Мангровые заросли

— Мангровое дерево это кустарник или дерево, которое растет в основном в прибрежных соленых или солоноватой воде . Мангровые заросли растут в экваториальном климате, обычно вдоль береговой линии и приливных рек. У них есть особые приспособления для поглощения дополнительного кислорода и удаления соли, что позволяет им переносить условия, которые могут привести к гибели большинства растений. Этот термин также используется для обозначения тропической прибрежной растительности, состоящей из таких видов. Мангровые заросли таксономически разнообразны в результате конвергентной эволюции нескольких семейств растений. Они встречаются по всему миру в тропиках и субтропиках и даже в некоторых прибрежных районах с умеренным климатом , в основном между 30° северной широты и 30° южной широты, с наибольшей площадью мангровых зарослей в пределах 5° от экватора . [1] [2] Семейства мангровых растений впервые появились в период от позднего мела до палеоцена и получили широкое распространение отчасти из-за движения тектонических плит . Самые старые известные окаменелости мангровых пальм датируются 75 миллионами лет назад. [2]
Мангровые заросли — это солеустойчивые деревья, кустарники и папоротники, также называемые галофитами , которые приспособлены к жизни в суровых прибрежных условиях. Они содержат сложную систему фильтрации соли и сложную корневую систему, способную выдерживать погружение в соленую воду и воздействие волн. Они адаптированы к условиям с низким содержанием кислорода в заболоченных илах. [3] но, скорее всего, они будут процветать в верхней половине приливной зоны . [4]
Мангровый биом , часто называемый мангровым лесом или мангалом, представляет собой особую среду обитания засоленных лесов или кустарников, характеризующуюся осадочной прибрежной средой, где мелкие отложения (часто с высоким содержанием органических веществ) собираются в районах, защищенных от воздействия волн высокой энергии. Мангровые леса служат жизненно важной средой обитания для множества водных видов, предлагая уникальную экосистему, поддерживающую сложное взаимодействие морской жизни и наземной растительности. Соленые условия, переносимые различными видами мангровых зарослей, варьируются от солоноватой воды до чистой морской воды (соленость от 3 до 4%) до воды, концентрированной за счет испарения, соленость которой более чем в два раза превышает соленость океанской морской воды (соленость до 9%). [5] [6]
Начиная с 2010 года технологии дистанционного зондирования и глобальные данные используются для оценки площадей, условий и темпов обезлесения мангровых зарослей по всему миру. [7] [1] [2] В 2018 году инициатива Global Mangrove Watch опубликовала новый глобальный базовый показатель, согласно которому общая площадь мангровых лесов в мире по состоянию на 2010 год оценивается в 137 600 км2. 2 (53 100 квадратных миль), охватывающий 118 стран и территорий. [2] [7] Исследование 2022 года, посвященное потерям и увеличению приливных водно-болотных угодий, оценивает площадь в 3700 км2. 2 (1400 квадратных миль) чистое сокращение площади мангровых зарослей в мире с 1999 по 2019 год. [8] Утрата мангровых лесов продолжается из-за деятельности человека: глобальные ежегодные темпы вырубки лесов оцениваются в 0,16%, а показатели по странам достигают 0,70%. Деградация качества сохранившихся мангровых зарослей также является серьезной проблемой. [2]
существует Интерес к восстановлению мангровых зарослей по нескольким причинам. Мангровые заросли поддерживают устойчивые прибрежные и морские экосистемы. Они защищают близлежащие районы от цунами и экстремальных погодных явлений. Мангровые леса также эффективны в улавливании и хранении углерода. [2] [9] [10] Успех восстановления мангровых зарослей может во многом зависеть от взаимодействия с местными заинтересованными сторонами, а также от тщательной оценки, позволяющей гарантировать, что условия выращивания будут подходить для выбранных видов. [4]
Международный день охраны мангровых экосистем отмечается ежегодно 26 июля. [11]
Этимология
[ редактировать ]

Этимология английского термина «мангровые деревья» может быть только спекулятивной и является спорной. [12] : 1–2 [13] Этот термин, возможно, пришел в английский язык из португальского mangue или испанского языка. калечить . [13] Еще дальше его можно отнести к Южной Америке, карибанским и аравакским языкам. [14] например, Тайно . [15] Другие возможности включают малайский язык мангги-манги. [13] [12] Английское использование может отражать искажение народной этимологии слов «мангроу» и «роща» . [14] [12] [16]
Слово «мангровые заросли» используется как минимум в трех значениях:
- В наиболее широком смысле это относится к среде обитания и всему растительному комплексу или мангалу . [13] [17] термины мангрового леса» «биом и «мангровое болото» ; для которых также используются
- Для обозначения всех деревьев и крупных кустарников мангрового болота ; [13] и
- только к мангровым деревьям рода Rhizophora семейства В узком смысле это относится Rhizophoraceae . [18]
Биология
[ редактировать ]По данным Хогарта (2015), среди признанных видов мангровых зарослей насчитывается около 70 видов 20 родов из 16 семейств , составляющих «настоящие мангровые заросли» – виды, которые встречаются почти исключительно в мангровых местообитаниях. [17] Демонстрируя конвергентную эволюцию , многие из этих видов нашли сходные решения для тропических условий с переменной соленостью, приливным диапазоном (наводнением), анаэробными почвами и интенсивным солнечным светом. Биоразнообразие растений в данном мангровом заросле обычно невелико. [19] Наибольшее биоразнообразие мангровых зарослей встречается в Юго-Восточной Азии , особенно на Индонезийском архипелаге . [20]

Адаптация к низкому содержанию кислорода
[ редактировать ]Красный мангровый лес ( Rhizophora mangle ) выживает на наиболее затопленных участках, опирается над уровнем воды с помощью ходулей или опорных корней, а затем поглощает воздух через чечевички в своей коре. [21] Черный мангровый лес ( Avicennia Germinans ) живет на возвышенностях и развивает множество специализированных корнеобразных структур, называемых пневматофорами , которые торчат из почвы, как соломинки для дыхания. [22] [23] Эти «дыхательные трубки» обычно достигают высоты до 30 см (12 дюймов), а у некоторых видов - более 3 м (9,8 футов). Корни также содержат широкую аэренхиму , облегчающую транспортировку внутри растения. [ нужна ссылка ]
Поглощение питательных веществ
[ редактировать ]Поскольку почва постоянно заболочена, свободного кислорода в ней мало. Анаэробные бактерии выделяют газообразный азот , растворимое железо (железо), неорганические фосфаты , сульфиды и метан , которые делают почву гораздо менее питательной. [ нужна ссылка ] Пневматофоры ( воздушные корни ) позволяют мангровым деревьям поглощать газы непосредственно из атмосферы, а другие питательные вещества, такие как железо, из негостеприимной почвы. Мангровые деревья хранят газы непосредственно внутри корней, перерабатывая их, даже когда корни погружаются под воду во время прилива.

Ограничение потребления соли
[ редактировать ]Красные мангровые деревья исключают соль, поскольку имеют значительно непроницаемые корни, которые сильно суберинизированы (пропитаны суберином ), действуя как механизм ультрафильтрации, исключающий натрия соли из остальной части растения. [ нужна ссылка ] One study found that roots of the Indian mangrove Avicennia officinalis exclude 90% to 95% of the salt in water taken up by the plant, depositing the excluded salt in the cortex of the root. An increase in the production of suberin and in the activity of a gene regulating cytochrome P450 were observed in correlation with an increase in the salinity of the water to which the plant was exposed.[24] In a frequently cited concept that has become known as the "sacrificial leaf", salt which does accumulate in the shoot (sprout) then concentrates in old leaves, which the plant then sheds. However, recent research on the Red mangrove Rhizophora mangle suggests that the older, yellowing leaves have no more measurable salt content than the other, greener leaves.[25]
 Pneumatophorous aerial roots of the grey mangrove (Avicennia marina)
Pneumatophorous aerial roots of the grey mangrove (Avicennia marina) Vivipary in Rhizophora mangle seeds
Vivipary in Rhizophora mangle seeds


Limiting water loss
[edit]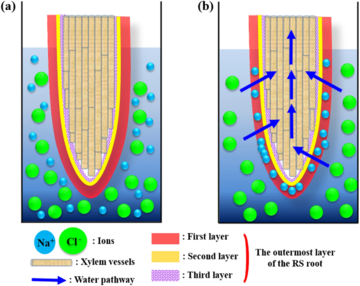
Because of the limited fresh water available in salty intertidal soils, mangroves limit the amount of water they lose through their leaves. They can restrict the opening of their stomata (pores on the leaf surfaces, which exchange carbon dioxide gas and water vapor during photosynthesis). They also vary the orientation of their leaves to avoid the harsh midday sun and so reduce evaporation from the leaves. A captive red mangrove grows only if its leaves are misted with fresh water several times a week, simulating frequent tropical rainstorms.[27]
Filtration of seawater
[edit]A 2016 study by Kim et al. investigated the biophysical characteristics of sea water filtration in the roots of the mangrove Rhizophora stylosa from a plant hydrodynamic point of view. R. stylosa can grow even in saline water and the salt level in its roots is regulated within a certain threshold value through filtration. The root possesses a hierarchical, triple layered pore structure in the epidermis and most Na+ ions are filtered at the first sublayer of the outermost layer. The high blockage of Na+ ions is attributed to the high surface zeta potential of the first layer. The second layer, which is composed of macroporous structures, also facilitates Na+ ion filtration. The study provides insights into the mechanism underlying water filtration through halophyte roots and could serve as a basis for the development of a bio-inspired method of desalination.[26]
Uptake of Na+ ions is desirable for halophytes to build up osmotic potential, absorb water and sustain turgor pressure. However, excess Na+ions may work on toxic element. Therefore, halophytes try to adjust salinity delicately between growth and survival strategies. In this point of view, a novel sustainable desalination method can be derived from halophytes, which are in contact with saline water through their roots. Halophytes exclude salt through their roots, secrete the accumulated salt through their aerial parts and sequester salt in senescent leaves and/or the bark.[28][29][30] Mangroves are facultative halophytes and Bruguiera is known for its special ultrafiltration system that can filter approximately 90% of Na+ions from the surrounding seawater through the roots.[31][32][33] The species also exhibits a high rate of salt rejection. The water-filtering process in mangrove roots has received considerable attention for several decades.[34][35] Morphological structures of plants and their functions have been evolved through a long history to survive against harsh environmental conditions.[36][26]
Increasing survival of offspring
[edit]
In this harsh environment, mangroves have evolved a special mechanism to help their offspring survive. Mangrove seeds are buoyant and are therefore suited to water dispersal. Unlike most plants, whose seeds germinate in soil, many mangroves (e.g. red mangrove) are viviparous,[37] meaning their seeds germinate while still attached to the parent tree. Once germinated, the seedling grows either within the fruit (e.g. Aegialitis, Avicennia and Aegiceras), or out through the fruit (e.g. Rhizophora, Ceriops, Bruguiera and Nypa) to form a propagule (a ready-to-go seedling) which can produce its own food via photosynthesis.
The mature propagule then drops into the water, which can transport it great distances. Propagules can survive desiccation and remain dormant for over a year before arriving in a suitable environment. Once a propagule is ready to root, its density changes so that the elongated shape now floats vertically rather than horizontally. In this position, it is more likely to lodge in the mud and root. If it does not root, it can alter its density and drift again in search of more favorable conditions.
Taxonomy and evolution
[edit]The following listings, based on Tomlinson, 2016, give the mangrove species in each listed plant genus and family.[38] Mangrove environments in the Eastern Hemisphere harbor six times as many species of trees and shrubs as do mangroves in the New World. Genetic divergence of mangrove lineages from terrestrial relatives, in combination with fossil evidence, suggests mangrove diversity is limited by evolutionary transition into the stressful marine environment, and the number of mangrove lineages has increased steadily over the Tertiary with little global extinction.[39]
True mangroves
[edit]| True mangroves (major components or strict mangroves) | ||||
|---|---|---|---|---|
| Following Tomlinson, 2016, the following 35 species are the true mangroves, contained in 5 families and 9 genera[38]: 29–30 Included on green backgrounds are annotations about the genera made by Tomlinson | ||||
| Family | Genus | Mangrove species | Common name | |
| Arecaceae | Monotypic subfamily within the family | |||
| Nypa | Nypa fruticans | Mangrove palm |  | |
| Avicenniaceae (disputed) | Old monogeneric family, now subsumed in Acanthaceae, but clearly isolated | |||
| Avicennia | Avicennia alba |  | ||
| Avicennia balanophora | ||||
| Avicennia bicolor | ||||
| Avicennia integra | ||||
| Avicennia marina | grey mangrove (subspecies: australasica, eucalyptifolia, rumphiana) | |||
| Avicennia officinalis | Indian mangrove | |||
| Avicennia germinans | black mangrove | |||
| Avicennia schaueriana | ||||
| Avicennia tonduzii | ||||
| Combretaceae | Tribe Lagunculariae (including Macropteranthes = non-mangrove) | |||
| Laguncularia | Laguncularia racemosa | white mangrove |  | |
| Lumnitzera | Lumnitzera racemosa | white-flowered black mangrove | ||
| Lumnitzera littorea | ||||
| Rhizophoraceae | Rhizophoraceae collectively form the tribe Rhizophorae, a monotypic group, within the otherwise terrestrial family | |||
| Bruguiera | Bruguiera cylindrica | |||
| Bruguiera exaristata | rib-fruited mangrove | |||
| Bruguiera gymnorhiza | oriental mangrove |  | ||
| Bruguiera hainesii | ||||
| Bruguiera parviflora | ||||
| Bruguiera sexangula | upriver orange mangrove | |||
| Ceriops | Ceriops australis | yellow mangrove |  | |
| Ceriops tagal | spurred mangrove |  | ||
| Kandelia | Kandelia candel | |||
| Kandelia obovata | ||||
| Rhizophora | Rhizophora apiculata | |||
| Rhizophora harrisonii | ||||
| Rhizophora mangle | red mangrove | |||
| Rhizophora mucronata | Asiatic mangrove |  | ||
| Rhizophora racemosa | ||||
| Rhizophora samoensis | Samoan mangrove | |||
| Rhizophora stylosa | spotted mangrove, | |||
| Rhizophora x lamarckii | ||||
| Lythraceae | Sonneratia | Sonneratia alba |  | |
| Sonneratia apetala | ||||
| Sonneratia caseolaris | ||||
| Sonneratia ovata | ||||
| Sonneratia griffithii | ||||
Minor components
[edit]| Minor components | ||||
|---|---|---|---|---|
| Tomlinson, 2016, lists about 19 species as minor mangrove components, contained in 10 families and 11 genera[38]: 29–30 Included on green backgrounds are annotations about the genera made by Tomlinson | ||||
| Family | Genus | Species | Common name | |
| Euphorbiaceae | This genus includes about 35 non-mangrove taxa | |||
| Excoecaria | Excoecaria agallocha | milky mangrove, blind-your-eye mangrove and river poison tree |  | |
| Lythraceae | Genus distinct in the family | |||
| Pemphis | Pemphis acidula | bantigue or mentigi | ||
| Malvaceae | Formerly in Bombacaceae, now an isolated genus in subfamily Bombacoideeae | |||
| Camptostemon | Camptostemon schultzii | kapok mangrove |  | |
| Camptostemon philippinense | ||||
| Meliaceae | Genus of 3 species, one non-mangrove, forms tribe Xylocarpaeae with Carapa, a non–mangrove | |||
| Xylocarpus | Xylocarpus granatum |  | ||
| Xylocarpus moluccensis | ||||
| Myrtaceae | An isolated genus in the family | |||
| Osbornia | Osbornia octodonta | mangrove myrtle | ||
| Pellicieraceae | Monotypic genus and family of uncertain phylogenetic position | |||
| Pelliciera | Pelliciera rhizophorae | tea mangrove | ||
| Plumbaginaceae | Isolated genus, at times segregated as family Aegialitidaceae | |||
| Aegialitis | Aegialitis annulata | club mangrove | ||
| Aegialitis rotundifolia | ||||
| Primulaceae | Formerly an isolated genus in Myrsinaceae | |||
| Aegiceras | Aegiceras corniculatum | black mangrove, river mangrove or khalsi | ||
| Aegiceras floridum | ||||
| Pteridaceae | A fern somewhat isolated in its family | |||
| Acrostichum | Acrostichum aureum | golden leather fern, swamp fern or mangrove fern | ||
| Acrostichum speciosum | mangrove fern | |||
| Rubiaceae | A genus isolated in the family | |||
| Scyphiphora | Scyphiphora hydrophylacea | nilad | ||
Species distribution
[edit]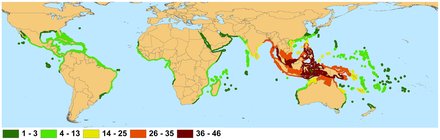
Mangroves are a type of tropical vegetation with some outliers established in subtropical latitudes, notably in South Florida and southern Japan, as well as South Africa, New Zealand and Victoria (Australia). These outliers result either from unbroken coastlines and island chains or from reliable supplies of propagules floating on warm ocean currents from rich mangrove regions.[38]: 57
"At the limits of distribution, the formation is represented by scrubby, usually monotypic Avicennia-dominated vegetation, as at Westonport Bay and Corner Inlet, Victoria, Australia. The latter locality is the highest latitude (38° 45'S) at which mangroves occur naturally. The mangroves in New Zealand, which extend as far south as 37°, are of the same type; they start as low forest in the northern part of the North Island but become low scrub toward their southern limit. In both instances, the species is referred to as Avicennia marina var. australis, although genetic comparison is clearly needed. In Western Australia, A. marina extends as far south as Bunbury (33° 19'S). In the northern hemisphere, scrubby Avicennia gerrninans in Florida occurs as far north as St. Augustine on the east coast and Cedar Point on the west. There are records of A. germinans and Rhizophora mangle for Bermuda, presumably supplied by the Gulf Stream. In southern Japan, Kandelia obovata occurs to about 31 °N (Tagawa in Hosakawa et al., 1977, but initially referred to as K. candel)."[38]: 57
Mangrove forests
[edit]
Mangrove forests, also called mangrove swamps or mangals, are found in tropical and subtropical tidal areas. Areas where mangroves occur include estuaries and marine shorelines.[19]
The intertidal existence to which these trees are adapted represents the major limitation to the number of species able to thrive in their habitat. High tide brings in salt water, and when the tide recedes, solar evaporation of the seawater in the soil leads to further increases in salinity. The return of tide can flush out these soils, bringing them back to salinity levels comparable to that of seawater.[2][4]
At low tide, organisms are also exposed to increases in temperature and reduced moisture before being then cooled and flooded by the tide. Thus, for a plant to survive in this environment, it must tolerate broad ranges of salinity, temperature, and moisture, as well as several other key environmental factors—thus only a select few species make up the mangrove tree community.[2][4]
About 110 species are considered mangroves, in the sense of being trees that grow in such a saline swamp,[19] though only a few are from the mangrove plant genus, Rhizophora. However, a given mangrove swamp typically features only a small number of tree species. It is not uncommon for a mangrove forest in the Caribbean to feature only three or four tree species. For comparison, the tropical rainforest biome contains thousands of tree species, but this is not to say mangrove forests lack diversity. Though the trees themselves are few in species, the ecosystem that these trees create provides a home (habitat) for a great variety of other species, including as many as 174 species of marine megafauna.[42]

Mangrove plants require a number of physiological adaptations to overcome the problems of low environmental oxygen levels, high salinity, and frequent tidal flooding. Each species has its own solutions to these problems; this may be the primary reason why, on some shorelines, mangrove tree species show distinct zonation. Small environmental variations within a mangal may lead to greatly differing methods for coping with the environment. Therefore, the mix of species is partly determined by the tolerances of individual species to physical conditions, such as tidal flooding and salinity, but may also be influenced by other factors, such as crabs preying on plant seedlings.[43]

Once established, mangrove roots provide an oyster habitat and slow water flow, thereby enhancing sediment deposition in areas where it is already occurring. The fine, anoxic sediments under mangroves act as sinks for a variety of heavy (trace) metals which colloidal particles in the sediments have concentrated from the water. Mangrove removal disturbs these underlying sediments, often creating problems of trace metal contamination of seawater and organisms of the area.[44]
Mangrove swamps protect coastal areas from erosion, storm surge (especially during tropical cyclones), and tsunamis.[45][46][47] They limit high-energy wave erosion mainly during events such as storm surges and tsunamis.[48]The mangroves' massive root systems are efficient at dissipating wave energy.[49] Likewise, they slow down tidal water so that its sediment is deposited as the tide comes in, leaving all except fine particles when the tide ebbs.[50] In this way, mangroves build their environments.[45] Because of the uniqueness of mangrove ecosystems and the protection against erosion they provide, they are often the object of conservation programs,[4] including national biodiversity action plans.[46]
The unique ecosystem found in the intricate mesh of mangrove roots offers a quiet marine habitat for young organisms.[51] In areas where roots are permanently submerged, the organisms they host include algae, barnacles, oysters, sponges, and bryozoans, which all require a hard surface for anchoring while they filter-feed. Shrimps and mud lobsters use the muddy bottoms as their home.[52] Mangrove crabs eat the mangrove leaves, adding nutrients to the mangal mud for other bottom feeders.[53] In at least some cases, the export of carbon fixed in mangroves is important in coastal food webs.[54]
Mangrove forests contribute significantly to coastal ecosystems by fostering complex and diverse food webs. The intricate root systems of mangroves create a habitat conducive to the proliferation of microorganisms, crustaceans, and small fish, forming the foundational tiers of the food chain. This abundance of organisms serves as a critical food source for larger predators like birds, reptiles, and mammals within the ecosystem. Additionally, mangrove forests function as essential nurseries for many commercially important fish species, providing a sheltered environment rich in nutrients during their early life stages. The decomposition of leaves and organic matter in the water further enhances the nutrient content, supporting overall ecosystem productivity. In summary, mangrove forests play a crucial and unbiased role in sustaining biodiversity and ecological balance within coastal food webs.[55]
Larger marine organisms benefit from the habitat as a nursery for their offspring. Lemon Sharks depend on mangrove creeks to give birth to their pups. The ecosystem provides little competition and minimizes threats of predation to juvenile lemon sharks as they use the cover of mangroves to practice hunting before entering the food web of the Ocean.[56]
Mangrove plantations in Vietnam, Thailand, Philippines, and India host several commercially important species of fish and crustaceans.[57]
The mangrove food chain extends beyond the marine ecosystem. Coastal bird species inhabit the tidal ecosystems feeding off small marine organisms and wetland insects. Common bird families found in mangroves around the world are egrets, kingfishers, herons, and hornbills, among many others dependent on ecological range.[58] Bird predation plays a key role in maintaining prey species along coastlines and within mangrove ecosystems.
Мангровые леса могут превратиться в торфяные отложения в результате грибковых и бактериальных процессов, а также под действием термитов . В хороших геохимических , осадочных и тектонических условиях он превращается в торф. [59] Природа этих отложений зависит от окружающей среды и типов мангровых зарослей. В Рико красные белые , черные и Пуэрто - мангровые заросли занимают разные экологические ниши и имеют немного разный химический состав, поэтому содержание углерода варьируется между видами, а также между различными тканями растения (например, в листьях и корнях). [59]
В Пуэрто-Рико наблюдается четкая последовательность этих трех деревьев от более низких возвышенностей, где преобладают красные мангровые заросли, до более глубоких территорий с более высокой концентрацией белых мангровых зарослей. [59] Мангровые леса являются важной частью круговорота и хранения углерода в тропических прибрежных экосистемах. [59] Зная это, ученые стремятся реконструировать окружающую среду и исследовать изменения в прибрежной экосистеме за тысячи лет, используя керны отложений. [60] Однако дополнительной сложностью является импортное морское органическое вещество, которое также откладывается в отложениях из-за приливных смывов мангровых лесов. Термиты играют важную роль в образовании торфа из мангровых материалов. [59] Они перерабатывают опавшую листву , корневую систему и древесину мангровых зарослей в торф для строительства своих гнезд и стабилизируют химический состав этого торфа, который составляет примерно 2% надземных запасов углерода в мангровых зарослях. Поскольку гнезда со временем закапываются, этот углерод сохраняется в отложениях, и углеродный цикл продолжается. [59]
Мангровые заросли являются важным источником голубого углерода . Во всем мире мангровые заросли хранят 4,19 Гт (9,2 × 10 12 фунтов) углерода в 2012 году. Два процента мирового углерода мангровых лесов были потеряны в период с 2000 по 2012 год, что эквивалентно максимальному потенциалу в 0,316996250 Гт (6,9885710 × 10 11 lb) выбросов углекислого газа в атмосферу Земли . [61]
Было доказано, что во всем мире мангровые заросли обеспечивают измеримую экономическую защиту прибрежных сообществ, пострадавших от тропических штормов. [62]
Мангровый микробиом
[ редактировать ]Микробиомы растений играют решающую роль в их здоровье и продуктивности мангровых зарослей. [63] Многие исследователи успешно применили полученные знания о микробиомах растений для производства специфических инокулятов для защиты сельскохозяйственных культур. [64] [65] Такая инокулят может стимулировать рост растений, высвобождая фитогормоны и улучшая усвоение некоторых минеральных питательных веществ (особенно фосфора и азота). [65] [66] [67] Однако большинство исследований микробиома растений были сосредоточены на модельном растении Arabidopsis thaliana и экономически важных сельскохозяйственных растениях, таких как рис , ячмень , пшеница , кукуруза и соя . О микробиомах древесных пород информации меньше. [63] [65] Микробиомы растений определяются факторами, связанными с растениями (например, генотипом , органом, видом и состоянием здоровья) и факторами окружающей среды (например, землепользованием, климатом и доступностью питательных веществ). [63] [67] Было показано, что два фактора, связанных с растениями, виды и генотипы растений, играют важную роль в формировании ризосферы и микробиомов растений, поскольку генотипы и виды деревьев связаны с конкретными микробными сообществами . [66] Различные органы растений также имеют специфические микробные сообщества в зависимости от факторов, связанных с растением (генотип растения, доступные питательные вещества и специфичные для органа физико-химические условия) и/или условий окружающей среды (связанных с надземными и подземными поверхностями и нарушениями). [68] [69] [70] [71]
Корневой микробиом
[ редактировать ]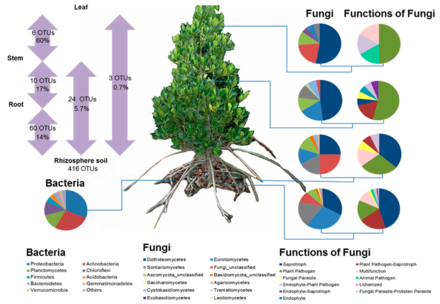
Корни мангровых зарослей содержат набор микробных таксонов , которые способствуют важным экологическим функциям в мангровых экосистемах. Подобно типичным наземным растениям, мангровые заросли зависят от взаимовыгодного взаимодействия с микробными сообществами. [72] В частности, микробы, обитающие в развитых корнях, могут помочь мангровым зарослям преобразовывать питательные вещества в полезные формы до ассимиляции растениями. [73] [74] Эти микробы также обеспечивают фитогормоны мангровых деревьев для подавления фитопатогенов. [75] или помочь мангровым деревьям противостоять жаре и солености. [72] В свою очередь, корневые микробы получают метаболиты углерода из растения через корневые экссудаты . [76] таким образом устанавливаются тесные связи между растением и микробами для их взаимной выгоды. [77] [78]
Уровень таксономического класса показывает, что большинство протеобактерий произошли от гаммапротеобактерий, за которыми следовали дельтапротеобактерии и альфапротеобактерии. Разнообразные функции и филогенная изменчивость гаммапротеобактерий, состоящих из таких отрядов, как Alteromonadales и Vibrionales, обнаружены в морских и прибрежных регионах и в большом количестве представлены в отложениях мангровых зарослей, выполняющих функцию переработчиков питательных веществ. Члены Deltaproteobacteria, обнаруженные в мангровых почвах, в основном связаны с серой, включая Desulfobacterales , Desulfuromonadales , Desulfovibrionales и Desulfarculales, среди других. [79] весьма разнообразные микробные сообщества (в основном бактерии и грибы ). Было обнаружено, что в корнях мангровых зарослей обитают и функционируют [80] [72] [81] Например, диазотрофные бактерии вблизи корней мангровых деревьев могут осуществлять биологическую фиксацию азота , что обеспечивает 40–60% общего азота, необходимого мангровым деревьям; [82] [83] почва, прикрепленная к корням мангровых деревьев, лишена кислорода, но богата органическими веществами, обеспечивая оптимальную микросреду для сульфатредуцирующих бактерий и метаногенов , [72] лигнинолитические , целлюлозолитические и амилолитические грибы; в корневой среде мангровых зарослей преобладают [72] Ризосферные грибы могут помочь мангровым деревьям выжить в заболоченной среде с ограниченным количеством питательных веществ. [84] Эти исследования предоставили все больше доказательств, подтверждающих важность корневых бактерий и грибов для роста и здоровья мангровых зарослей. [72] [73] [78]
Недавние исследования изучили детальную структуру микробных сообществ, связанных с корнями, на непрерывном мелкомасштабном уровне у других растений. [85] где микросреда обитания была разделена на четыре корневых отсека: эндосферу, [75] [86] [87] эписфера [75] ризосфера, [86] [88] и внеризосферные. [89] [90] Более того, сообщалось, что микробные сообщества в каждом отсеке обладают уникальными характеристиками. [75] [86] Ризосфера могла выделять корневые экссудаты, которые выборочно обогащали определенные микробные популяции; однако было обнаружено, что эти экссудаты оказывают лишь незначительное воздействие на микробы внеризосферной почвы. [91] [77] Более того, было отмечено, что корневая эписфера, а не ризосфера, несет основную ответственность за контроль проникновения определенных микробных популяций в корень. [75] что приводит к избирательному обогащению протеобактерий в эндосфере. [75] [92] Эти результаты дают новое представление о дифференциации ниш микробных сообществ, связанных с корнями. [75] [91] [77] [92] Тем не менее, профилирование сообществ на основе ампликонов может не дать функциональных характеристик корневых микробных сообществ в росте растений и биогеохимическом цикле. [93] Раскрытие функциональных закономерностей в четырех корневых отделах открывает большой потенциал для понимания функциональных механизмов, ответственных за взаимодействие корня и микробов в поддержку улучшения функционирования мангровых экосистем. [78]
Сообщается, что разнообразие бактерий в нарушенных мангровых зарослях выше, чем вхорошо сохранившиеся мангровые заросли [79] Исследования, сравнивающие мангровые заросли в различных состояниях сохранности, показывают, что бактериальный состав нарушенных мангровых отложений изменяет их структуру, приводя к функциональному равновесию, при этом динамика химических веществ в мангровых почвах приводит к ремоделированию их микробной структуры. [94]
Предложения по будущим исследованиям микробного разнообразия мангровых лесов
[ редактировать ]Несмотря на многочисленные достижения в области метагеномики бактерий в отложениях мангровых лесов,разнообразия в различных условиях за последние несколько лет, устраняя пробел в исследованиях ирасширение наших знаний в отношении взаимоотношений между микробами, состоящими в основном из бактерий, и их питательными кругами в отложениях мангровых зарослей, а также прямым и косвенным воздействием на рост мангровых зарослей и структуры насаждений в качестве прибрежных барьеров и других поставщиков экологических услуг. Таким образом, основываясь на исследованиях систематического обзора Лая и др., здесь они предлагают улучшить выборку и фундаментальный экологический индекс для дальнейшего использования. [79]
Мангровый виром
[ редактировать ]
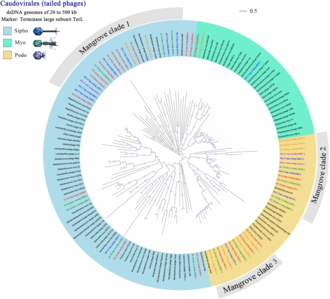
Мангровые леса являются одним из наиболее богатых углеродом биомов, на их долю приходится 11% общего поступления земного углерода в океаны. вирусы Считается, что существенно влияют на локальные и глобальные биогеохимические циклы , хотя по состоянию на 2019 год было доступно мало информации о структуре сообщества, генетическом разнообразии и экологической роли вирусов в мангровых экосистемах. [95]
Вирусы — самые распространенные биологические объекты на Земле, присутствующие практически во всех экосистемах. [96] [97] Лизируя своих хозяев, то есть разрывая их клеточные мембраны, вирусы контролируют численность хозяев и влияют на структуру сообществ хозяев. [98] Вирусы также влияют на разнообразие и эволюцию своих хозяев посредством горизонтального переноса генов , отбора на устойчивость и манипулирования бактериальным метаболизмом . [99] [100] [101] Важно отметить, что морские вирусы влияют на локальные и глобальные биогеохимические циклы посредством высвобождения значительных количеств органического углерода и питательных веществ из хозяев и помогают микробам управлять биогеохимическими циклами с помощью вспомогательных метаболических генов (АМГ). [102] [103] [104] [95]
Предполагается, что АМГ усиливают метаболизм инфицированного вирусом хозяина и способствуют образованию новых вирусов. [99] [105] AMG широко изучались у морских цианофагов и включают гены, участвующие в фотосинтезе, обмене углерода, поглощении фосфатов и реакции на стресс. [106] [107] [108] [109] Независимый от культивирования метагеномный анализ вирусных сообществ выявил дополнительные AMG, которые участвуют в подвижности, центральном углеродном метаболизме, фотосистеме I, энергетическом метаболизме, железо-серных кластерах, антиокислительном цикле серы и азота. [103] [110] [111] [112] Интересно, что недавний анализ данных тихоокеанского вирома выявил AMG, специализирующиеся на нише, которые способствуют адаптации хозяина с стратификацией по глубине. [113] Учитывая, что микробы управляют глобальными биогеохимическими циклами, и большая часть микробов в любой момент времени заражается вирусами, [114] кодируемые вирусами AMG должны играть важную роль в глобальной биогеохимии и микробной метаболической эволюции. [95]
Мангровые леса — единственные древесные галофиты , обитающие в соленой воде вдоль субтропических и тропических побережий мира. Мангровые заросли — одна из самых продуктивных и экологически важных экосистем на Земле. Темпы первичной продукции мангровых зарослей аналогичны темпам производства тропических влажных вечнозеленых лесов и коралловых рифов. [115] Являясь глобально важным компонентом углеродного цикла, мангровые леса поглощают около 24 миллионов метрических тонн углерода каждый год. [115] [116] Большая часть углерода мангровых деревьев хранится в почве и значительных подземных резервуарах мертвых корней, что способствует сохранению и переработке питательных веществ под лесами. [117] Хотя мангровые заросли покрывают лишь 0,5% прибрежной территории Земли, на их долю приходится 10–15% запасов углерода в прибрежных отложениях и 10–11% общего поступления наземного углерода в океаны. [118] Непропорциональный вклад мангровых зарослей в поглощение углерода сейчас воспринимается как важное средство уравновешивания выбросов парниковых газов. [95]
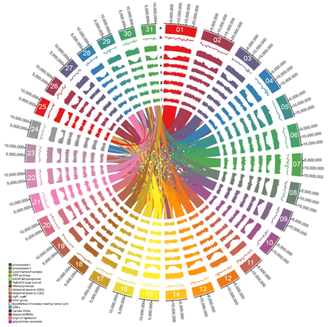
Несмотря на экологическую важность мангровых экосистем, знания о биоразнообразии мангровых лесов весьма ограничены. Предыдущие отчеты в основном исследовали биоразнообразие мангровых фауны, флоры и бактериальных сообществ. [120] [121] [122] В частности, мало информации доступно о вирусных сообществах и их роли в экосистемах мангровых почв. [123] [124] Учитывая важность вирусов в структурировании и регулировании сообществ-хозяев, а также в качестве посредников в биогеохимических циклах элементов, изучение вирусных сообществ в мангровых экосистемах имеет важное значение. Кроме того, периодическое затопление морской водой и, как следствие, резкая смена среды мангровых зарослей могут привести к существенному различию генетического и функционального разнообразия бактериальных и вирусных сообществ в мангровых почвах по сравнению с таковыми в других системах. [125] [95]
Секвенирование генома
[ редактировать ]- Rhizophoreae , выявленные с помощью полногеномного секвенирования [126]
См. также
[ редактировать ]- Прибрежное управление
- Экологическая ценность мангровых зарослей
- Краеугольные виды
- Аделаида К. Семеси
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Гири, К.; Очиенг, Э.; Тизен, LL; Чжу, З.; Сингх, А.; Лавленд, Т.; Масек, Дж.; Дюк, Н. (2011). «Состояние и распространение мангровых лесов в мире с использованием данных спутников наблюдения Земли: состояние и распространение мангровых лесов в мире» . Глобальная экология и биогеография . 20 (1): 154–159. дои : 10.1111/j.1466-8238.2010.00584.x .
- ^ Перейти обратно: а б с д и ж г час Фрисс, Д.А.; Роджерс, К.; Лавлок, CE; Краусс, КВ; Гамильтон, ЮВ; Ли, С.Ю.; Лукас, Р.; Примавера, Дж.; Раджкаран, А.; Ши, С. (2019). «Состояние мангровых лесов в мире: прошлое, настоящее и будущее» . Ежегодный обзор окружающей среды и ресурсов . 44 (1): 89–115. doi : 10.1146/annurev-environ-101718-033302 .
- ^ Флауэрс, Ти Джей; Колмер, Т.Д. (2015). «Солеустойчивость растений: адаптации галофитов» . Анналы ботаники . 115 (3): 327–331. дои : 10.1093/aob/mcu267 . ПМЦ 4332615 . ПМИД 25844430 .
- ^ Перейти обратно: а б с д и Циммер, Катарина (22 июля 2021 г.). «Многие реставрации мангровых зарослей терпят неудачу. Есть ли лучший способ?» . Знающий журнал . doi : 10.1146/knowable-072221-1 . Проверено 11 августа 2021 г.
- ^ «Морфологическая и физиологическая адаптация: сайт мангровых зарослей Флориды» . Nhmi.org. Архивировано из оригинала 4 февраля 2012 года . Проверено 8 февраля 2012 года .
- ^ Примавера, Дж. Х.; Саварис, JP; Баджойо, Бельгия; Кочинг, доктор медицинских наук; Курник, диджей; Гольбек, РЛ; Гузман, АТ; Хендерин, JQ; Ховен, Р.В.; Лома, РА; Колдевей, HJ (2012). Руководство по восстановлению мангровых зарослей на уровне общин (PDF) . Мангровое руководство. Зоологическое общество Лондона ZSL. Архивировано из оригинала (PDF) 1 января 2016 года . Проверено 15 августа 2021 г.
- ^ Перейти обратно: а б Бантинг, П.; Розенквист А.; Лукас, Р.; Ребело, Л.-М.; Хиларидес, Л.; Томас, Н.; Харди, А.; Ито, Т.; Шимада, М.; Финлейсон, К. (2018). «Глобальный обзор мангровых зарослей — новый глобальный базовый показатель распространенности мангровых зарослей на 2010 год» . Дистанционное зондирование . 10 (10): 1669. Бибкод : 2018RemS...10.1669B . дои : 10.3390/rs10101669 .
- ^ Мюррей, Нью-Джерси; Уортингтон, штат Техас; Бантинг, П.; Дуче, С.; Хаггер, В.; Лавлок, CE; Лукас, Р.; Сондерс, Мичиган; Шивс, М.; Сполдинг, М.; Уолтем, Нью-Джерси; Лайонс, МБ (2022 г.). «Картирование с высоким разрешением потерь и прироста приливных водно-болотных угодий Земли» . Наука . 376 (6594): 744–749. Бибкод : 2022Sci...376..744M . дои : 10.1126/science.abm9583 . hdl : 2160/55fdc0d4-aa3e-433f-8a88-2098b1372ac5 . ПМИД 35549414 . S2CID 248749118 .
- ^ Р., Кэрол; Карлович, М. (2019). «Новые спутниковые карты мангровых высот» . Проверено 15 мая 2019 г.
- ^ Симард, М.; Фатойинбо, Л.; Сметанка, Ц.; Ривера-Монрой, В.Х.; Кастаньеда-Мойя, Э.; Томас, Н.; Ван дер Стокен, Т. (2018). «Высота полога мангровых зарослей в глобальном масштабе связана с осадками, температурой и частотой циклонов». Природа Геонауки . 12 (1): 40–45. дои : 10.1038/s41561-018-0279-1 . hdl : 2060/20190029179 . S2CID 134827807 .
- ^ «Международный день охраны мангровых экосистем» . ЮНЕСКО . Проверено 9 июня 2023 г.
- ^ Перейти обратно: а б с Сенгер, П. (2013). Экология мангровых лесов, лесоводство и охрана (переиздание 2002 г.). Springer Science & Business Media. ISBN 9789401599627 .
- ^ Перейти обратно: а б с д и Макнэ, В. (1969). «Общий отчет о фауне и флоре мангровых болот и лесов Индо-Западно-Тихоокеанского региона» . Достижения морской биологии . 6 : 73–270. дои : 10.1016/S0065-2881(08)60438-1 . ISBN 9780120261062 . Проверено 13 августа 2021 г.
- ^ Перейти обратно: а б Герлах, М. (1 января 2003 г.). Английские слова за рубежом . Издательство Джона Бенджамина. п. 59. ИСБН 9027223319 . Проверено 13 августа 2021 г.
- ^ Рафинеск, CS (1836 г.). Американские нации . Том. 1. К.С. Рафинеск. п. 244.
- ^ Уикли, Эрнест (1967). Этимологический словарь современного английского языка . Том. 2 (Перепечатка изд. 1921 г.). Дувр. ISBN 9780486122861 . Проверено 13 августа 2021 г.
- ^ Перейти обратно: а б Хогарт, Питер Дж. (2015). Биология мангровых зарослей и морских трав . Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-871654-9 .
- ^ Остин, Д.Ф. (2004). Этноботаника Флориды . ЦРК Пресс. ISBN 978-0-203-49188-1 .
- ^ Перейти обратно: а б с Матиас, М.Э. «Мангал (Мангровые леса). Мировая растительность » . Ботанический сад Калифорнийского университета в Лос-Анджелесе . Botgard.ucla.edu. Архивировано из оригинала 9 февраля 2012 года . Проверено 8 февраля 2012 года .
- ^ «Распределение разнообразия кораллов, мангровых зарослей и морских водорослей» . Maps.grida.no. Архивировано из оригинала 5 марта 2010 года . Проверено 8 февраля 2012 года .
- ^ «Красный мангровый лес» . Министерство сельского хозяйства и рыболовства правительства Квинсленда . Январь 2013 года . Проверено 13 августа 2021 г.
- ^ «Черный мангровый лес ( Avicennia Germinans )» . Департамент окружающей среды и природных ресурсов правительства Бермудских островов . Проверено 13 августа 2021 г.
- ^ «Морфофизиологические адаптации» . Морской институт Ньюфаунд-Харбор . Проверено 13 августа 2021 г.
- ^ Кришнамурти, Паннага; Джьоти-Пракаш, Павитра А.; Цинь, Линь; Он, Цзе; Линь, Цинсун; Ло, Чианг-Шион; Кумар, Пракаш П. (июль 2014 г.). «Роль корневых гидрофобных барьеров в отторжении солей мангрового растения Avicennia officinalis » . Растение, клетка и окружающая среда . 37 (7): 1656–1671. дои : 10.1111/шт.12272 . ПМИД 24417377 .
- ^ Грей, Л. Джозеф; и др. (2010). «Гипотеза жертвенных листьев мангровых зарослей» (PDF) . Электронный журнал ISME/GLOMIS . ГЛОМИС . Проверено 21 января 2012 г.
- ^ Перейти обратно: а б с Ким, Кивунг; Со, Ынсок; Чанг, Сук-Кю; Пак, Тэ Чжон; Ли, Сан Джун (5 февраля 2016 г.). «Новая фильтрация соленой воды во внешнем слое корней мангровых деревьев» . Научные отчеты . 6 (1). Springer Science and Business Media LLC: 20426. Бибкод : 2016NatSR...620426K . дои : 10.1038/srep20426 . ISSN 2045-2322 . ПМЦ 4742776 . ПМИД 26846878 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Кальфо, Энтони (2006). «Мангровые заросли для морского аквариума» . Рифовое содержание . Риф Центральный. Архивировано из оригинала 1 февраля 2022 года . Проверено 8 февраля 2012 года .
- ^ Томлинсон, П. Ботаника мангровых зарослей. [116–130] (Издательство Кембриджского университета, Кембридж, 1986).
- ^ Чжэн, Вэнь-Цзяо; Ван, Вэнь-Цин; Линь, Пэн (1999). «Динамика содержания элементов в процессе развития гипокотилей и листьев некоторых видов мангровых деревьев». Журнал экспериментальной морской биологии и экологии . 233 (2): 247–257. Бибкод : 1999JEMBE.233..247Z . дои : 10.1016/S0022-0981(98)00131-2 .
- ^ Парида, Асиш Кумар; Джа, Бхаванатх (2010). «Механизмы солеустойчивости в мангровых зарослях: обзор». Деревья . 24 (2): 199–217. Бибкод : 2010Деревья..24..199P . дои : 10.1007/s00468-010-0417-x . S2CID 3036770 .
- ^ Кришнамурти, Паннага; Джьоти-Пракаш, Павитра А.; Цинь, ЛИН; Он, Цзе; Линь, Цинсун; Ло, Чианг-Шион; Кумар, Пракаш П. (2014). «Роль корневых гидрофобных барьеров в исключении солей мангрового растения Avicennia officinalis» . Растение, клетка и окружающая среда . 37 (7): 1656–1671. дои : 10.1111/шт.12272 . ПМИД 24417377 .
- ^ Шоландер, П.Ф. (1968). «Как мангровые заросли опресняют морскую воду». Физиология Плантарум . 21 : 251–261. дои : 10.1111/j.1399-3054.1968.tb07248.x .
- ^ Шоландер, П.Ф.; Брэдстрит, Эдда Д.; Хаммель, ХТ; Хеммингсен, Э.А. (1966). «Концентрация сока у галофитов и некоторых других растений» . Физиология растений . 41 (3): 529–532. дои : 10.1104/стр.41.3.529 . ПМК 1086377 . ПМИД 5906381 .
- ^ Дреннан, Филиппа; Памментер, Северо-Запад (1982). «Физиология солевого выделения в мангровых зарослях Авиценния Марина (Форск.) Вирх» . Новый фитолог . 91 (4): 597–606. дои : 10.1111/j.1469-8137.1982.tb03338.x .
- ^ Собрадо, Массачусетс (2001). «Влияние высокой внешней концентрации Na Cl на осмоляльность сока ксилемы, ткани листьев и секреции желез листьев мангровых деревьев Avicennia Germinans (L.) L». Флора . 196 (1): 63–70. Бибкод : 2001FMDFE.196...63S . дои : 10.1016/S0367-2530(17)30013-0 .
- ^ Фудзита, Мики; Фудзита, Ясунари; Нотоши, Ёситеру; Такахаси, Фуминори; Нарусака, Ёсихиро; Ямагучи-Синозаки, Кадзуко; Шинозаки, Кадзуо (2006). «Перекресток между абиотическими и биотическими реакциями на стресс: современный взгляд с точек конвергенции сетей передачи сигналов стресса». Современное мнение в области биологии растений . 9 (4): 436–442. Бибкод : 2006COPB....9..436F . дои : 10.1016/j.pbi.2006.05.014 . ПМИД 16759898 . S2CID 31166870 .
- ^ Хогарт, П.Дж. (1 января 2017 г.), «Мангровые экосистемы☆» , Справочный модуль по наукам о жизни , Elsevier, doi : 10.1016/b978-0-12-809633-8.02209-3 , ISBN 978-0-12-809633-8 , получено 1 марта 2024 г.
- ^ Перейти обратно: а б с д и Томлинсон, ПБ (2016). Ботаника мангровых зарослей . Кембридж, Соединенное Королевство: Издательство Кембриджского университета. ISBN 978-1-107-08067-6 . OCLC 946579968 .
- ^ Риклефс, RE; А. Шварцбах; СС Реннер (2006). «Скорость происхождения линий объясняет аномальное разнообразие мангровой растительности в мире» (PDF) . Американский натуралист . 168 (6): 805–810. дои : 10.1086/508711 . ПМИД 17109322 . S2CID 1493815 . Архивировано из оригинала (PDF) 16 июня 2013 года.
- ^ Перейти обратно: а б Полидоро, Бет А.; Карпентер, Кент Э.; Коллинз, Лорна; Дьюк, Норман К.; Эллисон, Аарон М.; Эллисон, Джоанна С.; Фарнсворт, Элизабет Дж.; Фернандо, Эдвино С.; Катиресан, Кандасами; Коедам, Нико Э.; Ливингстон, Сюзанна Р.; Мияги, Тоёхико; Мур, Грегг Э.; Нгок Нам, Вена; Онг, Джин Ёнг; Примавера, Юргенне Х.; Салмо, Северино Г.; Санчиангко, Джоннелл К.; Сукарджо, Сукристихоно; Ван, Ямин; Ён, Джин Ван Хонг (2010). «Потеря видов: риск исчезновения мангровых зарослей и географические районы, вызывающие глобальную озабоченность» . ПЛОС ОДИН . 5 (4): e10095. Бибкод : 2010PLoSO...510095P . дои : 10.1371/journal.pone.0010095 . ПМЦ 2851656 . ПМИД 20386710 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ «Картирование мангровых зарослей со спутника» . Earthobservatory.nasa.gov . 30 ноября 2010 г.
- ^ Сиверс, М.; Браун, CJ; Таллох, VJD; Пирсон, РМ; Хейг, Дж.А.; Тершвелл, член парламента; Коннолли, РМ (2019). «Роль прибрежных водно-болотных угодий с растительностью в сохранении морской мегафауны». Тенденции в экологии и эволюции . 34 (9): 807–817. Бибкод : 2019TEcoE..34..807S . дои : 10.1016/j.tree.2019.04.004 . hdl : 10072/391960 . ПМИД 31126633 . S2CID 164219103 .
- ^ Канниччи, С.; Фуси, М.; Симо, Ф.; Даду-Гебас, Ф.; Фратини, С. (2018). «Интерференционная конкуренция как ключевой фактор пространственного распределения мангровых крабов» . БМК Экология . 18 (1): 8. Бибкод : 2018BMCE...18....8C . дои : 10.1186/s12898-018-0164-1 . ПМК 5815208 . ПМИД 29448932 .
- ^ Сенгер, П.; МакКончи, Д. (2004). «Тяжелые металлы в мангровых зарослях: методология, мониторинг и управление». Лесной бюллетень Энвиса . 4 : 52–62. CiteSeerX 10.1.1.961.9649 .
- ^ Перейти обратно: а б Мазда, Ю.; Кобаши, Д.; Окада, С. (2005). «Приливная гидродинамика в мангровых болотах». Экология и управление водно-болотными угодьями . 13 (6): 647–655. Бибкод : 2005WetEM..13..647M . CiteSeerX 10.1.1.522.5345 . дои : 10.1007/s11273-005-0613-4 . S2CID 35322400 .
- ^ Перейти обратно: а б Дэниэлсен, Ф.; Соренсен, депутат Кнессета; Ольвиг, МФ; Сельвам, В.; Пэриш, Ф.; Берджесс, Северная Дакота; Хираиши, Т.; Карунагаран, В.М.; Расмуссен, MS; Хансен, Л.Б.; Куарто, А.; Сурьядипутра, Н. (2005). «Азиатское цунами: защитная роль прибрежной растительности». Наука . 310 (5748): 643. doi : 10.1126/science.1118387 . ПМИД 16254180 . S2CID 31945341 .
- ^ Такаги, Х.; Миками, Т.; Фуджи, Д.; Эстебан, М.; Куробе, С. (2016). «Мангровые леса против цунами, вызванного прорывом дамб, на быстро погружающихся побережьях» . Природные опасности и науки о системе Земли . 16 (7): 1629–1638. Бибкод : 2016NHESS..16.1629T . doi : 10.5194/nhess-16-1629-2016 .
- ^ Даду-Гебас, Ф.; Джаятисса, LP; Ди Нитто, Д.; Босире, Дж.О.; Ло Син, Д.; Коедам, Н. (2005). «Насколько эффективны мангровые заросли в качестве защиты от недавнего цунами?» . Современная биология . 15 (12): Р443–447. дои : 10.1016/j.cub.2005.06.008 . ПМИД 15964259 . S2CID 8772526 .
- ^ Массель, СР; Фурукава, К.; Бринкман, Р.М. (1999). «Распространение поверхностных волн в мангровых лесах». Исследование гидродинамики . 24 (4): 219. Бибкод : 1999FlDyR..24..219M . дои : 10.1016/s0169-5983(98)00024-0 . S2CID 122572658 .
- ^ Мазда, Ю.; Волански, Э.; Кинг, Б.; Сасе, А.; Оцука, Д.; Маги, М. (1997). «Сила сопротивления из-за растительности в мангровых болотах». Мангровые заросли и солончаки . 1 (3): 193. дои : 10.1023/A:1009949411068 . S2CID 126945589 .
- ^ Бос, Арканзас; Гуманао, Г.С.; Ван Катвейк, ММ; Мюллер, Б.; Саседа, ММ; Техада, РЛ (2010). «Онтогенетическое изменение среды обитания, рост популяции и роющее поведение индо-тихоокеанской пляжной звезды Archaster typicus (Echinodermata; Asteroidea)» . Морская биология . 158 (3): 639–648. дои : 10.1007/s00227-010-1588-0 . ПМЦ 3873073 . ПМИД 24391259 .
- ^ Энциклопедия Энкарта 2005 . «Берег моря», Хайди Непф .
- ^ Сков, МВт; Хартнолл, Р.Г. (2002). «Парадоксальное избирательное кормление на диете с низким содержанием питательных веществ: почему мангровые крабы едят листья?». Экология . 131 (1): 1–7. Бибкод : 2002Oecol.131....1S . дои : 10.1007/s00442-001-0847-7 . ПМИД 28547499 . S2CID 23407273 .
- ^ Абрантес, КГ; Джонстон, Р.; Коннолли, РМ; Шивс, М. (2015). «Значение углерода мангровых деревьев для водных пищевых сетей во влажных и сухих тропических эстуариях». Эстуарии и побережья . 38 (1): 383–399. Бибкод : 2015EstCo..38..383A . дои : 10.1007/s12237-014-9817-2 . hdl : 10072/141734 . ISSN 1559-2731 . S2CID 3957868 .
- ^ Муро-Торрес, Виктор М.; Амескуа, Фелипе; Сото-Хименес, Мартин; Баларт, Эдуардо Ф.; Сервьер-Сарагоса, Элиза; Грин, Люсинда; Райнохова, Яна (5 ноября 2020 г.). «Первичные источники и структура пищевой сети тропического водно-болотного угодья с высокой плотностью мангровых лесов» . Вода . 12 (11): 3105. дои : 10.3390/w12113105 . hdl : 1854/LU-01HV3XGJPZJE3Z72394VV0MRJB . ISSN 2073-4441 .
- ^ Ньюман, Сп; Хэнди, Роуд; Грубер, Ш (5 января 2010 г.). «Питание и предпочтения в отношении добычи молодых лимонных акул Negaprion brevirostris» . Серия «Прогресс в области морской экологии» . 398 : 221–234. Бибкод : 2010MEPS..398..221N . дои : 10.3354/meps08334 . ISSN 0171-8630 .
- ^ Гупта, Словакия; Гоял, MR (2017). Управление засолением почвы в сельском хозяйстве: технологические достижения и применение . ЦРК Пресс. ISBN 978-1-315-34177-4 .
- ^ Мохд-Тайб, Фарах Шафавати; Мохд-Салех, Варда; Асыха, Роша; Мансор, Мохаммад Сайфул; Ахмад-Мустафа, Муззина; Мустафа-Бакрай, Нур-Акила; Мод-Хусин, Шахрил; Мд-Шукор, Айса; Амат-Дарбис, Нурул Дарсани; Сулейман, Норела (июнь 2020 г.). «Влияние антропогенного воздействия на видовые комплексы птиц в глухих мангровых лесах» . Экология и управление водно-болотными угодьями . 28 (3): 479–494. Бибкод : 2020WetEM..28..479M . дои : 10.1007/s11273-020-09726-z . ISSN 0923-4861 . S2CID 218484236 .
- ^ Перейти обратно: а б с д и ж Вейн, Швейцария; Ким, AW; Мосс-Хейс, В.; Снейп, CE; Диас, MC; Хан, Н.С.; Энгельхарт, SE; Хортон, BP (2013). «Деградация тканей мангровых зарослей древесными термитами ( Nasutitermes acajutlae ) и их роль в цикле углерода мангровых деревьев (Пуэрто-Рико): химическая характеристика и происхождение органических веществ с использованием объемного δ13C, C/N, щелочного окисления CuO-GC/MS и твердого состояние" . Геохимия, геофизика, геосистемы . 14 (8): 3176. Бибкод : 2013ГГГ....14.3176В . дои : 10.1002/ggge.20194 .
- ^ Верстиг, Дж.Дж.; и др. (2004). «Тараксерол и пыльца ризофоры как индикаторы для отслеживания прошлых мангровых экосистем». Geochimica et Cosmochimica Acta . 68 (3): 411–22. Бибкод : 2004GeCoA..68..411V . дои : 10.1016/S0016-7037(03)00456-3 .
- ^ Гамильтон, ЮВ; Фрисс, Д.А. (2018). «Глобальные запасы углерода и потенциальные выбросы из-за вырубки мангровых лесов с 2000 по 2012 год». Природа Изменение климата . 8 (3): 240–244. arXiv : 1611.00307 . Бибкод : 2018NatCC...8..240H . дои : 10.1038/s41558-018-0090-4 . S2CID 89785740 .
- ^ Хочард, JP; Гамильтон, С.; Барбье, Э.Б. (2019). «Мангровые заросли защищают прибрежную экономическую деятельность от циклонов» . Труды Национальной академии наук . 116 (25): 12232–12237. Бибкод : 2019PNAS..11612232H . дои : 10.1073/pnas.1820067116 . ПМК 6589649 . ПМИД 31160457 .
- ^ Перейти обратно: а б с Пурахонг, Витун; Орру, Луиджи; Донати, Ирен; Перпетуини, Джорджия; Челлини, Антонио; Ламонтанара, Антонелла; Микелотти, Ваня; Таккони, Джанни; Спинелли, Франческо (2018). «Микробиом растений и его связь со здоровьем растений: виды-хозяева, органы и Pseudomonas syringae pv. Инфекция Actinidiae, формирующая бактериальные филлосферные сообщества растений киви» . Границы в науке о растениях . 9 : 1563. doi : 10.3389/fpls.2018.01563 . ПМК 6234494 . ПМИД 30464766 .
- ^ Афзал, А.; Бано, А. (2008). «Ризобии и бактерии, солюбилизирующие фосфаты, улучшают урожайность и поглощение фосфора пшеницей ( Triticum aestivum )» . Международный журнал сельского хозяйства и биологии (Пакистан) . 10 (1): 85–88. eISSN 1814-9596 . ISSN 1560-8530 .
- ^ Перейти обратно: а б с Басби, Пози Э.; Соман, Чинмей; Вагнер, Мэгги Р.; Фризен, Марен Л.; Кремер, Джеймс; Беннетт, Элисон; Морси, Мустафа; Эйзен, Джонатан А.; Лич, Ян Э.; Дангл, Джеффри Л. (2017). «Приоритеты исследований по использованию растительных микробиомов в устойчивом сельском хозяйстве» . ПЛОС Биология . 15 (3): e2001793. дои : 10.1371/journal.pbio.2001793 . ПМК 5370116 . ПМИД 28350798 .
- ^ Перейти обратно: а б Берендсен, Роланд Л.; Питерс, Корне MJ; Баккер, Питер АХМ (2012). «Ризосферный микробиом и здоровье растений». Тенденции в науке о растениях . 17 (8): 478–486. Бибкод : 2012TPS....17..478B . doi : 10.1016/j.tplants.2012.04.001 . hdl : 1874/255269 . ПМИД 22564542 . S2CID 32900768 .
- ^ Перейти обратно: а б Брингель, Франсуаза; Куэ, Иван (2015). «Основная роль микроорганизмов филлосферы на стыке между функционированием растений и динамикой газовых примесей в атмосфере» . Границы микробиологии . 06 : 486. дои : 10.3389/fmicb.2015.00486 . ПМК 4440916 . ПМИД 26052316 .
- ^ Коулман-Дерр, Девин; Дегаренн, Дамарис; Фонсека-Гарсия, Читлали; Гросс, Стивен; Клингенпил, Скотт; Войке, Таня; Норт, Гретхен; Визель, Аксель; Партида-Мартинес, Лейла П.; Тринге, Сюзанна Г. (2016). «Растительный компартмент и биогеография влияют на состав микробиома культивируемых и местных видов агавы» . Новый фитолог . 209 (2): 798–811. дои : 10.1111/nph.13697 . ПМК 5057366 . ПМИД 26467257 .
- ^ Креггер, Массачусетс; Вич, AM; Ян, ЗК; Крауч, MJ; Вилгалис, Р.; Тускан, Джорджия; Шадт, CW (2018). «Голобионт Populus: анализ влияния растительных ниш и генотипа на микробиом» . Микробиом . 6 (1): 31. дои : 10.1186/s40168-018-0413-8 . ПМК 5810025 . ПМИД 29433554 .
- ^ Хаккар, Стефан (2016). «Распутывание факторов, формирующих состав микробиоты холобионта растений» . Новый фитолог . 209 (2): 454–457. дои : 10.1111/nph.13760 . hdl : 11858/00-001M-0000-002B-166F-5 . ПМИД 26763678 .
- ^ Перейти обратно: а б Пурахонг, Витун; Садубсарн, Долая; Танунчай, Бенджаван; Вахдан, Сара Фарид Мохамед; Сансупа, Чакрия; Нолл, Матиас; Ву, Юй-Тин; Бускот, Франсуа (2019). «Первые сведения о микробиоме мангрового дерева выявляют значительные различия в таксономическом и функциональном составе между растительными и почвенными отсеками» . Микроорганизмы . 7 (12): 585. doi : 10.3390/microorganisms7120585 . ПМК 6955992 . ПМИД 31756976 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б с д и ж Татои, Хрудаянатх; Бехера, Бикаш Чандра; Мишра, Рашми Ранджан; Дутта, Сушил Кумар (2013). «Биоразнообразие и биотехнологический потенциал микроорганизмов мангровых экосистем: обзор» . Анналы микробиологии . 63 : 1–19. дои : 10.1007/s13213-012-0442-7 . S2CID 17798850 .
- ^ Перейти обратно: а б Лю, Синъюй; Ян, Чао; Ю, Сяоли; Ю, Хуан; Чжуан, Вэй; Гу, Ханг; Сюй, Куй; Чжэн, Сяфэй; Ван, Ченг; Сяо, Фаншу; Ву, Бо; Он, Чжили; Ян, Цинъюнь (2020). «Выявление структуры и сборки диазотрофного сообщества ризофитов-эндофитов в экосистеме мангровых зарослей после интродукции Sonneratia apetala и Laguncularia Racemosa». Наука об общей окружающей среде . 721 : 137807. Бибкод : 2020ScTEn.72137807L . doi : 10.1016/j.scitotenv.2020.137807 . ПМИД 32179356 . S2CID 212739128 .
- ^ Сюй, Цзинь; Чжан, Триведи, Панкай; Лю, Синь, Цзилиан; Колетта-Фильо, Куберо, Хайме; Сяолин, Вероника; Чжун, Балиан; Капоте, Виттория; Верньер, Кристиан; Аль-Сади, Абдулла М.; Цзинь, Тао, Ван, , ( ) 2018 Нянь . Ян Фан, Сюнь , 2018NatCo 9.4894X doi : 10.1038 . PMC 6244077 . / s41467-018-07343-2 ...
- ^ Перейти обратно: а б с д и ж г Дуран, Палома; Тиргарт, Торстен; Гарридо-Отер, Рубен; Аглер, Мэтью; Кемен, Эрик; Шульце-Леферт, Пол; Хаккар, Стефан (2018). «Взаимодействия микробных царств в корнях способствуют выживанию арабидопсиса» . Клетка . 175 (4): 973–983.e14. дои : 10.1016/j.cell.2018.10.020 . ПМК 6218654 . PMID 30388454 .
- ^ Сасс, Джоэль; Мартинойя, Энрико; Нортен, Трент (2018). «Накормите своих друзей: формируют ли растительные экссудаты корневой микробиом?» (PDF) . Тенденции в науке о растениях . 23 (1). Эльзевир Б.В.: 25–41. Бибкод : 2018TPS....23...25S . doi : 10.1016/j.tplants.2017.09.003 . ISSN 1360-1385 . ОСТИ 1532289 . ПМИД 29050989 . S2CID 205455681 .
- ^ Перейти обратно: а б с Байс, Харш П.; Вейр, Тиффани Л.; Перри, Лаура Г.; Гилрой, Саймон; Виванко, Хорхе М. (2006). «Роль корневых выделений во взаимодействии ризосферы с растениями и другими организмами». Ежегодный обзор биологии растений . 57 : 233–266. doi : 10.1146/annurev.arplant.57.032905.105159 . ПМИД 16669762 .
- ^ Перейти обратно: а б с Чжуан, Вэй; Ху, Жуйвэнь; Ло, Чивэнь; Чжэн, Сяо, Фаньшу; Хэ, Цян, Юн; Шу, Лунфэй; Ван, Ченг; Хэ, Чжили (2020). сообществ, связанных с корнями мангровых деревьев, в непрерывном мелкомасштабном масштабе» . и микробиомы . npj Биопленки «Разнообразие, функции и сборка микробных 52. дои : 10.1038 s41522-020-00164-6 PMC 7665043. 33184266 PMID . / Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б с Лай, Цзяюн; Чеа, Ви; Паланивело, Кишнеф; Сува, Ремпей; Шарма, Сахадев (16 декабря 2022 г.). «Систематический обзор физико-химического и микробного разнообразия хорошо сохранившихся, восстановленных и нарушенных мангровых лесов: что известно и каков путь вперед?» . Леса . 13 (12): 2160. дои : 10.3390/f13122160 .
- ^ Шрикант, Сандхья; Лам, Шон Кайхекулани Ямаути; Чен, Чжун (2016). «Корень мангрового дерева: адаптации и экологическое значение». Деревья . 30 (2): 451–465. Бибкод : 2016Деревья..30..451S . дои : 10.1007/s00468-015-1233-0 . S2CID 5471541 .
- ^ Макки, Карен Л. (1993). «Физико-химические закономерности почвы и распространение видов мангровых зарослей - взаимные эффекты?». Журнал экологии . 81 (3): 477–487. Бибкод : 1993JEcol..81..477M . дои : 10.2307/2261526 . JSTOR 2261526 .
- ^ Ольгин, Джина; Васкес, Патрисия; Башан, Йоав (2001). «Роль донных микроорганизмов в продуктивности, сохранении и восстановлении мангровых экосистем: обзор». Биология и плодородие почв . 33 (4): 265–278. Бибкод : 2001BioFS..33..265H . дои : 10.1007/s003740000319 . S2CID 10826862 .
- ^ Риф, Р.; Феллер, IC; Лавлок, CE (2010). «Питание мангровых зарослей» . Физиология дерева . 30 (9): 1148–1160. doi : 10.1093/treephys/tpq048 . ПМИД 20566581 .
- ^ Се, Сянъюй; Венг, Босен; Цай, Бангпинг; Донг, Иран; Ян, Чунлин (2014). «Влияние арбускулярной микоризной инокуляции и подачи фосфора на рост и усвоение питательных веществ сеянцев Kandelia obovata (Sheue, Liu & Yong) в автоклавированной почве». Прикладная экология почв . 75 : 162–171. Бибкод : 2014AppSE..75..162X . дои : 10.1016/j.apsoil.2013.11.009 .
- ^ Эдвардс, Джозеф; Джонсон, Кэмерон; Сантос-Медельин, Кристиан; Лурье, Евгений; Подишетти, Натрадж Кумар; Бхатнагар, Шриджак; Эйзен, Джонатан А.; Сундаресан, Венкатесан (20 января 2015 г.). «Структура, вариации и сборка корневых микробиомов риса» . Труды Национальной академии наук . 112 (8): Е911–Е920. Бибкод : 2015PNAS..112E.911E . дои : 10.1073/pnas.1414592112 . ISSN 0027-8424 . ПМЦ 4345613 . ПМИД 25605935 .
- ^ Перейти обратно: а б с Эдвардс, Джозеф; Джонсон, Кэмерон; Сантос-Медельин, Кристиан; Лурье, Евгений; Подишетти, Натрадж Кумар; Бхатнагар, Шриджак; Эйзен, Джонатан А.; Сундаресан, Венкатесан (2015). «Структура, вариации и сборка корневых микробиомов риса» . Труды Национальной академии наук . 112 (8): Е911–Е920. Бибкод : 2015PNAS..112E.911E . дои : 10.1073/pnas.1414592112 . ПМЦ 4345613 . ПМИД 25605935 .
- ^ Хартман, Кайл; Тринге, Сюзанна Г. (2019). «Взаимодействие между растениями и почвой, формирующее корневой микробиом в условиях абиотического стресса» . Биохимический журнал . 476 (19): 2705–2724. дои : 10.1042/BCJ20180615 . ПМК 6792034 . ПМИД 31654057 .
- ^ Рейнхольд-Хурек, Барбара; Бюнгер, Вибке; Бурбано, Клаудия София; Сабале, Мугдха; Хурек, Томас (2015). «Корни, формирующие их микробиом: глобальные горячие точки микробной активности». Ежегодный обзор фитопатологии . 53 : 403–424. doi : 10.1146/annurev-phyto-082712-102342 . ПМИД 26243728 .
- ^ Лю, Ялун; Ге, Тида; Да, Джун; Лю, Шулун; Шибистова, Ольга; Ван, Пин; Ван, Цзинкуань; Ли, Юн; Гуггенбергер, Георг; Кузяков, Яков ; Ву, Цзиньшуй (2019). «Первоначальное использование ризоотложений при выращивании риса на рисовых почвах: эффекты ризосферы и азотных удобрений». Геодерма . 338 : 30–39. Бибкод : 2019Geode.338...30L . дои : 10.1016/j.geoderma.2018.11.040 . S2CID 134648694 .
- ^ Йоханссон, Йонас Ф.; Пол, Лесли Р.; Финли, Роджер Д. (2004). «Микробные взаимодействия в микоризосфере и их значение для устойчивого сельского хозяйства» . ФЭМС Микробиология Экология . 48 (1): 1–13. Бибкод : 2004FEMME..48....1J . дои : 10.1016/j.femsec.2003.11.012 . ПМИД 19712426 . S2CID 22700384 .
- ^ Перейти обратно: а б Сасс, Джоэль; Мартинойя, Энрико; Нортен, Трент (2018). «Накормите своих друзей: формируют ли растительные экссудаты корневой микробиом?» (PDF) . Тенденции в науке о растениях . 23 (1): 25–41. Бибкод : 2018TPS....23...25S . doi : 10.1016/j.tplants.2017.09.003 . ОСТИ 1532289 . ПМИД 29050989 . S2CID 205455681 .
- ^ Перейти обратно: а б Офек-Лалзар, Майя; Села, Ноа; Гольдман-Воронов, Милана; Грин, Стефан Дж.; Хадар, Ицхак; Минц, Дрор (2014). «Нишевые и связанные с хозяином функциональные признаки микробиома поверхности корня» . Природные коммуникации . 5 : 4950. Бибкод : 2014NatCo...5.4950O . дои : 10.1038/ncomms5950 . ПМИД 25232638 .
- ^ Лю, Юн-Синь; Цинь, Юань; Чен, Тонг; Лу, Мэйпин; Цянь, Сюбо; Го, Сяосюань; Бай, Ян (2021). «Практическое руководство по ампликонному и метагеномному анализу данных микробиома» . Белок и клетка . 12 (5): 315–330. дои : 10.1007/s13238-020-00724-8 . ПМК 8106563 . ПМИД 32394199 .
- ^ Котта, Симоне Рапозо; Кадет, Луана Лира; Ван Эльзас, Ян Дирк; Андреоте, Фернандо Дини; Диас, Армандо Кавальканте Франко (2019). «Изучение функциональности бактерий в отложениях мангровых зарослей и их способности преодолевать антропогенную деятельность» . Бюллетень о загрязнении морской среды . 141 : 586–594. Бибкод : 2019MarPB.141..586C . doi : 10.1016/j.marpolbul.2019.03.001 . ПМИД 30955771 . S2CID 91872087 .
- ^ Перейти обратно: а б с д и ж Джин, Мин; Го, Сюнь; Чжан, Руй; Цюй, Ву; Гао, Болян; Цзэн, Руньин (2019). «Разнообразие и потенциальное биогеохимическое воздействие вирусов мангровых почв» . Микробиом . 7 (1): 58. дои : 10.1186/s40168-019-0675-9 . ПМК 6460857 . ПМИД 30975205 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Саттл, Кертис А. (2005). «Вирусы в море». Природа . 437 (7057): 356–361. Бибкод : 2005Natur.437..356S . дои : 10.1038/nature04160 . ПМИД 16163346 . S2CID 4370363 .
- ^ Холмфельдт, К.; Солоненко Н.; Шах, М.; Корье, К.; Риман, Л.; Верберкмоэс, Северная Каролина; Салливан, МБ (2013). «Двенадцать ранее неизвестных родов фагов повсеместно распространены в мировом океане» . Труды Национальной академии наук . 110 (31): 12798–12803. Бибкод : 2013PNAS..11012798H . дои : 10.1073/pnas.1305956110 . ПМЦ 3732932 . ПМИД 23858439 .
- ^ Симе-Нгандо, Телесфор (2014). «Экологические бактериофаги: Вирусы микробов в водных экосистемах» . Границы микробиологии . 5 : 355. дои : 10.3389/fmicb.2014.00355 . ПМК 4109441 . ПМИД 25104950 .
- ^ Перейти обратно: а б Брейтбарт, Майя (2012). «Морские вирусы: правда или действие». Ежегодный обзор морской науки . 4 : 425–448. Бибкод : 2012ARMS....4..425B . doi : 10.1146/annurev-marine-120709-142805 . ПМИД 22457982 .
- ^ Он, Тяньлян; Ли, Хунъюн; Чжан, Сяобо (2017). «Вирусы глубоководных гидротермальных источников компенсируют микробный метаболизм во взаимодействиях вирус-хозяин» . мБио . 8 (4). дои : 10.1128/mBio.00893-17 . ПМК 5513705 . ПМИД 28698277 .
- ^ Гурвиц, БЛ; Вествельд, АХ; Брум, младший; Салливан, МБ (2014). «Моделирование экологических факторов в морских вирусных сообществах с использованием сравнительной метагеномики и сетевого анализа» . Труды Национальной академии наук . 111 (29): 10714–10719. Бибкод : 2014PNAS..11110714H . дои : 10.1073/pnas.1319778111 . ПМЦ 4115555 . ПМИД 25002514 .
- ^ Анантараман, Картик; Дюэм, Мелисса Б.; Брейер, Джон А.; Вендт, Кэтлин А.; Тонер, Брэнди М.; Дик, Грегори Дж. (2014). «Гены окисления серы в разнообразных глубоководных вирусах». Наука . 344 (6185): 757–760. Бибкод : 2014Sci...344..757A . дои : 10.1126/science.1252229 . HDL : 1912/6700 . ПМИД 24789974 . S2CID 692770 .
- ^ Перейти обратно: а б Йорк, Эшли (2017). «Вирус водорослей увеличивает поглощение азота в океане» . Обзоры природы Микробиология . 15 (10): 573. doi : 10.1038/nrmicro.2017.113 . ПМИД 28900307 . S2CID 19473466 .
- ^ Ру, Саймон; Брум, Дженнифер Р.; Дутил, Лоу Э.; Сунагава, Синъити; Дюэм, Мелисса Б.; Лой, Александр; Пулос, Бонни Т.; Солоненко, Наталья; Лара, Елена; Пулен, Жюли; Пезан, Стефан; Кандельс-Льюис, Стефани; Димье, Селин; Пишераль, Марк; Сирсон, Сара; Круо, Коринн; Альберти, Адриана; Дуарте, Карлос М.; Газоль, Джозеф М.; Ваке, Боли; Борк, Пер; Ацинас, Сильвия Г.; Винкер, Патрик; Салливан, Мэтью Б. (2016). «Экогеномика и потенциальное биогеохимическое воздействие распространенных во всем мире океанических вирусов». Природа . 537 (7622): 689–693. Бибкод : 2016Natur.537..689. . дои : 10.1038/nature19366 . hdl : 1874/341494 . ПМИД 27654921 . S2CID 54182070 .
- ^ Ровер, Форест; Тербер, Ребекка Вега (2009). «Вирусы манипулируют морской средой». Природа . 459 (7244): 207–212. Бибкод : 2009Natur.459..207R . дои : 10.1038/nature08060 . ПМИД 19444207 . S2CID 4397295 .
- ^ Салливан, Мэтью Б.; Линделл, Дебби ; Ли, Джессика А.; Томпсон, Люк Р.; Белявский, Джозеф П.; Чисхолм, Салли В. (2006). «Распространенность и эволюция генов основной фотосистемы II в морских цианобактериальных вирусах и их хозяевах» . ПЛОС Биология . 4 (8): е234. дои : 10.1371/journal.pbio.0040234 . ПМЦ 1484495 . ПМИД 16802857 .
- ^ Томпсон, ЛР; Цзэн, К.; Келли, Л.; Хуанг, К.Х.; Сингер, Австралия; Стуббе, Дж.; Чисхолм, Юго-Запад (2011). «Вспомогательные метаболические гены фага и перенаправление углеродного метаболизма цианобактерий-хозяев» . Труды Национальной академии наук . 108 (39): Е757–Е764. дои : 10.1073/pnas.1102164108 . ПМК 3182688 . ПМИД 21844365 .
- ^ Цзэн, Цинлу; Чисхолм, Салли В. (2012). «Морские вирусы используют двухкомпонентную систему регулирования своего хозяина в ответ на ограничение ресурсов» . Современная биология . 22 (2): 124–128. Бибкод : 2012CBio...22..124Z . дои : 10.1016/j.cub.2011.11.055 . hdl : 1721.1/69047 . ПМИД 22244998 . S2CID 7692657 .
- ^ Фрэнк, Джереми А.; Лоример, Дон; Юл, Мерри; Витте, Пэм; Крейг, Тим; Абендрот, Ян; Ровер, Форест; Эдвардс, Роберт А.; Сигалл, Анка М.; Бургин, Алекс Б. (2013). «Структура и функция пептидной деформилазы, кодируемой цианофагом» . Журнал ISME . 7 (6): 1150–1160. Бибкод : 2013ISMEJ...7.1150F . дои : 10.1038/ismej.2013.4 . ПМЦ 3660681 . ПМИД 23407310 .
- ^ Юсеф, Сибу; и др. (2007). «Глобальная экспедиция по отбору проб океана Sorcerer II: расширение Вселенной белковых семейств» . ПЛОС Биология . 5 (3): е16. дои : 10.1371/journal.pbio.0050016 . ПМК 1821046 . ПМИД 17355171 .
- ^ Динсдейл, Элизабет А.; Эдвардс, Роберт А.; Холл, Дана; Англи, Флоран; Брейтбарт, Майя; Брюлк, Дженнифер М.; Фурлан, Майк; Десю, Кристель; Хейнс, Мэтью; Ли, Линлин; Макдэниел, Лорен; Моран, Мэри Энн; Нельсон, Карен Э.; Нильссон, Кристина; Олсон, Роберт; Пол, Джон; Брито, Бельтран Родригес; Жуань, Ицзюнь; Свон, Брэндон К.; Стивенс, Рик; Валентин, Дэвид Л.; Тербер, Ребекка Вега; Уэгли, Линда; Уайт, Брайан А.; Ровер, Форест (2008). «Функциональное метагеномное профилирование девяти биомов». Природа . 452 (7187): 629–632. Бибкод : 2008Natur.452..629D . дои : 10.1038/nature06810 . ПМИД 18337718 . S2CID 4421951 .
- ^ Розенвассер, Шило; Зив, Кармит; Кревелд, Шири Графф ван; Варди, Ассаф (2016). «Метаболизм вироклеток: метаболические инновации во время взаимодействия хозяина и вируса в океане». Тенденции в микробиологии . 24 (10): 821–832. дои : 10.1016/j.tim.2016.06.006 . ПМИД 27395772 .
- ^ Гурвиц, Бонни Л.; Брам, Дженнифер Р.; Салливан, Мэтью Б. (2015). «Глубинно-слоистая функциональная и таксономическая специализация ниш в «основном» и «гибком» вироме Тихого океана» . Журнал ISME . 9 (2): 472–484. Бибкод : 2015ISMEJ...9..472H . дои : 10.1038/ismej.2014.143 . ПМЦ 4303639 . ПМИД 25093636 .
- ^ Уоммак, К. Эрик; Колвелл, Рита Р. (2000). «Вириопланктон: вирусы в водных экосистемах» . Обзоры микробиологии и молекулярной биологии . 64 (1): 69–114. дои : 10.1128/ММБР.64.1.69-114.2000 . ПМК 98987 . ПМИД 10704475 .
- ^ Перейти обратно: а б Алонги, Дэниел М. (2012). «Связывание углерода в мангровых лесах» . Управление выбросами углерода . 3 (3): 313–322. Бибкод : 2012CarM....3..313A . дои : 10.4155/cmt.12.20 . S2CID 153827173 .
- ^ Дженнерджан, Тим С.; Иттеккот, Венугопалан (2002). «Значение мангровых зарослей для производства и отложения органических веществ вдоль тропических континентальных окраин». Naturwissenschaften . 89 (1): 23–30. Бибкод : 2002NW.....89...23J . дои : 10.1007/s00114-001-0283-x . ПМИД 12008969 . S2CID 33556308 .
- ^ Алонги, Дэниел М.; Клаф, Барри Ф.; Диксон, Пол; Тиренди, Фрэнк (2003). «Распределение и хранение питательных веществ в лесах засушливой зоны мангровых зарослей Rhizophora stylosa и Avicennia marina». Деревья . 17 (1): 51–60. Бибкод : 2003Деревья..17...51А . дои : 10.1007/s00468-002-0206-2 . S2CID 23613917 .
- ^ Алонги, Дэниел М. (2014). «Круговорот и хранение углерода в мангровых лесах» . Ежегодный обзор морской науки . 6 : 195–219. Бибкод : 2014ARMS....6..195A . doi : 10.1146/annurev-marine-010213-135020 . ПМИД 24405426 .
- ^ Натараджан, Пурушотаман; Муругесан, Ашок Кумар; Говиндан, Ганесан; Гопалакришнан, Айяру; Кумар, Равичандиран; Дурайсами, Пурушотаман; Баладжи, Раджу; Шьямли, Пухан Сушри; Парида, Аджай К.; Парани, Мадасами (8 июля 2021 г.). «Геном эталонного уровня идентифицирует гены солеустойчивости у солевыделяющих видов мангровых зарослей Avicennia marina» . Коммуникационная биология . 4 (1). Springer Science and Business Media LLC: 851. doi : 10.1038/s42003-021-02384-8 . ISSN 2399-3642 . ПМЦ 8266904 . ПМИД 34239036 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Марсиал Гомес, Ньютон К.; Борхес, Людмила Р.; Параньос, Родольфо; Пинто, Фернандо Н.; Мендонаса-Хаглер, Леда К.С.; Смолла, Корнелия (2008). «Изучение разнообразия бактериальных сообществ в отложениях городских мангровых лесов». ФЭМС Микробиология Экология . 66 (1): 96–109. Бибкод : 2008FEMME..66...96M . дои : 10.1111/j.1574-6941.2008.00519.x . ПМИД 18537833 . S2CID 40733636 .
- ^ Андреоте, Фернандо Дини; Хименес, Диего Хавьер; Чавес, Диего; Диас, Армандо Кавальканте Франко; Лувизотто, Дэнис Маззер; Дини-Андреоте, Франциско; Фасанелла, Кристиан Чипола; Лопес, Мариейми Варон; Баэна, Сандра; Такетани, Родриго Гувеа; Де Мело, Итамар Соарес (2012). «Микробиом отложений бразильских мангровых зарослей, выявленный метагеномикой» . ПЛОС ОДИН . 7 (6): е38600. Бибкод : 2012PLoSO...738600A . дои : 10.1371/journal.pone.0038600 . ПМК 3380894 . ПМИД 22737213 .
- ^ Риклефс, Роберт Э.; Шлютер, Дольф (1993). Видовое разнообразие экологических сообществ: историко-географические перспективы . Издательство Чикагского университета. ISBN 9780226718231 .
- ^ Пратама, Акбар Аджи; Ван Эльзас, Ян Дирк (2018). «Заброшенный» почвенный виром – потенциальная роль и влияние». Тенденции в микробиологии . 26 (8): 649–662. дои : 10.1016/j.tim.2017.12.004 . ПМИД 29306554 . S2CID 25057850 .
- ^ Уильямсон, Курт Э.; Фурманн, Джеффри Дж.; Уоммак, К. Эрик; Радосевич, Марк (2017). «Вирусы в почвенных экосистемах: неизвестное количество на неизведанной территории» . Ежегодный обзор вирусологии . 4 (1): 201–219. doi : 10.1146/annurev-virology-101416-041639 . ПМИД 28961409 .
- ^ Лян, Цзюнь-Бин; Чен, Юэ-Цинь; Лан, Чонг-Ю; Тэм, Нора Ф.Ю.; Зан, Ци-Цзе; Хуан, Ли-Нан (2007). «Восстановление нового бактериального разнообразия из отложений мангровых зарослей». Морская биология . 150 (5): 739–747. Бибкод : 2007МарБи.150..739Л . дои : 10.1007/s00227-006-0377-2 . S2CID 85384181 .
- ^ Сюй, Шаохуа; Чжан, Чжан; Го, Го, Уся; Ли, Цзяньфан; Ду, Чжэнлинь; Чжоу, Жэньчао; Дэвид Э; Мануэль; Ву, Чунг-И; Дьюк, Норман С.; Ши, Сухуа (5 июня 2017 г.). «Происхождение, диверсификация и адаптация основной клады мангровых деревьев (Rhizophoreae), выявленные с помощью полногеномного секвенирования». . National Science Review . 4 (5). Издательство Оксфордского университета (OUP): 721–734. : 10.1093 /nsr/nwx065 . ISSN 2095-5138 . PMC 6599620. . PMID 31258950 doi
Дальнейшее чтение
[ редактировать ]- Сенгер, Питер (2002). Экология мангровых лесов, лесоводство и охрана . Kluwer Academic Publishers, Дордрехт. ISBN 1-4020-0686-1 .
- Таникаимони, Ганапати (1986). Палинология мангровых зарослей ПРООН / ЮНЕСКО и Французский институт Пондичерри , ISSN 0073-8336 (E).
- Томлинсон, Филип Б. (1986). Ботаника мангровых зарослей . Издательство Кембриджского университета, Кембридж, ISBN 0-521-25567-8 .
- Чаи, HJ (1983). Биология и экология мангровых зарослей . W. Junk Publishers, Гаага. ISBN 90-6193-948-8 .
- Плазиа, Жан-Клод; Кавагнетто, Карла; Кенигер, Жан-Клод; Бальцер, Фредерик (2001). «История и биогеография мангровых экосистем, основанная на критической переоценке палеонтологических данных». Экология и управление водно-болотными угодьями . 9 (3): 161–180. дои : 10.1023/А:1011118204434 . S2CID 24980831 .
- Джаятисса, LP; Даду-Гебас, Ф.; Коедам, Н. (2002). «Обзор цветочного состава и распространения мангровых зарослей в Шри-Ланке» (PDF) . Ботанический журнал Линнеевского общества . 138 : 29–43. дои : 10.1046/j.1095-8339.2002.00002.x .
- Эллисон, Аарон М. (2000). «Восстановление мангровых зарослей: достаточно ли мы знаем?». Реставрационная экология . 8 (3): 219–229. Бибкод : 2000ResEc...8..219E . дои : 10.1046/j.1526-100x.2000.80033.x . S2CID 86352384 .
- Агравала, Шардул; Хагестад; Марка; Коши, Каяту; Ота, Томоко; Прасад, Биман; Рисби, Джеймс; Смит, Джоэл; Ван Алст, Мартен. 2003. Развитие и изменение климата на Фиджи: внимание прибрежным мангровым зарослям. Организация экономического сотрудничества и развития, Париж, Cedex 16, Франция.
- Барбье, Э.Б.; Сатиратаи, С. (2001). «Ценность сохранения мангровых зарослей в Южном Таиланде». Современная экономическая политика . 19 (2): 109–122. дои : 10.1111/j.1465-7287.2001.tb00054.x .
- Босире, Дж.О.; Даду-Гебас, Ф.; Джаятисса, LP; Коедам, Н.; Ло Син, Д.; Нитто, Ди Д. (2005). «Насколько эффективны были мангровые заросли в качестве защиты от недавнего цунами?» . Современная биология . 15 (12): Р443–Р447. дои : 10.1016/j.cub.2005.06.008 . ПМИД 15964259 . S2CID 8772526 .
- Боуэн, Дженнифер Л.; Валиэла, Иван; Йорк, Джоанна К. (2001). «Мангровые леса: одна из основных тропических сред мира, находящихся под угрозой исчезновения» . Бионаука . 51 (10): 807–815. doi : 10.1641/0006-3568(2001)051[0807:mfootw]2.0.co;2 .
- Джин-Ёнг, Онг (2004). «Экология сохранения и управления мангровыми зарослями». Гидробиология . 295 (1–3): 343–351. дои : 10.1007/BF00029141 . S2CID 26686381 .
- Гленн, ЧР 2006. «Существа Земли, находящиеся под угрозой исчезновения».
- Льюис, Рой Р. III (2004). «Экологическая инженерия для успешного управления и восстановления мангровых лесов». Экологическая инженерия . 24 (4): 403–418. дои : 10.1016/j.ecoleng.2004.10.003 .
- Кюнцер, К.; Блюмел, А.; Гебхардт, С.; Во Куок Т. и Деч С. (2011). «Дистанционное зондирование мангровых экосистем: обзор» . Дистанционное зондирование . 3 (5): 878–928. Бибкод : 2011RemS....3..878K . дои : 10.3390/rs3050878 .
- Люсьен-Брун, Х (1997). «Эволюция мирового производства креветок: рыболовство и аквакультура». Мировая аквакультура . 28 : 21–33.
- Твилли, Р.Р., В.Х. Ривера-Монрой, Э. Медина, А. Найман, Дж. Форет, Т. Маллак и Л. Ботеро. 2000. Особенности развития лесов в мангровых зарослях вдоль устья реки Сан-Хуан, Венесуэла. Лесная экология и управление
- Мюррей, MR; Зисман, SA; Ферли, Пенсильвания; Манро, DM; Гибсон, Дж.; Рэттер, Дж.; Бриджуотер, С.; Мити, компакт-диск; Плейс, CJ (2003). «Мангровые заросли Белиза: Часть 1. Распространение, состав и классификация». Лесная экология и управление . 174 (1–3): 265–279. Бибкод : 2003ForEM.174..265M . дои : 10.1016/s0378-1127(02)00036-1 .
- Во Куок, Т.; Кюнцер, К.; Во Куанг, М.; Модер Ф. и Оппельт Н. (декабрь 2012 г.). «Обзор методов оценки услуг мангровых экосистем». Экологические показатели . 23 : 431–446. Бибкод : 2012EcInd..23..431V . дои : 10.1016/j.ecolind.2012.04.022 .
- Сполдинг, Марк; Кайнума, Мами и Коллинз, Лорна (2010) Мировой атлас мангровых зарослей Earthscan, Лондон, ISBN 978-1-84407-657-4 ; 60 карт, показывающих распространение мангровых зарослей по всему миру.
- Уорн, Кеннеди (2013) Пусть едят креветки: трагическое исчезновение тропических лесов моря. Айленд Пресс, 2012, ISBN 978-1597263344
- Массо; Алеман, С.; Буржуа, К.; Аппельтанс, В.; Ванхоорн, Б.; Де Хаувер, Н.; Стоффелен, П.; Хигебарт, А.; Даду-Гебас, Ф. (2010). «Справочная база данных и гербарий по мангровым деревьям » (PDF) . Экология и эволюция растений . 143 (2): 225–232. дои : 10.5091/plecevo.2010.439 .
- Во Куок, Т.; Оппельт, Н.; Лейненкугель П. и Кюнцер К. (2013). «Дистанционное зондирование при картировании мангровых экосистем – объектно-ориентированный подход» . Дистанционное зондирование . 5 (1): 183–201. Бибкод : 2013RemS....5..183В . дои : 10.3390/rs5010183 .
Внешние ссылки
[ редактировать ] Определения из Викисловаря
Определения из Викисловаря  СМИ из Commons
СМИ из Commons
- «Информационный бюллетень о мангровых зарослях» . Институт Уэйта. Архивировано из оригинала 4 сентября 2015 года . Проверено 8 июня 2015 г.
- «Мангровые заросли» . Смитсоновский океанический портал. 30 апреля 2018 г.
- 10 лучших мангровых лесов в мире
- «Информационный бюллетень о мангровых зарослях» (PDF) . Рыболовство Западной Австралии. 2013. Архивировано из оригинала (PDF) 23 апреля 2013 года. * Rhizophoraceae в Curlie.
- Мангровые леса в Керли
- В мае 2011 года специальная английская служба « Голоса Америки » транслировала 15-минутную программу о мангровых лесах. Стенограмму и MP3-файл программы, предназначенной для изучающих английский язык, можно найти на сайте « Мангровые леса могут стать крупным игроком в торговле выбросами углерода».
- «Водный центр влажных тропиков Латинской Америки и Карибского бассейна» . Архивировано из оригинала 5 февраля 2012 года . Проверено 25 января 2014 г.
- «Программа просмотра океанических данных – ЮНЕП-WCMC» . Официальный сайт ЮНЕП-ВЦМК – Ocean Data Viewer . Проверено 27 ноября 2020 г.
- Прибрежные почки Квинсленда: мангровые заросли . Стейси Ларнер, блог библиотеки Джона Оксли. Государственная библиотека Квинсленда.
- «Возьмите убежище – мангровые деревья работают вместе, чтобы защитить Землю и ее воды. Чему они могут научить нас о сообществе и жертвоприношениях?» . Атмосфера . 16 февраля 2024 г.


