Бактериофаг



Бактериофаг , ( / b æ k ˈ t ɪər i oʊ f eɪ dʒ / ), также неофициально известный как фаг ( / ˈ f eɪ dʒ / ), представляет собой вирус который заражает и размножается внутри бактерий и архей . Этот термин произошел от слова «бактерии» и греческого φαγεῖν ( фагин ), что означает «пожирать». Бактериофаги состоят из белков геном , инкапсулирующих или . РНК , ДНК и могут иметь как простые, так и сложные структуры Их геномы могут кодировать от четырех генов (например, MS2 ) до сотен генов . Фаги реплицируются внутри бактерии после инъекции их генома в ее цитоплазму .
Бактериофаги относятся к числу наиболее распространенных и разнообразных объектов биосферы . [2] Бактериофаги — это вездесущие вирусы, встречающиеся везде, где существуют бактерии. По оценкам, их более 10 31 бактериофагов на планете больше, чем у любого другого организма на Земле, включая бактерии, вместе взятые. [3] Вирусы являются наиболее распространенным биологическим объектом в водной толще мирового океана и вторым по величине компонентом биомассы после прокариот . [4] где до 9х10 8 вирионы на миллилитр обнаружены в микробных матах на поверхности, [5] и до 70% морских бактерий могут быть инфицированы бактериофагами. [6]
Бактериофаги использовались с 1920-х годов в качестве альтернативы антибиотикам в бывшем Советском Союзе и Центральной Европе, а также во Франции. [7] [8] Их рассматривают как возможную терапию против с множественной лекарственной устойчивостью штаммов многих бактерий (см. Фаготерапию ). [9] [10] [11] [12]
Известно, что бактериофаги взаимодействуют с иммунной системой как опосредованно через бактериальную экспрессию кодируемых фагом белков, так и напрямую, влияя на врожденный иммунитет и бактериальный клиренс. [13] Взаимодействие фага с хозяином становится все более важной областью исследований. [14]
Классификация [ править ]
Бактериофаги широко распространены в биосфере, имеют разные геномы и образ жизни. Фаги классифицируются Международным комитетом по таксономии вирусов (ICTV) по морфологии и нуклеиновой кислоте.

| Заказ | Семья | Морфология | Нуклеиновая кислота | Примеры |
|---|---|---|---|---|
| Белфривиралес | Турривирусиды | Конвертированный, изометрический | Линейная дцДНК | |
| Каудовирал | Аккерманвирусиды | Необолоченный . , сократимый хвост | Линейная дцДНК | |
| Автографивирусиды | Хвост без оболочки, несокращающийся (короткий) | Линейная дцДНК | ||
| Часевирусиды | Линейная дцДНК | |||
| Демереквирусиды | Линейная дцДНК | |||
| Дрекслервирусиды | Линейная дцДНК | |||
| Гвенливириды | Линейная дцДНК | |||
| Гереллевирусиды | Необолочечный, сократительный хвост | Линейная дцДНК | ||
| Миовирусиды | Необолочечный, сократительный хвост | Линейная дцДНК | Т4 , Му , П1 , П2 | |
| Сифовирусиды | Хвост без оболочки, несокращающийся (длинный) | Линейная дцДНК | л , Т5 , НК97 , Н15 | |
| Подовирусиды | Хвост без оболочки, несокращающийся (короткий) | Линейная дцДНК | Т7 , Т3 , Ф29 , П22 | |
| Раунтривирусиды | Линейная дцДНК | |||
| Саласмавирусиды | Линейная дцДНК | |||
| щитовирусиды | Линейная дцДНК | |||
| Зобелльвирусиды | Линейная дцДНК | |||
| Галопанивирусы | Сферолиповирусиды | Конвертированный, изометрический | Линейная дцДНК | |
| Симуловирусиды | Конвертированный, изометрический | Линейная дцДНК | ||
| Мацушитавирусиды | Конвертированный, изометрический | Линейная дцДНК | ||
| Галорувиралес | Плеолиповирусиды | Обернутый, плеоморфный | Круглая оцДНК, кольцевая дцДНК или линейная дцДНК | |
| Каламавиралес | Тективирусиды | Без оболочки, изометрический | Линейная дцДНК | |
| Лигаменвирусы | Липотриксвирусиды | Обернутая, палочковидная | Линейная дцДНК | Вирус ацидиануса нитевидный 1 |
| Рудивирусиды | Без оболочки, палочковидной формы | Линейная дцДНК | Палочковидный вирус Sulfolobus Islandicus 1 | |
| Минидивиралы | Цистовирусиды | Обернутая, сферическая | Линейная дсРНК | F6 |
| Норзивирусы | Аткинсвирусиды | Без оболочки, изометрический | Линейная оцРНК | |
| Дуинвирусиды | Без оболочки, изометрический | Линейная оцРНК | ||
| Фиерсвирусиды | Без оболочки, изометрический | Линейная оцРНК | MS2 , Qβ | |
| Солспивиридиды | Без оболочки, изометрический | Линейная оцРНК | ||
| Петитвирусы | Микровирусиды | Без оболочки, изометрический | Круговая оцДНК | ΦX174 |
| Примавирусы | Тристромавирусы | Обернутая, палочковидная | Линейная дцДНК | |
| Тимловиралес | Блюмевирусиды | Без оболочки, изометрический | Линейная оцРНК | |
| Steitzviridae | Без оболочки, изометрический | Линейная оцРНК | ||
| Тубулавирусы | Иновириды | Без оболочки, нитевидные | Круговая оцДНК | М13 |
| Паулиновириды | Без оболочки, нитевидные | Круговая оцДНК | ||
| Плектровирусиды | Без оболочки, нитевидные | Круговая оцДНК | ||
| Винавирусы | Кортиковирусы | Без оболочки, изометрический | Круговая дцДНК | ПМ2 |
| дурнавиралес | Пикобирнавирусиды (предложение) | Без оболочки, изометрический | Линейная дсРНК | |
| Неназначенный | Ампуллавирусиды | Конвертированный, в форме бутылки. | Линейная дцДНК | |
| Аутоликивирусиды | Без оболочки, изометрический | Линейная дцДНК | ||
| Бикаудавирусиды | Без конверта, в форме лимона. | Круговая дцДНК | ||
| Клававирусиды | Без оболочки, палочковидной формы | Круговая дцДНК | ||
| Финнлейкевирусиды | Без оболочки, изометрический | Круговая оцДНК | Подбросить [15] | |
| Фузелловирусиды | Без конверта, в форме лимона. | Круговая дцДНК | Альфафузелловирус | |
| Глобуловирусиды | Конвертированный, изометрический | Линейная дцДНК | ||
| Гуттавирусиды | Без оболочки, яйцевидная | Круговая дцДНК | ||
| Халспивиридиды | Без конверта, в форме лимона. | Линейная дцДНК | ||
| Плазмавирусиды | Обернутый, плеоморфный | Круговая дцДНК | ||
| Портоглобовирусиды | Конвертированный, изометрический | Круговая дцДНК | ||
| Таспивирусиды | Без конверта, в форме лимона. | Линейная дцДНК | ||
| Спиравирусиды | Без оболочки, палочковидной формы | Круговая оцДНК |
Было высказано предположение, что представители Picobirnaviridae заражают бактерии, но не млекопитающих. [16]
Существует также множество неназначенных родов класса Leviviricetes : Chimpavirus , Hohglivirus , Mahrahvirus , Meihzavirus , Nicedsevirus , Sculuvirus , Skrubnovirus , Tetipavirus и Winunavirus, содержащих линейные геномы оцРНК. [17] и неназначенный род Lilyvirus порядка Caudovirales, содержащий линейный геном дцДНК.
История [ править ]

В 1896 году Эрнест Ханбери Ханкин сообщил, что что-то в водах рек Ганг и Ямуна в Индии обладает выраженным антибактериальным действием против холеры и может проходить через очень тонкий фарфоровый фильтр. [18] В 1915 году британский бактериолог Фредерик Творт , руководитель лондонского Института Брауна, обнаружил небольшой агент, который заражал и убивал бактерии. Он считал, что агент должен быть одним из следующих:
- этап жизненного цикла бактерий
- фермент , вырабатываемый самими бактериями, или
- вирус, который вырос на бактериях и уничтожил их [19]
Исследования Творта были прерваны началом Первой мировой войны , а также нехваткой финансирования и открытием антибиотиков.
Независимо, франко-канадский микробиолог Феликс д'Эрель , работавший в Институте Пастера в Париже , объявил 3 сентября 1917 года, что он обнаружил «невидимый микроб-антагонист дизентерийной палочки ». Для д'Эреля не было никаких сомнений относительно природы его открытия: «Я в мгновение ока понял: то, что вызвало мои ясные пятна, на самом деле было невидимым микробом... вирусом, паразитирующим на бактериях». [20] Д'Эрелль назвал вирус бактериофагом, пожирателем бактерий (от греческого phagein , что означает «пожирать»). Он также записал драматический рассказ о человеке, страдавшем дизентерией, которому бактериофаги вернули хорошее здоровье. [21] Именно д'Эрель провел многочисленные исследования бактериофагов и ввел концепцию фаговой терапии . [22] В 1919 году в Париже, Франция, д'Эрелль провел первое клиническое применение бактериофага, а первое сообщение о его использовании в Соединенных Штатах было сделано в 1922 году. [23]
за исследования премии присуждены фагов Нобелевские
В 1969 году Макс Дельбрюк , Альфред Херши и Сальвадор Лурия были удостоены Нобелевской премии по физиологии и медицине за открытия репликации вирусов и их генетической структуры. [24] В частности, работа Херши, участвовавшего в эксперименте Херши-Чейза в 1952 году, предоставила убедительные доказательства того, что ДНК, а не белок, является генетическим материалом жизни. Дельбрюк и Лурия провели эксперимент Лурии-Дельбрюка , который статистически продемонстрировал, что мутации у бактерий происходят случайным образом и, таким образом, следуют принципам Дарвина , а не Ламарка .
Использует [ править ]
Фаготерапия [ править ]

Было обнаружено, что фаги являются антибактериальными агентами и использовались в бывшей советской республике Грузия (первопроходцем там был Георгий Элиава с помощью одного из первооткрывателей бактериофагов Феликса д'Эреля ) в 1920-х и 1930-х годах для лечения бактериальных инфекций.
Д'Эрелль «быстро понял, что бактериофаги обнаруживаются везде, где процветают бактерии: в канализации, реках, собирающих сточные воды из труб, и в кале выздоравливающих пациентов». [25]
Они имели широкое применение, в том числе для лечения солдат Красной Армии . [26] Однако на Западе от них отказались от общего использования по нескольким причинам:
- Антибиотики были открыты и широко продавались. Их было легче изготавливать, хранить и прописывать.
- Медицинские испытания фагов были проведены, но элементарное непонимание фагов вызвало вопросы о достоверности этих испытаний. [27]
- Публикации исследований в Советском Союзе велись в основном на русском или грузинском языках и в течение многих лет не сопровождались на международном уровне.
- Советские технологии широко не поощрялись, а в некоторых случаях были незаконными из-за печально известной красной паники .
Использование фагов продолжается после окончания Холодной войны в России. [28] Грузия и другие страны Центральной и Восточной Европы. О первом регулируемом рандомизированном двойном слепом клиническом исследовании сообщалось в журнале Journal of Wound Care в июне 2009 года, в котором оценивалась безопасность и эффективность коктейля бактериофагов для лечения инфицированных венозных язв ног у пациентов-людей. [29] FDA одобрило исследование как клиническое исследование фазы I. Результаты исследования продемонстрировали безопасность терапевтического применения бактериофагов, но не показали эффективности. Авторы объяснили, что использование определенных химических веществ, входящих в стандартный уход за ранами (например, лактоферрина или серебра), могло повлиять на жизнеспособность бактериофагов. [29] было опубликовано сообщение о другом контролируемом клиническом исследовании в Западной Европе (лечение ушных инфекций, вызванных Pseudomonas aeruginosa ). Вскоре после этого в журнале Clinical Otolaryngology в августе 2009 года [30] Исследование пришло к выводу, что препараты бактериофагов безопасны и эффективны для лечения хронических ушных инфекций у людей. Кроме того, были проведены многочисленные исследования на животных и другие экспериментальные клинические испытания по оценке эффективности бактериофагов при различных заболеваниях, таких как инфицированные ожоги и раны, инфекции легких, связанные с муковисцидозом, и другие. [30] С другой стороны, фаги Inoviridae было показано, что усложняют биопленки, участвующие в пневмонии и муковисцидозе , и защищают бактерии от лекарств, предназначенных для искоренения болезней, тем самым способствуя стойкой инфекции. [31]
Тем временем исследователи бактериофагов разрабатывают модифицированные вирусы для преодоления устойчивости к антибиотикам и разрабатывают фаговые гены, ответственные за кодирование ферментов, которые разрушают матрикс биопленки, структурные белки фага и ферменты, ответственные за лизис бактериальной клеточной стенки. [5] [6] [7] Были результаты, показывающие, что фаги Т4 небольшого размера и с коротким хвостом могут быть полезны при обнаружении кишечной палочки в организме человека. [32]
Терапевтическая эффективность фагового коктейля оценивалась на мышиной модели с назальной инфекцией A. baumannii с множественной лекарственной устойчивостью (MDR) . Мыши, получавшие фаговый коктейль, показали в 2,3 раза более высокую выживаемость по сравнению с мышами, не получавшими лечения, через семь дней после заражения. [33]
В 2017 году 68-летний пациент с диабетом и некротизирующим панкреатитом, осложненным псевдокистой, инфицированной штаммами MDR A. baumannii, в течение 4 месяцев лечился коктейлем из азитромицина, рифампицина и колистина, но безрезультатно и в целом его состояние быстро ухудшалось.
Поскольку началось обсуждение клинической бесполезности дальнейшего лечения, новый препарат для экстренного исследования (eIND) был подан в качестве последней попытки, по крайней мере, получить ценные медицинские данные из ситуации, и был одобрен, поэтому он был подвергнут фаговой терапии с использованием чрескожно (ПК) инъецируемый коктейль, содержащий девять различных фагов, которые были идентифицированы как эффективные против штамма первичной инфекции с помощью методов быстрой изоляции и тестирования (процесс, который занял менее одного дня). Это оказалось эффективным в течение очень короткого периода времени, хотя пациент не реагировал, и его здоровье продолжало ухудшаться; вскоре изоляты штамма A. baumannii были собраны из дренажа кисты, который показал устойчивость к этому коктейлю, и был добавлен второй коктейль, эффективность которого была проверена против этого нового штамма, на этот раз путем внутривенной (IV) инъекции, как стало ясно, что инфекция оказалась более распространенной, чем первоначально предполагалось. [34]
После сочетания внутривенной и ПК-терапии нисходящая клиническая траектория пациента изменилась, и в течение двух дней он вышел из комы и стал реагировать. Когда его иммунная система начала функционировать, его пришлось временно исключить из коктейля, поскольку его температура поднялась до более чем 104 °F (40 °C), но через два дня фаговые коктейли были повторно введены на уровне, который он мог переносить. . Первоначальный коктейль из трех антибиотиков был заменен миноциклином после того, как выяснилось, что бактериальный штамм не устойчив к нему, и он быстро полностью восстановил сознание, хотя его выписали из больницы примерно через 145 дней после начала фаготерапии. К концу терапии было обнаружено, что бактерии стали устойчивыми к обоим исходным фаговым коктейлям, но их продолжали, поскольку они, по-видимому, предотвращали развитие резистентности к миноциклину в собранных бактериальных образцах и оказывали полезный синергетический эффект. [34]
Другое [ править ]
Пищевая промышленность [ править ]
Фаги все чаще используются для обеспечения безопасности пищевых продуктов и предотвращения появления бактерий, вызывающих порчу . [35] С 2006 года Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) и Министерство сельского хозяйства США (USDA) одобрили несколько продуктов бактериофагов. LMP-102 (Intralytix) был одобрен для обработки готовых к употреблению (RTE) продуктов из птицы и мяса. В том же году FDA одобрило LISTEX (разработанный и производимый Micreos ) с использованием бактериофагов на сыре для уничтожения бактерий Listeria monocytogenes , чтобы придать им общепризнанный статус безопасных (GRAS). [36] В июле 2007 года один и тот же бактериофаг был одобрен для использования во всех пищевых продуктах. [37] В 2011 году Министерство сельского хозяйства США подтвердило, что LISTEX является средством обработки чистой этикетки и включено в Министерство сельского хозяйства США. [38] Продолжаются исследования в области безопасности пищевых продуктов, чтобы выяснить, являются ли литические фаги жизнеспособным вариантом борьбы с другими патогенами пищевого происхождения в различных пищевых продуктах. [39]
Индикаторы воды [ править ]
Бактериофаги, в том числе специфичные для Escherichia coli , использовались в качестве индикаторов фекального загрязнения источников воды. Благодаря общим структурным и биологическим характеристикам колифаги могут служить индикаторами вирусного фекального загрязнения и присутствия патогенных вирусов, таких как ротавирус, норовирус и ВГА. Исследования, проведенные в системах очистки сточных вод, выявили значительные различия в поведении колифагов по сравнению с фекальными колиформами, демонстрируя четкую корреляцию с восстановлением патогенных вирусов по завершении очистки. Исследования установили безопасный порог сброса, согласно которому сбросы ниже 3000 БОЕ/100 мл считаются безопасными с точки зрения ограничения выброса патогенных вирусов. [40]
Диагностика [ править ]
В 2011 году FDA разрешило первый продукт на основе бактериофага для использования в диагностике in vitro. [41] В тесте культуры крови KeyPath MRSA/MSSA используется смесь бактериофагов для обнаружения Staphylococcus aureus в положительных культурах крови и определения резистентности или чувствительности к метициллину . Тест дает результаты примерно через пять часов по сравнению с двумя-тремя днями для стандартных методов идентификации микробов и определения чувствительности. Это был первый ускоренный тест на чувствительность к антибиотикам, одобренный FDA. [42]
Противодействие биологическому оружию и токсинам [ править ]
Правительственные учреждения на Западе в течение нескольких лет обращались к Грузии и бывшему Советскому Союзу за помощью в использовании фагов для противодействия биологическому оружию и токсинам, таким как сибирская язва и ботулизм . [43] В исследовательских группах в США продолжаются разработки. Другие области применения включают распыление в садоводстве для защиты растений и овощной продукции от гниения и распространения бактериальных заболеваний. Другими применениями бактериофагов являются биоциды для поверхностей окружающей среды, например, в больницах, а также профилактическая обработка катетеров и медицинских устройств перед использованием в клинических условиях. В настоящее время существует технология нанесения фагов на сухие поверхности, например, на униформу, занавески или даже хирургические швы. Клинические исследования опубликованы в журнале Clinical Otolaryngology. [30] продемонстрировали успехи в ветеринарном лечении домашних собак с отитами .
и бактерий идентификация Обнаружение
Метод обнаружения и идентификации бактерий , запускаемый фагом ионных каскадов (SEPTIC), использует эмиссию ионов и ее динамику во время фаговой инфекции и обеспечивает высокую специфичность и скорость обнаружения. [44]
Фаговый дисплей [ править ]
Фаговый дисплей — это другое использование фагов, включающее библиотеку фагов с переменным пептидом, связанным с поверхностным белком. Геном каждого фага кодирует вариант белка, отображаемый на его поверхности (отсюда и название), обеспечивая связь между вариантом пептида и кодирующим его геном. Варианты фагов из библиотеки могут быть выбраны по их аффинности связывания с иммобилизованной молекулой (например, токсином ботулизма) для ее нейтрализации. Связанные выбранные фаги можно размножать путем повторного заражения восприимчивого бактериального штамма, что позволяет им извлечь закодированные в них пептиды для дальнейшего изучения. [45]
Открытие противомикробного препарата [ править ]
Фаговые белки часто обладают антимикробной активностью и могут служить основой для создания пептидомиметиков , то есть препаратов, имитирующих пептиды. [46] В технологии фаговых лигандов фаговые белки используются для различных целей, таких как связывание бактерий и бактериальных компонентов (например, эндотоксина ) и лизис бактерий. [47]
Фундаментальные исследования [ править ]
Бактериофаги — важные модельные организмы для изучения принципов эволюции и экологии . [48]
Вред [ править ]
Молочная промышленность [ править ]
Бактериофаги, присутствующие в окружающей среде, могут привести к тому, что сыр не забродит. Чтобы избежать этого, можно использовать закваски смешанных штаммов и режимы ротации культур. [49] Генная инженерия культурных микробов, особенно Lactococcus Lactis и Streptococcus thermophilus , изучалась с целью генетического анализа и модификации с целью повышения устойчивости к фагам . Особое внимание уделялось плазмидным и рекомбинантным хромосомным модификациям. [50] [35]
Некоторые исследования были сосредоточены на потенциале бактериофагов как антимикробного средства против патогенов пищевого происхождения и образования биопленок в молочной промышленности. Поскольку распространение устойчивости к антибиотикам является основной проблемой молочной промышленности, фаги могут служить многообещающей альтернативой. [51]
Репликация [ править ]

Жизненный цикл бактериофагов имеет тенденцию быть либо литическим , либо лизогенным циклом . Кроме того, некоторые фаги проявляют псевдолизогенное поведение. [13]
С помощью литических фагов, таких как фаг Т4 , бактериальные клетки вскрываются (лизируются) и уничтожаются после немедленной репликации вириона. Как только клетка разрушается, потомство фага может найти новых хозяев для заражения. [13] Литические фаги более подходят для фаготерапии . Некоторые литические фаги подвергаются явлению, известному как ингибирование лизиса, при котором законченное потомство фага не сразу высвобождается из клетки, если внеклеточные концентрации фага высоки. Этот механизм не идентичен механизму фага умеренного пояса и обычно носит временный характер. покоя [52]
Напротив, лизогенный цикл не приводит к немедленному лизису клетки-хозяина. Фаги, способные подвергаться лизогении, известны как умеренные фаги . Их вирусный геном будет интегрироваться с ДНК хозяина и реплицироваться вместе с ней, относительно безвредно, или даже может утвердиться в виде плазмиды . Вирус остается в состоянии покоя до тех пор, пока состояние хозяина не ухудшится, возможно, из-за истощения питательных веществ, затем эндогенные фаги (известные как профаги ) становятся активными. В этот момент они начинают репродуктивный цикл, приводящий к лизису клетки-хозяина. Поскольку лизогенный цикл позволяет клетке-хозяину продолжать выживать и размножаться, вирус реплицируется во всех потомках клетки. Примером бактериофага, который, как известно, следует лизогенному циклу и литическому циклу, является фаг лямбда E. coli. [53]
Иногда профаги могут приносить пользу бактериям-хозяевам, пока они находятся в состоянии покоя, добавляя новые функции к бактериальному геному , в явлении, называемом лизогенной конверсией . Примерами могут служить превращение бактериофагами безвредных штаммов Corynebacterium diphtheriae или Vibrio cholerae в высоковирулентные, вызывающие дифтерию или холеру соответственно. [54] [55] Были предложены стратегии борьбы с некоторыми бактериальными инфекциями путем воздействия на профаги, кодирующие токсин. [56]
Привязанность и проникновение [ править ]
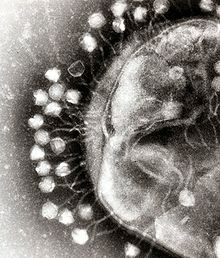
Бактериальные клетки защищены клеточной стенкой из полисахаридов , которые являются важными факторами вирулентности, защищающими бактериальные клетки как от иммунной защиты хозяина, так и от антибиотиков . [57] Чтобы проникнуть в клетку-хозяина, бактериофаги связываются со специфическими рецепторами на поверхности бактерий, включая липополисахариды , тейхоевые кислоты , белки или даже жгутики . Эта специфичность означает, что бактериофаг может инфицировать только определенные бактерии, несущие рецепторы, с которыми они могут связываться, что, в свою очередь, определяет круг хозяев фага. Ферменты, расщепляющие полисахариды, представляют собой белки, связанные с вирионами, которые ферментативно разрушают внешний слой капсулы своих хозяев на начальном этапе жестко запрограммированного процесса фаговой инфекции. [ нужна ссылка ] Условия роста хозяина также влияют на способность фага прикрепляться к ним и проникать в них. [58] Поскольку вирионы фага не движутся независимо, в растворе они должны полагаться на случайные встречи с правильными рецепторами, такими как кровь, лимфатическая циркуляция, ирригация, почвенная вода и т. д. [ нужна ссылка ]
Бактериофаги миовирусов используют движение, напоминающее подкожный шприц, для инъекции своего генетического материала в клетку. После контакта с соответствующим рецептором хвостовые волокна изгибаются, приближая базовую пластинку к поверхности клетки. Это известно как обратимое связывание. После полного прикрепления начинается необратимое связывание, и хвост сокращается, возможно, с помощью АТФ, присутствующего в хвосте. [6] введение генетического материала через бактериальную мембрану. [59] Инъекция осуществляется посредством своего рода изгибающего движения в стержне, когда он движется в сторону, сжимаясь ближе к клетке и толкаясь обратно вверх. У подовирусов нет удлиненной хвостовой оболочки, как у миовирусов, поэтому вместо этого они используют свои маленькие, похожие на зубы волокна хвоста ферментативно, чтобы разрушить часть клеточной мембраны перед внедрением своего генетического материала.
Синтез белков и нуклеиновых кислот [ править ]
Через несколько минут бактериальные рибосомы начинают транслировать вирусную мРНК в белок. Для фагов на основе РНК РНК-репликаза синтезируется на ранних стадиях процесса. Белки модифицируют бактериальную РНК-полимеразу , поэтому она преимущественно транскрибирует вирусную мРНК. Нормальный синтез белков и нуклеиновых кислот хозяина нарушается, и вместо этого он вынужден производить вирусные продукты. Эти продукты впоследствии становятся частью новых вирионов внутри клетки, белками-помощниками, которые способствуют сборке новых вирионов, или белками, участвующими в лизисе клеток . В 1972 году Уолтер Фирс ( Университет Гента , Бельгия ) первым установил полную нуклеотидную последовательность гена, а в 1976 году — вирусного генома бактериофага MS2 . [60] Некоторые бактериофаги дцДНК кодируют рибосомальные белки, которые, как полагают, модулируют трансляцию белков во время фаговой инфекции. [61]
Сборка вириона [ править ]
В случае фага Т4 построение новых вирусных частиц осуществляется с помощью хелперных белков, которые действуют каталитически во время морфогенеза фага . [62] Сначала собираются опорные пластины, а затем на них надстраиваются хвостовики. Головные капсиды, построенные отдельно, самопроизвольно соединяются с хвостами. Во время сборки фага Т4 вириона морфогенетические белки, кодируемые фаговыми генами, взаимодействуют друг с другом в характерной последовательности. Поддержание соответствующего баланса в количествах каждого из этих белков, вырабатываемых во время вирусной инфекции, по-видимому, имеет решающее значение для нормального морфогенеза фага Т4 . [63] ДНК эффективно упакована в головах. [64] Весь процесс занимает около 15 минут.
Высвобождение вирионов [ править ]
Фаги могут высвобождаться посредством лизиса клеток, экструзии или, в некоторых случаях, путем почкования. Лизис хвостатых фагов достигается с помощью фермента эндолизина , который атакует и разрушает пептидогликан клеточной стенки . Совершенно другой тип фага, нитчатый фаг , заставляет клетку-хозяина постоянно секретировать новые вирусные частицы. Высвободившиеся вирионы описываются как свободные и, если они не являются дефектными, способны инфицировать новую бактерию. Почкование связано с определенными фагами микоплазмы . В отличие от высвобождения вирионов, фаги, демонстрирующие лизогенный цикл, не убивают хозяина, а вместо этого становятся долговременными резидентами в качестве профагов . [65]
Общение [ править ]
Исследования 2017 года показали, что бактериофаг Φ3T вырабатывает короткий вирусный белок, который сигнализирует другим бактериофагам о том, что они находятся в состоянии покоя, а не убивают бактерию-хозяина. Арбитриум — это название, данное этому белку исследователями, открывшими его. [66] [67]
Структура генома [ править ]
Учитывая миллионы различных фагов в окружающей среде, фаговые геномы бывают самых разных форм и размеров. РНК-фаги, такие как MS2, имеют самые маленькие геномы, всего несколько тысяч оснований. Однако некоторые ДНК-фаги, такие как Т4, могут иметь большие геномы с сотнями генов; размер и форма капсида варьируются вместе с размером генома. [68] Самые крупные геномы бактериофагов достигают размера 735 т.п.н. [69]

Геномы бактериофагов могут быть очень мозаичными , то есть геном многих видов фагов состоит из множества отдельных модулей. Эти модули могут быть обнаружены у других видов фагов в различных формах. Микобактериофаги , бактериофаги с микобактериальными хозяевами, предоставили прекрасные примеры этого мозаицизма. У этих микобактериофагов генетический ассортимент может быть результатом повторяющихся случаев сайт-специфической рекомбинации и незаконной рекомбинации (результат приобретения фаговым геномом генетических последовательностей бактериального хозяина). [71] Эволюционные механизмы формирования геномов бактериальных вирусов различаются у разных семейств и зависят от типа нуклеиновой кислоты, особенностей структуры вириона, а также способа жизненного цикла вируса. [72]
Некоторые морские фаги розеобактерий содержат в своей геномной ДНК дезоксиуридин (dU) вместо дезокситимидина (dT). Есть некоторые свидетельства того, что этот необычный компонент является механизмом обхода бактериальных защитных механизмов, таких как эндонуклеазы рестрикции и системы CRISPR/Cas , которые эволюционировали, чтобы распознавать и расщеплять последовательности внутри вторгающихся фагов, тем самым инактивируя их. Давно известно, что другие фаги используют необычные нуклеотиды. В 1963 году Такахаши и Мармур идентифицировали фаг Bacillus , в геноме которого dU заменял dT. [73] а в 1977 г. Кирнос и др. идентифицировали цианофаг , содержащий 2-аминоаденин (Z) вместо аденина (А). [74]
Системная биология
Область системной биологии исследует сложные сети взаимодействий внутри организма, обычно с использованием вычислительных инструментов и моделирования. [75] Например, фаговый геном, проникающий в бактериальную клетку-хозяина, может экспрессировать сотни фаговых белков, которые будут влиять на экспрессию многочисленных генов хозяина или его метаболизм . Все эти сложные взаимодействия можно описать и смоделировать с помощью компьютерных моделей. [75]
Например, заражение Pseudomonas aeruginosa умеренным фагом РаР3 изменило экспрессию 38% (2160/5633) генов хозяина. Многие из этих эффектов, вероятно, являются косвенными, поэтому задача состоит в выявлении прямых взаимодействий между бактериями и фагами. [76]
Было предпринято несколько попыток картировать белок-белковые взаимодействия между фагом и его хозяином. Например, было обнаружено, что бактериофаг лямбда взаимодействует со своим хозяином, E. coli , посредством десятков взаимодействий. Опять же, значение многих из этих взаимодействий остается неясным, но эти исследования показывают, что, скорее всего, существует несколько ключевых взаимодействий и множество непрямых взаимодействий, роль которых остается невыясненной. [77]
Сопротивление хозяина [ править ]
Бактериофаги представляют собой серьезную угрозу для бактерий, а прокариоты выработали многочисленные механизмы, позволяющие блокировать инфекцию или блокировать репликацию бактериофагов в клетках-хозяевах. Система CRISPR — один из таких механизмов, как ретроны и кодируемая ими антитоксиновая система. [78] Известно, что защитная система Thoeris использует уникальную стратегию борьбы с бактериальной устойчивостью к антифагам посредством деградации НАД+ . [79]
- бактериофаг хозяин Симбиоз
Умеренные фаги — это бактериофаги, которые интегрируют свой генетический материал в хозяина в виде внехромосомных эписом или профагов во время лизогенного цикла . [80] [81] [82] Некоторые умеренные фаги могут придавать своему хозяину преимущества в приспособленности различными способами, включая придание устойчивости к антибиотикам за счет переноса или введения генов устойчивости к антибиотикам (ARG), [81] [83] защита хозяев от фагоцитоза, [84] [85] защита хозяев от вторичной инфекции путем исключения суперинфекции, [86] [87] [88] усиление патогенности хозяина, [80] [89] или усиление бактериального метаболизма или роста. [90] [91] [92] [93] Симбиоз бактериофаг-хозяин может принести пользу бактериям, предоставляя селективные преимущества при пассивной репликации генома фага. [94]
В окружающей среде [ править ]
Метагеномика позволила обнаружить бактериофаги в воде, что раньше было невозможно. [95]
Кроме того, бактериофаги использовались для гидрологического отслеживания и моделирования речных систем, особенно там, где происходит взаимодействие поверхностных и подземных вод . Использование фагов предпочтительнее более традиционных маркеров- красителей , поскольку они значительно меньше поглощаются при прохождении через грунтовые воды и легко обнаруживаются при очень низких концентрациях. [96] Незагрязненная вода может содержать примерно 2×10 8 бактериофагов на мл. [97]
Считается, что бактериофаги вносят значительный вклад в горизонтальный перенос генов в естественной среде, главным образом посредством трансдукции , но также и посредством трансформации . [98] Исследования, основанные на метагеномике, также показали, что виромы из различных сред содержат гены устойчивости к антибиотикам, в том числе те, которые могут придавать множественную лекарственную устойчивость . [99]
Недавние открытия позволили составить карту сложного и переплетенного арсенала средств антифаговой защиты у бактерий окружающей среды. [100]
У людей [ править ]
Хотя фаги не заражают человека, в организме человека имеется бесчисленное множество фаговых частиц, учитывая наш обширный микробиом . Нашу популяцию фагов назвали человеческим фагеомом , включая «фагом здорового кишечника» (HGP) и «фагом больного человека» (DHP). [101] По оценкам , активный фагом здорового человека (т.е. активно реплицирующийся, а не нереплицирующийся интегрированный профаг ) включает от десятков до тысяч различных вирусов. [102] Имеются доказательства того, что бактериофаги и бактерии взаимодействуют в микробиоме кишечника человека как антагонистически, так и полезно. [103]
Предварительные исследования показали, что обычные бактериофаги обнаруживаются в среднем у 62% здоровых лиц, тогда как их распространенность снижается в среднем на 42% и 54% у больных язвенным колитом (ЯК) и болезнью Крона (БК). [101] Обилие фагов также может снижаться у пожилых людей. [103]
Наиболее распространенными фагами в кишечнике человека, обнаруженными во всем мире, являются crAssphages . КрАсфаги передаются от матери к ребенку вскоре после рождения, и есть некоторые данные, свидетельствующие о том, что они могут передаваться локально. У каждого человека формируются свои уникальные кластеры crAssphage. CrAss-подобные фаги могут присутствовать не только у человека, но и у приматов . [103]
Обычно изучаемые бактериофаги
Среди бесчисленного множества фагов лишь немногие изучены подробно, в том числе некоторые исторически важные фаги, открытые на заре микробной генетики. Они, особенно Т-фаг, помогли открыть важные принципы структуры и функции генов.
Базы данных и ресурсы по бактериофагам [ править ]
См. также [ править ]
- Антибиотик
- Бактериоядное животное
- КрАссфаг
- КРИСПР
- ДНК-вирусы
- Макрофаг
- Экология фага
- Монографии о фагах (полный список монографий о фагах и связанных с ними фагах, с 1921 г. по настоящее время)
- Фагемида
- Полифаг
- РНК-вирусы
- Трансдукция
- Вириом
- Вирофаг – вирусы, заражающие другие вирусы.
Ссылки [ править ]
- ^ Падилья-Санчес V (2021). «Структурная модель бактериофага Т4» . Викижурнал науки . 4 (1): 5. doi : 10.15347/WJS/2021.005 . S2CID 238939621 .
- ^ Jump up to: Перейти обратно: а б МакГрат С., ван Синдерен Д., ред. (2007). Бактериофаг: генетика и молекулярная биология (1-е изд.). Кайстер Академик Пресс. ISBN 978-1-904455-14-1 .
- ^ ЛаФи С., Бушман Х. (25 апреля 2017 г.). «Новая фаготерапия спасает пациентов с бактериальной инфекцией с множественной лекарственной устойчивостью» . UC Health – Калифорнийский университет в Сан-Диего . Проверено 13 мая 2018 г.
- ^ Саттл, Калифорния (сентябрь 2005 г.). «Вирусы в море». Природа . 437 (7057): 356–361. Бибкод : 2005Natur.437..356S . дои : 10.1038/nature04160 . ПМИД 16163346 . S2CID 4370363 .
- ^ Jump up to: Перейти обратно: а б Уоммак К.Е., Колвелл Р.Р. (март 2000 г.). «Вириопланктон: вирусы в водных экосистемах» . Обзоры микробиологии и молекулярной биологии . 64 (1): 69–114. дои : 10.1128/ММБР.64.1.69-114.2000 . ПМК 98987 . ПМИД 10704475 .
- ^ Jump up to: Перейти обратно: а б с м Прескотт Л. (1993). Микробиология . Вм. Издательство К. Браун. ISBN 0-697-01372-3 .
- ^ Jump up to: Перейти обратно: а б Бантинг Дж. (1997). «Вирус, который лечит» . Би-би-си Горизонт . BBC Worldwide Ltd. OCLC 224991186 . – Документальный фильм об истории фаговой медицины в России и на Западе.
- ^ Боррелл Б., Фишетти В. (август 2012 г.). «Научный разговор: Фаговый фактор». Научный американец . стр. 80–83. JSTOR 26016042 .
- ^ Кортрайт К.Е., Чан Б.К., Кофф Дж.Л., Тернер П.Е. (февраль 2019 г.). «Фаготерапия: новый подход к борьбе с бактериями, устойчивыми к антибиотикам» . Клетка-хозяин и микроб . 25 (2): 219–232. дои : 10.1016/j.chom.2019.01.014 . ПМИД 30763536 . S2CID 73439131 .
- ^ Гордилло Альтамирано, Флорида, Барр Джей Джей (апрель 2019 г.). «Фаготерапия в эпоху постантибиотиков» . Обзоры клинической микробиологии . 32 (2). дои : 10.1128/CMR.00066-18 . ПМК 6431132 . ПМИД 30651225 .
- ^ Гонсалес-Мора А., Эрнандес-Перес Х., Икбал Х.М., Рито-Паломарес М., Бенавидес Х. (сентябрь 2020 г.). «Вакцины на основе бактериофагов: мощный подход к доставке антигенов» . Вакцина . 8 (3): 504. doi : 10.3390/vaccines8030504 . ПМЦ 7565293 . ПМИД 32899720 .
- ^ Кин ЕС (2012). «Фаготерапия: концепция лечения» . Границы микробиологии . 3 : 238. дои : 10.3389/fmicb.2012.00238 . ПМК 3400130 . ПМИД 22833738 .
- ^ Jump up to: Перейти обратно: а б с Попеску М., Ван Беллегем Дж.Д., Хосрави А., Боллыки П.Л. (сентябрь 2021 г.). «Бактериофаги и иммунная система» . Ежегодный обзор вирусологии . 8 (1): 415–435. doi : 10.1146/annurev-virology-091919-074551 . ПМИД 34014761 .
- ^ Стоун Э., Кэмпбелл К., Грант И., Маколифф О. (июнь 2019 г.). «Понимание и использование взаимодействий фаг-хозяин» . Вирусы . 11 (6): 567. дои : 10.3390/v11060567 . ПМК 6630733 . ПМИД 31216787 .
- ^ Лаанто Э., Мянтюнен С., Де Колибус Л., Марьякангас Дж., Гиллум А., Стюарт Д.И. и др. (август 2017 г.). «Вирус, обнаруженный в бореальном озере, связывает вирусы оцДНК и дцДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (31): 8378–8383. Бибкод : 2017PNAS..114.8378L . дои : 10.1073/pnas.1703834114 . ПМЦ 5547622 . ПМИД 28716906 .
- ^ Кришнамурти С.Р., Ван Д. (март 2018 г.). «Обширная консервация прокариотических сайтов связывания рибосом у известных и новых пикобирнавирусов» . Вирусология . 516 : 108–114. дои : 10.1016/j.virol.2018.01.006 . ПМИД 29346073 .
- ^ Калланан Дж., Стокдейл С.Р., Адриансенс Э.М., Кун Дж.Х. (январь 2021 г.). «Переименуйте один класс (Leviviricetes — ранее Allassoviricetes), переименуйте один отряд (Norzivirales — ранее Levivirales), создайте один новый порядок (Timlovirales) и расширите класс в общей сложности до шести семейств, 420 родов и 883 видов» . Исследовательские ворота . дои : 10.13140/RG.2.2.25363.40481 .
- ^ Ханкин Э.Х. (1896). «Бактерицидное действие вод Джамны и Ганга на холерный вибрион» . Анналы Института Пастера (на французском языке). 10 :511–23.
- ^ Творт Ф.В. (1915). «Исследование природы ультрамикроскопических вирусов» . Ланцет . 186 (4814): 1241–43. дои : 10.1016/S0140-6736(01)20383-3 .
- ^ д'Эрель Ф (1917). «О невидимом микробе, антагонисте дизентерийной палочки» (PDF) . Доклады Парижской академии наук . 165 :373–5. Архивировано (PDF) из оригинала 11 мая 2011 года . Проверено 5 сентября 2010 г.
- ^ д'Эрель Ф (1949). «Бактериофаг» (PDF) . Новости науки . 14 :44–59 . Проверено 5 сентября 2010 г.
- ^ Кин EC (декабрь 2012 г.). «Феликс д'Эрель и наше микробное будущее». Будущая микробиология . 7 (12): 1337–1339. дои : 10.2217/fmb.12.115 . ПМИД 23231482 .
- ^ Асуани В.Х., Шукла СК (июнь 2021 г.). «Ранняя история фаготерапии в Соединенных Штатах: пора ли пересмотреть свое мнение?» . Клиническая медицина и исследования . 19 (2): 82–89. дои : 10.3121/cmr.2021.1605 . ПМЦ 8231696 . ПМИД 34172535 .
- ^ «Нобелевская премия по физиологии и медицине 1969 года» . Нобелевский фонд . Проверено 28 июля 2007 г.
- ^ Кучмент А. (2012), Забытое лекарство: прошлое и будущее фаготерапии , Springer, с. 11, ISBN 978-1-4614-0250-3
- ^ Мельников Д. (октябрь 2018 г.). «Альтернативное лечение: внедрение и выживание бактериофаговой терапии в СССР, 1922-1955 гг.» . Журнал истории медицины и смежных наук . 73 (4): 385–411. дои : 10.1093/jhmas/jry024 . ПМК 6203130 . ПМИД 30312428 .
- ^ Каттер Э., Де Вос Д., Гвасалия Г., Алавидзе З., Гогохия Л., Куль С. и др. (январь 2010 г.). «Фаготерапия в клинической практике: лечение инфекций человека». Современная фармацевтическая биотехнология . 11 (1): 69–86. дои : 10.2174/138920110790725401 . ПМИД 20214609 . S2CID 31626252 .
- ^ Golovin S (2017). "Бактериофаги: убийцы в роли спасителей" [Bacteriophages: killers as saviors]. Наука и жизнь [ Nauka I Zhizn (Science and life) ] (in Russian) (6): 26–33.
- ^ Jump up to: Перейти обратно: а б Роудс Д.Д., Уолкотт Р.Д., Кусковски М.А., Уолкотт Б.М., Уорд Л.С., Сулаквелидзе А. (июнь 2009 г.). «Бактериофаговая терапия венозных язв ног у человека: результаты I фазы исследования безопасности». Журнал ухода за ранами . 18 (6): 237–8, 240–3. дои : 10.12968/jowc.2009.18.6.42801 . ПМИД 19661847 .
- ^ Jump up to: Перейти обратно: а б с Райт А., Хокинс Ч.Х., Анггард Э.Э., Харпер Д.Р. (август 2009 г.). «Контролируемое клиническое исследование терапевтического препарата бактериофага при хроническом отите, вызванном устойчивой к антибиотикам Pseudomonas aeruginosa; предварительный отчет об эффективности» . Клиническая отоларингология . 34 (4): 349–357. дои : 10.1111/j.1749-4486.2009.01973.x . ПМИД 19673983 .
- ^ Свир Дж.М., Ван Беллегем Дж.Д., Ишак Х., Бах М.С., Попеску М., Сункари В. и др. (март 2019 г.). «Бактериофаги вызывают противовирусный иммунитет и предотвращают устранение бактериальной инфекции» . Наука . 363 (6434): eaat9691. дои : 10.1126/science.aat9691 . ПМК 6656896 . ПМИД 30923196 .
- ^ Тавил Н., Захер Э., Мандевиль Р., Менье М. (май 2012 г.). «Поверхностно-плазмонное резонансное обнаружение E. coli и метициллин-резистентного S. aureus с использованием бактериофагов» (PDF) . Биосенсоры и биоэлектроника . 37 (1): 24–29. дои : 10.1016/j.bios.2012.04.048 . ПМИД 22609555 . Архивировано (PDF) из оригинала 2 февраля 2023 года.
- ^ Ча К., О Х.К., Чан Дж.Ю., Джо Ю., Ким В.К., Ха ГУ и др. (10 апреля 2018 г.). «Характеристика двух новых бактериофагов, инфицирующих Acinetobacter baumannii с множественной лекарственной устойчивостью (MDR) , и оценка их терапевтической эффективности in vivo » . Границы микробиологии . 9 : 696. дои : 10.3389/fmicb.2018.00696 . ПМЦ 5932359 . ПМИД 29755420 .
- ^ Jump up to: Перейти обратно: а б Скули Р.Т., Бисвас Б., Гилл Дж.Дж., Эрнандес-Моралес А., Ланкастер Дж., Лессор Л. и др. (октябрь 2017 г.). «Разработка и применение персонализированных лечебных коктейлей на основе бактериофагов для лечения пациента с диссеминированной резистентной инфекцией Acinetobacter baumannii» . Антимикробные средства и химиотерапия . 61 (10). дои : 10.1128/AAC.00954-17 . ПМК 5610518 . ПМИД 28807909 .
- ^ Jump up to: Перейти обратно: а б О'Салливан Л., Болтон Д., Маколифф О., Коффи А. (март 2019 г.). «Бактериофаги в пищевой промышленности: от врага к другу». Ежегодный обзор пищевой науки и технологий . 10 (1). Годовые обзоры : 151–172. doi : 10.1146/annurev-food-032818-121747 . ПМИД 30633564 . S2CID 58620015 .
- ^ US FDA/CFSAN: Ответное письмо агентства, уведомление GRAS № 000198.
- ^ (US FDA/CFSAN: ответное письмо агентства, уведомление GRAS № 000218)
- ^ «Директива FSIS 7120: Безопасные и подходящие ингредиенты, используемые при производстве продуктов из мяса, птицы и яиц» (PDF) . Служба безопасности и инспекции пищевых продуктов . Вашингтон, округ Колумбия: Министерство сельского хозяйства США. Архивировано из оригинала (PDF) 18 октября 2011 года.
- ^ Хан FM, Чен Дж. Х., Чжан Р., Лю Б (2023). «Всесторонний обзор применения эндолизинов, полученных из бактериофагов, для борьбы с пищевыми бактериальными патогенами и безопасности пищевых продуктов: последние достижения, проблемы и перспективы на будущее» . Границы микробиологии . 14 : 1259210. дои : 10.3389/fmicb.2023.1259210 . ПМЦ 10588457 . ПМИД 37869651 .
- ^ Чакон Л., Баррантес К., Сантамария-Уллоа С., Солано М.Рейес Л., Тейлор Л.Валиенте С., Симондс Э.М., Ачи Р. 2020. Пороговый подход соматического колифага для улучшения управления сточными водами очистных сооружений с активным илом в регионах с ограниченными ресурсами. Appl Environ Microbiol 86:e00616-20. https://doi.org/10.1128/AEM.00616-20/
- ^ «Предварительное уведомление FDA 510(k)» . Управление по контролю за продуктами и лекарствами США.
- ^ Чакон Л., Баррантес К., Сантамария-Уллоа С., Солано М., Рейес Л., Тейлор Л. и др. (6 мая 2011 г.). «FDA одобрило первый тест для быстрой диагностики и различения MRSA и MSSA» . Прикладная и экологическая микробиология . 86 (17). Управление по контролю за продуктами и лекарствами США. дои : 10.1128/aem.00616-20 . ПМЦ 7440787 . ПМИД 32591380 . Архивировано из оригинала 5 апреля 2024 года.
- ^ Вайсман Д. (25 мая 2007 г.). «Изучение сибирской язвы в лаборатории советских времен – при западном финансировании» . Нью-Йорк Таймс .
- ^ Добози-Кинг М., Со С., Ким Джу, Ён Р., Ченг М., Киш Л.Б. (2005). «Быстрое обнаружение и идентификация бактерий: сенсорирование ионного каскада, запускаемого фагами (SEPTIC)» (PDF) . Журнал биологической физики и химии . 5 : 3–7. дои : 10.4024/1050501.jbpc.05.01 . Архивировано из оригинала (PDF) 26 сентября 2018 года . Проверено 19 декабря 2016 г.
- ^ Смит Г.П., Петренко В.А. (апрель 1997 г.). «Фаговый дисплей». Химические обзоры . 97 (2): 391–410. дои : 10.1021/cr960065d . ПМИД 11848876 .
- ^ Лю Дж., Деби М., Моек Г., Архин Ф., Бауда П., Бержерон Д. и др. (февраль 2004 г.). «Открытие противомикробных препаратов посредством геномики бактериофагов». Природная биотехнология . 22 (2): 185–191. дои : 10.1038/nbt932 . ПМИД 14716317 . S2CID 9905115 .
- ^ «Технологические основы технологии Фаго-лигандов» . биоМерье .
- ^ Кин EC (январь 2014 г.). «Компромиссы в истории жизни бактериофагов» . Бактериофаг . 4 (1): e28365. дои : 10.4161/bact.28365 . ПМЦ 3942329 . ПМИД 24616839 .
- ^ Атамер З., Самтлебе М., Неве Х., Дж. Хеллер К., Хинрикс Дж. (16 июля 2013 г.). «Обзор: устранение бактериофагов в сыворотке и сывороточных продуктах» . Границы микробиологии . 4 : 191. дои : 10.3389/fmicb.2013.00191 . ПМЦ 3712493 . ПМИД 23882262 .
- ^ Коффи А., Росс Р.П. (август 2002 г.). «Системы устойчивости к бактериофагам в штаммах молочной закваски: молекулярный анализ к применению». Антони ван Левенгук . 82 (1–4). Спрингер : 303–321. дои : 10.1023/а:1020639717181 . ПМИД 12369198 . S2CID 7217985 .
- ^ Фернандес Л., Эскобедо С., Гутьеррес Д., Портилья С., Мартинес Б., Гарсиа П. и др. (ноябрь 2017 г.). «Бактериофаги в молочной среде: от врагов к союзникам» . Антибиотики . 6 (4). [MDPI]: 27. doi : 10.3390/antibiotics6040027 . ПМЦ 5745470 . ПМИД 29117107 .
- ^ Абедон СТ (октябрь 2019 г.). «Посмотрите, кто говорит: ингибирование лизиса T-даже фага, дедушка исследований межклеточной коммуникации вирус-вирус» . Вирусы . 11 (10): 951. дои : 10.3390/v11100951 . ПМЦ 6832632 . ПМИД 31623057 .
- ^ Мейсон К.А., Лосос Дж.Б., Сингер С.Р., Рэйвен П.Х., Джонсон ГБ (2011). Биология . Нью-Йорк: МакГроу-Хилл. п. 533. ИСБН 978-0-07-893649-4 .
- ^ Мокроусов И (январь 2009 г.). «Corynebacterium diphtheriae: разнообразие генома, структура популяции и перспективы генотипирования». Инфекция, генетика и эволюция . 9 (1): 1–15. Бибкод : 2009InfGE...9....1M . дои : 10.1016/j.meegid.2008.09.011 . ПМИД 19007916 .
- ^ Чарльз Р.К., Райан Э.Т. (октябрь 2011 г.). «Холера в XXI веке». Современное мнение об инфекционных заболеваниях . 24 (5): 472–477. дои : 10.1097/QCO.0b013e32834a88af . ПМИД 21799407 . S2CID 6907842 .
- ^ Кин EC (декабрь 2012 г.). «Парадигмы патогенеза: воздействие на мобильные генетические элементы болезней» . Границы клеточной и инфекционной микробиологии . 2 : 161. дои : 10.3389/fcimb.2012.00161 . ПМЦ 3522046 . ПМИД 23248780 .
- ^ Друлис-Кава З., Майковска-Скробек Г., Мациевска Б. (2015). «Бактериофаги и фаговые белки – подходы к применению» . Современная медицинская химия . 22 (14): 1757–1773. дои : 10.2174/0929867322666150209152851 . ПМК 4468916 . PMID 25666799 .
- ^ Габашвили И.С., Хан С.А., Хейс С.Дж., Сервер П. (октябрь 1997 г.). «Полиморфизм бактериофага Т7». Журнал молекулярной биологии . 273 (3): 658–667. дои : 10.1006/jmbi.1997.1353 . ПМИД 9356254 .
- ^ Магсуди А., Чаттерджи А., Андрисиоаи I, Перкинс, Северная Каролина (декабрь 2019 г.). «Как работает механизм инъекции фага Т4, включая энергетику, силы и динамический путь» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (50): 25097–25105. Бибкод : 2019PNAS..11625097M . дои : 10.1073/pnas.1909298116 . ПМК 6911207 . ПМИД 31767752 .
- ^ Фирс В., Контрерас Р., Дуринк Ф., Хегеман Г., Изерентант Д., Меррегарт Дж. и др. (апрель 1976 г.). «Полная нуклеотидная последовательность РНК бактериофага MS2: первичная и вторичная структура гена репликазы». Природа . 260 (5551): 500–507. Бибкод : 1976Natur.260..500F . дои : 10.1038/260500a0 . ПМИД 1264203 . S2CID 4289674 .
- ^ Мизуно СМ, Гийомар С, Ру С, Лавин Р, Родригес-Валера Ф, Салливан МБ и др. (февраль 2019 г.). «Многие культивируемые и некультивируемые вирусы кодируют рибосомальные белки» . Природные коммуникации . 10 (1): 752. Бибкод : 2019NatCo..10..752M . дои : 10.1038/s41467-019-08672-6 . ПМК 6375957 . ПМИД 30765709 .
- ^ Снустад Д.П. (август 1968 г.). «Взаимодействия доминирования в клетках Escherichia coli, смешанно инфицированных бактериофагом T4D дикого типа и янтарными мутантами, и их возможные последствия в отношении типа функции гена-продукта: каталитическая или стехиометрическая». Вирусология . 35 (4): 550–563. дои : 10.1016/0042-6822(68)90285-7 . ПМИД 4878023 .
- ^ Этаж Е (февраль 1970 г.). «Взаимодействие морфогенетических генов бактериофага Т4». Журнал молекулярной биологии . 47 (3): 293–306. дои : 10.1016/0022-2836(70)90303-7 . ПМИД 4907266 .
- ^ Петров А.С., Харви С.С. (июль 2008 г.). «Упаковка двухспиральной ДНК в вирусные капсиды: структуры, силы и энергетика» . Биофизический журнал . 95 (2): 497–502. Бибкод : 2008BpJ....95..497P . дои : 10.1529/biophysj.108.131797 . ПМК 2440449 . ПМИД 18487310 .
- ^ Анро С., Пети Массачусетс (ноябрь 2022 г.). «Сигналы, запускающие индукцию профагов в микробиоте кишечника» . Молекулярная микробиология . 118 (5): 494–502. дои : 10.1111/mmi.14983 . ПМЦ 9827884 . ПМИД 36164818 . S2CID 252542284 .
- ^ Каллауэй Э (2017). «Вы говорите по-вирусному? Фаги поймали отправку химических сообщений» . Природа . дои : 10.1038/nature.2017.21313 .
- ^ Эрез З., Стейнбергер-Леви И., Шамир М., Дорон С., Стокар-Авихайль А., Пелег Ю. и др. (январь 2017 г.). «Коммуникация между вирусами определяет решения по лизису-лизогении» . Природа . 541 (7638): 488–493. Бибкод : 2017Natur.541..488E . дои : 10.1038/nature21049 . ПМЦ 5378303 . ПМИД 28099413 .
- ^ Блэк Л.В., Томас Дж.А. (2012). «Конденсированная структура генома». Вирусные молекулярные машины . Достижения экспериментальной медицины и биологии. Том. 726. Спрингер. стр. 469–87. дои : 10.1007/978-1-4614-0980-9_21 . ISBN 978-1-4614-0979-3 . ПМЦ 3559133 . ПМИД 22297527 .
- ^ Аль-Шайеб Б., Сачдева Р., Чен Л.К., Уорд Ф., Мунк П., Девото А. и др. (февраль 2020 г.). «Клады огромных фагов со всей экосистемы Земли» . Природа . 578 (7795): 425–431. Бибкод : 2020Natur.578..425A . дои : 10.1038/s41586-020-2007-4 . ПМК 7162821 . ПМИД 32051592 .
- ^ Хаус Р., Блаше С., Докланд Т., Хаггорд-Люнгквист Е., фон Брунн А., Салас М. и др. (2012). «Белко-белковые взаимодействия бактериофагов» . Достижения в области исследования вирусов . 83 : 219–298. дои : 10.1016/B978-0-12-394438-2.00006-2 . ISBN 978-0-12-394438-2 . ПМЦ 3461333 . ПМИД 22748812 .
- ^ Моррис П., Маринелли Л.Дж., Джейкобс-Сера Д., Хендрикс Р.В., Хэтфулл Г.Ф. (март 2008 г.). «Геномная характеристика микобактериофага Джайлза: доказательства приобретения фагом ДНК хозяина путем незаконной рекомбинации» . Журнал бактериологии . 190 (6): 2172–2182. дои : 10.1128/JB.01657-07 . ПМК 2258872 . ПМИД 18178732 .
- ^ Крупович М., Прангишвили Д., Хендрикс Р.В., Бэмфорд Д.Х. (декабрь 2011 г.). «Геномика бактериальных и архейных вирусов: динамика в прокариотической виросфере» . Обзоры микробиологии и молекулярной биологии . 75 (4): 610–635. дои : 10.1128/MMBR.00011-11 . ПМЦ 3232739 . ПМИД 22126996 .
- ^ Такахаси I, Мармур Дж (февраль 1963 г.). «Замена тимидиловой кислоты на дезоксиуридиловую кислоту в дезоксирибонуклеиновой кислоте трансдуцирующего фага Bacillus subtilis». Природа . 197 (4869): 794–795. Бибкод : 1963Natur.197..794T . дои : 10.1038/197794a0 . ПМИД 13980287 . S2CID 4166988 .
- ^ Кирнос М.Д., Худяков И.Ю., Александрушкина Н.И., Ванюшин Б.Ф. (ноябрь 1977 г.). «2-аминоаденин представляет собой аденин, заменяющий основание в ДНК цианофага S-2L». Природа . 270 (5635): 369–370. Бибкод : 1977Natur.270..369K . дои : 10.1038/270369a0 . ПМИД 413053 . S2CID 4177449 .
- ^ Jump up to: Перейти обратно: а б Клип Е (2009). Системная биология: учебник . Вайнхайм: Wiley-VCH. ISBN 978-3-527-31874-2 . OCLC 288986435 .
- ^ Чжао X, Чен С, Шен В, Хуан Г, Ле С, Лу С и др. (январь 2016 г.). «Глобальный транскриптомный анализ взаимодействий между Pseudomonas aeruginosa и бактериофагом PaP3» . Научные отчеты . 6 : 19237. Бибкод : 2016NatSR...619237Z . дои : 10.1038/srep19237 . ПМК 4707531 . ПМИД 26750429 .
- ^ Блаше С., Вухти С., Раджагопала С.В., Уец П. (декабрь 2013 г.). «Сеть взаимодействия белков бактериофага лямбда с его хозяином, Escherichia coli» . Журнал вирусологии . 87 (23): 12745–12755. дои : 10.1128/JVI.02495-13 . ПМЦ 3838138 . ПМИД 24049175 .
- ^ Бобонис Дж., Митош К., Матеус А., Керхер Н., Критикос Г., Селькриг Дж. и др. (сентябрь 2022 г.). «Бактериальные ретроны кодируют трехсторонние системы токсин-антитоксин, защищающие фага». Природа . 609 (7925): 144–150. Бибкод : 2022Natur.609..144B . дои : 10.1038/s41586-022-05091-4 . ПМИД 35850148 . S2CID 250643138 .
- ^ Ка Д, О Х, Пак Э, Ким Дж. Х., Пэ Э (июнь 2020 г.). «Структурные и функциональные доказательства бактериальной антифаговой защиты защитной системой Теериса через НАД. + деградация» . Nature Communications . 11 (1): 2816. Bibcode : 2020NatCo..11.2816K . doi : 10.1038/s41467-020-16703- . PMC 7272460. . PMID 32499527 w
- ^ Jump up to: Перейти обратно: а б Чеслик М., Багиньска Н., Йоньчик-Матысяк Е., Венгжин А., Венгжин Г., Гурски А. (май 2021 г.). «Умеренные бактериофаги — мощные непрямые модуляторы эукариотических клеток и иммунных функций» . Вирусы . 13 (6): 1013. дои : 10.3390/v13061013 . ПМЦ 8228536 . ПМИД 34071422 .
- ^ Jump up to: Перейти обратно: а б Вендлинг CC, Рефардт Д., Холл А.Р. (февраль 2021 г.). «Польза от фитнеса для бактерий, несущих профаги и кодируемые профагами гены устойчивости к антибиотикам, достигает пика в различных средах» . Эволюция; Международный журнал органической эволюции . 75 (2): 515–528. дои : 10.1111/evo.14153 . ПМЦ 7986917 . ПМИД 33347602 .
- ^ Кирш Дж.М., Бжозовский Р.С., Фейт Д., Раунд Дж.Л., Secor PR, Дюркоп Б.А. (сентябрь 2021 г.). «Взаимодействие бактериофагов и бактерий в кишечнике: от беспозвоночных до млекопитающих» . Ежегодный обзор вирусологии . 8 (1): 95–113. doi : 10.1146/annurev-virology-091919-101238 . ПМЦ 8484061 . ПМИД 34255542 .
- ^ Бренчани А, Баччалья А, Виньяроли С, Пуньялони А, Варальдо П.Е., Джованетти Е (январь 2010 г.). «Phim46.1, основной элемент Streptococcus pyogenes, несущий гены mef(A) и tet(O)» . Антимикробные средства и химиотерапия . 54 (1): 221–229. дои : 10.1128/AAC.00499-09 . ПМК 2798480 . ПМИД 19858262 .
- ^ Ян М.Т., Архипова К., Маркерт С.М., Стиглохер С., Лахнит Т., Пита Л. и др. (октябрь 2019 г.). «Фаговый белок помогает бактериальным симбионтам уклоняться от иммунного ответа у эукариот» . Клетка-хозяин и микроб . 26 (4): 542–550.е5. дои : 10.1016/j.chom.2019.08.019 . ПМИД 31561965 . S2CID 203580138 .
- ^ Ли Б.А. (октябрь 2019 г.). «Сотрудничество в условиях конфликта: профаги защищают бактерии от фагоцитоза» . Клетка-хозяин и микроб . 26 (4): 450–452. дои : 10.1016/j.chom.2019.09.003 . ПМИД 31600498 . S2CID 204243652 .
- ^ Али Й., Коберг С., Хесснер С., Сан Х., Рабе Б., Бэк А. и др. (2014). «Умеренные фаги Streptococcus thermophilus, экспрессирующие белки исключения суперинфекции типа Ltp» . Границы микробиологии . 5 : 98. дои : 10.3389/fmicb.2014.00098 . ПМЦ 3952083 . ПМИД 24659988 .
- ^ МакГрат С., Фицджеральд Г.Ф., ван Синдерен Д. (январь 2002 г.). «Идентификация и характеристика генов фагорезистентности у лактококковых бактериофагов умеренного климата» . Молекулярная микробиология . 43 (2): 509–520. дои : 10.1046/j.1365-2958.2002.02763.x . ПМИД 11985726 . S2CID 7084706 .
- ^ Дауве М., МакГрат Дж., Фицджеральд С., ван Синдерен Г.Ф. Идентификация и характеристика генов исключения суперинфекции, переносимых лактококковыми профагами▿ † . Американское общество микробиологии (ASM). OCLC 679550931 .
- ^ Брюссов Х., Канчая С., Хардт В.Д. (сентябрь 2004 г.). «Фаги и эволюция бактериальных патогенов: от геномных перестроек к лизогенной конверсии» . Обзоры микробиологии и молекулярной биологии . 68 (3): 560–602, оглавление. дои : 10.1128/MMBR.68.3.560-602.2004 . ПМК 515249 . ПМИД 15353570 .
- ^ Эдлин Г., Лин Л., Кудрна Р. (июнь 1975 г.). «Лямбда-лизогены E. coli размножаются быстрее, чем нелизогены». Природа . 255 (5511): 735–737. Бибкод : 1975Natur.255..735E . дои : 10.1038/255735a0 . ПМИД 1094307 . S2CID 4156346 .
- ^ Секулович О., Фортье LC (февраль 2015 г.). Шаффнер Д.В. (ред.). «Глобальный транскрипционный ответ Clostridium difficile, несущего профаг CD38» . Прикладная и экологическая микробиология . 81 (4): 1364–1374. Бибкод : 2015ApEnM..81.1364S . дои : 10.1128/АЕМ.03656-14 . ПМК 4309704 . ПМИД 25501487 .
- ^ Россманн Ф.С., Рачек Т., Вобсер Д., Пучалка Дж., Рабенер Э.М., Рейгер М. и др. (февраль 2015 г.). «Фагово-опосредованное распространение биопленки и распространение бактериальных генов вирулентности индуцируется ощущением кворума» . ПЛОС Патогены . 11 (2): e1004653. дои : 10.1371/journal.ppat.1004653 . ПМК 4338201 . ПМИД 25706310 .
- ^ Обенг Н., Пратама А.А., Эльзас Дж.Д. (июнь 2016 г.). «Значение мутуалистических фагов для бактериальной экологии и эволюции» (PDF) . Тенденции в микробиологии . 24 (6): 440–449. дои : 10.1016/j.tim.2015.12.009 . ПМИД 26826796 . S2CID 3565635 .
- ^ Ли Дж., Кортес М.Х., Душофф Дж., Вайц Дж.С. (июль 2020 г.). «Когда следует соблюдать умеренность: о пользе лизиса для фитнеса по сравнению с лизогенией» . Эволюция вирусов . 6 (2): veaa042. bioRxiv 10.1101/709758 . дои : 10.1093/ve/veaa042 . ПМЦ 9532926 . ПМИД 36204422 .
- ^ Брейтбарт М. , Саламон П., Андресен Б., Махаффи Дж.М., Сегалл А.М., Мид Д. и др. (октябрь 2002 г.). «Геномный анализ некультивируемых морских вирусных сообществ» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (22): 14250–14255. Бибкод : 2002PNAS...9914250B . дои : 10.1073/pnas.202488399 . ПМК 137870 . ПМИД 12384570 .
- ^ Мартин С. (1988). «Применение методов отслеживания бактериофагов в водах юго-запада». Журнал «Вода и окружающая среда» . 2 (6): 638–642. Бибкод : 1988WaEnJ...2..638M . дои : 10.1111/j.1747-6593.1988.tb01352.x .
- ^ Берг О., Борсхайм К.Ю., Братбак Г., Хелдал М. (август 1989 г.). «Высокое количество вирусов обнаружено в водной среде». Природа . 340 (6233): 467–468. Бибкод : 1989Natur.340..467B . дои : 10.1038/340467a0 . ПМИД 2755508 . S2CID 4271861 .
- ^ Кин Э.К., Блисковский В.В., Малагон Ф., Бейкер Дж.Д., Принс Дж.С., Клаус Дж.С. и др. (январь 2017 г.). «Новые бактериофаги-суперраспространители способствуют горизонтальному переносу генов путем трансформации» . мБио . 8 (1): e02115–16. дои : 10.1128/mBio.02115-16 . ПМК 5241400 . ПМИД 28096488 .
- ^ Лекунберри И., Субиратс Х., Боррего К.М., Балькасар Х.Л. (январь 2017 г.). «Изучение вклада бактериофагов в устойчивость к антибиотикам». Загрязнение окружающей среды . 220 (Часть Б): 981–984. Бибкод : 2017EPoll.220..981L . дои : 10.1016/j.envpol.2016.11.059 . hdl : 10256/14115 . ПМИД 27890586 .
- ^ Беавоги А., Лакруа А., Виарт Н., Пулен Дж., Дельмонт Т.О., Паоли Л. и др. (март 2024 г.). «Защита сложных бактериальных сообществ» . Природные коммуникации . 15 (1): 2146. Бибкод : 2024NatCo..15.2146B . дои : 10.1038/s41467-024-46489-0 . ПМЦ 10924106 . ПМИД 38459056 .
- ^ Jump up to: Перейти обратно: а б Манрике П., Болдук Б., Уок С.Т., ван дер Ост Дж., де Вос В.М., Янг М.Дж. (сентябрь 2016 г.). «Здоровый кишечный фагом человека» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (37): 10400–10405. Бибкод : 2016PNAS..11310400M . дои : 10.1073/pnas.1601060113 . ПМК 5027468 . ПМИД 27573828 .
- ^ Минот С., Синха Р., Чен Дж., Ли Х., Кейлбо С.А., Ву Г.Д. и др. (октябрь 2011 г.). «Виром кишечника человека: индивидуальные вариации и динамическая реакция на диету» . Геномные исследования . 21 (10): 1616–1625. дои : 10.1101/гр.122705.111 . ПМК 3202279 . ПМИД 21880779 .
- ^ Jump up to: Перейти обратно: а б с Кирш Дж.М., Бжозовский Р.С., Фейт Д., Раунд Дж.Л., Secor PR, Дюркоп Б.А. (сентябрь 2021 г.). «Взаимодействие бактериофагов и бактерий в кишечнике: от беспозвоночных до млекопитающих» . Ежегодный обзор вирусологии . 8 (1): 95–113. doi : 10.1146/annurev-virology-091919-101238 . ПМЦ 8484061 . ПМИД 34255542 .
- ^ Штраус Дж. Х., Зиншаймер Р. Л. (июль 1963 г.). «Очистка и свойства бактериофага MS2 и его рибонуклеиновой кислоты». Журнал молекулярной биологии . 7 (1): 43–54. дои : 10.1016/S0022-2836(63)80017-0 . ПМИД 13978804 .
- ^ Миллер Э.С., Каттер Э., Мосиг Г., Арисака Ф., Кунисава Т., Рюгер В. (март 2003 г.). «Геном бактериофага Т4» . Обзоры микробиологии и молекулярной биологии . 67 (1): 86–156, оглавление. дои : 10.1128/ММБР.67.1.86-156.2003 . ПМК 150520 . ПМИД 12626685 .
- ^ Акерманн Х.В., Криш Х.М. (6 апреля 2014 г.). «Каталог бактериофагов типа Т4». Архив вирусологии . 142 (12): 2329–2345. дои : 10.1007/s007050050246 . ПМИД 9672598 . S2CID 39369249 .
- ^ Ван Р.Х., Ян С., Лю З, Чжан Ю, Ван Х, Сюй З и др. (январь 2024 г.). «PhageScope: хорошо аннотированная база данных бактериофагов с автоматическим анализом и визуализацией» . Исследования нуклеиновых кислот . 52 (Д1): Д756–Д761. дои : 10.1093/nar/gkad979 . ПМЦ 10767790 . ПМИД 37904614 .
Библиография [ править ]
- Хаузер А.Р., Мексас Дж., Мойр Д.Т. (июль 2016 г.). «За пределами антибиотиков: новые терапевтические подходы к бактериальным инфекциям» . Клинические инфекционные болезни . 63 (1): 89–95. дои : 10.1093/cid/ciw200 . ПМК 4901866 . ПМИД 27025826 .
- Стратди С., Паттерсон Т. (2019). Идеальный хищник . Книги Хачетта . ISBN 978-0-316-41808-9 .
- Хойслер Т. (2006). Вирусы против супербактерий: решение кризиса антибиотиков? . Лондон: Макмиллан. ISBN 978-1-4039-8764-8 .
Внешние ссылки [ править ]
- Абедон СТ. «Группа экологии бактериофагов» . Университет штата Огайо. Архивировано из оригинала 3 июня 2013 года.
- Туртерель С., Блуэн Ю. «Иллюстрации бактериофагов и геномика» . Веб-сайт фагов Орсе . Архивировано из оригинала 29 октября 2013 года . Проверено 24 октября 2013 г.
- «QuipStories: Бактериофаги захватывают свою добычу» (PDF) . ПДБе .
- Флатов I (апрель 2008 г.). «Использование фаговых вирусов для борьбы с инфекцией» . Подкаст «Научная пятница» . ЭНЕРГЕТИЧЕСКИЙ ЯДЕРНЫЙ РЕАКТОР. Архивировано из оригинала 17 апреля 2008 года.
- «Анимация научно правильного бактериофага Т4, нацеленного на бактерии E. coli» . Ютуб. 21 мая 2019 г.
- «Бактериофаг Т4, нацеленный на бактерии E. coli » . Анимация от Hybrid Animation Medical . 21 декабря 2009 г.
- Бактериофаги: что это такое. Презентация профессора Грэма Хэтфулла из Питтсбургского университета на YouTube
