Кембрийская революция в субстрате
| Часть серии о |
| Кембрийский взрыв |
|---|
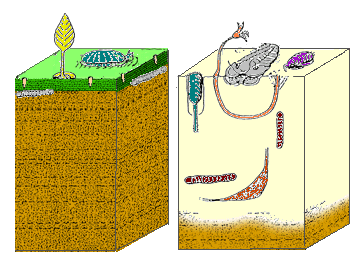
« Кембрийская субстратная революция ». [1] или « Агрономическая революция », [2] свидетельствуют Следы окаменелостей о внезапном разнообразии роющих животных животных в раннем кембрийском периоде.
До этого «расширения поведенческого репертуара» [3] донные животные в основном паслись на микробных матах , выстилавших поверхность субстрата, ползая сверху (как до сих пор делают пресноводные улитки ) или зарывая норы чуть ниже них. Эти микробные маты создавали барьер между водой и отложениями под ними , которые были менее заболоченными, чем современное морское дно, и почти полностью бескислородными (не содержали кислорода ). В результате субстрат был заселен сульфатредуцирующими бактериями , чьи выбросы сероводорода (H 2 S) сделали субстрат токсичным для большинства других организмов. [4]
Примерно в начале кембрия организмы начали рыть норки вертикально, образуя большое разнообразие различных форм и следов окаменелых нор, проникая в отложения в целях защиты или питания. [5] Эти роющие животные разрушали и ослабляли микробные маты, позволяя воде и кислороду проникать на значительное расстояние под поверхность. Это ограничило сульфатредуцирующие бактерии и их выбросы H 2 S в более глубокие слои, сделав верхние слои морского дна пригодными для обитания гораздо более широкого круга организмов. Верхний уровень морского дна стал более влажным и мягким, поскольку его постоянно вспахивали норы. [1]
Закапывание до кембрия
[ редактировать ]
Следы передвижения организмов по микробным матам, покрывавшим дно Эдиакарского моря, и непосредственно под ними сохранились с эдиакарского периода , около 565 миллионов лет назад . Единственные эдиакарские норы расположены горизонтально, на поверхности или чуть ниже ее, и были сделаны животными, которые питались над поверхностью, но зарывали норы, чтобы спрятаться от хищников. [6] Если эти норы биогенны (созданы организмами), это означает наличие подвижных организмов с головами, которые, вероятно, были билатеранами (двусторонне-симметричными животными). [7] Предполагаемые «норы», датируемые еще 1100 миллионами лет назад , могли быть проложены животными, которые питались нижней стороной микробных матов, которые защищали их от химически неприятного океана; [8] однако их неравномерная ширина и сужающиеся концы не позволяют поверить в то, что они были созданы живыми организмами. [9] и первоначальный автор предположил, что мениски лопнувших пузырей, скорее всего, создали следы, которые он наблюдал. [10] Обнаруженные до сих пор эдиакарские норы демонстрируют простое поведение, а сложные и эффективные следы питания, характерные для начала кембрия, отсутствуют.
Некоторые простые докембрийские горизонтальные следы могли быть оставлены крупными одноклеточными организмами; эквивалентные следы производятся сегодня протистами. [11]
Раннекембрийское разнообразие форм нор.
[ редактировать ]С самого начала кембрийского периода [12] (около 538,8 миллионов лет назад ) [13] Впервые появляются многие новые типы следов, в том числе хорошо известные вертикальные норы, такие как Diplocraterion и Skolithos , а также следы, обычно приписываемые членистоногим , таким как Cruziana и Rusophycus . Вертикальные норы указывают на то, что червеобразные животные приобрели новое поведение и, возможно, новые физические способности. Некоторые окаменелости кембрийского периода указывают на то, что их создатели обладали твердыми (хотя и не обязательно минерализованными) экзоскелетами . [14]
Важно проводить различие между закапыванием нор, при котором в отложения поступает кислород (биорорригация), и теми, которые просто перемещают зерна отложений (биосмешение). Последний может фактически уменьшить количество кислорода, доступного в отложениях, перенося органический материал на глубину, где он будет вдыхаться анаэробно. С точки зрения ихноразнообразия, одинаковые пропорции этих двух режимов (преимущественно биосмешивания) наблюдаются по обе стороны границы эдиакарско-кембрийского периода – даже если случаи биоорошения становятся относительно более частыми в терренёвском периоде. [15]
Преимущества рытья
[ редактировать ]Кормление
[ редактировать ]Многие организмы роют норы, чтобы получить пищу либо в виде других роющих организмов, либо в виде органического вещества. Остатки планктонных организмов опускаются на морское дно, обеспечивая источник питания; если эти органические вещества смешаны с осадком, ими можно питаться. Однако возможно, что до кембрия планктон был слишком мал, чтобы тонуть, поэтому на морское дно не было поступления органического углерода. [16] Однако похоже, что организмы питались самими отложениями только после кембрия. [17]
Анкоридж
[ редактировать ]Преимуществом жизни внутри субстрата будет защита от смыва потоками. [ нужна проверка ]
Защита
[ редактировать ]Организмы также роют норы, чтобы избежать нападения хищников. Хищническое поведение впервые появилось более 1 миллиарда лет назад, но хищничество крупных организмов, по-видимому, впервые стало значительным незадолго до начала кембрия. Докембрийские норы выполняли защитную функцию, поскольку животные, их создавшие, кормились над поверхностью; они развивались в то же время, когда другие организмы начали формировать минерализованные скелеты. [6]
Включение рытья
[ редактировать ]Микробные маты образовывали покров, отрезая нижележащие отложения от океанской воды наверху. Это означало, что отложения были бескислородными и сероводородными ( H 2 S ) было в изобилии. Свободный обмен поровых вод с кислородсодержащей океанской водой был необходим для того, чтобы сделать отложения пригодными для жизни. Этот обмен стал возможен благодаря действиям мелких животных: слишком маленькие, чтобы создавать собственные норы, эта мейофауна населяла пространства между песчинками в микробных матах. Их биотурбация – движение, которое смещало зерна и нарушало устойчивость устойчивых биоматов – разрушало маты, позволяя воде и химикатам сверху и снизу смешиваться. [5]
Последствия революции
[ редактировать ]Кембрийская революция субстрата представляла собой долгий и неоднородный процесс, который протекал с разной скоростью в разных местах на протяжении большей части кембрия . [18]
Воздействие на экосистемы
[ редактировать ]После агрономической революции микробные маты , покрывавшие дно Эдиакарского моря, стали все более ограничиваться ограниченным кругом сред:
- Очень суровые условия, такие как гиперсоленые лагуны или солоноватые устья рек, которые были непригодны для обитания роющих организмов, разбивающих маты. [5]
- Каменистые грунты, куда не могли проникнуть норы. [1]
- Глубины океанов, где роющая деятельность сегодня находится на таком же уровне, как и в мелководных прибрежных морях до революции. [1]
Первые норы, вероятно, питались микробными матами, одновременно закапываясь под них в целях защиты; это рытье привело к падению циновок, на которых они питались. [6]
До революции донные организмы делились на четыре категории: [1]
- «инкрустеры на коврике», которые были постоянно прикреплены к коврику;
- «царапатели ковриков», которые задевали поверхность коврика, не разрушая его;
- «наклейки на коврик», подвесные кормушки, частично встроенные в коврик; и
- «подземные шахтеры», которые зарывали норы под мат и питались разлагающимся материалом мата.
«Подземные горняки», судя по всему, вымерли к середине кембрийского периода. [5] «Инкрустеры на ковриках» и «наклейки на коврики» либо вымерли, либо появились более надежные крепления , предназначенные для мягких или твердых оснований. «Когтеточки» были ограничены каменистым субстратом и глубинами океана, где могли выжить и они, и циновки. [1]

Ранние сидячие иглокожие в основном представляли собой «матовые наклейки». Геликоплакоиды не смогли адаптироваться к новым условиям и вымерли; эдриоастероиды выжили , и эокриноиды разработав опоры для прикрепления к твердым субстратам и стебли, которые поднимали их питающий аппарат над большей частью мусора, который роющие норы поднимали на более рыхлом морском дне. Подвижные иглокожие ( стилофоры , гомостелы , гомойостелены и ктеноцистоиды ) существенно не пострадали от революции субстрата. [1]
Ранние моллюски , по-видимому, паслись на микробных циновках, поэтому естественно предположить, что пасущиеся моллюски также были ограничены областями, где циновки могли выжить. Самые ранние известные окаменелости моллюсков -моноплакофоров («одинарных пластинок») датируются ранним кембрием, когда они паслись на микробных матах. Большинство современных моноплакофор живут на мягких субстратах в глубоководных частях морей, хотя один род обитает на твердых субстратах по краям континентальных шельфов . К сожалению, самые старые известные окаменелости полиплакофоров (моллюсков с множественными панцирными пластинами) относятся к позднему кембрию, когда революция субстратов существенно изменила морскую среду. Поскольку они встречаются в строматолитах (коротких столбах, построенных некоторыми типами колоний микробных матов), предполагается, что полиплакофоры паслись на микробных матах. Современные полиплакофоры в основном пасутся на циновках на скалистых берегах, хотя некоторые из них живут и в глубоком море. [1] обнаружено не было Никаких окаменелостей аплакофоров (моллюсков без раковины), которые обычно считаются наиболее примитивными из ныне живущих моллюсков, . Некоторые зарываются в морское дно глубоких вод, питаясь микроорганизмами и детритом; другие живут на рифах и поедают коралловые полипы. [19]
Палеонтологическое значение
[ редактировать ]Революция положила конец условиям, которые позволили исключительно сохранившимся ископаемым пластам или лагерштеттенам, таким как сланцы Бёрджесс . образоваться [5] Прямое потребление туш имело относительно незначительное значение для снижения окаменения по сравнению с изменениями в химическом составе, пористости и микробиологии отложений, что затрудняло развитие химических градиентов, необходимых для развития минерализации мягких тканей. [20] Как и в случае с микробными матами, среда, которая могла вызвать этот способ окаменения, все больше ограничивалась более суровыми и глубокими областями, где землекопы не могли закрепиться; с течением времени степень закапывания нор увеличилась настолько, что этот способ сохранения стал невозможным. [20] Посткембрийские лагерштеттены такого типа обычно встречаются в очень необычных условиях.
Рост популярности нор имеет еще одно значение, поскольку норы являются убедительным доказательством существования сложных организмов; их также гораздо легче сохранить, чем окаменелости тел, до такой степени, что отсутствие следов окаменелостей использовалось для обозначения подлинного отсутствия крупных подвижных донных организмов. Это расширяет понимание палеонтологами раннего кембрия и предоставляет дополнительные доказательства того, что кембрийский взрыв представляет собой реальную диверсификацию, а не является консервационным артефактом - даже если его время не совпадало напрямую с агрономической революцией. [5]
Рост рытья нор представляет собой настолько фундаментальное изменение в экосистеме , что появление сложной норы Treeptichnus pedum используется для обозначения основания кембрийского периода. [12]
Геохимическое значение
[ редактировать ]Повышенный уровень биотурбации означал, что сера, которая постоянно поступает в океаническую систему из вулканов и речных стоков, легче окислялась - вместо того, чтобы быстро захораниваться и оставаться в восстановленной форме (сульфид), роющие организмы постоянно подвергали ее воздействию кислорода. , что позволяет окислить его до сульфата. Предполагается, что эта деятельность объясняет внезапное повышение концентрации сульфатов, наблюдаемое у основания кембрия; это можно записать в геохимических записях как с помощью δ, так и с помощью δ 34 S- изотопные индикаторы и путем количественного определения содержания сульфатного минерала гипса . [21]
См. также
[ редактировать ]Дальнейшее чтение
[ редактировать ]Кэллоу, RHT; Брейзер, доктор медицины (2009). «Замечательная сохранность микробных матов в неопротерозойских кремниевых обломках: значение для эдиакарских тафономических моделей». Обзоры наук о Земле . 96 (3): 207–219. Бибкод : 2009ESRv...96..207C . CiteSeerX 10.1.1.426.2250 . doi : 10.1016/j.earscirev.2009.07.002 .
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час Боттьер, диджей; Хагадорн, JW; Дорнбос, SQ (сентябрь 2000 г.). «Кембрийская революция субстратов» (PDF) . ГСА сегодня . Том. 10, нет. 9. С. 1–9 . Проверено 28 июня 2008 г.
- ^ Сейлахер, А.; Пфлюгер, Ф. (1994). «От биоматов к придонному сельскому хозяйству: биоисторическая революция». В Крумбайне, МЫ; Петерсон, Д.М.; Сталь, ЖЖ (ред.). Биостабилизация осадков . Библиотека и информационная система Ольденбургского университета имени Карла фон Осецкого. стр. 97–105. ISBN 3-8142-0483-2 .
- ^ С. Конвей Моррис, 2001 г.
- ^ Бейли, СП; Корсетти, ФА; Боттьер, диджей; Маренко, КН (июнь 2006 г.). «Микробно-опосредованное влияние окружающей среды на колонизацию многоклеточными животными экосистем грунта: данные формации Харклесс нижнего кембрия». ПАЛЕОС . 21 (3): 215. Бибкод : 2006Palai..21..215B . дои : 10.2110/palo.2005-p05-51e . S2CID 130690224 .
- ^ Jump up to: а б с д и ж Зейлахер, Адольф; Луис А. Буатоисб; М. Габриэла Мангано (07 октября 2005 г.). «Следы окаменелостей в период эдиакарско-кембрийского перехода: поведенческая диверсификация, экологический оборот и экологический сдвиг». Палеогеография, Палеоклиматология, Палеоэкология . 227 (4): 323–356. Бибкод : 2005PPP...227..323S . дои : 10.1016/j.palaeo.2005.06.003 .
- ^ Jump up to: а б с Дзик, Дж. (2007), «Синдром Вердена: одновременное возникновение защитных доспехов и инфаунистических укрытий на этапе докембрия-кембрия», в Викерс-Рич, Патрисия; Комарауэр, Патрисия (ред.), Взлет и падение эдиакарской биоты , Специальные публикации, том. 286, Лондон: Геологическое общество, стр. 405–414, номер doi : 10.1144/SP286.30 , ISBN. 978-1-86239-233-5 , OCLC 156823511
- ^ Федонкин, М.А. (1992). Фауны венда и ранняя эволюция Metazoa . Спрингер. стр. 87–129. ISBN 978-0-306-44067-0 . Проверено 8 марта 2007 г.
- ^ Сейлахер, А .; Бозе, ПК; Пфлюгер, Ф. (2 октября 1998 г.). «Триплобластические животные, жившие более 1 миллиарда лет назад: следы ископаемых свидетельств из Индии». Наука . 282 (5386): 80–83. Бибкод : 1998Sci...282...80S . дои : 10.1126/science.282.5386.80 . ПМИД 9756480 .
- ^ Бадд, GE; Дженсен, С. (2000). «Критическая переоценка летописи окаменелостей двусторонних типов» (аннотация) . Биологические обзоры . 75 (2): 253–295. дои : 10.1111/j.1469-185X.1999.tb00046.x . ПМИД 10881389 . S2CID 39772232 . Проверено 27 июня 2007 г.
- ^ Дженсен, С. (2008). «ПАЛЕОНТОЛОГИЯ: Чтение поведения по камням». Наука . 322 (5904): 1051–1052. дои : 10.1126/science.1166220 . S2CID 129734373 .
- ^ Мац, В.; Франк, М.; Маршалл, Дж.; Виддер, А.; Джонсен, С. (декабрь 2008 г.). «Гигантский глубоководный протист оставляет следы, похожие на двусторонние» . Современная биология . 18 (23): 1849–1854. дои : 10.1016/j.cub.2008.10.028 . ISSN 0960-9822 . ПМИД 19026540 . S2CID 8819675 .
- ^ Jump up to: а б хотя с тех пор он был обнаружен в нижних, технически «докембрийских», слоях.
- Гелинг, Дж.; Дженсен, СР; Дрозер, М.; Мироу, П.; Нарбонн, Г. (март 2001 г.). «Роясь под базальным кембрийским GSSP, Форчун-Хед, Ньюфаундленд». Геологический журнал . 138 (2): 213–218. Бибкод : 2001ГеоМ..138..213Г . дои : 10.1017/S001675680100509X . S2CID 131211543 .
- ^ «Последняя версия международной хроностратиграфической карты» . Международная комиссия по стратиграфии . Проверено 7 апреля 2024 г.
- ^ Дженсен, С. (2003). «Протерозойские и самые ранние кембрийские следы ископаемых; закономерности, проблемы и перспективы» . Интегративная и сравнительная биология . 43 (1): 219–28. дои : 10.1093/icb/43.1.219 . ПМИД 21680425 .
- ^ Элисон Т. Крибб; Себастьян Дж. ван де Вельде; Уильям М. Берельсон; Дэвид Дж. Боттьер; Фрэнк А. Корсетти (22 февраля 2023 г.). «Эдиакарско-кембрийская биотурбация не привела к значительному насыщению кислородом отложений в мелководных морских экосистемах» . Геобиология (Препринт). 21 (4): 435–453. Бибкод : 2023Gbio...21..435C . дои : 10.1111/gbi.12550 . ПМИД 36815223 .
- ^ Баттерфилд, Нью-Джерси (2000). «Экология и эволюция кембрийского планктона». У Журавлев А.; Райдинг, Р. (ред.). Экология кембрийской радиации . Издательство Колумбийского университета. стр. 200–216. дои : 10.7312/zhur10612-009 . ISBN 978-0-231-50516-1 .
- ^
- ^ Дорнбос, С.; Боттьер, Д.; Чен, Ж.-Ю. (июнь 2004 г.). «Доказательства существования микробных матов морского дна и связанного с ними образа жизни многоклеточных животных в нижнекембрийских фосфоритах Юго-Западного Китая» (PDF) . Летайя . 37 (2): 127–137. дои : 10.1080/00241160410004764 . Архивировано из оригинала (PDF) 6 января 2007 г. Проверено 4 августа 2008 г.
- ^ «Аплакофора» . Музей палеонтологии Калифорнийского университета . Проверено 3 июля 2008 г.
- ^ Jump up to: а б Орр, ПиДжей; Бентон, MJ ; Бриггс, DEG (2003). «Посткембрийское закрытие тафономического окна глубоководного склона-бассейна». Геология . 31 (9): 769. Бибкод : 2003Geo....31..769O . дои : 10.1130/G19193.1 . S2CID 129502810 .
- ^ Кэнфилд, Э.; Фаркуар, Дж. (май 2009 г.). «Эволюция животных, биотурбация и концентрация сульфатов в океанах» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (20): 8123–8127. Бибкод : 2009PNAS..106.8123C . дои : 10.1073/pnas.0902037106 . ISSN 0027-8424 . ПМЦ 2688866 . ПМИД 19451639 .
- Маренко, Кэтрин Н.; Боттьер, Дэвид Дж. (2008). «Важность планолитов в кембрийской субстратной революции» . Палеогеография, Палеоклиматология, Палеоэкология . 258 (3): 189–199. Бибкод : 2008PPP...258..189M . дои : 10.1016/j.palaeo.2007.05.025 .
- Боттьер, Дэвид Дж. (2010). «Кембрийская революция субстрата и ранняя эволюция типов». Журнал наук о Земле . 21 : 21–24. дои : 10.1007/s12583-010-0160-7 . S2CID 129851901 .
- Дорнбос, Стивен К.; Боттьер, Дэвид Дж. (2000). «Эволюционная палеоэкология самых ранних иглокожих: геликоплакоиды и кембрийская субстратная революция» . Геология . 28 (9): 839. Бибкод : 2000Geo....28..839D . doi : 10.1130/0091-7613(2000)28<839:EPOTEE>2.0.CO;2 .