Микробный коврик

Микробный мат — это многослойный лист или биопленка микробных колоний и , состоящая преимущественно из бактерий /или архей . Микробные маты растут на границах между различными типами материалов, в основном на затопленных или влажных поверхностях , но некоторые из них выживают в пустынях. [ 1 ] встречаются как эндосимбионты животных Некоторые из них .
Хотя микробные маты имеют толщину всего несколько сантиметров, они создают широкий спектр внутренней химической среды и, следовательно, обычно состоят из слоев микроорганизмов, которые могут питаться или, по крайней мере, переносить доминирующие химические вещества на своем уровне и которые обычно принадлежат к близкородственным видам. . Во влажных условиях коврики обычно скрепляются слизистыми веществами, выделяемыми микроорганизмами. Во многих случаях некоторые бактерии образуют запутанную сеть нитей , которая делает коврик более жестким. Наиболее известными физическими формами являются плоские маты и короткие столбы, называемые строматолитами , но существуют также сферические формы.
Микробные маты — самая ранняя форма жизни на Земле, существование которой подтверждено убедительными ископаемыми свидетельствами 3500 миллионов лет назад планеты , и они были наиболее важными членами и хранителями экосистем . Первоначально они зависели от гидротермальных источников в качестве источника энергии и химической «пищи», но развитие фотосинтеза позволило матам размножаться за пределами этих сред, используя более широко доступный источник энергии — солнечный свет. Последним и наиболее значимым этапом этого освобождения стало развитие фотосинтеза, производящего кислород, поскольку основными химическими веществами для этого являются углекислый газ и вода.
В результате микробные маты начали создавать ту атмосферу, которую мы знаем сегодня, в которой свободный кислород является жизненно важным компонентом. Примерно в то же время они, возможно, также были местом рождения более сложного эукариот типа клеток , из которых все многоклеточные организмы. состоят [ 2 ] Микробные маты были в изобилии на мелководном морском дне до кембрийской субстратной революции , когда животные, живущие в мелководных морях, увеличили свои способности к рытью норы и, таким образом, разрушили поверхность матов и пропустили насыщенную кислородом воду в более глубокие слои, отравив обитавшие там непереносимые к кислороду микроорганизмы. . Хотя эта революция вытеснила циновки с мягкого дна мелководных морей, они все еще процветают во многих средах, где рытье нор ограничено или невозможно, включая каменистое морское дно и берега, а также гиперсоленые и солоноватые лагуны. Их находят также на дне глубоких океанов.
Из-за способности микробных ковриков использовать в качестве «пищи» практически все, существует значительный интерес к промышленному использованию ковриков, особенно для очистки воды и очистки загрязнений .
Описание
[ редактировать ]
Микробные маты также могут называться водорослевыми матами и бактериальными матами. Они представляют собой тип биопленки , достаточно большой, чтобы ее можно было увидеть невооруженным глазом, и достаточно прочной, чтобы выдерживать умеренные физические нагрузки. Эти колонии бактерий многих типов образуются на поверхностях на границах , например, между водой и отложениями или камнями на дне, между воздухом и породой или отложениями, между почвой и коренной породой и т. д. Такие границы раздела образуют вертикальные химические градиенты , т.е. вертикальные вариации химического состава, которые делают разные уровни пригодными для разных типов бактерий и, таким образом, делят микробные маты на слои, которые могут быть четко выражены или могут более постепенно сливаться друг с другом. [ 3 ] Различные микробы способны преодолевать пределы диффузии, используя «нанопровода» для переноса электронов из метаболических реакций на глубину до двух сантиметров в осадок – например, электроны могут переноситься из реакций с участием сероводорода, находящихся глубже в осадке, в кислород в воде, который действует как акцептор электронов. [ 4 ]
Наиболее известными типами микробного мата могут быть плоские слоистые маты, которые образуются примерно на горизонтальных поверхностях, и строматолиты , короткие столбы, образующиеся по мере того, как микробы медленно движутся вверх, чтобы избежать удушения осадками, осаждаемыми на них водой. Однако существуют также сферические маты, некоторые из которых находятся снаружи комков камня или другого твердого материала, а другие внутри сфер осадка. [ 3 ]
Структура
[ редактировать ]Микробный мат состоит из нескольких слоев, в каждом из которых преобладают определенные виды микроорганизмов , преимущественно бактерий . Хотя состав отдельных матов варьируется в зависимости от окружающей среды, по общему правилу побочные продукты каждой группы микроорганизмов служат «пищей» для других групп. По сути, каждый мат образует свою собственную пищевую цепь с одной или несколькими группами на вершине пищевой цепи, поскольку их побочные продукты не потребляются другими группами. Различные типы микроорганизмов доминируют в разных слоях в зависимости от их сравнительного преимущества для жизни в этом слое. Другими словами, они живут там, где они могут превзойти другие группы, а не там, где им было бы абсолютно комфортно: экологические отношения между различными группами представляют собой комбинацию конкуренции и сотрудничества. Поскольку метаболические возможности бактерий (что они могут «есть» и какие условия они могут переносить) обычно зависят от их филогении (т. е. наиболее близкие группы имеют наиболее схожий метаболизм), разные слои мата делятся как по своим различный метаболический вклад в сообщество и их филогенетические взаимоотношения.
Во влажной среде, где солнечный свет является основным источником энергии, в самых верхних слоях обычно преобладают аэробные фотосинтезирующие цианобактерии (сине-зеленые бактерии, цвет которых обусловлен наличием в них хлорофилла ), тогда как в самых нижних слоях обычно преобладают анаэробные сульфатредуцирующие бактерии. бактерии . [ 5 ] Иногда встречаются промежуточные (окисляемые только в дневное время) слои, населенные факультативно-анаэробными бактериями. Например, в гиперсоленых прудах близ Герреро-Негро (Мексика) были исследованы различные виды матов. Встречаются маты со средним пурпурным слоем, населенным фотосинтезирующими пурпурными бактериями. [ 6 ] Некоторые другие маты имеют белый слой, населенный хемотрофными сероокисляющими бактериями , а под ним оливковый слой, населенный фотосинтезирующими зелеными серными бактериями и гетеротрофными бактериями. [ 7 ] Однако структура этого слоя не является неизменной в течение дня: некоторые виды цианобактерий утром мигрируют в более глубокие слои, а вечером возвращаются обратно, чтобы избежать интенсивного солнечного света и УФ-излучения в полдень. [ 7 ] [ 8 ]
Микробные маты обычно скрепляются и связываются со своими субстратами с помощью слизистых внеклеточных полимерных веществ, которые они выделяют. Во многих случаях некоторые бактерии образуют нити (нити), которые спутываются и тем самым увеличивают структурную прочность колоний, особенно если нити имеют оболочки (жесткие внешние покрытия). [ 3 ]
Такое сочетание слизи и спутанных нитей привлекает другие микроорганизмы, которые становятся частью сообщества коврика, например, простейших , некоторые из которых питаются бактериями, образующими коврик, и диатомеями , которые часто покрывают поверхности погруженных микробных ковриков тонким пергаментным слоем . как покрытия. [ 3 ]
Морские маты могут достигать нескольких сантиметров в толщину, из которых только верхние несколько миллиметров насыщены кислородом. [ 9 ]
Типы колонизированной среды
[ редактировать ]Подводные микробные маты описываются как слои, которые живут за счет использования и в некоторой степени изменения локальных химических градиентов , то есть изменений химического состава. Более тонкие и менее сложные биопленки живут во многих субаэральных средах, например, на камнях, на минеральных частицах, таких как песок, и в почве . Им приходится выживать в течение длительного времени без жидкой воды, часто в спящем состоянии. Микробные маты, обитающие в приливных зонах, например, в солончаках Сиппевиссетт , часто содержат большую долю подобных микроорганизмов, которые могут выжить в течение нескольких часов без воды. [ 3 ]
Микробные маты и менее сложные виды биопленок встречаются в диапазоне температур от –40 °C до +120 °C, поскольку изменения давления влияют на температуру, при которой вода остается жидкой. [ 3 ]
Они даже появляются как эндосимбионты у некоторых животных, например в задних кишках некоторых морских ежей . [ 10 ]
Эколого-геологическое значение
[ редактировать ]


Микробные коврики используют все типы метаболизма и стратегии питания, которые развились на Земле: аноксигенный и кислородный фотосинтез ; анаэробная и аэробная хемотрофия (использование химических веществ вместо солнечного света в качестве источника энергии); органическое и неорганическое дыхание и ферментация (т.е. преобразование пищи в энергию с использованием кислорода в процессе или без него); автотрофия (производство пищи из неорганических соединений) и гетеротрофия (производство пищи только из органических соединений за счет некоторой комбинации хищничества и детритофагии ). [ 3 ]
Большинство осадочных пород и рудных месторождений образовались в результате рифоподобных наростов, а не «выпадения» из воды, и на это наращивание, по крайней мере, повлияло, а иногда, возможно, и вызвало действие микробов. Строматолиты , биогермы (купола или колонны, внутренне похожие на строматолиты) и биостромы (отдельные пласты осадка) относятся к числу таких образований, подвергшихся микробному влиянию. [ 3 ] Другие типы микробного мата создали морщинистую текстуру «слоновьей кожи» в морских отложениях, хотя прошло много лет, прежде чем эти текстуры были признаны следами окаменелостей матов. [ 12 ] Микробные маты увеличили концентрацию металла во многих рудных месторождениях, без этого их добыча была бы невозможна — например, месторождения железа (как сульфидные, так и оксидные), урана, меди, серебра и золота. [ 3 ]
Роль в истории жизни
[ редактировать ]История жизни | ||||||||||||||||||
−4500 — – −4000 — – −3500 — – −3000 — – −2500 — – −2000 — – −1500 — – −1000 — – −500 — – 0 — |
| |||||||||||||||||
Очень краткая история жизни на Земле. Масштаб по оси указан в миллионах лет назад. | ||||||||||||||||||
Самые ранние коврики
[ редактировать ]Микробные маты являются одними из старейших явных признаков жизни, поскольку осадочные структуры, вызванные микробами (MISS), образовавшиеся 3480 миллионов лет назад были обнаружены в западной Австралии . [ 3 ] [ 13 ] [ 14 ] На этом раннем этапе структура матов, возможно, уже была подобна структуре современных матов, не включающих фотосинтезирующие бактерии. Возможно даже, что нефотосинтезирующие маты существовали еще 4000 миллионов лет назад . Если это так, то их источником энергии могли быть гидротермальные источники высокого давления ( горячие источники вокруг затопленных вулканов эволюционный раскол между бактериями и археями . ), и примерно в это же время мог произойти [ 15 ]
Самые ранние маты, возможно, представляли собой небольшие одновидовые биопленки хемотрофов , которые полагались на гидротермальные источники для снабжения как энергией, так и химической «пищей». В течение короткого времени (по геологическим меркам) накопление мертвых микроорганизмов создало бы экологическую нишу для пожирающих гетеротрофов , возможно, выделяющих метан и сульфатредуцирующих организмов, которые сформировали бы новые слои в матах и обогатили бы запас биологических ресурсов. полезные химикаты. [ 15 ]
Фотосинтез
[ редактировать ]Принято считать, что фотосинтез , биологическое производство химической энергии из света, развился вскоре после 3000 миллионов лет назад (3 миллиарда). [ 15 ] Однако изотопный анализ показывает, что кислородный фотосинтез мог быть широко распространен еще 3500 миллионов лет назад . [ 15 ] Существует несколько различных типов фотосинтетических реакций, и анализ бактериальной ДНК показывает, что фотосинтез впервые возник у аноксигенных пурпурных бактерий , тогда как кислородный фотосинтез, наблюдаемый у цианобактерий и гораздо позже у растений, развился последним. [ 16 ]
Самый ранний фотосинтез, возможно, осуществлялся с помощью инфракрасного света с использованием модифицированных версий пигментов , первоначальная функция которых заключалась в обнаружении инфракрасного теплового излучения из гидротермальных источников. Развитие фотосинтетической генерации энергии позволило микроорганизмам сначала колонизировать более обширные территории вокруг жерл, а затем использовать солнечный свет в качестве источника энергии. Роль гидротермальных источников теперь сводилась к поставке восстановленных металлов в океаны в целом, а не к основным источникам жизни в определенных местах. [ 16 ] Гетеротрофные падальщики сопровождали фотосинтезаторов в их миграции из «гидротермального гетто». [ 15 ]
Эволюция пурпурных бактерий, которые не производят и не используют кислород, но могут его переносить, позволила матам колонизировать районы с относительно высокими концентрациями кислорода, токсичного для организмов, не адаптированных к нему. [ 17 ] Микробные маты были бы разделены на окисленные и восстановленные слои, и такая специализация повысила бы их продуктивность. [ 15 ] Возможно, эту модель удастся подтвердить, проанализировав соотношения изотопов углерода и серы в отложениях, отложенных на мелководье. [ 15 ]
Последним крупным этапом эволюции микробных матов стало появление цианобактерий — фотосинтезаторов, которые производят и используют кислород. Это придало подводным матам их типичную современную структуру: богатый кислородом верхний слой цианобактерий; слой фотосинтезирующих пурпурных бактерий, способных переносить кислород; и бескислородные, с преобладанием H 2 S нижние слои гетеротрофных падальщиков, в основном выделяющих метан и сульфатредуцирующих организмов. [ 15 ]
Подсчитано, что появление оксигенного фотосинтеза увеличило биологическую продуктивность в 100–1000 раз. Все фотосинтетические реакции требуют восстановителя , но значение кислородного фотосинтеза состоит в том, что в качестве восстановителя используется вода , а воды гораздо больше, чем восстановителей, образующихся геологическим путем, от которых раньше зависел фотосинтез. Возникающее в результате увеличение популяций фотосинтезирующих бактерий в верхних слоях микробных матов должно было вызвать соответствующий рост популяций хемотрофных и гетеротрофных микроорганизмов, населявших нижние слои и питавшихся соответственно побочными продуктами фотосинтезаторов, а также трупами и трупами. / или живые тела других маточных организмов. Такое увеличение сделало бы микробные маты доминирующими экосистемами планеты. С этого момента сама жизнь производила значительно больше необходимых ей ресурсов, чем геохимические процессы. [ 18 ]
Кислородный фотосинтез в микробных матах также увеличил бы содержание свободного кислорода в атмосфере Земли, как непосредственно за счет выделения кислорода, так и потому, что маты выделяли молекулярный водород (H 2 ), часть которого ускользнула бы из атмосферы Земли, прежде чем он смог бы повторно восстановиться. в сочетании со свободным кислородом образуется больше воды. Таким образом, микробные маты сыграли важную роль в эволюции организмов, которые сначала могли переносить свободный кислород, а затем использовать его в качестве источника энергии. [ 18 ] Кислород токсичен для организмов, не адаптированных к нему, но значительно повышает метаболическую эффективность адаптированных к кислороду организмов. [ 17 ] — например, анаэробное , внутреннего « топлива брожение дает чистый выход двух молекул аденозинтрифосфата » клеток, на молекулу глюкозы , тогда как аэробное дыхание дает чистый выход 36. [ 19 ] Насыщение атмосферы кислородом было предпосылкой для эволюции более сложного эукариот типа клеток все многоклеточные организмы. , из которых построены [ 20 ]
Цианобактерии обладают наиболее полным биохимическим «инструментарием» из всех маткообразующих организмов: механизмами фотосинтеза как зеленых, так и пурпурных бактерий; производство кислорода; и цикл Кальвина , который преобразует углекислый газ и воду в углеводы и сахара . Вполне вероятно, что они приобрели многие из этих подсистем от существующих маточных организмов путем некоторой комбинации горизонтального переноса генов и эндосимбиоза с последующим слиянием. Какими бы ни были причины, цианобактерии являются наиболее самодостаточными из маточных организмов и были хорошо приспособлены к тому, чтобы действовать самостоятельно как в виде плавающих матов, так и в качестве первого представителя фитопланктона , который составляет основу большинства морских пищевых цепей . [ 15 ]
Происхождение эукариотов
[ редактировать ]Время, когда впервые появились эукариоты, до сих пор не установлено: существуют убедительные доказательства того, что окаменелости, датированные периодом от 1600 представляют до 2100 миллионов лет назад, собой эукариотов. [ 21 ] но присутствие стеранов в австралийских сланцах может указывать на то, что эукариоты присутствовали 2700 миллионов лет назад . [ 22 ] До сих пор ведутся споры о происхождении эукариот, и многие теории сосредоточены на идее о том, что бактерия сначала стала эндосимбионтом анаэробного архея, а затем слилась с ним, чтобы стать одним организмом. Если бы такой эндосимбиоз был важным фактором, микробные маты способствовали бы этому. [ 2 ] Известны два варианта этого сценария:
- Граница между насыщенной кислородом и бескислородной зонами коврика сдвинулась бы вверх, когда фотосинтез прекращался ночью, и опускалась вниз, когда фотосинтез возобновлялся после следующего восхода солнца. Симбиоз независимых аэробных и анаэробных организмов позволил бы обоим комфортно жить в зоне, подверженной кислородным «приливам», а последующий эндосимбиоз сделал бы такие партнерства более мобильными. [ 15 ]
- Первоначальное партнерство могло возникнуть между анаэробными археями, которым требовался молекулярный водород (H 2 ), и гетеротрофными бактериями, которые его продуцировали и могли жить как с кислородом, так и без него. [ 15 ] [ 23 ]
Жизнь на суше
[ редактировать ]Микробные коврики, образовавшиеся примерно 1200 миллионов лет назад, являются первым свидетельством существования жизни на Земле. [ 24 ]
Самые ранние многоклеточные «животные»
[ редактировать ]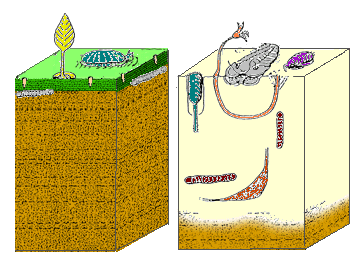
является Биота эдиакары самым ранним широко признанным свидетельством существования многоклеточных «животных». Большинство эдиакарских слоев с текстурой «слоновьей кожи», характерной для микробных матов, содержат окаменелости, а эдиакарские окаменелости почти никогда не встречаются в слоях, не содержащих этих микробных матов. [ 25 ] Адольф Зейлахер классифицировал «животных» как: «инкрустаторы циновки», которые были постоянно прикреплены к циновке; «царапатели ковриков», которые задевали поверхность коврика, не разрушая его; «наклейки на коврик», подвесные кормушки, частично встроенные в коврик; и «подземные шахтеры», которые зарывали норы под мат и питались разлагающимся материалом мата. [ 26 ]
Кембрийская субстратная революция
[ редактировать ]Однако в раннем кембрии организмы начали рыть вертикальные норы в поисках защиты или пищи, разрушая микробные маты и, таким образом, позволяя воде и кислороду проникать на значительное расстояние под поверхность и убивать непереносимые кислород микроорганизмы в нижних слоях. В результате кембрийской субстратной революции морские микробные маты оказались ограничены средами, в которых норное рытье отсутствует или незначительно: [ 27 ] очень суровые условия, такие как гиперсоленые лагуны или солоноватые устья рек, непригодные для обитания роющих организмов, разбивающих маты; [ 28 ] каменистые «полы», сквозь которые не могут проникнуть норы; [ 27 ] глубины океанов, где роющая деятельность сегодня находится на том же уровне, что и в мелких прибрежных морях до революции. [ 27 ]
Текущий статус
[ редактировать ]Хотя кембрийская субстратная революция открыла для животных новые ниши, она не была катастрофической для микробных матов, но значительно сократила их распространение.
Использование микробных матов в палеонтологии.
[ редактировать ]В большинстве окаменелостей сохранились только твердые части организмов, например, раковины. Редкие случаи сохранения мягкотелых окаменелостей (останков мягкотелых организмов, а также мягких частей организмов, у которых обычно обнаруживаются только твердые части, такие как раковины), чрезвычайно ценны, поскольку дают информацию об организмах, которые едва ли можно считать достоверными. когда-либо окаменевших, и гораздо больше информации, чем обычно имеется, о тех, у которых обычно сохраняются только твердые части. [ 29 ] Микробные коврики помогают сохранить мягкие окаменелости за счет:
- Захват трупов на липких поверхностях ковриков и таким образом предотвращение их всплывания или уноса. [ 29 ]
- Физическая защита их от поедания падальщиками и разрушения роющими животными, а также защита отложений, содержащих ископаемые, от эрозии. Например, скорость течения воды, необходимая для размывания связанного матом осадка, в 20–30 раз превышает скорость, необходимую для размывания голого осадка. [ 29 ]
- Предотвращение или уменьшение гниения как путем физической защиты останков от бактерий, вызывающих гниение, так и путем создания химических условий, враждебных к бактериям, вызывающим гниение. [ 29 ]
- Сохранение следов и нор путем защиты их от эрозии. [ 29 ] Многие следы окаменелостей датируются значительно раньше, чем окаменелости тел животных, которые, как считается, были способны их создавать, и таким образом улучшают оценки палеонтологов относительно того, когда впервые появились животные с такими способностями. [ 30 ]
Промышленное использование
[ редактировать ]Способность сообществ микробного мата использовать широкий спектр «пищи» недавно привела к интересу к промышленному использованию. Были проведены испытания микробных матов для очистки воды как для использования человеком, так и в рыбоводстве . [ 31 ] [ 32 ] и исследования их потенциала для очистки разливов нефти . [ 33 ] В результате растущего коммерческого потенциала были поданы заявки и выданы патенты, касающиеся выращивания, установки и использования микробных матов, в основном для очистки загрязняющих веществ и отходов. [ 34 ]
См. также
[ редактировать ]- Биологическая почвенная корка
- Кембрийская революция в субстрате
- Цианобактерии
- Сохранение эдиакарского типа
- Эволюционная история жизни
- Микробиологический коврик Sippewissett
Примечания
[ редактировать ]- ^ Шибер, Дж.; Бозе, П; Эрикссон, П.Г.; Банерджи, С.; Саркар, С.; Альтерманн, В.; Катуняну, О. (2007). Атлас особенностей микробного мата, сохранившихся в летописи кремнеобломочных пород . Эльзевир. ISBN 978-0-444-52859-9 . Проверено 1 июля 2008 г.
- ^ Перейти обратно: а б Нобс, Стефани-Джейн; Маклауд, Фрейзер И.; Вонг, Хон Лун; Бернс, Брендан П. (01 мая 2022 г.). «Эукария-химера: эукариоты, вторичное нововведение двух областей жизни?» . Тенденции в микробиологии . 30 (5): 421–431. дои : 10.1016/j.tim.2021.11.003 . ISSN 0966-842X . ПМИД 34863611 . S2CID 244823103 .
- ^ Перейти обратно: а б с д и ж г час я дж Крумбейн, МЫ; Брем, У.; Гердес, Г.; Горбушина А.А.; Левит, Г.; Палинска, К.А. (2003). «Биопленка, Биодиктион, Биомат Микробиалиты, Оолиты, Строматолиты, Геофизиология, Глобальный механизм, Парагистология». В Крумбайне, МЫ; Патерсон, DM; Заварзин Г.А. (ред.). Ископаемые и современные биопленки: естественная история жизни на Земле (PDF) . Клювер Академик. стр. 1–28. ISBN 978-1-4020-1597-7 . Архивировано из оригинала (PDF) 6 января 2007 года . Проверено 9 июля 2008 г.
- ^ Нильсен, Л.; Рисгаард-Петерсен, Н.; Фоссинг, Х.; Кристенсен, П.; Саяма, М. (2010). «Электрические токи соединяют пространственно разделенные биогеохимические процессы в морских отложениях». Природа . 463 (7284): 1071–1074. Бибкод : 2010Natur.463.1071N . дои : 10.1038/nature08790 . ПМИД 20182510 . S2CID 205219761 .
- Кэтрин Сандерсон (24 февраля 2010 г.). «Бактерии жужжат на морском дне». Новости природы . дои : 10.1038/news.2010.90 .
- ^ Рисатти, Дж.Б.; Кэпман, туалет; Шталь, Д.А. (11 октября 1994 г.). «Структура сообщества микробного мата: филогенетическое измерение» . Труды Национальной академии наук . 91 (21): 10173–7. Бибкод : 1994PNAS...9110173R . дои : 10.1073/pnas.91.21.10173 . ПМК 44980 . ПМИД 7937858 .
- ^ Лукас Дж. Сталь: Физиологическая экология цианобактерий в микробных матах и других сообществах, Новый фитолог (1995), 131, 1–32.
- ^ Перейти обратно: а б Гарсия-Пихель Ф., Мехлинг М., Кастенхольц Р.В., Дильские миграции микроорганизмов в бентосном сообществе гиперсоленого мата , Appl. и окр. Микробиология, май 1994 г., стр. 1500–1511.
- ^ Бебут Б.М., Гарсия-Пичел Ф., Вертикальные миграции цианобактерий в микробном мате , индуцированные УФ-В, Appl. Окружающая среда. Микробиол., декабрь 1995 г., 4215–4222, Том 61, № 12.
- ^ Че, Л.М.; Андрефуэ. С.; Боторель, В.; Гезеннек, М.; Ружо, Х.; Гезеннек, Ж.; Десландес, Э.; Трише, Дж.; Матерон, Р.; Ле Кэмпион, Т.; Пайри, К.; Кометт, П. (2001). «Физические, химические и микробиологические характеристики микробных матов (КОПАРА) на атоллах южной части Тихого океана Французской Полинезии» . Канадский журнал микробиологии . 47 (11): 994–1012. дои : 10.1139/cjm-47-11-994 . ПМИД 11766060 . Проверено 18 июля 2008 г. [ постоянная мертвая ссылка ]
- ^ Темара, А.; де Риддер, К.; Куэнен, Дж.Г.; Робертсон, Луизиана (февраль 1993 г.). «Сульфидокисляющие бактерии роющего ежа Echinocardium cordatum (Echinodermata)» . Морская биология . 115 (2): 179. Бибкод : 1993МарБи.115..179Т . дои : 10.1007/BF00346333 . S2CID 85351601 .
- ^ Порада Х.; Гергут Дж.; Буугри Эль Х. (2008). «Морщинистые структуры типа Киннея - критический обзор и модель формирования». ПАЛЕОС 23 (2): 65–77. Бибкод : 2008Представитель..23...65П . дои : 10.2110/palo.2006.p06-095r . S2CID 128464944 .
- ^ Перейти обратно: а б Мантен, А.А. (1966). «Некоторые проблемные мелководно-морские структуры» . Морской геолог . 4 (3): 227–232. Бибкод : 1966МГеол...4..227М . дои : 10.1016/0025-3227(66)90023-5 . hdl : 1874/16526 . S2CID 129854399 . Архивировано из оригинала 21 октября 2008 г. Проверено 18 июня 2007 г.
- ^ Боренштейн, Сет (13 ноября 2013 г.). «Найдена самая старая окаменелость: познакомьтесь со своей микробной мамой» . АП Новости . Проверено 15 ноября 2013 г.
- ^ Ноффке, Нора ; Христианин, христианин; Уэйси, Дэвид; Хейзен, Роберт М. (8 ноября 2013 г.). «Микробно-индуцированные осадочные структуры, фиксирующие древнюю экосистему формации Дрессер возрастом около 3,48 миллиардов лет, Пилбара, Западная Австралия» . Астробиология . 13 (12): 1103–24. Бибкод : 2013AsBio..13.1103N . дои : 10.1089/ast.2013.1030 . ПМК 3870916 . ПМИД 24205812 .
- ^ Перейти обратно: а б с д и ж г час я дж к Нисбет, Э.Г. и Фаулер, CMR (7 декабря 1999 г.). «Архейская метаболическая эволюция микробных матов» . Труды Королевского общества Б. 266 (1436): 2375. doi : 10.1098/rspb.1999.0934 . ПМК 1690475 . – аннотация со ссылкой на бесплатный полный контент (PDF)
- ^ Перейти обратно: а б Бланкеншип, RE (1 января 2001 г.). «Молекулярные доказательства эволюции фотосинтеза». Тенденции в науке о растениях . 6 (1): 4–6. дои : 10.1016/S1360-1385(00)01831-8 . ПМИД 11164357 .
- ^ Перейти обратно: а б Абеле, Д. (7 ноября 2002 г.). «Токсичный кислород: радикальный датель жизни» (PDF) . Природа . 420 (27): 27. Бибкод : 2002Natur.420...27A . дои : 10.1038/420027a . ПМИД 12422197 . S2CID 4317378 .
- ^ Перейти обратно: а б Хелер, ТМ; Бебут, Б.М.; Де Марэ, диджей (19 июля 2001 г.). «Роль микробных матов в производстве восстановленных газов на ранней Земле». Природа . 412 (6844): 324–7. Бибкод : 2001Natur.412..324H . дои : 10.1038/35085554 . ПМИД 11460161 . S2CID 4365775 .
- ^ «Введение в аэробное дыхание» . Калифорнийский университет в Дэвисе. Архивировано из оригинала 8 сентября 2008 года . Проверено 14 июля 2008 г.
- ^ Хеджес, Южная Каролина; Блэр, Дж. Э.; Вентури, ML; Шу, JL (28 января 2004 г.). «Молекулярная временная шкала эволюции эукариот и возникновения сложной многоклеточной жизни» . Эволюционная биология BMC . 4 :2. дои : 10.1186/1471-2148-4-2 . ПМК 341452 . ПМИД 15005799 .
- ^ Нолл, Эндрю Х.; Яво, Э.Дж.; Хьюитт, Д.; Коэн, П. (2006). «Эукариотические организмы в протерозойских океанах» . Философские труды Королевского общества Б. 361 (1470): 1023–38. дои : 10.1098/rstb.2006.1843 . ПМЦ 1578724 . ПМИД 16754612 .
- ^ Брокс, Джей-Джей; Логан, Джорджия; Бьюик, Р.; Вызов, RE (13 августа 1999 г.). «Архейские молекулярные окаменелости и раннее появление эукариотов». Наука . 285 (5430): 1033–6. Бибкод : 1999Sci...285.1033B . CiteSeerX 10.1.1.516.9123 . дои : 10.1126/science.285.5430.1033 . ПМИД 10446042 .
- ^ Мартин В. и Мюллер, М. (март 1998 г.). «Водородная гипотеза первых эукариот» . Природа . 392 (6671): 37–41. Бибкод : 1998Natur.392...37M . дои : 10.1038/32096 . ПМИД 9510246 . S2CID 338885 . Проверено 16 июля 2008 г.
- ^ Праве, А.Р. (2002). «Жизнь на суше в протерозое: свидетельства торридонских пород северо-западной Шотландии». Геология . 30 (9): 811–812. Бибкод : 2002Geo....30..811P . doi : 10.1130/0091-7613(2002)030<0811:LOLITP>2.0.CO;2 . ISSN 0091-7613 .
- ^ Раннегар, Б.Н.; Федонкин, М.А. (1992). «Окаменелости тел протерозойских многоклеточных животных». В Шопфе, WJ; Кляйн, К. (ред.). Протерозойская биосфера . Издательство Кембриджского университета. стр. 369–388. ISBN 978-0-521-36615-1 .
- ^ Сейлахер, А. (1999). «Образ жизни, связанный с биоматом, в докембрии» . ПАЛЕОС . 14 (1): 86–93. Бибкод : 1999Палай..14...86С . дои : 10.2307/3515363 . JSTOR 3515363 . Проверено 17 июля 2008 г.
- ^ Перейти обратно: а б с Боттьер, диджей; Хагадорн, JW; Дорнбос, ЮК «Кембрийская революция субстрата» (PDF) . Амхерстский колледж . Архивировано из оригинала (PDF) 9 сентября 2006 г. Проверено 28 июня 2008 г.
- ^ Зейлахер, Адольф; Луис А. Буатоисб; М. Габриэла Мангано (07 октября 2005 г.). «Следы окаменелостей в период эдиакарско-кембрийского перехода: поведенческая диверсификация, экологический оборот и экологический сдвиг». Палеогеография, Палеоклиматология, Палеоэкология . 227 (4): 323–56. Бибкод : 2005PPP...227..323S . дои : 10.1016/j.palaeo.2005.06.003 .
- ^ Перейти обратно: а б с д и Бриггс, DEG (2003). «Роль биопленок в окаменении небиоминерализованных тканей». В Крумбайне, МЫ; Патерсон, DM; Заварзин Г.А. (ред.). Ископаемые и современные биопленки: естественная история жизни на Земле . Клювер Академик. стр. 281–290. ISBN 978-1-4020-1597-7 . Проверено 9 июля 2008 г.
- ^ Сейлахер, А. (1994). «Насколько достоверна стратиграфия Круцианы?». Международный журнал наук о Земле . 83 (4): 752–8. Бибкод : 1994ГеоРу..83..752С . дои : 10.1007/BF00251073 . S2CID 129504434 .
- ^ Поттс, Д.А.; Патенауд, Эль; Гёррес, Дж. Х.; Амадор, Дж. А. «Реконструкция сточных вод и гидравлические характеристики низкопрофильной системы выщелачивания» (PDF) . ГеоМатрикс, Инк . Проверено 17 июля 2008 г. [ мертвая ссылка ]
- ^ Бендер, Дж. (август 2004 г.). «Система очистки сточных вод на основе микробных матов для марикультуры черноморского окуня Centropristis striata на оборотной воде» . Аквакультурная инженерия . 31 (1–2): 73–82. Бибкод : 2004AqEng..31...73B . дои : 10.1016/j.aquaeng.2004.02.001 . Проверено 17 июля 2008 г.
- ^ «Роль микробных матов в биоремедиации прибрежных зон, загрязненных углеводородами» . ISTworld. Архивировано из оригинала 23 июля 2011 г. Проверено 17 июля 2008 г.
- ^ «Композиции и способы использования сконструированных микробных матов – Патент США 6033559» . Проверено 17 июля 2008 г. ; «Система и способ силосно-микробного мата - патент США 5522985» . Проверено 17 июля 2008 г. ; «ГеоМат» . ГеоМатрикс, Инк . Проверено 17 июля 2008 г. [ мертвая ссылка ] цитирует патенты США 7351005 и 7374670.
Ссылки
[ редактировать ]- Зекбах С. (2010) Микробные коврики: современные и древние микроорганизмы в стратифицированных системах Springer, ISBN 978-90-481-3798-5 .
Внешние ссылки
[ редактировать ]- Юрген Шибер. «Страница с микробным ковриком» . Проверено 1 июля 2008 г. – контуры микробных матов и изображения матов в различных ситуациях и при различных увеличениях.
