Опабиния
| Опабиния Временной диапазон: средний кембрий ( Улюань ), | |
|---|---|
 | |
| Ископаемый образец ( голотип ) на выставке в Смитсоновском институте в Вашингтоне, округ Колумбия. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Членистоногие |
| Сорт: | † Динокаридид |
| Семья: | † Опабинииды |
| Род: | † Опабиния Уолкотт , 1912 год. |
| Разновидность: | † О. regalis |
| Биномиальное имя | |
| † Королевский двор Уолкотт , 1912 год. | |
Opabinia regalis — вымершее , стеблевой группы членистоногое обнаруженное в среднекембрийских сланцах Бёрджесс -Лагерштетте (505 миллионов лет назад) в Британской Колумбии . [1] Опабиния представляла собой мягкотелое животное длиной тела до 7 см, его сегментированное туловище имело створки по бокам и веерообразный хвост. Голова имеет необычные черты: пять глаз , рот под головой, обращенный назад, и когтистый хоботок , который, вероятно, передал пищу в рот. Опабиния, вероятно, жила на морском дне, используя хоботок для поиска маленькой мягкой пищи. [2] Описано менее двадцати хороших экземпляров; 3 экземпляра Opabinia известно Из слоя Greater Phyllopod , где они составляют менее 0,1% сообщества. [3]
Когда первое тщательное исследование опабинии в 1975 году выявило ее необычные особенности, считалось, что она не связана ни с одним известным типом . [4] или, возможно, родственник предков членистоногих и кольчатых червей . [2] Однако более поздние исследования, начиная с конца 1990-х годов, последовательно подтверждают его родство с базальными членистоногими, наряду с близкородственными радиодонтами ( Anomalocaris и родственники) и жаберными лобоподами ( Kerygmachela и Pambdelurion ). [5] [6] [7] [8] [1] [9] [10]
В 1970-х годах продолжались споры о том, появились ли многоклеточные животные внезапно во время раннего кембрия, в результате события, называемого кембрийским взрывом , или возникли раньше, но не оставив после себя окаменелостей. Поначалу Опабиния рассматривалась как убедительное доказательство «взрывной» гипотезы. [4] Позднее открытие целого ряда подобных лобоподовых животных, некоторые из которых имели более близкое сходство с членистоногими, и развитие идеи о стволовых группах позволили предположить, что ранний кембрий был временем относительно быстрой эволюции , но такой, которую можно было понять, не предполагая любые уникальные эволюционные процессы. [11]
История открытия
[ редактировать ]В 1911 году Чарльз Дулиттл Уолкотт обнаружил в сланцах Бёрджесс девять почти полных окаменелостей Opabinia regalis и несколько окаменелостей, которые он классифицировал как Opabinia? СМИ и опубликовал описание всего этого в 1912 году. [12] Родовое название происходит от перевала Опабин между горами Хунгаби и горами Биддл , к юго-востоку от озера О'Хара , Британская Колумбия, Канада . [12] В 1966–1967 годах Гарри Б. Уиттингтон нашел еще один хороший экземпляр. [13] а в 1975 году он опубликовал подробное описание, основанное на тщательном вскрытии некоторых экземпляров и фотографиях этих экземпляров, освещенных под разными углами. Анализ Уиттингтона не охватывал Опабинию? СМИ ; Экземпляры этого вида Уолкотта не удалось обнаружить в его коллекции. [2] В 1960 году российские палеонтологи описали экземпляры, найденные ими в Норильском районе Сибири , и назвали Opabinia norilica . [14] но эти окаменелости плохо сохранились, и Уиттингтон не считал, что они дают достаточно информации, чтобы отнести их к роду Opabinia . [2]
возникновение
[ редактировать ]Все известные экземпляры опабинии , найденные до сих пор, происходят из « слоя филлопод » сланцев Бёрджесс в канадских Скалистых горах Британской Колумбии. [2]
В 1997 году Бриггс и Недин сообщили из Южной Австралии Emu Bay Shale о новом экземпляре Myoscolex , который сохранился гораздо лучше, чем предыдущие экземпляры, что привело их к выводу, что это близкий родственник Opabinia. [15] - хотя позже эта интерпретация была подвергнута сомнению Дзиком, который вместо этого пришел к выводу, что Миосколекс был кольчатым червем. [16]
Морфология
[ редактировать ]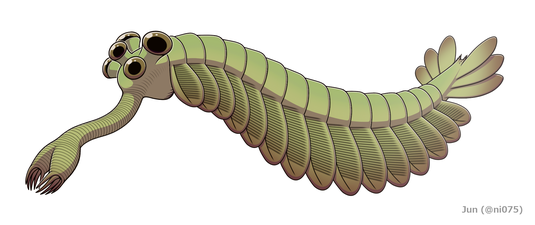 Реставрация
Реставрация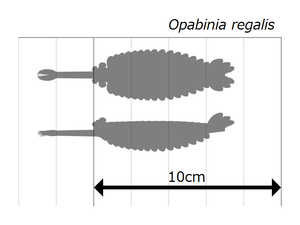 Оценка размера
Оценка размера


Опабиния выглядела настолько странно, что публика на первой презентации анализа Уиттингтона рассмеялась. [2] Длина Opabinia regalis от головы (без хоботка) до конца хвоста колебалась от 4 сантиметров (1,6 дюйма) до 7 сантиметров (2,8 дюйма). [2] Одним из наиболее отличительных признаков опабинии является полый хоботок , общая длина которого составляла около одной трети длины тела и выступал вниз из-под головы. Хоботок был поперечно-полосатый, как шланг пылесоса , гибкий и заканчивался когтеобразной структурой, конечные края которой имели пять шипов, выступающих внутрь и вперед. Двусторонняя симметрия и латеральная (вместо вертикальной, как реконструировал Уиттингтон 1975 г.) [2] ) расположение клешни позволяет предположить, что он представляет собой пару сросшихся лобных придатков, сравнимых с таковыми у радиодонтов и жаберных лобоподий . [17] [5] [18] [19] На голове было пять глаз-стебельков: два спереди и довольно близко к середине головы, направленные вверх и вперед; два более крупных глаза с более длинными стебельками возле заднего и внешнего краев головы, направленные вверх и в стороны; и единственный глаз между большей парой глаз на стебельках, направленный вверх. Предполагалось, что все глаза были сложными других членистоногих . , как и боковые глаза [2] но эта реконструкция, не подкрепленная никакими доказательствами, [2] является «несколько причудливым». [20] Рот находился под головой, за хоботком, и был направлен назад , так что пищеварительный тракт на своем пути к задней части животного образовывал U-образный изгиб. Хоботок оказался достаточно длинным и гибким, чтобы доходить до рта. [2]
Основная часть тела обычно имела ширину около 5 миллиметров (0,20 дюйма) и имела 15 сегментов, на каждом из которых были пары створок (долей), направленных вниз и наружу. Закрылки перекрывались так, что передняя часть каждого закрывалась задней кромкой предыдущего. Тело заканчивалось чем-то похожим на один конический сегмент, несущий три пары перекрывающихся лопастей хвостового вентилятора, направленных вверх и наружу, образуя хвост, похожий на двойной V-образный вентилятор. [2]
Интерпретации других особенностей окаменелостей Опабинии различаются. Поскольку у животных не было ни минерализованной брони, ни даже прочного органического экзоскелета , как у других членистоногих, их тела были уплощенными, когда они были захоронены и окаменели, а более мелкие или внутренние особенности проявлялись в виде отметин внутри очертаний окаменелостей. [2] [5]
![Различные интерпретации створок и жаберных структур Opabinia regalis A: Whittington (1975), [2] B: Bergström (1986), [17] C: Budd (1996), [5] D: Zhang & Briggs (2007), [7] Э: Бадд и Дейли (2011)[8]](//upload.wikimedia.org/wikipedia/commons/thumb/3/34/20210809_Opabinia_regalis_flap_gill_interpretation.png/585px-20210809_Opabinia_regalis_flap_gill_interpretation.png)
![Поперечное сечение Опабинии на основе Бадда и Дейли (2011) [8]](//upload.wikimedia.org/wikipedia/commons/thumb/3/30/20210807_Opabinia_regalis_trunk_cross_section.png/315px-20210807_Opabinia_regalis_trunk_cross_section.png) Опабинии по данным Бадда и Дейли (2011). Поперечное сечение [8]
Опабинии по данным Бадда и Дейли (2011). Поперечное сечение [8]
![Различные интерпретации створок и жаберных структур Opabinia regalis A: Whittington (1975), [2] B: Bergström (1986), [17] C: Budd (1996), [5] D: Zhang & Briggs (2007), [7] Э: Бадд и Дейли (2011)[8]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/34/20210809_Opabinia_regalis_flap_gill_interpretation.png/585px-20210809_Opabinia_regalis_flap_gill_interpretation.png)
![Поперечное сечение Опабинии на основе Бадда и Дейли (2011) [8]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/30/20210807_Opabinia_regalis_trunk_cross_section.png/315px-20210807_Opabinia_regalis_trunk_cross_section.png)
Уиттингтон (1975) интерпретировал жабры как парные расширения, прикрепленные дорсально к основаниям всех створок, кроме первых, с каждой стороны, и полагал, что эти жабры снизу плоские, а сверху имеют перекрывающиеся слои. [2] Бергстрем (1986) обнаружил, что «перекрывающиеся слои» представляют собой ряды отдельных лопастей, и интерпретировал створки как часть спинных покрытий ( тергитов ) на верхней поверхности тела, с лопастями, прикрепленными под каждым из них. [17] [8] Бадд (1996) считал, что жаберные лопасти прикреплены по передним краям на спинной стороне всех, кроме первых створок. Он также обнаружил следы внутри передних краев створок, которые он интерпретировал как внутренние каналы, соединяющие жабры с внутренней частью тела, так же, как Уиттингтон интерпретировал след на хоботке как внутренний канал. [5] Чжан и Бриггс (2007), однако, интерпретировали, что все створки имеют заднее расстояние между местами прикрепления жаберных лопастей. [7] Бадд и Дейли (2011) отвергают реконструкцию Чжана и Бриггса, показывая, что лоскуты имеют полные задние края, как и в предыдущих реконструкциях. В основном они следуют реконструкции Бадда (1996) с модификациями некоторых деталей (например, первая пара створок также имеет жабры; точка прикрепления жаберных лопастей расположена более назад, чем считалось ранее). [8]

Уиттингтон (1975) обнаружил признаки почти треугольных особенностей вдоль тела и пришел к выводу, что это внутренние структуры, скорее всего, боковые продолжения кишечника ( дивертикулы ). [2] Чен и др. (1994) интерпретировали их как находящиеся внутри долей по бокам. [21] Бадд (1996) считал, что «треугольники» слишком широки, чтобы поместиться в опабинии тонком теле , и что изображения в поперечном сечении показали, что они прикреплены отдельно от долей и ниже их и простираются ниже тела. [5] Позже он нашел образцы, у которых, по-видимому, сохранилась внешняя кутикула ног. Поэтому он интерпретировал «треугольники» как короткие, мясистые, конические ноги (лобоподы). Он также нашел небольшие минерализованные пятна на кончиках некоторых из них и интерпретировал их как когти. В рамках этой реконструкции жаберный лоскут и лобопод были гомологизированы внешней жаберной ветвью и внутренней ветвью ноги двуветвистых конечностей членистоногих, наблюдаемых у Марреллы , трилобитов и ракообразных . [5] Чжан и Бриггс (2007) проанализировали химический состав «треугольников» и пришли к выводу, что они имеют тот же состав, что и кишечник, и поэтому согласились с Уиттингтоном в том, что они являются частью пищеварительной системы. Вместо этого они считали и жабр опабинии расположение лопастей ранней формой конечностей членистоногих до того, как оно разделилось на двуветвистую структуру. [7] Однако подобный химический состав связан не только с пищеварительным трактом; Бадд и Дейли (2011) предполагают, что это минерализация, образующаяся в заполненных жидкостью полостях внутри тела, что соответствует полым лобоподам, как это видно на однозначных окаменелостях лобоподий. Они также уточняют, что дивертикулы кишечника Opabinia представляют собой серию круглых кишечных желез, индивидуализированных из «треугольников». Хотя они согласились с отсутствием конечных когтей, присутствие лобоподов в Опабинии остается правдоподобной интерпретацией. [8]
Образ жизни
[ редактировать ]
То, как животные из сланцев Берджесс были погребены оползнем или течением с наносами, которое действовало как песчаная буря, позволяет предположить, что они жили на поверхности морского дна. [2] Опабиния, вероятно, использовала свой хоботок, чтобы искать в осадке частицы пищи и передавать их в рот. [2] Поскольку нет никаких признаков чего-либо, что могло бы функционировать как челюсти, его пища, по-видимому, была маленькой и мягкой. [2] Парные дивертикулы кишечника могут повысить эффективность переваривания и поступления пищи. [22] Уиттингтон (1975), полагая, что у опабинии нет ног, считал, что она ползет на своих лепестках и что она также могла медленно плавать, взмахивая лопастями, особенно если она синхронизировала движения, чтобы создать волну с метахронным движением своих лепестков. [2] С другой стороны, он считал, что тело недостаточно гибкое, чтобы позволять рыбьим колебаниям всего тела. [2]
Классификация
[ редактировать ]
| ||||||||||||||||||||||||||||||||||||||||||
| Обобщенная филогения опабинии и других таксонов экдизозоев . [10] |






палеонтологами Учитывая, насколько различаются реконструкции опабинии , неудивительно, что классификация этого животного широко обсуждалась в 20 веке. [5] Чарльз Дулиттл Уолкотт , первоначальный описатель , в 1912 году считал это аностраком ракообразным- . [12] Эту идею поддержала Дж. Эвелин Хатчинсон в 1930 году, впервые реконструировав опабинию как аностракана, плавающего вверх ногами. [23] Альберто Симонетта представил новую реконструкцию Opabinia в 1970 году, сильно отличающуюся от реконструкции Хатчинсона, с множеством членистоногих особенностей ( например, спинной экзоскелет и суставчатые конечности), которые напоминают Yohoia и Leanchoilia . [24] Лейф Стёрмер , следуя более ранним работам Перси Рэймонда , считал, что опабинии принадлежат к так называемым «трилобитоидам» ( трилобитам и подобным таксонам). После тщательного анализа Гарри Б. Уиттингтон в 1975 году пришел к выводу, что опабиния не была членистоногим, поскольку он не нашел доказательств существования членистоногих конечностей и что у членистоногих не было известно ничего похожего на гибкий, вероятно, заполненный жидкостью хоботок. [2] Хотя он оставил Опабинии классификацию выше уровня семейства открытой, кольчатое, но не сочлененное тело и необычные боковые створки с жабрами убедили его, что это мог быть представитель предкового запаса от кольчатых червей и членистоногих. [2] два отдельных типа животных ( Lophotrochozoan и Ecdysozoan все еще считались близкими родственниками (объединенными под названием Articulata ). соответственно), которые в то время [25]
В 1985 году Дерек Бриггс и Уиттингтон опубликовали крупное переописание Anomalocaris , также из сланцев Бёрджесс. [26] Вскоре после этого шведский палеонтолог Ян Бергстрем , отметив в 1986 году сходство Anomalocaris и Opabinia , предположил, что эти два животных были родственными, поскольку у них были общие многочисленные особенности ( например, боковые створки, жаберные лопасти, стебельчатые глаза и специализированные лобные придатки). Он отнес их к примитивным членистоногим, хотя считал, что членистоногие не представляют собой единого типа . [17]
В 1996 году Грэм Бадд обнаружил, по его мнению, свидетельство коротких, несвязанных ног у Опабинии . [5] Его исследование жаберного лобопода Kerygmachela из Сириус-Пассет лагерштетте , произошедшего около 518 миллионов лет назад и более чем на 10 миллионов лет старше сланцев Бёрджесс, убедило его в том, что у этого экземпляра были похожие ноги. [27] [28] Он считал ноги этих двух родов из сланцев Берджесс очень похожими на ноги лобоподов Aysheaia и современных онихофоранов (бархатных червей), которые считаются носителями многочисленных наследственных черт, общих с предками членистоногих. Изучив несколько наборов черт, общих для этих и подобных им лобоподий, он разработал «широкомасштабную реконструкцию стволовой группы членистоногих », то есть членистоногих и того, что он считал их эволюционными базальными членами. [5] Одной из поразительных особенностей этого генеалогического древа является то, что современные тихоходки (водяные медведи) могут быть родственниками Опабинии ближайшими ныне живущими эволюционными . [5] С другой стороны, Хоу и др. (1995, 2006) предположили, что Opabinia является представителем необычных циклоневральных червей с конвергентными чертами членистоногих. [29] [18]
Хотя Чжан и Бриггс (2007) не согласились с диагнозом Бадда о том, что «треугольники» Опабинии расположением были ногами, сходство, которое они увидели между Опабинии . лопастей и жабр и двуветвистыми конечностями членистоногих, привело их к выводу, что Опабиния очень тесно связана с членистоногими . На самом деле они представили генеалогическое древо, очень похожее на генеалогическое древо Бадда, за исключением того, что в их генеалогическом древе не упоминались тихоходки. [7] Независимо от различных морфологических интерпретаций, все крупные повторные исследования, начиная с 1980-х годов, аналогичным образом пришли к выводу, что сходство между Opabinia и членистоногими ( например, стебельчатые глаза, спинная сегментация, задний рот, сросшиеся придатки, жаберные ветви конечностей) таксономически значимы. [17] [5] [7] [8] [1]
С 2010-х годов предполагаемая тесная связь между Opabinia и тихоходками/циклоневралами больше не поддерживается, в то время как родство Opabinia как членистоногих стеблевой группы наряду с Radiodonta (клада, которая включает Anomalocaris и ее родственников) [30] ) и жаберных лобоподий широко распространены, [1] [9] [10] как последовательно показывают многочисленные филогенетические анализы, [31] [32] [33] [34] [35] [36] [37] [38] [39] [40] [41] [42] а также новые открытия, такие как наличие кишечных желез, подобных членистоногим. [8] [22] и промежуточный таксон Kylinxia . [40]
В 2022 году палеонтологи описали похожее животное, которое было обнаружено в скалах кембрийского возраста в штате Юта . [43] Окаменелость была названа Utaurora comosa и была найдена в сланцах Уиллер . [43] На самом деле членистоногое стебель было впервые описано в 2008 году, но в то время его первоначально считали экземпляром Anomalocaris. [44] Это открытие может указывать на то, что существовали и другие животные, похожие на опабинию, и их семейство могло быть более разнообразным. [43]
Теоретическая значимость
[ редактировать ]
Опабиния ясно показала, как мало было известно о мягкотелых животных, которые обычно не оставляют окаменелостей. [2] Когда Уиттингтон описал это в середине 1970-х годов, уже велись бурные дебаты о ранней эволюции животных . Престон Клауд утверждал в 1948 и 1968 годах, что этот процесс был «взрывным». [45] а в начале 1970-х годов Найлз Элдридж и Стивен Джей Гулд разработали свою теорию прерывистого равновесия , которая рассматривает эволюцию как длинные интервалы почти стазиса, «перемежающиеся» короткими периодами быстрых изменений. [46] С другой стороны, примерно в то же время Уятт Дарем и Мартин Глесснер утверждали, что животное царство имело долгую протерозойскую историю, скрытую из-за отсутствия окаменелостей. [47] [48] Уиттингтон (1975) пришел к выводу, что Opabinia и другие таксоны , такие как Marrella и Yohoia , не могут быть включены в современные группы. Это была одна из основных причин, почему Гулд в своей книге о сланцах Бёрджесс « Чудесная жизнь » считал, что жизнь раннего кембрия была гораздо более разрозненной и «экспериментальной», чем любая более поздняя группа животных, и что кембрийский взрыв был поистине драматическим событием. возможно, обусловлено необычными эволюционными механизмами. [4] Он считал Опабинию настолько важной для понимания этого явления, что хотел назвать свою книгу « Посвящение Опабинии» . [49]
Однако вскоре последовали другие открытия и анализы, в результате которых были обнаружены похожие на вид животные, такие как Anomalocaris из сланцев Берджесс и Kerygmachela из Сириус-Пассет . [17] [27] Другое животное из Берджесс-Шейла, айшея , считалось очень похожим на современных онихофор . [50] которые считаются близкими родственниками членистоногих. [51] Палеонтологи определили группу под названием лобоподии, в которую вошли ископаемые панартроподы, которые считаются близкими родственниками онихофоров, тихоходок и членистоногих, но не имеют членистых конечностей. Позже эта группа была широко принята как парафилетический класс, что привело к возникновению современных типов панартропод.

- — = Линии спуска
- = Базальный узел
- = Узел короны
- = Общая группа
- = Группа короны
- = Группа ствола
В то время как дискуссия о конкретных окаменелостях, таких как Opabinia и Anomalocaris, продолжалась в конце 20 века, была введена концепция стволовых групп, охватывающая эволюционных «тетушек» и «кузенов». Коронная группа — это группа близкородственных живых животных плюс их последний общий предок плюс все его потомки. Стеблевая группа содержит ответвления от представителей линии, предшествующей последнему общему предку кронной группы; это относительное понятие, например, тихоходки — это живые животные, которые сами по себе образуют кронную группу, но Бадд (1996) считал их также стеблевой группой по отношению к членистоногим. [5] [52] Рассматривая таким образом странные на вид организмы, такие как опабинии, можно увидеть, что, хотя кембрийский взрыв был необычным, его можно понять с точки зрения нормальных эволюционных процессов. [11]
См. также
[ редактировать ]- План тела - набор морфологических особенностей, общих для представителей одного типа животных.
- Radiodonta - вымерший отряд базальных членистоногих.
- Палеобиота сланцев Бёрджесс
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Бриггс, Дерек Э.Г. (19 апреля 2015 г.). «Необычайные окаменелости раскрывают природу кембрийской жизни: комментарий к Уиттингтону (1975) «Загадочное животное Opabinia regalis, средний кембрий, сланцы Бёрджесс, Британская Колумбия» » . Философские труды Королевского общества B: Биологические науки . 370 (1666): 20140313. doi : 10.1098/rstb.2014.0313 . ПМК 4360120 . ПМИД 25750235 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и Уиттингтон, HB (июнь 1975 г.). «Загадочное животное Opabinia regalis , сланцы Бёрджесс среднего кембрия, Британская Колумбия». Философские труды Королевского общества Б. 271 (910): 1–43 271. Бибкод : 1975RSPTB.271....1W . дои : 10.1098/rstb.1975.0033 . JSTOR 2417412 . Бесплатная аннотация на Уиттингтон, HB (1975). «Загадочное животное Opabinia regalis , средний кембрий, сланцы Бёрджесс, Британская Колумбия». Философские труды Королевского общества Б. 271 (910): 1–43. Бибкод : 1975RSPTB.271....1W . дои : 10.1098/rstb.1975.0033 .
- ^ Карон, Жан-Бернар; Джексон, Дональд А. (октябрь 2006 г.). «Тафономия сообщества больших слоев филлопод, сланцы Берджесс». ПАЛЕОС . 21 (5): 451–65. Бибкод : 2006Палай..21..451C . дои : 10.2110/palo.2003.P05-070R . JSTOR 20173022 . S2CID 53646959 .
- ^ Перейти обратно: а б с Гулд, С.Дж. (1989). Прекрасная жизнь . Хатчинсон Радиус. стр. 124–136 и далее. ISBN 978-0-09-174271-3 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Бадд, GE (1996). «Морфология Opabinia regalis и реконструкция стеблевой группы членистоногих». Летайя . 29 (1): 1–14. дои : 10.1111/j.1502-3931.1996.tb01831.x .
- ^ Бадд, Грэм Э. (1998). «Морфология и филогенетическое значение Kerygmachela kierkegaardi Budd (формация Буэн, нижний кембрий, Северная Гренландия)» . Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 89 (4): 249–290. дои : 10.1017/S0263593300002418 . ISSN 1473-7116 . S2CID 85645934 .
- ^ Перейти обратно: а б с д и ж Чжан, X.; Бриггс, DEG (2007). «Природа и значение придатков Опабинии из среднекембрийских сланцев Бёрджесс». Летайя . 40 (2): 161–173. дои : 10.1111/j.1502-3931.2007.00013.x .
- ^ Перейти обратно: а б с д и ж г час Бадд, GE; Дейли, AC (2011). «Доли и лобоподы Opabinia regalis из среднекембрийских сланцев Берджесс». Летайя . 45 : 83–95. дои : 10.1111/j.1502-3931.2011.00264.x .
- ^ Перейти обратно: а б Ортега-Эрнандес, Хавьер (2016). «Осмысление «нижней» и «верхней» стеблевой группы Euarthropoda с комментариями о строгом использовании названия Arthropoda von Siebold, 1848» . Биологические обзоры . 91 (1): 255–273. дои : 10.1111/brv.12168 . ПМИД 25528950 . S2CID 7751936 .
- ^ Перейти обратно: а б с Ортега-Эрнандес, Хавьер; Янссен, Ральф; Бадд, Грэм Э. (01 мая 2017 г.). «Происхождение и эволюция головы панартропод - палеобиологическая перспектива и перспектива развития» . Строение и развитие членистоногих . 46 (3): 354–379. дои : 10.1016/j.asd.2016.10.011 . ISSN 1467-8039 . ПМИД 27989966 .
- ^ Перейти обратно: а б Бадд, GE (2003). «Кембрийские окаменелости и происхождение типов» . Интегративная и сравнительная биология . 43 (1): 157–165. дои : 10.1093/icb/43.1.157 . ПМИД 21680420 .
- ^ Перейти обратно: а б с УОЛКОТТ, CD 1912. Среднекембрийские Branchiopoda, Malacostraca, Trilobita и Merostomata . Разные коллекции Смитсоновского института, 57: 145–228.
- ^ Гулд, С.Дж. (1990). Прекрасная жизнь . Лондон: Радиус Хатчинсона. п. 77 и с. 189. ИСБН 978-0-09-174271-3 .
- ^ Мирошников Л.Д. и Кразов А.Г. (1960). Редкие палеонтологические остатки и следы жизни в позднекембрийских отложениях северо-запада Сибирской платформы. Палеонтология и биостратиграфия Советской Арктики, 3, стр. 28–41.
- ^ Бриггс, Германия ; Недин, К. (январь 1997 г.). «Тафономия и сходство проблемного ископаемых миосколексов из нижнекембрийских сланцев залива Эму в Южной Австралии». Журнал палеонтологии . 71 (1): 22–32. Бибкод : 1997JPal...71...22B . дои : 10.1017/S0022336000038919 . JSTOR 1306537 . S2CID 131851540 .
- ^ Дзик, Дж. (2004). «Анатомия и взаимоотношения раннекембрийского червя Myoscolex ». Зоологика Скрипта . 33 (1): 57–69. дои : 10.1111/j.1463-6409.2004.00136.x . S2CID 85216629 .
- ^ Перейти обратно: а б с д и ж Бергстрем, Дж. (1986). « Опабиния и Аномалокарис , уникальные кембрийские членистоногие». Летайя . 19 (3): 241–246. дои : 10.1111/j.1502-3931.1986.tb00738.x .
- ^ Перейти обратно: а б Сянгуан, Хоу; Бергстрём, январь (2006). «Динокаридиды – аномальные членистоногие или членистоногие черви?». В Цзяюй, Ронг; Цзунцзе, Фан; Чжанхэ, Чжоу; Ренбин, Жан; Сяндун, Ван; Сюньлай, Юань (ред.). Происхождение, радиация и изменения биоразнообразия – свидетельства китайской летописи окаменелостей . стр. 139–158. CiteSeerX 10.1.1.693.5869 .
- ^ Чипман, Ариэль Д. (18 декабря 2015 г.). «Эмбриологический взгляд на летопись окаменелостей ранних членистоногих» . Эволюционная биология BMC . 15 (1): 285. дои : 10.1186/s12862-015-0566-z . ISSN 1471-2148 . ПМЦ 4683962 . ПМИД 26678148 .
- ^ Паулюс, Х.Ф. (2000). «Филогения многоножек-ракообразных-насекомых: новая попытка использования структуры фоторецепторов *» . Журнал зоологической систематики и эволюционных исследований . 38 (3): 189–208. дои : 10.1046/j.1439-0469.2000.383152.x .
- ^ Чен, JY.; Рамшельд, Л.; Гуй-цин Чжоу, GQ. (май 1994 г.). «Доказательства монофилии и близости кембрийских гигантских хищников к членистоногим». Наука . 264 (5163): 1304–1308. Бибкод : 1994Sci...264.1304C . дои : 10.1126/science.264.5163.1304 . ПМИД 17780848 . S2CID 1913482 .
- ^ Перейти обратно: а б Ваннье, Жан; Лю, Цзянни; Лерози-Обрил, Руди; Винтер, Якоб; Дэйли, Эллисон К. (2 мая 2014 г.). «Сложная пищеварительная система у ранних членистоногих» . Природные коммуникации . 5 (1): 3641. Бибкод : 2014NatCo...5.3641V . дои : 10.1038/ncomms4641 . ISSN 2041-1723 . ПМИД 24785191 . S2CID 205324774 .
- ^ Хатчинсон, Джордж Эвелин (1930). Повторное исследование некоторых окаменелостей сланцев Берджесс . Труды Национального музея США 78 (2854): 1–24. doi:10.5479/si.00963801.78-2854.1.
- ^ Симонетта AM. 1970 г. Исследования нетрилобитовых членистоногих сланцев Бёрджесс (средний кембрий). Палеонтогр. Итал. 66, 35–45.
- ^ Эджкомб, Грегори Д. (2009). «Палеонтологические и молекулярные данные, связывающие членистоногих, онихофоров и других экдизозоа» . Эволюция: образование и информационно-пропагандистская деятельность . 2 (2): 178–190. дои : 10.1007/s12052-009-0118-3 . ISSN 1936-6434 .
- ^ Уиттингтон, Гарри Блэкмор; Бриггс, Дерек Эрнест Гилмор (14 мая 1985 г.). «Самое крупное кембрийское животное, Anomalocaris, Берджесс-Шейл, Британская Колумбия» . Философские труды Лондонского королевского общества. Б. Биологические науки . 309 (1141): 569–609. Бибкод : 1985RSPTB.309..569W . дои : 10.1098/rstb.1985.0096 .
- ^ Перейти обратно: а б Бадд, GE (1993). «Кембрийский жаберный лобопод из Гренландии». Природа . 364 (6439): 709–711. Бибкод : 1993Natur.364..709B . дои : 10.1038/364709a0 . S2CID 4341971 .
- ^ Бадд, GE (1997). «Членистоногие стволовой группы из фауны Сириуса Пассета нижнего кембрия Северной Гренландии». в Форти, РА ; Томас, Р.Х. (ред.). Взаимоотношения членистоногих – Специальная серия томов 55 . Ассоциация систематиков.
- ^ Сиань-Гуан, Хоу; Бергстрем, Ян; Альберг, Пер (1 сентября 1995 г.). «Аномалокарис и другие крупные животные фауны Чэнцзян нижнего кембрия на юго-западе Китая» . ГФФ . 117 (3): 163–183. дои : 10.1080/11035899509546213 . ISSN 1103-5897 .
- ^ Коллинз, Десмонд (1996). «Эволюция» Anomalocaris и его отнесение к классу членистоногих Dinocarida (nov.) и отряду Radiodonta (nov.)» . Журнал палеонтологии . 70 (2): 280–293. Бибкод : 1996JPal...70..280C . дои : 10.1017/S0022336000023362 . ISSN 0022-3360 . S2CID 131622496 .
- ^ Легг, Дэвид А.; Саттон, Марк Д.; Эджкомб, Грегори Д.; Карон, Жан-Бернар (7 декабря 2012 г.). «Кембрийские двустворчатые членистоногие раскрывают происхождение артродизации» . Труды Королевского общества B: Биологические науки . 279 (1748): 4699–4704. дои : 10.1098/рспб.2012.1958 . ПМЦ 3497099 . ПМИД 23055069 .
- ^ Легг, Дэвид (2013). «Многочлениковые членистоногие из среднего кембрия Британской Колумбии (Канада)» . Журнал палеонтологии . 87 (3): 493–501. Бибкод : 2013JPal...87..493L . дои : 10.1666/12-112.1 . ISSN 0022-3360 . S2CID 86725173 .
- ^ Легг, Дэвид А.; Ваннье, Жан (2013). «Сродство космополитических членистоногих Isoxys и его значение для происхождения членистоногих» . Летайя . 46 (4): 540–550. дои : 10.1111/лет.12032 . ISSN 1502-3931 .
- ^ Легг, Дэвид А.; Саттон, Марк Д.; Эджкомб, Грегори Д. (30 сентября 2013 г.). «Данные об окаменелостях членистоногих повышают соответствие морфологической и молекулярной филогении» . Природные коммуникации . 4 (1): 2485. Бибкод : 2013NatCo...4.2485L . дои : 10.1038/ncomms3485 . ISSN 2041-1723 . ПМИД 24077329 .
- ^ Смит, Мартин Р.; Ортега-Эрнандес, Хавьер (2014). «Онихофороподобные когти Hallucigenia и случай Tactopoda» . Природа . 514 (7522): 363–366. Бибкод : 2014Natur.514..363S . дои : 10.1038/nature13576 . ISSN 1476-4687 . ПМИД 25132546 . S2CID 205239797 .
- ^ Ван Рой, Питер; Дейли, Эллисон С.; Бриггс, Дерек Э.Г. (2015). «Гомология конечностей туловища аномалокаридид, выявленная гигантским фильтратором с парными створками» . Природа . 522 (7554): 77–80. Бибкод : 2015Natur.522...77V . дои : 10.1038/nature14256 . ISSN 1476-4687 . ПМИД 25762145 . S2CID 205242881 .
- ^ Смит, Мартин Р.; Карон, Жан-Бернар (2015). «Голова галлюцигении и глоточный каркас ранних экдизозоев» . Природа . 523 (7558): 75–78. Бибкод : 2015Natur.523...75S . дои : 10.1038/nature14573 . ISSN 1476-4687 . ПМИД 26106857 . S2CID 205244325 .
- ^ Карон, Жан-Бернар; Ария, Седрик (31 января 2017 г.). «Кембрийские лобоподии, питающиеся взвесью, и ранняя радиация панартропод» . Эволюционная биология BMC . 17 (1): 29. дои : 10.1186/s12862-016-0858-y . ISSN 1471-2148 . ПМЦ 5282736 . ПМИД 28137244 .
- ^ Сиветер, Дерек Дж.; Бриггс, Дерек Э.Г.; Сиветер, Дэвид Дж.; Саттон, Марк Д.; Легг, Дэвид (2018). «Трехмерно сохранившийся лобопод из Херефордширского (силурийского) Лагерштетте, Великобритания» . Королевское общество открытой науки . 5 (8): 172101. дои : 10.1098/rsos.172101 . ПМК 6124121 . ПМИД 30224988 .
- ^ Перейти обратно: а б Цзэн, Хан; Чжао, Фанчэн; Ню, Кеченг; Чжу, Маоянь; Хуан, Диин (2020). «Раннекембрийский эуартропод с хищными придатками, похожими на радиодонтов» . Природа . 588 (7836): 101–105. Бибкод : 2020Natur.588..101Z . дои : 10.1038/s41586-020-2883-7 . ISSN 1476-4687 . ПМИД 33149303 . S2CID 226248177 .
- ^ Андерсон, Эван П.; Шиффбауэр, Джеймс Д.; Жаке, Сара М.; Ламсделл, Джеймс К.; Клюссендорф, Джоан; Микулич, Дональд Г. (2021). «Незнакомец, чем скорпион: переоценка Parioscorpio venator , проблемного членистоногого из лландоверского Waukesha Lagerstätte» . Палеонтология . 64 (3): 429–474. Бибкод : 2021Palgy..64..429A . дои : 10.1111/пала.12534 . ISSN 1475-4983 . S2CID 234812878 .
- ^ Мойсюк, Иосиф; Карон, Жан-Бернар (2020). «Исключительная многофункциональность в питающем аппарате среднекембрийского радиодонта» . Палеобиология . 47 (4): 704–724. Бибкод : 2021Pbio...47..704M . дои : 10.1017/pab.2021.19 . ISSN 0094-8373 . S2CID 236552819 .
- ^ Перейти обратно: а б с Пейтс, Стивен; Вулф, Джоанна М.; Лерози-Обрил, Руди; Дейли, Эллисон С.; Ортега-Эрнандес, Хавьер (9 февраля 2022 г.). «Новый опабиниид разнообразит самые странные чудеса стволовой группы эуартропод» . Труды Королевского общества B: Биологические науки . 289 (1968): 20212093. doi : 10.1098/rspb.2021.2093 . ПМЦ 8826304 . ПМИД 35135344 .
- ^ Бриггс, Дерек Э.Г.; Либерман, Брюс С.; Хендрикс, Джонатан Р.; Хальгедал, Сьюзен Л.; Джаррард, Ричард Д. (2008). «Среднекембрийские членистоногие из Юты» . Журнал палеонтологии . 82 (2): 238–254. Бибкод : 2008JPal...82..238B . дои : 10.1666/06-086.1 . ISSN 0022-3360 . S2CID 31568651 .
- ^ Клауд, ЧП (1948). «Некоторые проблемы и закономерности эволюции на примере ископаемых беспозвоночных». Эволюция . 2 (4): 322–350. дои : 10.2307/2405523 . JSTOR 2405523 . ПМИД 18122310 . и Клауд, ЧП (1968). «Дометазоа эволюция и происхождение многоклеточных». В Дрейке, ET (ред.). Эволюция и окружающая среда . Нью-Хейвен, Коннектикут: Издательство Йельского университета. стр. 1–72.
- ^ Элдридж, Н.; Гулд, С.Дж. (1989). «ПРИЛОЖЕНИЕ: Прерывистые равновесия: альтернатива филетическому градуализму». Временные рамки: эволюция прерывистых равновесий . Элдридж, Н., стр. 193–224. дои : 10.1515/9781400860296.193 . ISBN 9781400860296 .
- ^ Бенгтсон, Стефан (2004). «Ранние скелетные окаменелости» . В Липпсе, Джере Х .; Ваггонер, Бенджамин М. (ред.). Неопротерозойско-кембрийские биологические революции: представлено в виде краткого курса палеонтологического общества на ежегодном собрании Геологического общества Америки, Денвер, Колорадо, 6 ноября 2004 г. Документы Палеонтологического общества. Том. 10. Нью-Хейвен, Коннектикут: Служба репрографии и визуализации Йельского университета; Палеонтологическое общество . ISSN 1089-3326 . OCLC 57481790 . Проверено 6 февраля 2015 г.
- ^ Дарем, JW (1971). «Летопись окаменелостей и происхождение вторичноротых». Труды Североамериканской палеонтологической конвенции, Часть H : 1104–1132. и Глесснер, МФ (1972). «Докембрийская палеозоология». В Джонсе, Дж. Б.; Макгоуран, Б. (ред.). Стратиграфические проблемы позднего докембрия и раннего кембрия . Том. 1. Университет Аделаиды. стр. 43–52.
- ^ Нолл, АХ (2004). «Кембрийский редукс» . Первые три миллиарда лет эволюции на Земле . Издательство Принстонского университета. п. 192. ИСБН 978-0-691-12029-4 . Проверено 22 апреля 2009 г.
- ^ Робисон, РА (январь 1985 г.). «Сродство Aysheaia (Onychophora) с описанием нового кембрийского вида». Журнал палеонтологии . 59 (1): 226–235. JSTOR 1304837 .
- ^ Джейкобс, ДК; Рэй, К.Г.; Ведин, CJ; Кострикен, Р.; ДеСалле, Р.; Стейтон, Дж.Л.; Гейтс, РД; Линдберг, Д.Р. (2000). «Экспрессия моллюсков, серийная организация и эволюция раковины». Эволюция и развитие . 2 (6): 340–347. дои : 10.1046/j.1525-142x.2000.00077.x . ПМИД 11256378 . S2CID 25274057 .
- ^ Перейти обратно: а б Краск, Эй Джей; Джеффрис, RPS (1989). «Новый митрат из верхнего ордовика Норвегии и новый подход к подразделению плесиона». Палеонтология . 32 : 69–99.
Дальнейшее чтение
[ редактировать ]- Бергстрем, Дж. (1987). «Кембрийские опабинии и аномалокарисы ». Летайя . 20 (2): 187–188. дои : 10.1111/j.1502-3931.1987.tb02037.x .
- Бриггс, DEG; Уиттингтон, HB (1987). «Сродство кембрийских животных Anomalocaris и Opabinia ». Летайя . 20 (2): 185–186. дои : 10.1111/j.1502-3931.1987.tb02036.x .
Внешние ссылки
[ редактировать ]- « Опабиния королевская » . Галерея окаменелостей сланцев Бёрджесс . Виртуальный музей Канады. 2011. Архивировано из оригинала 12 ноября 2020 г.
- Страница Смитсоновского института об Опабинии с фотографией ископаемого сланца Бёрджесс.
