Вымирание солончаков

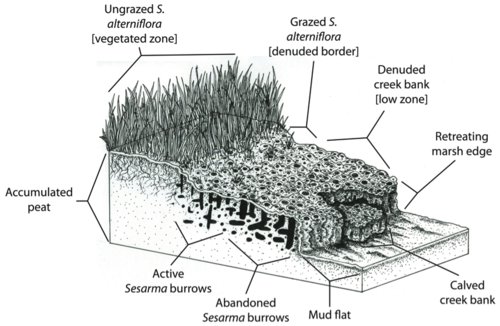
Вымирание солончаков - это термин, который использовался в США и Великобритании для описания гибели солончаков кордовой травы , ведущей к последующей деградации среды обитания, особенно в низинных зонах солончаков на побережьях Западной Атлантики. Кордграсс обычно закрепляет отложения в солончаках; его потеря приводит к снижению твердости субстрата, усилению эрозии и обрушению берегов ручьев в воду, что в конечном итоге приводит к снижению здоровья и продуктивности болот.
Отмирание может затронуть несколько видов кордграсса и (род Spartina ), включая S. alterniflora , S. densiflora S. Townsendii . Существует несколько конкурирующих гипотез, предсказывающих причины и механизмы вымирания солончаков по всей западной Атлантике. Эти гипотезы уделяют разное внимание влиянию нисходящих или восходящих процессов на вымирание солончаков. В сочетании с вымиранием солончаков в верховьях вымирание солончаков представляет собой серьезную угрозу для экосистемных услуг , которые болота предоставляют местным прибрежным сообществам.
История «сверху вниз» и «снизу вверх»
[ редактировать ]В свете своего влияния на общественные процессы, поведение и экологические взаимодействия, потребительские взаимодействия являются одними из наиболее широко изучаемых концепций в экологии . По этой причине ученые используют пищевые сети, чтобы изобразить все пищевые цепи и трофические отношения в экологическом сообществе .

Пищевые сети могут контролироваться силами, восходящими или нисходящими, которые определяют, регулируются ли структура пищевой сети и динамика популяции питательными веществами (запас фиксированного углерода ) и первичным производством или высшими хищниками , соответственно.
Большая часть энергии теряется при переходе от одного трофического уровня к другому (около 90%); следовательно, успех более высоких уровней связан с более низкими и их снабжением ресурсами (Lindeman 1942). [1] Однако на численность и распределение организмов в экосистеме также влияет плотность потребителей, которая ограничивает успех организмов на более низких трофических уровнях и тем самым влияет на численность этих организмов (Hairston et al. 1960). [2]
Многие экологи утверждают, что контроль снизу вверх и сверху вниз не играет одинаково важной роли в структуре и динамике популяций в экосистеме; однако данные свидетельствуют о том, что как восходящие, так и нисходящие силы влияют на структуру пищевых сетей, а также на пространственную и временную ( Бертнесс 2007 ) численность и распределение организмов , [3] хотя в какой степени каждый из них играет роль, до конца не понятно.
Исторически в экологической мысли преобладал акцент на контроле снизу вверх во многих экосистемах, часто исключая потребительский контроль (Strong 1992). [4] Во многих экосистемах, в которых потребительский контроль классически считался тривиальным, преобладают растения (например, леса, луга и солончаки) и они обычно имеют зеленый цвет. В 1960 году Хейрстон и его коллеги предложили противоположную точку зрения, в которой особое внимание уделялось контролю над потребителями. Они утверждали, что «мир зеленый», потому что более высокие трофические уровни регулируют численность травоядных (Hairston et al. 1960). [2]
Критики отмечали, что мир не всегда зеленый, и что когда это так, травоядные животные не обязательно играют важную роль в структурировании растительных сообществ (Ehrlich and Birch, 1967). [5] Другие утверждали, что зеленое не всегда съедобно или имеет достаточно высокое качество, чтобы позволить увеличить популяцию травоядных (Dixon 1966, Murdoch 1966). [6] [7] Споры продолжаются, но среди экологов по-прежнему преобладает мнение, что, хотя потребители влияют на многие аспекты продуктивности растений и экологии, контроль сверху вниз не влияет на продуктивность целых растительных экосистем.
Однако совсем недавно примеры явного контроля потребителей над целыми экосистемами появились в различных средах обитания, включая озера (Carpenter et al. 1985), [8] реки (Пауэр 1992), [9] и морской (Эстес и Даггинс, 1995). [10] места обитания. Основные виды растений могут быть заменены другими видами или субстратом, полностью лишенным растительности , а насекомые могут уничтожить целые мангровые заросли (Феллер, 2002). [11]
Классический пример взаимодействия сверху вниз, определяющего структуру и функции сообщества, можно найти в работе Боба Пейна в Вашингтоне, который установил, что удаление морской звезды Pisaster вызвало трофический каскад, в котором популяции голубых мидий ( Mytilus ) резко возросли из-за освобождения от давления хищников ( Пейн 1966) [12]
Другой влиятельный пример нисходящего контроля появился в экспериментах Джейн Любченко на скалистых берегах Новой Англии , которые продемонстрировали, что травоядная улитка L.littorea осуществляет контроль над разнообразием и сменой сообществ водорослей бассейнов приливных (Lubchenco and Menge 1978). [13] Одна гипотеза , возникшая на основе работы Любченко (Литтл и Китчинг, 1996). [14] Было то, что хищничество зеленого краба ( Carcinus maenas ) влияет на сообщества водорослей скалистых берегов, регулируя численность L.littorea .
Экологи приводят эти примеры как свидетельство того, что регулирование прав потребителей является более мощным и преобладающим, чем считалось ранее.
Исторические парадигмы теории солончаков
[ редактировать ]
Что касается солончаков, первые экологи, такие как Юджин Одум и Джон Тил, положили начало нынешней восходящей парадигме в экологии, работая на острове Сапело, штат Джорджия (США), в которой подчеркивалась доминирующая роль физических факторов, таких как температура , соленость и питательные вещества , в регулировании первичной растительности. продуктивность и структура экосистемы (Teal 1962, Odum 1971). [15] [16] Экологи отметили, что болотные растения не подвергались интенсивному выпасу и казались относительно неприятными на вкус, и, таким образом, утверждали, что большинство растений попадают в пищевые цепи солончаков в виде детрита (Teal 1962). [15] Следствием этой догмы является то, что потребители играют незначительную или тонкую роль в контроле первичной продукции солончаков (Smalley 1960, Teal 1962). [15] [17] Эта парадигма была широко принята на протяжении десятилетий и применялась к другим экосистемам, таким как мангровые заросли и заросли морских водорослей (Бертнесс, 2007). [3] и, таким образом, «укоренился в концептуальном понимании прибрежных экосистем» (цитата из Бертнесс и Силлиман, 2008; Смолли, 1960, Никсон, 1982). [17] [18] [19]
Однако недавние работы продемонстрировали сильный нисходящий контроль над растительными сообществами солончаков со стороны широкого круга потребителей, включая улиток , крабов и гусей (Джеффрис 1997, Бортолус и Ирибарн 1999, Силлиман и Бертнесс 2002, Холдридж и др. 2009). [20] [21] [22] [23] К болотным пасущимся относятся также дикие лошади (Фурбиш и Альбано, 1994), [24] крупный рогатый скот , зайцы , насекомые и грызуны , некоторые из которых способны сильно подавлять рост растений.
Сильный контроль над болотами сверху вниз был продемонстрирован в различных болотных системах. Потребительский контроль осуществляется крабом -грапсидом ( Chasmagnathus granulata ) в солончаках Аргентины и Бразилии на атлантическом побережье Южной Америки (Bortolus and Iribarne 1999). [21] Другие эксперименты в Аргентине способствовали увеличению количества работ, подтверждающих контроль потребителей на солончаках (Альберти и др., 2007). [25] Также предполагается, что травоядные животные играют важную роль в болотах юго-западной Атлантики (Бортолус и Ирибарн, 1999, Альберти и др., 2007). [21] [25] Потребители важны не только в небольших масштабах, как показали эксперименты в клетках в различных местах (Силлиман и Зиман, 2001, Силлиман и Бертнесс, 2002, Силлиман и др., 2005), [22] [26] [27] но они также влияют на первичную продуктивность на больших географических территориях (Silliman and Zieman 2001). [26]
Бертнесс и Силлиман также предположили, что, хотя солончаки исторически могли быть ограничены питательными веществами и контролироваться снизу вверх, антропогенные нарушения, такие как эвтрофикация и истощение численности хищников, в последние десятилетия сместили эти системы в режим контроля сверху вниз (Бертнесс и Силлиман, 2008). [18]
Региональные причины вымирания солончаков
[ редактировать ]
Как нисходящие, так и восходящие силы были предложены в качестве основных причин вымирания солончаков по всей западной Атлантике. Относительная важность этих факторов зависит от абиотических и биотических условий каждой местной или региональной болотной системы и ее окружающей среды.
Взаимодействие сверху вниз
[ редактировать ]Человеческая деятельность может запускать трофические каскады (Джеффрис, 1997), [20] которые происходят, когда хищники, которые ограничивают численность добычи и тем самым повышают выживаемость следующего более низкого трофического уровня , истощаются (Strong 1992). [4] Истощение высших хищников освобождает их добычу от контроля потребителей и приводит к сокращению популяции следующего более низкого трофического уровня, часто основных производителей . Трофические каскады возникают как минимум на трех трофических уровнях и могут влиять на динамику сообществ в различных системах (Estes et al. 1998). [28] Трофические каскады могут вызвать вымирание солончаков и превратить зеленые ландшафты в пустыни (Эстес и Даггинс, 1995, Силлиман и др., 2005). [10] [27] Основные триггеры трофических каскадов в результате деятельности человека включают внедрение инвазивных видов , чрезмерную эксплуатацию и изменение климата (Джексон и др., 2001, Лотце и др., 2006, Гедан и др., 2009, Гедан и Бертнесс, 2011). [29] [30]
Инвазивные виды
[ редактировать ]
Интродукция человеком чужеродных видов также может способствовать нисходящему контролю над болотными системами (Фурбиш и Альбано, 1994, Гоф и Грейс, 1998). [24] [31] Интродуцированные дикие лошади на барьерных островах Мэриленд, Вирджиния и Каролина полностью оголили болотные экосистемы (Фурбиш и Альбано, 1994). [24] а завезенные грызуны -нутрии на болота побережья Мексиканского залива в Луизиане могут снизить первичную продукцию болотных растений (Gough and Grace 1998). [31]
Инвазивный зеленый краб Carcinus maenas может отключить контроль сверху вниз, способствуя восстановлению экосистем солончаков. В болотах, которые вымерли в результате трофического каскада, вызванного чрезмерным выловом рыбы, пурпурные болотные крабы Sesarma reticulatum могут быть выселены более крупными зелеными крабами, которые охотятся на пурпурных крабов. Таким образом, зеленые крабы могут косвенно уменьшить травоядность пурпурных крабов и способствовать восстановлению кордграсса. Эти результаты обнадеживают, поскольку предполагают, что инвазивные виды, которые, как классически считается, оказывают в основном негативное воздействие на экосистемы, в которые они вторгаются, иногда могут действительно способствовать восстановлению деградировавших экосистем. [32]
Грибковый
[ редактировать ]Вымирание потенциально можно объяснить патогенами растений солончаков в некоторых районах: виды грибов были обнаружены на участках вымирания в Великобритании, а также на участках побережья Атлантического океана и Персидского залива США (Elmer 2013). [33] Деградация болот, вызванная грибками, является документально подтвержденной проблемой, особенно в Юго-Восточной Атлантике. Болотные улитки Littoraria irrorata во время выпаса делают небольшие надрезы в кордовой траве. Эти порезы способствуют росту грибка, а при высокой плотности улиток могут привести к массовым инфекциям , увеличению выпаса и последующей гибели (Silliman and Bertness 2002, Silliman and Newell 2003, Silliman et al. 2005). [22] [27] [34]
В прибрежной части Новой Англии эти доказательства слабее. Некоторые грибные возбудители S. alterniflora чаще обнаруживались в местах гибели. Эти патогены имеют различную степень вирулентности , и есть некоторые свидетельства связи с видами Fusarium и областями их гибели. Однако, хотя эта связь предполагает частичную причинно-следственную связь, особенно у растений, предрасположенных к стрессу, отсутствуют убедительные доказательства того, что такие грибковые патогены являются основной причиной вымирания растений в Новой Англии (Elmer 2013). [33]
Чрезмерная эксплуатация
[ редактировать ]Чрезмерный промысел привел к разрушению различных мелководных морских экосистем по всему миру (Джексон и др., 2001 г.). [29] включая коралловые рифы (Хьюз и др., 2003), [35] заросли морской травы и леса из водорослей (Эстес и др., 1998). [28]
Коммерческое рыболовство
[ редактировать ]
Исследования солончаковой улитки Littoraria irrorata и ее влияния на продуктивность болотных растений предоставили убедительные доказательства контроля над болотами со стороны потребителей, вызванного чрезмерной эксплуатацией. Эта улитка способна превращать стебли кордовой травы (Spartina alterniflora ) (высотой более 2,5 м) в ил за 8 месяцев, что составляет менее одного вегетационного периода (Silliman and Bertness 2002). [22] Как уже упоминалось ранее, болотные улитки наносят порезы на листья кордовой травы, когда они пасутся, обеспечивая субстрат и питательные вещества для грибка . При высокой плотности улиток кордграсс может поражаться грибковыми инфекциями , что может повлечь за собой вымирание болот (Силлиман и Бертнесс, 2002, Силлиман и Ньюэлл, 2003, Силлиман и др., 2005). [22] [27] [34] Естественными хищниками болотной улитки являются голубые крабы ( Callinectes sapidus ) и черепахи ( Malaclemys terrapin ), которые исторически регулировали численность улиток. Однако эти хищники подверглись чрезмерной коммерческой эксплуатации и теперь страдают от болезней из-за небольшого размера популяции, что освобождает улиток от потребительского давления и позволяет улиткам сеять хаос в популяциях солончаковой травы (Silliman and Zieman 2001). [22] [26]

Классический пример трофического каскада возник в результате чрезмерной эксплуатации каланов в 1980-х годах (Estes & Duggins 1995). [10] Морские выдры поедают ежей , которые, в свою очередь, питаются макроводорослями в пищевой цепочке, изменение которой может привести к бесплодию ежей . На каланов на Аляске охотились почти до полного исчезновения из-за их шкур . Там, где популяции каланов сохранились, они подавляют численность ежей и тем самым оказывают косвенное положительное влияние на плотность макроводорослей. Напротив, в местах, где каланы отсутствуют, популяция морских ежей резко возросла, что привело к образованию пустошей для ежей. Везде, где были восстановлены каланы, популяция морских ежей сокращалась, а плотность водорослей увеличивалась, возвращая среду обитания в исходное здоровое состояние. Этот пример демонстрирует потенциал восстановления всей экосистемы с восстановлением потребителей (Эстес и Даггинс, 1995). [10]
Другие трофические каскады, например, вызванные крабами, такими как Chasmagnathus granulata в Южной Америке, по крайней мере частично вызваны чрезмерным выловом высших хищников (Bortolus и Iribarne 1999, Alberti et al. 2007). [21] [25]
Любительская рыбалка
[ редактировать ]
В Новой Англии истощение численности хищников привело к обширному вымиранию болот в берегах солончаковых ручьев. Травоядность уже затронула до 90% берегов ручьев в более чем 70% внешних болот Кейп-Код. Отсутствовавшая в 1997 году гибель птиц недавно распространилась на залив Наррагансетт , штат Род-Айленд, затронув более 85% берегов ручьев. Ночной рытья пурпурный болотный краб Sesarma reticulatum играет важную роль в этом вымирании из-за увеличения нор и травоядности из-за освобождения от давления хищников. Имеющиеся данные указывают на то, что контроль сверху вниз, вызванный вмешательством человека, является основным фактором, способствующим вымиранию.
Альтьери и его коллеги провели серию экспериментов (Альтьери и др., 2012). [36] призван более конкретно объяснить механизмы, вызывающие вымирание болот. На крабов Sesarma обычно охотятся синие крабы ( Callinectes sapidus ) и рыба , в том числе полосатый окунь ( Morone saxatilis ) и гладкая акула ( Mustelus canis ). Хотя каскадные последствия чрезмерного вылова рыбы были продемонстрированы в различных экосистемах (Myers and Worm 2003), исследования по истощению численности хищников были сосредоточены почти исключительно на воздействии крупномасштабного коммерческого перелова (Worm et al. 2009). Однако Альтиери и его коллеги (2012) [36] продемонстрировали, что хищники Сесармы подвергались и продолжают подвергаться чрезмерной эксплуатации рыболовами-любителями . Их результаты показывают, что гибель и болота с растительностью резко различались по нагрузке рекреационного рыболовства, поскольку рыболовы наблюдались только в местах гибели.
Вымершие болота имели половину биомассы хищников высшего уровня, обнаруженных на участках с растительностью, тогда как биомасса непромысловых консументов не различалась на участках с вымиранием и зарослями. Кроме того, уровень хищничества Sesarma на участках с растительностью был в три раза выше, чем на участках с вымиранием, а крабы потребляли в четыре раза больше травы на участках с вымиранием, чем на участках с растительностью. Исторические реконструкции показали, что небольшая чистая потеря болот (<5%) с 1939 по 2005 год произошла на участках, покрытых в настоящее время растительностью. Напротив, вымирание болот началось в середине 70-х годов, с постоянным увеличением потери растительности до 2005 года, когда >20% общей площади болот было потеряно из-за вымирания и >80% зона кордграсса была лишена растительности. Это расхождение между вымиранием и болотами с растительностью в потере растительности совпадает с периодом быстрого увеличения количества причалов и стоянок лодок до середины 70-х годов, что привело к созданию> 70% той рыболовной инфраструктуры, которая в настоящее время присутствует на вымирающих болотах. вне болот. Эти результаты свидетельствуют о том, что Высвобождение сесармы под давлением хищников со стороны крабов и рыб из-за чрезмерного вылова рыбы рыболовами приводит к возникновению трофического каскада, который ответственен за массовое вымирание болот на юге Новой Англии (Altieri et al. 2012). [36]
Альтиери и коллеги (2012) [36] далее выдвинул гипотезу, что историческая крупномасштабная промышленная чрезмерная эксплуатация рыбы в северо-западной Атлантике (Лотце и др., 2006) [30] болот повышенная уязвимость к последствиям локализованного любительского рыболовства до такой степени, что последовало крупномасштабное вымирание, и что, как следствие, локальное вымирание может перерасти в полное вымирание болот в масштабах всего региона, если чрезмерная эксплуатация основных потребителей будет продолжаться (Altieri et al. . 2012). [36]
Совсем недавно Бертнесс и его коллеги (Бертнесс и др., 2014b) [37] провели полевой эксперимент по исключению хищников с участков на пастбищной границе болота. В течение одного вегетационного периода исключение высших хищников вызвало вымирание из-за увеличения численности крабов Sesarma и интенсивности травоядности. Для дальнейшей проверки гипотезы трофического каскада Бертнесс и его коллеги использовали распространение вымирания в заливе Наррагансетт, чтобы оценить все предполагаемые причины вымирания, в том числе: 1) эвтрофикация снижает вклад растений в подземную биомассу, вызывая гибель растений, 2) следы от лодок разрушают берега ручьев, 3) загрязнение или болезни влияют на здоровье растений, 4) твердость субстрата контролирует распространение травоядных крабов и 5) трофическая дисфункция освобождает Sesarma от контроля хищников. Доступность азота, интенсивность волн и рост растений не объясняют каких-либо изменений в гибели растений. Однако травоядность объясняет более 70% различий в вымирании между участками. (Бертнесс и др., 2014a) [38] Эта работа выявила один конкретный пример, когда экспериментально было показано, что нисходящие взаимодействия являются основной движущей силой изменения состояния экологического сообщества.
Взаимодействие снизу вверх
[ редактировать ]Гиперзасоленные и бескислородные почвы
[ редактировать ]Одна из теорий контроля «снизу вверх» заключается в различном почвы химическом составе на участках с растительностью и на участках с отмиранием. Было высказано предположение, что связанные с засухой изменения в химическом составе почвы связаны с областями вымирания. Засухи часто связаны с повышенным засолением и кислотности стрессом – почвы окисляются при ограничении пресной воды или приливно-отливной влажности , что приводит к повышению кислотности. Гиперсоленость связана с уменьшением выживаемости S. alterniflora (Brown et al., 2005). [39] Однако растения солончаков, как правило, толерантны к широкому диапазону уровней солености, и повышенная засоленность не всегда наблюдается в местах вымирания (Альбер и др., 2008). [40]
Существует неопределенность, связанная со связью между различными почвенными условиями и местами вымирания по нескольким причинам. Во-первых, измерения не могут проводиться через достаточно частые интервалы, чтобы уловить кратковременные колебания состояния почвы, связанные с вымиранием. Во-вторых, объяснение причинно-следственной связи с изменением почвенных условий и вымиранием потенциально сомнительно. Обратная причинно-следственная связь является потенциальным объяснением; то есть различия в химическом составе почвы могут быть результатом, а не фактором вымирания. Например, одно исследование отметило более высокие уровни солености в районах вымирания в некоторые месяцы, но это, возможно, было связано с изменением динамики корней или испарения из-за вымирания (Marsh 2007).
Эвтрофикация
[ редактировать ]Обогащение питательными веществами представляет собой широко распространенную глобальную угрозу как морским, так и наземным экосистемам (Галлоуэй и др., 2008 г., Верховен и др., 2006 г.). [41] [42] В морских экосистемах увеличение потока азота может спровоцировать сильное цветение водорослей, бескислородные условия и широкомасштабные потери рыболовства (Диас и Розенберг, 2008). [43] Предполагается, что в солончаках, важной экосистеме, разделяющей землю и море, добавление питательных веществ способствует повсеместному вымиранию ручьев (Диган и др., 2012). [44]
Чтобы проверить эту гипотезу, Линда Диган и ее коллеги провели девятилетнее исследование в одном из центров Массачусетса. Исследователи обнаружили, что обогащение приливных ручьев азотом и фосфором привело к уменьшению инвестиций в подземные корни и корневища, собирающие питательные вещества, усилению микробного разложения органического вещества и, в конечном итоге, к обрушению ручья и исчезновению солончаков (Диган и др., 2012). [44]
Однако аналогичные исследования в Коннектикуте не смогли повторить эти результаты. Шимон Анисфельд и Трой Хилл провели 5-летний эксперимент по удобрению солончака в проливе Лонг-Айленд и обнаружили, что ни азотные, ни фосфорные удобрения не привели к потере высоты, снижению содержания углерода в почве или снижению подземной первичной продукции. Они предполагают, что высокие уровни питательных веществ могут существенно изменить болотные процессы и увеличить общие потери углерода из отложений, но другие процессы могут компенсировать это воздействие. В результате этого исследования не ожидается никаких чистых вредных последствий нагрузки питательными веществами на хранение углерода или стабильность болот (Анисфельд и Хилл, 2012). [45]
Другие исследования эвтрофикации, проведенные в заливе Наррагансетт, штат Род-Айленд, показали, что экспериментальное добавление азота первоначально увеличивает продуктивность растений, но в конечном итоге приводит к снижению биомассы растений из-за травоядности насекомых (Bertness et al. 2008). [46] В дополнение к этому эксперименту исследователи провели обследование 20 солончаков, чтобы изучить взаимодействие между уровнем питательных веществ в болотах и давлением травоядных животных, и обнаружили, что поступление болотного азота является хорошим предиктором повреждения растений травоядными животными. Это исследование предполагает, что эвтрофикация в настоящее время вызывает подавление потребителями первичной продуктивности солончаков Новой Англии и может в конечном итоге представлять угрозу для предоставления экосистемных услуг солончаков (Бертнесс и др., 2008). [46]
Потенциал синергетического эффекта
[ редактировать ]Прибрежные экосистемы страдают от различных антропогенных воздействий , таких как крупномасштабная эвтрофикация , изменение пищевой сети , неконтролируемые потребительские эффекты, изменение климата , разрушение среды обитания и болезни . Эти факторы редко действуют изолированно. Часто исследователи обнаруживают аддитивное или синергетическое взаимодействие между воздействиями, усугубляющими степень деградации экосистемы . Один из таких примеров находится в солончаках на юго-востоке США. Истощение численности высших хищников в этих системах привело к чрезмерному выпасу улитками кордовой травы солончаков и последующему вымиранию (как объяснялось выше). Однако это вымирание было связано с сильной засухой и, как следствие, усилением солевого и кислотного стресса (Silliman et al. 2005). [27] Стресс засухи, который повышает засоление почвы и повышает уязвимость кордовой травы к нисходящему контролю, может быть продуктом изменения климата (Силлиман и Бертнесс, 2002, Силлиман и др., 2005). [22] [27] Подобные случаи показывают, как абиотические и биотические взаимодействия могут влиять на здоровье экосистемы .
Антропогенные действия также могут вызвать эвтрофикацию или увеличить нагрузку на морские экосистемы питательными веществами за счет стока в систему, содержащую удобрения , сточные воды , мыло для посудомоечной машины и другие вещества, богатые азотом и фосфором. Эвтрофикация широко распространена в прибрежных морских экосистемах (Лотце и др., 2006 г.). [30] и может косвенно инициировать трофические каскады и усилить потребительский контроль над растениями. Например, травоядность насекомых на болотах положительно коррелирует с наличием питательных веществ в солончаках Атлантического океана (Бертнесс и др., 2008). [46] В заливе Наррагансетт травоядные насекомые подавляют первичную продукцию нарушенных человеком солончаков почти на 40%. Добавление азота в результате деятельности человека может настолько увеличить численность травоядных насекомых, что первичное производство подавляется почти на 60%, в то время как болота, береговая линия которых не освоена человеком, остаются исключительно под контролем снизу вверх (Бертнесс и др., 2008). [46] Повышенное снабжение питательными веществами может привести к неконтролируемому развитию травоядных животных и в других системах (Gough and Grace 1998, Silliman and Zieman 2001). [26] [31]
Например, эвтрофикация инициирует контроль сверху вниз через воздействие снежных гусей на арктические болота Гудзонова залива (Джеффрис, 1997). [20] К 1980-м годам снежные гуси, которые первоначально ежегодно мигрировали в Гудзонов залив, переключились с кормления на умеренного пояса водно-болотных угодьях на кормление на сильно удобренных сельскохозяйственных полях. За 30 лет популяция гусей резко возросла. Снежные гуси с тех пор полностью уничтожили сотни тысяч акров водно-болотных угодий Гудзонова залива. Гуси грызли корни болотных растений, испарение увеличивалось, что приводило к последующему повышению засоленности, а без растений, насыщающих почву кислородом, субстрат становился бескислородным. Эта нежелательная среда для болотных растений препятствует сексуальной вербовке в этот район. Снежные гуси опустошили болота до тех пор, пока повторное заселение их клоновыми бегунами не сможет восстановить болотную растительность, что может занять годы. Из-за использования искусственных удобрений на сельскохозяйственных полях умеренной зоны был инициирован трофический каскад (Jefferies 1997). [20]
Основной целью экологии в следующем столетии будет понимание того, как экосистемы будут реагировать на нынешние и будущие воздействия человека, а также на аддитивные или синергетические взаимодействия между ними.
Последствия управления и сохранение
[ редактировать ]
Экосистемные услуги – это выгоды, которые люди извлекают из экологических систем. Сегодня одним из аргументов в пользу защиты солончаков является повышение качества и количества этих услуг. Солончаки улавливают азот, фильтруя сточные воды и уменьшая поступление азота в устья рек (Valiela and Cole 2002). [47] Солончаки также служат важным убежищем для молоди рыб и ракообразных , обеспечивая прибрежное рыболовство (Boesch and Turner 1984). [48] на долю которых приходится 90% мирового улова рыбы (ЮНЕП, 2006 г.). [49] Солончаки также улавливают углерод , который станет важной экосистемной услугой по мере усиления изменения климата (Chmura et al. 2003). [50] Вероятно, самая важная экосистемная услуга, которую предоставляют солончаки, заключается в том, что они действуют как естественные морские барьеры, поскольку травы связывают почву, предотвращают эрозию береговой линии , ослабляют волны и уменьшают прибрежные наводнения (Костанца и др., 2008). [51]
Однако естественная способность болота защищаться от эрозии и наводнений может быть снижена из-за отмирания берегов ручьев. Поскольку Spartina alterniflora ответственна за связывание отложений и отложение торфа (Redfield 1965), [52] Отмирание кордграсса может поставить под угрозу способность солончаков успевать за повышением уровня моря . Кроме того, концентрация нор Sesarma в торфе солончаков Новой Англии может напрямую спровоцировать эрозию и разрушение торфяного основания болот.
На Кейп-Коде берега болотных ручьев часто настолько пронизаны норами Сесармы , что они разрушаются, подвергая свежий торф дальнейшему закапыванию и эрозии (Бертнесс и др., 2008). [46] Таким образом, обратная связь между травоядностью крабов и отмиранием кордовой травы может привести к отступлению краев болот. Например, Ковердейл и его коллеги обнаружили, что 20 лет вымирания на Кейп-Коде привели к потере более чем 200-летнего прироста болот и потере сотен акров болот. [53] Пагубное воздействие трофических каскадов на болота не только снижает биоразнообразие , здоровье и эстетическую привлекательность этих экосистем, но и ставит под угрозу способность болот предоставлять ключевые экосистемные услуги человеческому населению.
Выводы
[ редактировать ]Несмотря на растущее количество свидетельств обратного, контроль снизу вверх остается центральной догмой, определяющей по управлению , сохранению и восстановлению усилия болот. В настоящее время управление солончаками осуществляется так, как будто они регулируются исключительно физическими факторами. Однако глобальное и прибрежное истощение высших хищников, ведущее к высвобождению загадочных или недооцененных травоядных животных, может быть самой большой текущей угрозой для солончаков. Зависимость от теории (подсознательное предпочтение выявлению и/или изучению природных явлений, которые имеют тенденцию скорее подтверждать, чем опровергать текущую парадигму системы исследования [Kuhn 1962] [54] ) и демонстрации, а не фальсификация науки, были главными виновниками этого упущения. Следовательно, угрозы солончакам, которые находятся под контролем сверху, игнорируются, что может иметь разрушительные последствия.
Трофические каскады — это мощные взаимодействия, которые жестко регулируют биоразнообразие , структуру сообщества и функции экосистемы. Первоначально считалось, что трофические каскады встречаются редко, но стало ясно, что они встречаются в разнообразных наземных, пресноводных и морских экосистемах как в малых, так и в больших пространственных и временных масштабах. Трофические каскады являются обычными и широко распространенными аспектами многих сообществ, которые, как долгое время считалось, находятся под контролем восходящих сил и/или устойчивы к контролю со стороны потребителей.
Таким образом, усилия по сохранению экосистем, затронутых трофическими каскадами, не включают должным образом нисходящий контроль в планы и усилия по управлению и восстановлению, но новые исследования подчеркивают, что это необходимо для защиты услуг, предоставляемых этими экосистемами, и восстановления этих экосистем до их первоначального состояния. условия. Несоблюдение этого требования может привести к возникновению трофических каскадов, превращающих весьма разнообразные и продуктивные растительные сообщества в бесплодные равнины.
Кроме того, неспособность переоценить действующую парадигму и признать, что как восходящие, так и нисходящие силы влияют на многие особенности структуры и функционирования экосистемы и что эти силы скорее дополняют друг друга, чем противоречат друг другу, может помешать любому включению обоих механизмов в любой план управления. и снизить успех усилий по сохранению еще до того, как они начнутся.
Ссылки
[ редактировать ]- ^ Линдеман, Р.Л. 1942. Трофический динамический аспект экологии. Экология 23: 399–418.
- ^ Перейти обратно: а б Хейрстон, Н.Г., Э.Д. Фредерик и Б.С. Лоуренс. 1960. Структура сообщества, контроль численности населения и конкуренция. Американский натуралист, 94 (879): 421–425.
- ^ Перейти обратно: а б Бертнесс, доктор медицинских наук, 2007. Берега Атлантического океана: естественная история и экология. Принстон, еврейский Джерси: Издательство Принстонского университета.
- ^ Перейти обратно: а б Strong DR 1992. Все ли трофические каскады связаны с влажной дифференцировкой и контролем доноров в конкретных экосистемах. Экология 73 (3): 747–754.
- ^ Эрлих, PR и LC Birch. 1967. «Природный баланс» и «контроль численности населения». Американский натуралист, 101: 97–107.
- ^ Диксон, AFF 1966. Влияние плотности популяции и пищевого статуса хозяина на летнюю репродуктивную активность платановой тли Drepanosiphum platanoides. Дж. Аним. Экол. 35 105–112.
- ^ Мердок, WW 1966. Структура сообщества, контроль рождаемости и конкуренция: критика. Американский натуралист, 100: 219–226.
- ^ Карпентер, SR, Дж. Ф. Китчелл и Дж. Р. Ходжсон. Каскадные трофические взаимодействия и продуктивность озер. BioScience 35, 634–639 (1985).
- ^ Power, ME 1992. Неоднородность среды обитания и функциональное значение рыб в речных пищевых сетях. Экология, 73: 1675–1688.
- ^ Перейти обратно: а б с д Эстес, Дж. А. и Д. О. Даггинс. 1995. Морские выдры и леса водорослей на Аляске: общность и вариации экологической парадигмы сообщества. Экологические монографии 65, 75–100.
- ^ Феллер, IC 2002. Роль травоядных насекомых-древоточцев в мангровых экосистемах Белиза. Ойкос 97, 167–176.
- ^ Пейн, RT 1966. Сложность сети и видовое разнообразие. Американский натуралист, 100 (910): 65–75.
- ^ Любченко Дж. и Б.А. Менге. 1978. Развитие и устойчивость сообщества в невысокой каменистой приливной зоне. Экологические монографии 48: 67–94.
- ^ Литтл, К. и Дж. А. Китчинг. 1996. Биология Скалистых берегов. Оксфорд, Великобритания: Издательство Оксфордского университета.
- ^ Перейти обратно: а б с Тил, Дж. М. 1962. Поток энергии в экосистеме солончаков Джорджии. Экология 43: 614–624.
- ^ Одум, EP 1971. Основы экологии. Филадельфия: Сондерс.
- ^ Перейти обратно: а б Смолли А.Е. 1960. Энергетический поток популяции солончаковых кузнечиков. Экология 41:672–77.
- ^ Перейти обратно: а б Бертнесс, доктор медицинских наук и Б.Р. Силлиман. 2008. Потребительский контроль над солончаками, вызванный вмешательством человека. Биология сохранения 22: 618–623.
- ^ Никсон, SW 1982. Экология высоких солончаков Новой Англии: профиль сообщества. Вашингтон, округ Колумбия: Деп. США. Межд. Рыба Вайлдл. Серв. FWS/OBS-81/55. 70 стр.
- ^ Перейти обратно: а б с д Джеффрис, Р.Л. 1997. Долгосрочный ущерб субарктическим экосистемам гусей: экологические индикаторы и меры дисфункции экосистемы. В Кроуфорде, RMM (ред.). Нарушение и восстановление арктических земель: экологическая перспектива. НАТО, Серия ASI Vol. 25: 151–165. Клювер: 1997.
- ^ Перейти обратно: а б с д Бортолус А., Ирибарн О. 1999. Воздействие роющего краба Chasmagnathus granulata юго-западной Атлантики на солончак Спартина. Море. Экол. Программа. Быть. 178: 79–88.
- ^ Перейти обратно: а б с д и ж г Силлиман, Б.Р. и доктор медицинских наук Бертнесс. 2002. Трофический каскад регулирует первичную продукцию солончаков. Труды Национальной академии наук США 99: 10500-10505.
- ^ Холдридж, К., А. Альтиери и доктор медицины Бертнесс. 2009. Роль травоядных крабов в вымирании солончаков Новой Англии. Биология сохранения 23: 672–679.
- ^ Перейти обратно: а б с Фербиш, CE и М. Альбано. 1994. Избирательная структура травоядных и растительных сообществ среднеатлантического солончака. Экология 75: 1015–1022.
- ^ Перейти обратно: а б с Альберти Дж., М. Эскапа, П. Далео, О. Ирибарн, Б. Р. Силлиман, доктор медицинских наук Бертнесс. 2007. Местные и географические различия в интенсивности выпаса растительноядных крабов в солончаках на юго-западе Атлантического океана. Мар Экол. Прог. Сер. 349: 235–43.
- ^ Перейти обратно: а б с д Силлиман Б.Р., Зиман Дж.К. 2001. Контроль сверху вниз за производством Spartina alterniflora путем выпаса барвинка на солончаке Вирджинии. Экология 82:2830–45.
- ^ Перейти обратно: а б с д и ж Силлиман Б.Р., ван де Коппель Дж., Бертнесс М.Д., Стэнтон Л.Е., Мендельсон И.А. 2005. Засуха , улитки и крупномасштабное вымирание солончаков на юге США. Наука 310:1803–6.
- ^ Перейти обратно: а б Эстес, Дж. А., М. Т. Тинкер, Т. М. Уильямс и Д. Ф. Доук. 1998. Хищничество косаток на каланов, связывающее океанические и прибрежные экосистемы. Наука 282: 473–476.
- ^ Перейти обратно: а б Джексон, JBC и др. 2001. Исторический чрезмерный вылов рыбы и недавний коллапс прибрежных экосистем. Наука 293: 629–638.
- ^ Перейти обратно: а б с Лотце, Х.К., Ленихан Х.С., Бурк Б.Дж., Брэдбери Р.Х. и другие. 2006. Истощение, деградация и потенциал восстановления эстуариев и прибрежных морей . Наука 312: 1806–1809.
- ^ Перейти обратно: а б с Гоф Л. и Дж. Б. Грейс. 1998. Влияние наводнений, засоленности и травоядности на прибрежные растительные сообщества, Луизиана, США. Экология 117:527–35.
- ^ Бертнесс, доктор медицинских наук и Т.К. Ковердейл. 2013. Инвазивный вид способствует восстановлению экосистем солончаков на Кейп-Коде. Экология 94:1937–1943.
- ^ Перейти обратно: а б Элмер, WH и др. 2013. Внезапное вымирание растительности в солончаках Атлантического океана и побережья Мексиканского залива. Болезни растений, 436–445.
- ^ Перейти обратно: а б Силлиман Б.Р., Ньюэлл С.Ю. 2003. Выращивание грибов у улитки. Учеб. Натл. акад. наук. 100:15643–48.
- ^ Хьюз, Т.П. и др. 2003. Изменение климата, воздействие человека и устойчивость коралловых рифов. Наука 301: 929–933.
- ^ Перейти обратно: а б с д и Альтьери, А.Х., доктор медицинских наук Бертнесс, Т.С. Ковердейл, Н.К. Херрманн, К. Анджелини. 2012. Трофический каскад вызывает коллапс солончаковой экосистемы с интенсивным любительским рыболовством. Экология 93 (6): 1402–1410.
- ^ Бертнесс, доктор медицинских наук, К.П. Бриссон * Т.К. Ковердейл, М.К. Бевил, С.М. Кротти и Э.Р. Суглиа. 2014 г. Экспериментальное уничтожение хищников приводит к быстрому вымиранию солончаков. Письма по экологии 17 (7): 830–835. два : 10.1111/ele.12287 .
- ^ Бертнесс, доктор медицинских наук, К. Бриссон, М. Бевил и С. Кротти. 2014. Травоядность способствует распространению вымирания солончаков. PLoS ONE 9(3): e92916.
- ^ Браун, CE, С.Р. Пезешки, Р.Д. ДеЛон. Влияние засоления и высыхания почвы на поглощение питательных веществ и рост S. alterniflora в моделируемой приливной системе. Экологическая и экспериментальная ботаника 58 (1–3): 140–148.
- ^ Альбер М., Э. Свенсон, С. Адамович, И. Мендельсон, 2008. Отмирание солончаков: обзор недавних событий в США. Эстуарий, Наука о прибрежных зонах и шельфе , 8(1):1–11.
- ^ Галлоуэй, Дж. и др. Трансформация азотного цикла: последние тенденции, вопросы и потенциальные решения. Наука 320, 889–892 (2008).
- ^ Верховен, JT и др. Региональные и глобальные опасения по поводу водно-болотных угодий и качества воды. Тенденции Экол. Эвол. 21, 96–103 (2006)
- ^ Диас, Р.Дж. и Р. Розенберг. Распространение мертвых зон и последствия для морских экосистем. Наука 321, 926–929 (2008).
- ^ Перейти обратно: а б Диган, Л.А., Боуэн, Дж.Л., Дрейк, Д., Флигер, Дж.В., Фридрихс, Коннектикут, Хобби, Дж.Э., Хопкинсон, К., Джонсон, Д.С., Джонсон, Дж.М., ЛеМэй, Л.Е., Миллер, Э., Петерсон, Б.Дж. , Пикард К., Шелдон С., Сазерленд М., Валлино Дж. и С. Уоррен. 2012. Прибрежная эвтрофикация как причина исчезновения солончаков. Природа 490:388–392.
- ^ Анисфельд, С. и Т. Хилл. 2012. Влияние удобрений на изменение высоты и подземный баланс углерода в приливных болотах пролива Лонг-Айленд. Эстуарии и побережья 35: 201–211.
- ^ Перейти обратно: а б с д и Бертнесс, доктор медицинских наук, К.М. Крейн, К. Холдридж и Н. Сала. 2008. Эвтрофикация и контроль потребителей первичной продукции солончаков Новой Англии. Биология сохранения 22: 131–139.
- ^ Валиела И. и М.Л. Коул. 2002. Сравнительные данные о том, что солончаки и мангровые заросли могут защитить луга, заросшие водорослями, от нагрузки азота, поступающей с суши. Экосистемы 5: 92–102.
- ^ Боеш Д.Ф. и Р.Э. Тернер. 1984. Зависимость промысловых видов от солончаков: роль пищи и убежища. Эстуарии 7: 460–468.
- ^ ЮНЕП. 2006. Морские и прибрежные экосистемы и благополучие человека: сводный отчет, основанный на результатах Оценки экосистем на пороге тысячелетия . ЮНЕП.
- ^ Чмура Г.Л., С.К. Анисфельд, Д.Р. Кахун и Дж.К. Линч. 2003. Глобальная секвестрация углерода в приливно-засоленных почвах водно-болотных угодий. Глобальные биогеохимические циклы 17: 1111–1133.
- ^ Костанца Р., О. Перес-Макео, М.Л. Мартинес, П. Саттон, С.Дж. Андерсон и К. Малдер. 2008. Значение прибрежных водно-болотных угодий для защиты от ураганов. Амбио 37: 241–248.
- ^ Редфилд, AC 1965 Онтогенез устья солончака. Наука 147: 50–55.
- ^ Ковердейл, Т.К., Э.В. Янг, С.Ф. Инь, К.П. Бриссон, Дж. П. Доннелли и доктор медицинских наук Бертнесс. 2014. Косвенное антропогенное воздействие обращает вспять многовековую секвестрацию углерода и образование солончаков. PLoS One, 9(3): e93296.
- ^ Кун, Т. 1962. Структура научных революций. Издательство Чикагского университета, Чикаго, Иллинойс.