [ 46 ] По-видимому, гидрофобные 17-альфа-заместители, такие как 17-альфа-этил и 17-альфа (1'-пентинил), вызывают антипрогестагенную активность, превосходящую активность мифепристона. [ 43 ] Было также обнаружено , что замена в 17-альфа-положении, включающая фенильную группу, небольшими электроноакцепторными заместителями, такими как F и CF 3 , в пара -положении значительно увеличивает селективность в отношении глюкокортикоидного рецептора, а также эффективность получаемых соединений. Такое же замещение в орто- или мета -положении приводило к снижению селективности. Объемные заместители, такие как трет -бутил, в этой области снижают эффективность прогестерона. [ 45 ]
Имеющиеся биологические и рентгенологические данные позволяют предположить, что замещение 4-(диметиламино)фенильной группы в положении С11 (рис. 4) определяет степень агонистической и антагонистической активности. [ 41 ] [ 42 ] Небольшие заместители, такие как метил или винил, вызывают мощные агонистические свойства рецептора прогестерона. [ 42 ] тогда как замещенные фенильные производные проявляют разную степень антагонистической активности. [ 42 ] [ 43 ] [ 44 ] Имеются указания, что при замещении различными азотистыми гетероциклами наиболее агонистическими являются соединения с отчетливым максимумом отрицательного электрического потенциала в области мета- и пара -атомов арильного кольца. [ 47 ] тогда как соединения, у которых отсутствует центр электроотрицательности в этой области, обладают наибольшей антагонистической активностью. [ 26 ] [ 47 ]
Модификация основной стероидной структуры влияет на способ связывания с рецептором прогестерона. [ 45 ] [ 48 ] Было исследовано замещение C7 (рис. 4) атомом кислорода, и эти мифепристоноподобные оксастероиды показали повышенную селективность по отношению к глюкокортикоидным рецепторам, но были менее эффективными, чем мифепристон. [ 45 ] [ 49 ]
Нестероидные СПРМ
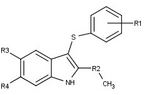
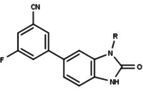
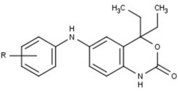
[ редактировать ]Модуляторы рецепторов прогестерона с уникальными нестероидными структурами в настоящее время находятся на ранних стадиях разработки (рис. 5-12). Сообщалось о множестве новых типов антагонистов рецепторов прогестерона с разной степенью эффективности, которые демонстрируют замечательное структурное разнообразие, которое можно увидеть в таблице ниже. Различные соединения свинца также были идентифицированы как новые агонисты рецепторов прогестерона. Их также можно посмотреть в таблице. [ 26 ]
| Антагонисты | 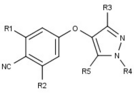 |
 |
 |
 |
 |
|---|---|---|---|---|---|
| Агонисты | 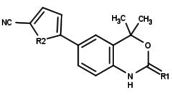 |
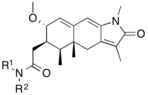 |
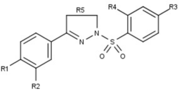 |
Наркотики
[ редактировать ]В число членов входят:
- Улипристала ацетат («Элла»)
- Асоприснил (J867; статус неизвестен)
- Телапристон (CDB-4124; Proellex, Progenta; в разработке)
СПРМ были предложены для различных гинекологических применений, таких как контрацепция и экстренная контрацепция, лечение эндометриоза , лейомиомы матки , а также в качестве гормонозамещающей терапии у женщин в постменопаузе. [ 50 ] Активность SPRM в основном опосредуется через рецептор прогестерона, где эндометрий является основной тканью-мишенью. В отличие от обычных антагонистов прогестерона, SPRM исключают возможность прерывания беременности из-за их смешанного профиля антагонистов/агонистов. Поскольку SPRM имеют низкое сродство к рецептору эстрогена, считается, что они не вызывают потерю костной массы, связанную с постменопаузой. [ 12 ] Использование СПРМ было связано с метаплазией эндометрия , что требует необходимости долгосрочной оценки безопасности. [ 12 ]
| Сложный | Химическая структура |
|---|---|
| Улипристала ацетат |  |
| Асоприснил | 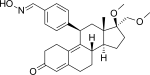 |
| Телапристон |  |
Улипристала ацетат
[ редактировать ]Улипристала ацетат (также известный как CDB-2914) [ 51 ] (Рис. 13) представляет собой 11-β арилзамещенный СПРМ, который доступен в качестве средства экстренной контрацепции в Европе с 2009 года и был одобрен FDA в 2010 году. [ 52 ] Он также продается как средство для лечения лейомиомы матки в Северной Америке и Европе. В качестве средства экстренной контрацепции улипристала ацетат оказался эффективным в течение 120 часов после незащищенного полового акта по сравнению с 72-часовой эффективностью современных средств экстренной контрацепции. [ 50 ] В постменопаузальном эндометрии это соединение, по-видимому, оказывает антагонистическое действие на рецепторы прогестерона, что указывает на его потенциальное использование в лечении менопаузы, но это еще не подтверждено. [ 12 ]
Асоприснил
[ редактировать ]Асоприснил (J867) представляет собой стероидный 11β-бензальдоксим-замещенный SPRM (рис. 14). [ 15 ] Предполагается, что геометрия его оксимной группы играет важную роль в эффективности in vitro . [ 26 ] Было предложено для лечения лейомиомы и эндометриоза. [ 53 ] и это первый SPRM в клинической разработке лечения эндометриоза, достигший продвинутой фазы. [ 54 ]
Телапристон
[ редактировать ]Телапристон (CDB-4124), также известный как Проеллекс (рис. 15), вступил во II фазу клинических исследований по лечению миомы матки в 2014 году. [ 55 ] и планирует провести II фазу клинических исследований по облегчению симптомов эндометриоза в начале 2016 года. [ 56 ] [ 57 ] Также было высказано предположение, что он оказывает химиопрофилактическое действие. [ 58 ]
Использование
[ редактировать ]SPRM разрабатываются для следующих целей:
- Асоприснил и телапристон находятся на стадии исследования (2005 г.) для лечения лейомиомы матки . [ 59 ] [ 60 ]
- Proellex завершил ряд клинических исследований по лечению эндометриоза и миомы матки. [ 61 ]
Хотя эти СПРМ были эффективны для лечения миомы матки, развитие побочных эффектов, таких как утолщение эндометрия, ограничило их применение сроком не более трех-четырех месяцев. [ 60 ]
Будущее
[ редактировать ]Благодаря своей антиглюкокортикоидной активности мифепристон исследуется на предмет его терапевтического потенциала при таких показаниях, как синдром Кушинга , болезнь Альцгеймера или психоз . Кроме того, разрабатываются СПРМ для различных гинекологических применений, включая безэстрогенную контрацепцию, лечение лейомиомы матки и эндометриоза. [ 62 ]
См. также
[ редактировать ]- Фитопрогестаген
- Селективный модулятор андрогенных рецепторов
- Селективный модулятор рецепторов эстрогена
- Селективный агонист глюкокортикоидных рецепторов
- Селективный модулятор рецепторов
Ссылки
[ редактировать ]- ^ Jump up to: а б Хвалиш К., Перес М.К., Деманно Д., Винкель С., Шуберт Г., Элгер В. (май 2005 г.). «Разработка и использование селективного модулятора рецепторов прогестерона в лечении лейомиомы и эндометриоза» . Эндокринные обзоры . 26 (3): 423–38. дои : 10.1210/er.2005-0001 . ПМИД 15857972 .
- ^ Мисрахи М., Лусфелт Х., Атгер М., Мериэль С., Зера В., Дессен П., Милгром Э. (июнь 1988 г.). «Организация всей мРНК кроличьего рецептора прогестерона, а также промотора и 5'-фланкирующей области гена» . Исследования нуклеиновых кислот . 16 (12): 5459–72. дои : 10.1093/нар/16.12.5459 . ПМК 336778 . ПМИД 3387238 .
- ^ Аллен В.М. (август 1935 г.). «Выделение кристаллического прогестина». Наука . 82 (2118): 89–93. Бибкод : 1935Sci....82...89A . дои : 10.1126/science.82.2118.89 . ПМИД 17747122 .
- ^ Каррер П., Шварценбах Г. (январь 1934 г.). «Дополнение о кислотности и восстанавливающей способности аскорбиновой кислоты». Helvetica Chimica Acta . 17 (1): 58–59. дои : 10.1002/hlca.19340170111 . ISSN 1522-2675 .
- ^ Шерман М.Р., Корвол П.Л., О'Мэлли Б.В. (ноябрь 1970 г.). «Прогестерон-связывающие компоненты яйцевода кур. I. Предварительная характеристика цитоплазматических компонентов» . Журнал биологической химии . 245 (22): 6085–96. дои : 10.1016/S0021-9258(18)62667-5 . ПМИД 5484467 .
- ^ Филибер Д., Дерэдт Р., Дойч Г. (1981). RU 38486: Мощный антиглюкокортикоид in vivo . VII Международный конгресс фармакологов. Япония: Токио.
- ^ Jump up to: а б Гройер А., Ле Бук И., Джоав И., Раданьи С., Ренуар Ж.М., Робель П., Болье Э.Э. (июнь 1985 г.). «Глюкокортикостероидный рецептор куриного яйцевода. Специфическое связывание синтетического стероида RU 486 и иммунологические исследования с антителами к прогестероновому рецептору яйцевода цыпленка» . Европейский журнал биохимии . 149 (2): 445–51. дои : 10.1111/j.1432-1033.1985.tb08945.x . ПМИД 3996417 .
- ^ Гасс Е.К., Леонхардт С.А., Нордин С.К., Эдвардс Д.П. (апрель 1998 г.). «Антагонисты RU486 и ZK98299 стимулируют связывание рецептора прогестерона с дезоксирибонуклеиновой кислотой in vitro и in vivo, но оказывают различное влияние на конформацию рецептора» . Эндокринология . 139 (4): 1905–19. дои : 10.1210/endo.139.4.5944 . ПМИД 9528977 .
- ^ Лазарус Г., Лазарус Г., Хустик Э., Сомнение Э., Агарвал М.К. (июнь 1995 г.). «Влияние антиглюкокортикоидов на стресс и шок». Анналы Нью-Йоркской академии наук . 761 (1): 276–95. Бибкод : 1995SCIENCE.761..276L . doi : 10.1111/j.1749–6632.1995.tb31384.x . ПМИД 7625726 . S2CID 40422188 .
- ^ Шпиц И.М., Хвалиш К. (август 2010 г.). «Модуляторы рецепторов прогестерона и антагонисты прогестерона в женском здоровье». Стероиды . 65 (10–11): 807–15. дои : 10.1016/S0039-128X(00)00194-X . ПМИД 11108892 . S2CID 27699000 .
- ^ Чен В., Охара Н., Ван Дж., Сюй Ц., Лю Дж., Морикава А., Сасаки Х., Ёсида С., Деманно Д.А., Чвалиш К., Маруо Т. (апрель 2006 г.). «Новый селективный модулятор рецептора прогестерона азоприснил (J867) ингибирует пролиферацию и индуцирует апоптоз в культивируемых клетках лейомиомы матки человека при отсутствии сопоставимого воздействия на клетки миометрия» . Журнал клинической эндокринологии и метаболизма . 91 (4): 1296–304. дои : 10.1210/jc.2005-2379 . ПМИД 16464945 .
- ^ Jump up to: а б с д и ж Шаббер-Бюффе Н., Медури Г., Бушар П., Шпиц И.М. (2005). «Селективные модуляторы рецепторов прогестерона и антагонисты прогестерона: механизмы действия и клиническое применение». Обновление репродукции человека . 11 (3): 293–307. дои : 10.1093/humupd/dmi002 . ПМИД 15790602 .
- ^ Элгер В., Бартли Дж., Шнайдер Б., Кауфман Г., Шуберт Г., Хвалиш К. (01.10.2000). «Эндокринная фармакологическая характеристика антагонистов прогестерона и модуляторов рецепторов прогестерона в отношении PR-агонистической и антагонистической активности». Стероиды . 65 (10–11): 713–23. дои : 10.1016/S0039-128X(00)00178-1 . ПМИД 11108882 . S2CID 46138800 .
- ^ Палмер С., Кампен К.А., Аллан Г.Ф., Рыбчински П., Хейнс-Джонсон Д., Хатчинс А., Крафт П., Киддо М., Лай М., Ломбарди Э., Педерсен П., Ходген Г., Комбс Д.В. (декабрь 2000 г.). «Лиганды нестероидных рецепторов прогестерона с беспрецедентной селективностью рецепторов». Журнал биохимии стероидов и молекулярной биологии . 75 (1): 33–42. дои : 10.1016/S0960-0760(00)00134-5 . ПМИД 11179906 . S2CID 19167595 .
- ^ Jump up to: а б ДеМанно Д., Элджер В., Гарг Р., Ли Р., Шнайдер Б., Хесс-Стумп Х., Шуберт Г., Хвалиш К. (ноябрь 2003 г.). «Асоприснил (J867): селективный модулятор рецепторов прогестерона для гинекологической терапии». Стероиды . 68 (10–13): 1019–32. doi : 10.1016/j.steroids.2003.09.008 . ПМИД 14667995 . S2CID 23074350 .
- ^ «Безопасность лечения миомы матки асоприснилом – полнотекстовый просмотр – ClinicalTrials.gov» . www.clinicaltrials.gov . Проверено 11 января 2016 г.
- ^ Доннес Дж., Васкес Ф., Томашевски Дж., Нури К., Бушар П., Фаузер Б.С., Барлоу Д.Х., Паласиос С., Доннес О., Бестель Е., Остерло И., Лумайе Е. (июнь 2014 г.). «Длительное лечение миомы матки улипристала ацетатом» . Фертильность и бесплодие . 101 (6): 1565–73.e1–18. doi : 10.1016/j.fertnstert.2014.02.008 . ПМИД 24630081 .
- ^ «Отчет об оценке Эллаоне» (PDF) . Европейское агентство лекарственных средств. 2009 . Проверено 4 апреля 2022 г.
- ^ Мангельсдорф Д.Д., Таммел С., Беато М., Херрлих П., Шютц Г., Умесоно К., Блумберг Б., Кастнер П., Марк М., Шамбон П., Эванс Р.М. (декабрь 1995 г.). «Суперсемейство ядерных рецепторов: второе десятилетие» . Клетка . 83 (6): 835–9. дои : 10.1016/0092-8674(95)90199-X . ПМК 6159888 . ПМИД 8521507 .
- ^ Jump up to: а б Коннили О.М., Максвелл Б.Л., Тофт Д.О., Шрейдер В.Т., О'Мэлли Б.В. (декабрь 1987 г.). «Формы A и B куриного рецептора прогестерона возникают в результате попеременной инициации трансляции уникальной мРНК». Связь с биохимическими и биофизическими исследованиями . 149 (2): 493–501. дои : 10.1016/0006-291X(87)90395-0 . ПМИД 3426587 .
- ^ Jump up to: а б Кастнер П., Краст А., Теркотт Б., Стропп У., Тора Л., Гронемейер Х., Шамбон П. (май 1990 г.). «Два различных промотора, регулируемых эстрогеном, генерируют транскрипты, кодирующие две функционально разные формы человеческого рецептора прогестерона А и В» . Журнал ЭМБО . 9 (5): 1603–14. дои : 10.1002/j.1460-2075.1990.tb08280.x . ПМК 551856 . ПМИД 2328727 .
- ^ Хирата С., Шода Т., Като Дж., Хоши К. (декабрь 2002 г.). «Новые изоформы мРНК рецепторов женских половых стероидных гормонов человека». Журнал биохимии стероидов и молекулярной биологии . 83 (1–5): 25–30. дои : 10.1016/S0960-0760(02)00255-8 . ПМИД 12650698 . S2CID 23588169 .
- ^ Хирата С., Шода Т., Като Дж., Хоши К. (апрель 2003 г.). «Изоформы/варианты мРНК для рецепторов половых стероидных гормонов у человека». Тенденции в эндокринологии и обмене веществ . 14 (3): 124–9. дои : 10.1016/S1043-2760(03)00028-6 . ПМИД 12670738 . S2CID 46220794 .
- ^ Jump up to: а б с д Бринтон Р.Д., Томпсон Р.Ф., Фой М.Р., Бодри М., Ван Дж., Финч С.Э., Морган Т.Е., Пайк С.Дж., Мак В.Дж., Станчик Ф.З., Нильсен Дж. (май 2008 г.). «Рецепторы прогестерона: форма и функции в мозге» . Границы нейроэндокринологии . 29 (2): 313–39. doi : 10.1016/j.yfrne.2008.02.001 . ПМЦ 2398769 . ПМИД 18374402 .
- ^ Jump up to: а б с д Эллманн С., Штихт Х., Тиль Ф., Бекманн М.В., Стрик Р., Стриссель П.Л. (август 2009 г.). «Рецепторы эстрогена и прогестерона: от молекулярных структур к клиническим мишеням» . Клеточные и молекулярные науки о жизни . 66 (15): 2405–26. дои : 10.1007/s00018-009-0017-3 . ПМЦ 11115849 . ПМИД 19333551 . S2CID 19975774 .
- ^ Jump up to: а б с д и ж г час я дж к л Виннекер Р.К., Фенсом А., Чжан П., Юдт М.Р., МакКомас CC, Унвалла Р.Дж. (август 2008 г.). «Новое поколение модуляторов рецепторов прогестерона». Стероиды . 73 (7): 689–701. doi : 10.1016/j.steroids.2008.03.005 . ПМИД 18472121 . S2CID 25094160 .
- ^ Jump up to: а б Эдвардс Д.П., Альтманн М., ДеМарзо А., Чжан Ю., Вайгель Н.Л., Бек К.А. (июнь 1995 г.). «Рецептор прогестерона и механизм действия антагонистов прогестерона». Журнал биохимии стероидов и молекулярной биологии . 53 (1–6): 449–58. дои : 10.1016/0960-0760(95)00091-д . ПМИД 7626494 . S2CID 41957428 .
- ^ Jump up to: а б с д Лушер С.Дж., Рааймейкерс ХК, Ву-Фам Д., Каземир Б., Бош Р., МакГуайр Р., Азеведо Р., Хамерсма Х., Дехеринг К., Обри А., ван Дуин М., де Влиг Дж. (июнь 2012 г.). «Рентгеновские структуры лигандсвязывающего домена рецептора прогестерона в его агонистическом состоянии выявляют различные механизмы смешанных профилей 11β-замещенных стероидов» . Журнал биологической химии . 287 (24): 20333–43. дои : 10.1074/jbc.M111.308403 . ПМК 3370215 . ПМИД 22535964 .
- ^ Джексон Т.А., Ричер Дж.К., Бэйн Д.Л., Такимото Г.С., Тунг Л., Хорвиц К.Б. (июнь 1997 г.). «Частичная агонистическая активность стероидных рецепторов, занятых антагонистами, контролируется новым коактиватором связывания шарнирного домена L7/SPA и корепрессорами N-CoR или SMRT». Молекулярная эндокринология . 11 (6): 693–705. дои : 10.1210/mend.11.6.0004 . ПМИД 9171233 . S2CID 21356045 .
- ^ Смит CL, О'Мэлли BW (февраль 2004 г.). «Функция корегулятора: ключ к пониманию тканевой специфичности селективных модуляторов рецепторов» . Эндокринные обзоры . 25 (1): 45–71. дои : 10.1210/er.2003-0023 . ПМИД 14769827 .
- ^ Оньяте С.А., Эстес П.А., Уэлч В.Дж., Нордин С.К., Эдвардс Д.П. (декабрь 1991 г.). «Доказательства того, что белок теплового шока-70, связанный с рецепторами прогестерона, не участвует в связывании рецептора с ДНК» . Молекулярная эндокринология . 5 (12): 1993–2004. doi : 10.1210/mend-5-12-1993 . ПМИД 1791844 .
- ^ Смит Д.Ф., Стенсгард Б.А., Уэлч В.Дж., Тофт Д.О. (январь 1992 г.). «Сборка рецептора прогестерона с белками теплового шока и активация рецептора являются событиями, опосредованными АТФ» . Журнал биологической химии . 267 (2): 1350–6. дои : 10.1016/S0021-9258(18)48438-4 . ПМИД 1730655 .
- ^ ДеМарзо А.М., Бек К.А., Онате С.А., Эдвардс Д.П. (январь 1991 г.). «Димеризация рецепторов прогестерона млекопитающих происходит в отсутствие ДНК и связана с высвобождением белка теплового шока массой 90 кДа» . Труды Национальной академии наук Соединенных Штатов Америки . 88 (1): 72–6. Бибкод : 1991ПНАС...88...72Д . дои : 10.1073/pnas.88.1.72 . ПМК 50750 . ПМИД 1986383 .
- ^ Гиошон-Мантель А., Лусфелт Х., Лескоп П., Сар С., Атгер М., Перро-Аппланат М., Милгром Э. (июнь 1989 г.). «Механизмы ядерной локализации рецептора прогестерона: доказательства взаимодействия между мономерами» . Клетка . 57 (7): 1147–54. дои : 10.1016/0092-8674(89)90052-4 . ПМИД 2736623 .
- ^ О'Мэлли BW, Цай MJ (февраль 1992 г.). «Молекулярные пути действия стероидных рецепторов» . Биология размножения . 46 (2): 163–7. дои : 10.1095/biolreprod46.2.163 . ПМИД 1536890 .
- ^ Багчи М.К., Цай М.Дж., О'Мэлли Б.В., Цай С.Ю. (август 1992 г.). «Анализ механизма активации генов, зависимых от рецепторов стероидных гормонов, в бесклеточных системах». Эндокринные обзоры . 13 (3): 525–35. дои : 10.1210/edrv-13-3-525 . ПМИД 1425487 .
- ^ Маккенна, штат Нью-Джерси, О'Мэлли, BW (февраль 2002 г.). «Комбинаторный контроль экспрессии генов ядерными рецепторами и корегуляторами» . Клетка . 108 (4): 465–74. дои : 10.1016/S0092-8674(02)00641-4 . ПМИД 11909518 .
- ^ Оньяте С.А., Цай С.Ю., Цай М.Дж., О'Мэлли Б.В. (ноябрь 1995 г.). «Последовательность и характеристика коактиватора суперсемейства рецепторов стероидных гормонов». Наука . 270 (5240): 1354–7. Бибкод : 1995Sci...270.1354O . дои : 10.1126/science.270.5240.1354 . ПМИД 7481822 . S2CID 28749162 .
- ^ Вагнер Б.Л., Норрис Дж.Д., Ноттс Т.А., Вайгель Н.Л., Макдоннелл Д.П. (март 1998 г.). «Ядерные корепрессоры NCoR и SMRT являются ключевыми регуляторами как лиганд-, так и 8-бромциклического АМФ-зависимой транскрипционной активности человеческого рецептора прогестерона» . Молекулярная и клеточная биология . 18 (3): 1369–78. дои : 10.1128/mcb.18.3.1369 . ПМК 108850 . ПМИД 9488452 .
- ^ Лю З, Обёф Д., Вонг Дж., Чен Дж.Д., Цай С.Ю., Цай М.Дж., О'Мэлли Б.В. (июнь 2002 г.). «Соотношения коактиватор/корепрессор модулируют PR-опосредованную транскрипцию с помощью селективного модулятора рецептора RU486» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (12): 7940–4. Бибкод : 2002PNAS...99.7940L . дои : 10.1073/pnas.122225699 . ПМК 122999 . ПМИД 12048256 .
- ^ Jump up to: а б с д Никиш К., Элджер В., Сесак Дж., Кесаварам Н., Дас Б., Гарфилд Р., Ши С.К., Амелькина О., Мейстер Р. (февраль 2013 г.). «Синтез и биологическая оценка частично фторированных антипрогестинов и мезопрогестинов». Стероиды . 78 (2): 255–67. doi : 10.1016/j.steroids.2012.09.010 . ПМИД 23178161 . S2CID 17158416 .
- ^ Jump up to: а б с д и ж Никиш К., Элджер В., Сантамма Б., Гарфилд Р., Киллин З., Амелькина О., Шнайдер Б., Мейстер Р. (декабрь 2014 г.). «Синтез и биологическая оценка 11'-имидазолил-антипрогестинов и мезопрогестинов». Стероиды . 92 : 45–55. doi : 10.1016/j.steroids.2014.08.017 . ПМИД 25174783 . S2CID 6311225 .
- ^ Jump up to: а б с д Рао П.Н., Ван З., Сесак Дж.В., Розенберг Р.С., Дженкинс DJ, Диамандис EP (октябрь 1998 г.). «Новые 11 бета-арилзамещенных стероидов проявляют как прогестагенную, так и антипрогестационную активность». Стероиды . 63 (10): 523–30. дои : 10.1016/S0039-128X(98)00060-9 . ПМИД 9800283 . S2CID 42703467 .
- ^ Jump up to: а б Вагнер Б.Л., Поллио Г., Леонхардт С., Вани М.К., Ли Д.И., Имхоф М.О., Эдвардс Д.П., Кук С.Э., Макдоннелл Д.П. (август 1996 г.). «16 альфа-замещенных аналогов антипрогестина RU486 вызывают уникальную конформацию рецептора прогестерона человека, что приводит к смешанной агонистической активности» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (16): 8739–44. Бибкод : 1996PNAS...93.8739W . дои : 10.1073/pnas.93.16.8739 . ПМЦ 38743 . ПМИД 8710941 .
- ^ Jump up to: а б с д Кан Ф.А., Гуан Дж., Джайн Н., Аллан Г., Линтон О., Танненбаум П., Чен Х, Сюй Дж., Чжу П., Ганнет Дж., Демарест К., Лундин С., Суй З. (май 2007 г.). «Параллельный синтез и исследование SAR новых оксастероидов как мощных и селективных антагонистов рецепторов прогестерона». Письма по биоорганической и медицинской химии . 17 (9): 2531–4. дои : 10.1016/j.bmcl.2007.02.013 . ПМИД 17317167 .
- ^ Кан Ф.А., Аллан Дж., Гуан Дж., Джайн Н., Линтон О., Танненбаум П., Сюй Дж., Чжу П., Ганнет Дж., Чен Икс, Демарест К., Лундин С., Суй З. (февраль 2007 г.). «Синтез и идентификация новых оксастероидов как антагонистов рецепторов прогестерона». Письма по биоорганической и медицинской химии . 17 (4): 907–10. дои : 10.1016/j.bmcl.2006.11.062 . ПМИД 17169557 .
- ^ Jump up to: а б Ревинкель Дж., Энтховен М., Гольштейн И., ван дер Рейст М., Шолтен А., ван Тилборг М., де Вейс Д., Виссе Дж., Хамерсма Х. (март 2008 г.). «11-(пиридинилфенил)стероиды - новый класс агонистов/антагонистов прогестерона смешанного профиля». Биоорганическая и медицинская химия . 16 (6): 2753–63. дои : 10.1016/j.bmc.2008.01.010 . ПМИД 18243712 .
- ^ Джайн Н., Аллан Дж., Линтон О., Танненбаум П., Чен Икс, Сюй Дж., Чжу П., Ганнет Дж., Демарест К., Лундин С., Мюррей В., Суй З. (июль 2009 г.). «Синтез и исследование SAR новых псевдостероидов как мощных и селективных антагонистов рецепторов прогестерона». Письма по биоорганической и медицинской химии . 19 (14): 3977–80. дои : 10.1016/j.bmcl.2009.01.095 . ПМИД 19217285 .
- ^ Кан Ф.А., Чен X, Джайн Н., Аллан Дж., Танненбаум П., Лундин С., Суй З. (июль 2008 г.). «Из молекулярного моделирования различных конформаций и SAR природных стероидов и неприродных 7-окса-стероидов». Письма по биоорганической и медицинской химии . 18 (13): 3687–90. дои : 10.1016/j.bmcl.2008.05.070 . ПМИД 18539027 .
- ^ Jump up to: а б Бенаджано Дж., Бастианелли С., Фаррис М., Бросенс I (июль 2014 г.). «Селективные модуляторы рецепторов прогестерона: обновление». Экспертное заключение по фармакотерапии . 15 (10): 1403–15. дои : 10.1517/14656566.2014.914494 . ПМИД 24787486 . S2CID 31746942 .
- ^ Браш В., Кочон Л., Джесам К., Мальдонадо Р., Сальватьерра А.М., Леви Д.П., Гейнер Э., Кроксато Х.Б. (сентябрь 2010 г.). «Немедленное предовуляторное введение 30 мг улипристала ацетата значительно задерживает разрыв фолликула». Репродукция человека . 25 (9): 2256–63. дои : 10.1093/humrep/deq157 . ПМИД 20634186 .
- ^ Эйкен А.Р., Трасселл Дж. (2014). «Последние достижения в области контрацепции» . Отчеты F1000Prime . 6 : 113. дои : 10.12703/p6-113 . ПМК 4251416 . ПМИД 25580267 .
- ^ Шпиц ИМ (ноябрь 2003 г.). «Антагонисты прогестерона и модуляторы рецепторов прогестерона: обзор». Стероиды . 68 (10–13): 981–93. doi : 10.1016/j.steroids.2003.08.007 . ПМИД 14667991 . S2CID 23054270 .
- ^ Линдси С.Ф., Лучано Д.Е., Лучано А.А. (сентябрь 2015 г.). «Новые методы лечения эндометриоза». Мнение экспертов о новых лекарствах . 20 (3): 449–61. дои : 10.1517/14728214.2015.1051966 . ПМИД 26050551 . S2CID 19276904 .
- ^ «Фаза 2, исследование по оценке безопасности и эффективности Proellex® (ацетат телапристона) при вагинальном введении при лечении миомы матки - полнотекстовый просмотр - ClinicalTrials.gov» . www.clinicaltrials.gov . Проверено 11 января 2016 г.
- ^ Экстайн Н., Хаас Б., Хасс, доктор медицинских наук, Пфайфер В. (август 2014 г.). «Системная терапия синдрома Кушинга» . Сиротский журнал редких заболеваний . 9 (1): 122. дои : 10.1186/s13023-014-0122-8 . ПМЦ 4237936 . ПМИД 25091295 .
- ^ Тейлор Д.К., Холтаузер К., Сегарс Дж.Х., Лепперт ПК (2015). «Последние научные достижения в исследованиях лейомиомы (миомы матки) способствуют лучшему пониманию и лечению» . F1000Исследования . 4 (F1000 Faculty Rev): 183. doi : 10.12688/f1000research.6189.1 . ПМЦ 4513689 . ПМИД 26236472 .
- ^ Пабла Б., Биссоннетт М., Конда В.Дж. (октябрь 2015 г.). «Рак толстой кишки и рецептор эпидермального фактора роста: современные парадигмы лечения, важность диеты и роль химиопрофилактики» . Всемирный журнал клинической онкологии . 6 (5): 133–41. дои : 10.5306/wjco.v6.i5.133 . ПМК 4600187 . ПМИД 26468449 .
- ^ Охара Н (2008). «Действие модуляторов рецепторов прогестерона на лейомиому матки». Клиническая и экспериментальная акушерство и гинекология . 35 (3): 165–6. ПМИД 18754282 .
- ^ Jump up to: а б Шпиц ИМ (август 2009 г.). «Клиническая полезность модуляторов рецепторов прогестерона и их влияние на эндометрий». Современное мнение в акушерстве и гинекологии . 21 (4): 318–24. дои : 10.1097/GCO.0b013e32832e07e8 . ПМИД 19602929 . S2CID 2121292 .
- ^ «Проеллекс» . ClinicalTrials.gov . Национальные институты здравоохранения США . Проверено 1 апреля 2021 г.
- ^ Бушар П., Шаббер-Бюффе Н., Фаузер BC (ноябрь 2011 г.). «Селективные модуляторы рецепторов прогестерона в репродуктивной медицине: фармакология, клиническая эффективность и безопасность» . Фертильность и бесплодие . 96 (5): 1175–89. doi : 10.1016/j.fertnstert.2011.08.021 . ПМИД 21944187 .