Скучный миллиард
−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||||||||||||||||||||
« Скучный миллиард» , также известный как средний протерозой и земное средневековье , представляет собой неформальный геологический период времени между 1,8 и 0,8 миллиарда лет назад ( млрд лет назад ) в течение среднего протерозоя периоды , охватывающий от статерийского до тонийского периода , характеризующийся более или менее тектоническая стабильность, климатический застой и медленная биологическая эволюция . Хотя он граничит с двумя различными событиями насыщения кислородом ( Великим событием окисления и Неопротерозойским событием окисления ) и двумя глобальными ледниковыми событиями ( гуронским и криогенным оледенениями ), в самом периоде Бурного миллиарда на самом деле был очень низкий уровень кислорода и не было никаких геологических свидетельств оледенений.
Океаны во времена «Скучного миллиарда» могли быть бедными кислородом, бедными питательными веществами и сульфидными ( эуксиния ), населенными в основном аноксигенными пурпурными бактериями , типом фотосинтезирующих бактерий на основе бактериохлорофилла используют сероводород (H 2 S) для фиксации углерода. , которые вместо этого воды кислорода производит серу в качестве побочного продукта вместо и . Это известно как океан Кэнфилда , и такой состав, возможно, привел к тому, что океаны стали окрашены в черный и молочно -бирюзовый цвет , а не в синий или зеленый, как позже. (Напротив, во время гораздо более ранней фазы Пурпурной Земли в архее фотосинтез осуществлялся в основном колониями архей с использованием сетчатке глаза на протонных насосов , которые поглощают зеленый свет, и океаны были пурпурно - фиолетовыми .)
Несмотря на такие неблагоприятные условия, эукариоты , возможно, эволюционировали примерно в начале «Скучного миллиарда» и приняли несколько новых адаптаций, таких как различные органеллы , многоклеточность и, возможно, половое размножение , а диверсифицировались в водоросли , грибы и ранних животных в конце этого временного интервала . . [1] Такие достижения, возможно, были важными предшественниками эволюции крупной и сложной жизни позже во время Эдиакарского Авалонского взрыва и последующего фанерозойского кембрийского взрыва . Тем не менее, прокариотические цианобактерии были доминирующими автотрофными формами жизни в это время и, вероятно, поддерживали энергетически бедную пищевую сеть с небольшим количеством протистов на вершинном уровне . На суше, вероятно, обитали прокариотические цианобактерии и эукариотические протолишайники , причем последние здесь более успешны, вероятно, из-за большей доступности питательных веществ, чем в прибрежных водах океана.
Описание
[ редактировать ]В 1995 году геологи Роджер Бьюик, Дэвис Де Марэ и Эндрю Нолл рассмотрели очевидное отсутствие крупных биологических, геологических и климатических событий в течение мезопротерозойской эры от 1,6 до 1 миллиарда лет назад (Ga) и, таким образом, описали ее как « самое скучное время в истории Земли». [2] Термин «скучный миллиард» был придуман палеонтологом Мартином Бразье для обозначения периода между 2 и 1 млрд лет назад, который характеризовался геохимическим застоем и застоем ледников. [3] В 2013 году геохимик Грант Янг использовал термин «Бесплодный миллиард» для обозначения периода очевидного ледникового застоя и отсутствия отклонений изотопов углерода от 1,8 до 0,8 млрд лет назад. [4] В 2014 году геологи Питер Кавуд и Крис Хоксворт назвали время между 1,7 и 0,75 млрд лет «Средневековьем Земли» из-за отсутствия доказательств тектонического движения . [5]
В настоящее время считается, что «Скучный миллиард» охватывает период от 1,8 до 0,8 млрд лет и содержится в протерозойском эоне , в основном в мезопротерозое. «Скучный миллиард» характеризуется геологическим, климатическим и вообще эволюционным застоем с низким содержанием питательных веществ. [6] [7] [8]
Во время, предшествовавшее «Скучному миллиарду», Земля пережила Великое событие оксигенации из-за эволюции кислородных фотосинтезирующих цианобактерий и возникшего в результате гуронского оледенения ( Земля-снежок ), образования УФ-излучение , блокирующего озонового слоя , и окисления нескольких металлов. [9] Считается, что уровень кислорода во время «Скучного миллиарда» был заметно ниже, чем во время Великого события окисления, возможно, от 0,1% до 10% от современного уровня. [10] Это закончилось распадом суперконтинента Родиния в тонианском периоде (1000–720 млн лет назад), вторым событием насыщения кислородом и еще одной Землей-снежком в криогенный период. [5] [11]
Тектонический застой
[ редактировать ]
Эволюция биосферы , атмосферы и гидросферы Земли уже давно связана с циклом суперконтинентов , когда континенты объединяются, а затем расходятся. «Скучный миллиард» стал свидетелем эволюции двух суперконтинентов: Колумбии (или Нуны) и Родинии . [8] [12]
Суперконтинент Колумбия образовался между 2,0 и 1,7 млрд лет назад и оставался нетронутым по крайней мере до 1,3 млрд лет назад. Геологические и палеомагнитные данные позволяют предположить, что Колумбия претерпела лишь незначительные изменения, чтобы сформировать суперконтинент Родиния в период с 1,1 по 0,9 млрд лет назад. Палеогеографические реконструкции предполагают, что комплекс суперконтинентов располагался в экваториальные и умеренные мало или вообще нет климатические зоны, а свидетельств существования фрагментов континентов в полярных регионах . [12]
Из-за отсутствия свидетельств накопления отложений (на пассивных окраинах), которое могло бы произойти в результате рифтогенеза , [13] Суперконтинент, вероятно, не распался, а скорее представлял собой просто совокупность расположенных рядом протоконтинентов и кратонов . Нет никаких свидетельств рифтогенеза до образования Родинии, 1,25 млрд лет в Северной Лаврентии и 1 млрд лет в Восточной Балтике и Южной Сибири . [8] Распад произошел только 0,75 млрд лет назад, что ознаменовало конец «Скучного миллиарда». [5] Этот тектонический застой мог быть связан с химией океана и атмосферы. [8] [6]
Вполне возможно, что астеносфера Земли — расплавленный слой мантии , по которому, по существу, плавают и движутся тектонические плиты — была слишком горячей, чтобы поддерживать современную тектонику плит в то время. Вместо энергичной переработки плит в зонах субдукции , плиты были связаны друг с другом на протяжении миллиардов лет, пока мантия достаточно не остыла. Возникновению этого компонента тектоники плит, возможно, способствовало охлаждение и утолщение земной коры , которое, однажды начавшись, сделало субдукцию плит аномально сильной, произошедшую в конце «Скучного миллиарда». [5]
Тем не менее, крупные магматические события все же произошли, такие как образование (посредством магматического шлейфа ) 220 000-километрового вулкана. 2 (85 000 квадратных миль) центральной австралийской провинции Масгрейв от 1,22 до 1,12 млрд лет назад, [14] и 2 700 000 км. 2 (1 000 000 квадратных миль) Канадская крупная магматическая провинция Маккензи, 1,27 млрд лет назад. [15] Тектоника плит все еще была достаточно активной, чтобы образовать горы с несколькими складками , включая Гренвиллскую складчатость , [16] происходящее в то время.
Климатическая стабильность
[ редактировать ]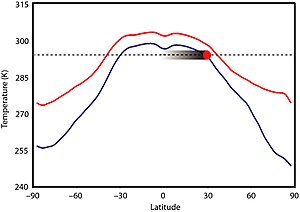
Существует мало свидетельств значительной изменчивости климата в этот период времени. [4] [17] Климат, вероятно, в первую очередь не диктовался солнечной яркостью, потому что Солнце было на 5–18% менее ярким, чем сегодня, но нет никаких доказательств того, что климат Земли был значительно прохладнее. [18] [19] Фактически, в «Скучном миллиарде», похоже, отсутствуют какие-либо свидетельства продолжительных оледенений, которые можно наблюдать с регулярной периодичностью в других частях геологической истории Земли. [19] Высокий уровень CO 2 не мог быть основным фактором потепления, поскольку его уровни должны были быть в 30–100 раз выше, чем доиндустриальные уровни . [18] и вызвало значительное закисление океана [19] для предотвращения образования льда, которого также не произошло. Уровни CO 2 в мезопротерозое могли быть сопоставимы с уровнями фанерозоя , возможно, в 7–10 раз выше современных уровней. [20] Первая запись льда за этот период времени была зарегистрирована в 2020 году в шотландской формации Диабайг, образовавшейся 1 млрд лет назад, в группе Торридон , где образования дропстоуна , вероятно, образовались из обломков ледяного наплыва ; эта территория, которая тогда располагалась между 35–50 ° ю.ш., представляла собой озеро (возможно , возвышенное), которое, как полагают, замерзало зимой и таяли летом, а во время весеннего таяния происходил сплав. [21]
Более высокое содержание других парниковых газов, а именно метана, вырабатываемого прокариотами, возможно, компенсировало низкие уровни CO 2 ; мир практически свободен ото льда, что достигается за счет концентрации метана в атмосфере 140 частей на миллион (ppm). [20] [18] Метаногенные прокариоты не могли производить столько метана, а это означает, что содержание какого-то другого парникового газа, вероятно, закиси азота , было повышено, возможно, до 3 частей на миллион (в 10 раз выше сегодняшнего уровня). Судя по предполагаемым концентрациям парниковых газов, экваториальные температуры в мезопротерозое могли составлять около 295–300 К (22–27 ° C; 71–80 ° F), в тропиках 290 К (17 ° C; 62 ° F), при 60 ° 265–280 К (-8–7 ° C; 17–44 ° F) и полюса 250–275 К (-23–2 ° C; -10–35 ° F); [22] а средняя глобальная температура около 19 ° C (66 ° F), что на 4 ° C (7,2 ° F) теплее, чем сегодня. Зимой температура на полюсах опускалась ниже нуля, что позволяло временно образовывать морской лед и выпадать снегопады, но постоянных ледниковых щитов, скорее всего, не было. [7]
Также было высказано предположение, что, поскольку было показано, что интенсивность космических лучей положительно коррелирует с облачным покровом, а облачный покров отражает свет в космос и снижает глобальные температуры, более низкие темпы бомбардировки в это время из-за замедления звездообразования в Галактика вызвала уменьшение облачности и предотвратила оледенение, поддерживая теплый климат. [19] [23] Кроме того, некоторая комбинация интенсивности выветривания, которая привела бы к снижению уровня CO 2 за счет окисления открытых металлов, охлаждения мантии и уменьшения геотермального тепла и вулканизма, а также увеличения солнечной интенсивности и солнечного тепла, могла достичь равновесия, исключая образование льда. [4]
И наоборот, движение ледников более миллиарда лет назад, возможно, не оставило сегодня много остатков, и очевидное отсутствие доказательств может быть связано с неполнотой летописи окаменелостей, а не с ее отсутствием. Кроме того, низкий уровень кислорода и интенсивность солнечной энергии могли препятствовать образованию озонового слоя , предотвращая парниковых газов попадание в атмосферу и нагревание Земли за счет парникового эффекта , что могло бы вызвать оледенение. [24] [25] [26] Хотя для поддержания озонового слоя необходимо не так много кислорода, и его уровни во время «Скучного миллиарда» могли быть достаточно высокими для этого, [27] Земля, возможно, подвергалась более сильной бомбардировке УФ-излучением , чем сегодня. [28]
Океанический состав
[ редактировать ]Судя по всему, в океанах были низкие концентрации ключевых питательных веществ, которые считались необходимыми для сложной жизни, а именно молибдена , железа, азота и фосфора , в значительной степени из-за нехватки кислорода и, как следствие, окисления, необходимого для этих геохимических циклов . [29] [30] [31] Питательных веществ могло быть больше в наземной среде, например, в озерах или прибрежных районах, расположенных ближе к континентальному стоку. [32]
В целом океаны могли иметь насыщенный кислородом поверхностный слой, сульфидный средний слой, [33] [34] [35] и субоксический нижний слой. [36] [37] Преимущественно сульфидный состав, возможно, привел к тому, что океаны приобрели черный и молочно-бирюзовый цвет вместо синего. [38]
Кислород
[ редактировать ]Геологические данные Земли указывают на два события, связанные со значительным увеличением уровня кислорода на Земле: одно произошло между 2,4 и 2,1 млрд лет назад, известное как Великое событие окисления (GOE), а второе произошло примерно 0,8 млрд лет назад, известное как неопротерозойское событие окисления. (НОЭ). [39] Считается, что в промежуточный период, во время «Скучного миллиарда», уровень кислорода был низким (с небольшими колебаниями), что привело к широкому распространению бескислородных вод . [34]
Океаны могли иметь отчетливую стратификацию, а поверхностные воды были насыщены кислородом. [33] [34] [35] а глубокая вода является субкислородной (менее 1 мкМ кислорода), [37] последнее, возможно, поддерживается за счет более низких уровней выхода водорода (H 2 ) и H 2 S из глубоководных гидротермальных источников , которые в противном случае были бы химически восстановлены кислородом, т.е. эвксиновыми водами . [36] Даже на самом мелководье значительные количества кислорода могли быть ограничены главным образом участками вблизи побережья. [40] Разложение . тонущего органического вещества также привело бы к выщелачиванию кислорода из глубоких вод [41] [34]
Внезапное падение содержания O 2 после Великого события оксигенации, о котором свидетельствуют уровни δ13C , которое представляло собой потерю в 10–20 раз большего объема атмосферного кислорода, известно как событие Ломагунди-Джатули и является наиболее заметным связанным с изотопами углерода событием, . в истории Земли. [42] [43] [44] Уровень кислорода мог составлять менее 0,1–1% от современного уровня. [45] что фактически остановило бы эволюцию сложной жизни во времена «Скучного миллиарда». [39] [35] Однако предполагается, что мезопротерозойское событие оксигенации (MOE), во время которого содержание кислорода временно повышалось примерно до 4% PAL в разные моменты времени, произошло в период с 1,59 по 1,36 млрд лет назад. [46] В частности, некоторые данные из формации Гаоючжуан предполагают повышение содержания кислорода около 1,57 млрд лет назад. [47] Велкерри в группе Ропер Северной территории Австралии в то время как формация , [48] the Kaltasy Formation ( Russian : Калтасинская свита ) of Volgo-Uralia , Russia , [40] и формация Сямалин на севере Северо-Китайского кратона. [49] [50] указывают на заметную оксигенацию около 1,4 млрд лет назад, хотя степень, в которой это отражает глобальный уровень кислорода, неясна. [48] Кислородные условия стали бы преобладать в НЭ, вызывая распространение аэробной активности над анаэробной . [33] [34] [41] но широко распространенные субоксические и бескислородные условия, вероятно, продолжались примерно до 0,55 млрд лет назад, что соответствовало эдиакарской биоте и кембрийскому взрыву . [51] [52]
сера
[ редактировать ]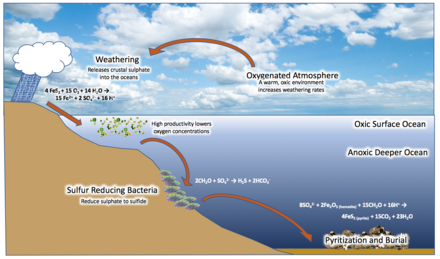
В 1998 году геолог Дональд Кэнфилд предложил то, что сейчас известно как гипотеза океана Кэнфилда . [33] Кэнфилд утверждал, что повышение уровня кислорода в атмосфере во время Великого оксигенационного события могло бы вступить в реакцию с отложениями континентального железного пирита (FeS 2 ) и окислить их с сульфатом (SO 4 2− ) как побочный продукт, который был вынесен в море. [53] Сульфатредуцирующие микроорганизмы превратили его в сероводород (H 2 S), разделив океан на несколько кислородный поверхностный слой и сульфидный слой под ним, с аноксигенными бактериями, живущими на границе, метаболизирующими H 2 S и производящими серу в качестве отходов. продукт. Это создало широко распространенные эвксинические условия в средних водах, бескислородное состояние с высокой концентрацией серы, которое поддерживалось бактериями. [54] [41] [35] Многие месторождения Бурового миллиарда содержат изотопные соотношения ртути, характерные для эвксиний фотической зоны. [55] Более систематические геохимические исследования среднего протерозоя показывают, что океаны были в основном железистыми с тонким поверхностным слоем слабокислородных вод. [56] а эвксиния могла встречаться на относительно небольших участках, возможно, менее 7% морского дна. [57] Очень низкие концентрации молибдена в мезопротерозое можно было бы в достаточной мере объяснить даже при таком относительно низком проценте эвксинного содержания морского дна. [34] Эвксиния расширялась и сжималась, иногда достигая фотической зоны, а иногда опускаясь в более глубокие воды. [58] Данные из бассейна Макартур на севере Австралии показывают, что внутриконтинентальные условия, в частности, периодически имели низкое содержание сульфидов. [59]
Железо
[ редактировать ]Среди пород, относящихся к «Скучному миллиарду», заметно отсутствие полосчатых железных образований , которые образуются из железа в верхних слоях воды (поступающего из глубокого океана), реагирующего с кислородом и выпадающего в осадок из воды. По-видимому, они прекратились по всему миру после 1,85 млрд лет назад. Кэнфилд утверждал, что океанические SO 2- 4 восстановил все железо в бескислородных морских глубинах. [33] Железо могло метаболизироваться аноксигенными бактериями. [60] Также было высказано предположение, что удар метеорита Садбери 1,85 млрд лет назад смешал ранее стратифицированный океан посредством цунами, взаимодействия между испаренной морской водой и насыщенной кислородом атмосферой, океанической кавитации и массивного стока разрушенных континентальных окраин в море. Образовавшиеся в результате субкислородные глубокие воды (из-за смешивания насыщенных кислородом поверхностных вод с ранее бескислородными глубокими водами) окислили бы глубоководное железо, предотвращая его транспортировку и отложение на окраинах континента. [36]
Тем не менее, богатые железом воды все же существовали, такие как формация Сямалин возрастом 1,4 млрд лет в Северном Китае, которая, возможно, питалась глубоководными гидротермальными жерлами. Богатые железом условия также указывают на бескислородную придонную воду в этой области, поскольку кислородные условия окислили бы все железо. [60]
Формы жизни
[ редактировать ]Низкое содержание питательных веществ могло способствовать фотосимбиозу (когда один организм способен к фотосинтезу, а другой метаболизирует продукты жизнедеятельности) среди прокариот ( бактерий и архей ), а также появлению эукариотов . Бактерии, археи и эукариоты — три домена , имеющие высший таксономический рейтинг. Эукариоты отличаются от прокариотов ядром и мембраносвязанными органеллами, и почти все многоклеточные организмы являются эукариотами. [61]
Прокариоты
[ редактировать ]
Прокариоты были доминирующей формой жизни на протяжении всего «Скучного миллиарда». [9] [62] [33] Микрофоссилии указывают на присутствие цианобактерий, зеленых и пурпурных серных бактерий, архей, производящих метан, бактерий, метаболизирующих сульфат, архей или бактерий, метаболизирующих метан , бактерий, метаболизирующих железо, бактерий, метаболизирующих азот , и аноксигенных фотосинтезирующих бактерий. [63]
Считается, что аноксигенные цианобактерии были доминирующими фотосинтезаторами, метаболизируя обильный H 2 S в океанах. В водах, богатых железом, цианобактерии могли страдать от отравления железом , особенно в морских водах, где богатые железом глубокие воды смешивались с поверхностными водами и, таким образом, были вытеснены другими бактериями, которые могли метаболизировать как железо, так и H 2 S. Однако отравление железом можно было бы уменьшить с помощью кремнеземом воды, богатой , или биоминерализации железа внутри клетки. [63]
Одноклеточные планктонные линии цианобактерий развивались в пресной воде в середине мезопротерозоя , а в неопротерозое как бентосные морские, так и некоторые пресноводные предки дали начало морским планктонным цианобактериям (как азотфиксирующим, так и неазотфиксирующим), способствуя оксигенации Докембрийские океаны. [64] [65]
Исследования цианобактерий в лаборатории показали, что фермент нитрогеназа, который используется для фиксации атмосферного азота, перестает работать, когда уровень кислорода превышает 10% от текущего уровня в атмосфере. Отсутствие азота из-за повышенного количества кислорода создало бы петлю отрицательной обратной связи , при которой уровень кислорода в атмосфере стабилизировался на уровне 2%, который начал меняться около 600 миллионов лет назад, когда наземные растения начали выделять кислород. К 408 миллионам лет назад у азотфиксирующих цианобактерий развились гетероцисты, защищающие свою нитрогеназу от кислорода. [66] [67]
Эукариоты
[ редактировать ]Эукариоты, возможно, возникли примерно в начале «Скучного миллиарда». [1] совпадающее с аккрецией Колумбии, что могло каким-то образом повысить уровень кислорода в океане. [11] Хотя были заявлены о существовании эукариот еще 2,1 миллиарда лет назад, они были сочтены сомнительными, поскольку самые старые однозначные останки эукариот датируются примерно 1,8-1,6 миллиарда лет назад в Китае. [68] После этого эволюция эукариот шла довольно медленно. [9] возможно, из-за эвксиновых условий океана Кэнфилд и нехватки ключевых питательных веществ и металлов. [5] [1] что препятствовало развитию большой, сложной жизни с высокими энергетическими потребностями. [24] Эвксиновые условия также снизили бы растворимость железа. [33] и молибден , [69] незаменимые металлы в фиксации азота . Отсутствие растворенного азота дало бы преимущество прокариотам перед эукариотами, поскольку первые могут метаболизировать газообразный азот. [70] Альтернативная гипотеза об отсутствии разнообразия среди эукариот предполагает высокие температуры во время «Скучного миллиарда», а не низкие уровни кислорода, утверждая, что тот факт, что события оксигенации до позднего неопротерозоя не дали толчок эволюции эукариот, предполагает, что это не было основным ограничивающим фактором, тормозящим ее. . [71]

Тем не менее, диверсификация эукариотических макроорганизмов кроновой группы, по-видимому, началась примерно 1,6–1 млрд лет назад, что, по-видимому, совпало с увеличением концентраций ключевых питательных веществ. [1] По данным молекулярно-часового анализа, растения отделились от животных и грибов около 1,6 млрд лет назад; животные и грибы около 1,5 млрд лет; Губки от других животных разошлись примерно в 1,35 млрд лет назад; [73] Билатерии и книдарии (животные соответственно с двусторонней симметрией и без нее ) около 1,3 млрд лет назад; и Ascomycota и Basidiomycota (два подразделения грибов подцарства Dikarya ) 0,97 млрд лет назад. [73] Авторы статьи заявляют, что их оценки времени не соответствуют научному консенсусу.
Окаменелости позднего палеопротерозоя и раннего мезопротерозоя Виндхийского осадочного бассейна Индии, [74] группа Руян Северного Китая, [75] [76] [77] и котуйканская свита Анабарского щита Сибири, [78] среди прочего указывают на высокие темпы (по доэдиакарским меркам) эукариотической диверсификации между 1,7 и 1,4 млрд лет назад, [79] хотя большая часть этого разнообразия представлена ранее неизвестными, уже не существующими кладами эукариот. [78] Самые ранние известные маты из красных водорослей датируются 1,6 млрд лет назад. [72] Самый ранний известный гриб датируется 1,01–0,89 млрд лет назад из Северной Канады. [80] Многоклеточные эукариоты, которые считаются потомками колониальных одноклеточных агрегатов, вероятно, возникли примерно 2–1,4 млрд лет назад. [81] [82] Точно так же ранние многоклеточные эукариоты, вероятно, в основном агрегировали в строматолитовые маты. [11]
Красная водоросль Bangiomorpha — самая ранняя известная форма жизни, размножающаяся половым путем и мейотическая . [83] и развился к 1,047 млрд лет назад. [84] Исходя из этого, эти адаптации развивались между ок. 2–1,4 млрд лет. [1] С другой стороны, они могли возникнуть задолго до появления последнего общего предка эукариот, учитывая, что мейоз осуществляется с использованием одних и тех же белков у всех эукариот, возможно, начиная с гипотетического мира РНК . [85]
клетки Органеллы , вероятно, произошли от свободноживущих цианобактерий ( симбиогенез ). [86] [87] [1] возможно, после развития фагоцитоза (поглощения других клеток) с удалением жесткой клеточной стенки , которая была необходима только для бесполого размножения. [9] Митохондрии уже развились во время Великого события оксигенации, но считается, что пластиды, используемые в приморастениях для фотосинтеза, появились примерно 1,6–1,5 млрд лет назад. [73] Гистоны , вероятно, появились во время «Скучного миллиарда», чтобы помочь организовать и упаковать растущее количество ДНК в эукариотических клетках в нуклеосомы . [9] Гидрогеносомы, используемые в анаэробной деятельности, возможно, произошли в то время от археев. [88] [86]
Учитывая эволюционные ориентиры, достигнутые эукариотами, этот период времени можно считать важным предшественником кембрийского взрыва около 0,54 млрд лет назад и эволюции относительно крупной и сложной жизни. [9]
Экология
[ редактировать ]Из-за маргинализации крупных пищевых частиц, таких как водоросли, в пользу цианобактерий и прокариот, которые не передают столько энергии на более высокие трофические уровни , сложная пищевая сеть, вероятно, не сформировалась, и крупные формы жизни с высокими потребностями в энергии не могли развиваться. . Такая пищевая сеть, вероятно, поддерживала лишь небольшое количество простейших , которые в некотором смысле были высшими хищниками . [62]
Предположительно кислородные фотосинтезирующие эукариотические акритархи , возможно, разновидность микроводорослей , населяли поверхностные воды мезопротерозоя. [89] Их популяция, возможно, была в значительной степени ограничена наличием питательных веществ, а не хищничеством, поскольку, как сообщается, виды выживали в течение сотен миллионов лет, но после 1 млрд лет продолжительность существования вида упала примерно до 100 млн лет назад, возможно, из-за увеличения травоядности ранних простейших. Это согласуется с падением выживаемости видов до 10 млн лет назад сразу после кембрийского взрыва и распространения травоядных животных. [90]
Относительно низкие концентрации молибдена в океане по всему «Скучному миллиарду» были предложены в качестве основного ограничивающего фактора, который удерживал на низком уровне популяции азотфиксирующих микроорганизмов открытого океана, которым требуется молибден для производства нитрогеназ , хотя пресноводные и прибрежные среды близки к речным источникам молибдена. растворенный молибден, возможно, все еще содержал значительные сообщества азотфиксаторов. Низкая скорость фиксации азота, которая закончилась только в криогенный период с появлением планктонных азотфиксаторов, означала, что свободный аммоний был в дефиците в этот временной интервал, что серьезно ограничивало эволюцию и разнообразие многоклеточной биоты. [91]
Жизнь на суше
[ редактировать ]Некоторые из самых ранних свидетельств прокариотической колонизации суши датируются периодом до 3 млрд лет назад. [92] возможно, уже 3,5 млрд лет назад. [93] Во времена «Скучного миллиарда» земля, возможно, была населена в основном цинобактериями. [94] [95] [96] [97] Пыль могла бы обеспечить обилие питательных веществ и средство распространения обитающих на поверхности микробов, хотя микробные сообщества могли также формироваться в пещерах, пресноводных озерах и реках. [28] [98] К 1,2 млрд лет назад микробные сообщества могли стать достаточно многочисленными, чтобы повлиять на выветривание, эрозию , седиментацию и различные геохимические циклы. [95] а обширные микробные маты могут указывать на биологической почвенной корки . обилие [28]
Самыми ранними наземными эукариотами, возможно, были лишайниковые грибы, возникшие около 1,3 млрд лет назад. [99] которые паслись на микробных матах. [28] Обильные эукариотические микрофоссилии из пресноводной шотландской группы Торридон, по-видимому, указывают на доминирование эукариот в неморских средах обитания к 1 млрд лет назад. [100] вероятно, из-за увеличения доступности питательных веществ в районах, расположенных ближе к континентам, и континентального стока. [32] Эти лишайники, возможно, позже каким-то образом способствовали колонизации растений 0,75 млрд лет назад. [99] Масштабное увеличение земной фотосинтетической биомассы, по-видимому, произошло около 0,85 млрд лет назад, о чем свидетельствует приток углерода земного происхождения, который мог привести к увеличению уровня кислорода, достаточному для поддержания экспансии многоклеточных эукариот. [101]
См. также
[ редактировать ]- Докембрий - История Земли 4600–539 миллионов лет назад.
- Эдиакарская биота - Жизнь эдиакарского периода
- Франвильская биота - возможные палеопротерозойские многоклеточные окаменелости из Габона.
- Бассейн Л'Аталанте - бассейн с бескислородной гиперсоленой водой на дне Средиземного моря.
- Земля-снежок - Эпизоды мирового оледенения во время протерозоя.
- естественные ядерные реакторы деления на территории нынешнего Окло , Габон. В то время действовали
Внешние ссылки
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж Мукерджи, И.; Большой, RR; Коркри, Р.; Данюшевский, Л.В. (2018). «Скучный миллиард: рогатка для сложной жизни на Земле» . Научные отчеты . 8 (4432): 4432. Бибкод : 2018НатСР...8.4432М . дои : 10.1038/s41598-018-22695-x . ПМЦ 5849639 . ПМИД 29535324 .
- ^ Бьюик, Р.; Де Марэ, диджей; Нолл, АХ (1995). «Стабильный изотопный состав карбонатов мезопротерозойской группы Бангмолл, северо-запад Австралии». Химическая геология . 123 (1–4): 153–171. Бибкод : 1995ЧГео.123..153Б . дои : 10.1016/0009-2541(95)00049-R . ПМИД 11540130 .
- ^ Брасье, М. (2012). Тайные камеры: внутренняя история клеток и сложной жизни . Издательство Оксфордского университета. п. 211. ИСБН 978-0-19-964400-1 .
- ^ Jump up to: а б с Янг, Грант М. (2013). «Докембрийские суперконтиненты, оледенения, насыщение атмосферы кислородом, эволюция многоклеточных животных и воздействие, которое могло изменить вторую половину истории Земли» . Геонаучные границы . 4 (3): 247–261. Бибкод : 2013GeoFr...4..247Y . дои : 10.1016/j.gsf.2012.07.003 .
- ^ Jump up to: а б с д и Кавуд, Питер А.; Хоксворт, Крис Дж. (1 июня 2014 г.). «Средний возраст Земли» . Геология . 42 (6): 503–506. Бибкод : 2014Geo....42..503C . дои : 10.1130/G35402.1 . hdl : 10023/4569 . ISSN 0091-7613 .
- ^ Jump up to: а б Холланд, Генрих Д. (29 июня 2006 г.). «Окисление атмосферы и океанов» . Философские труды Лондонского королевского общества B: Биологические науки . 361 (1470): 903–915. дои : 10.1098/rstb.2006.1838 . ISSN 0962-8436 . ПМЦ 1578726 . ПМИД 16754606 .
- ^ Jump up to: а б Пэн, Л.; Юнган, Л.; Ёнгюн, Х.; Юн, Ю.; Писаревский, С.А. (2019). «Теплый климат в эпоху «скучного миллиарда»» . Акта Геологика Синика . 93 (С3): 40–43. Бибкод : 2019AcGlS..93R..40L . дои : 10.1111/1755-6724.14239 .
- ^ Jump up to: а б с д Робертс, Нью-Мексико (2013). «Скучный миллиард? – Тектоника крышек, рост континентов и изменения окружающей среды, связанные с суперконтинентом Колумбия» . Геонаучные границы . Тематический раздел: Антарктида – окно в далекую землю. 4 (6): 681–691. Бибкод : 2013GeoFr...4..681R . дои : 10.1016/j.gsf.2013.05.004 .
- ^ Jump up to: а б с д и ж Лентон, Т.; Уотсон, А. (2011). «Не такой уж и скучный миллиард». Революции, сотворившие Землю . стр. 242–261. doi : 10.1093/acprof:oso/9780199587049.003.0013 . ISBN 978-0-19-958704-9 .
- ^ Планавский, Ной Дж.; Коул, Девон Б.; Иссон, Терри Т.; Рейнхард, Кристофер Т.; Крокфорд, Питер В.; Шелдон, Натан Д.; Лайонс, Тимоти В. (9 августа 2018 г.). «Дело о низком уровне кислорода в атмосфере в средней истории Земли» . Новые темы в науках о жизни . 2 (2): 149–159. дои : 10.1042/etls20170161 . ISSN 2397-8554 . ПМИД 32412619 . S2CID 210099422 .
- ^ Jump up to: а б с Брук, JL (2014). Изменение климата и ход глобальной истории: трудный путь . Издательство Кембриджского университета. стр. 40–42. ISBN 978-0-521-87164-8 .
- ^ Jump up to: а б Эванс, ПАПА (2013). «Реконструкция допангейских суперконтинентов». Бюллетень Геологического общества Америки . 125 (11–12): 1735–1751. Бибкод : 2013GSAB..125.1735E . дои : 10.1130/b30950.1 .
- ^ Брэдли, Дуайт К. (1 декабря 2008 г.). «Пассивные окраины в истории Земли» . Обзоры наук о Земле . 91 (1–4): 1–26. Бибкод : 2008ESRv...91....1B . doi : 10.1016/j.earscirev.2008.08.001 .
- ^ Горчик, В.; Смитис, Х.; Корхонен, Ф.; Ховард, Х.; де Громар, RQ (2015). «Сверхгорячая мезопротерозойская эволюция внутриконтинентальной центральной Австралии» . Геонаучные границы . 6 (1): 23–37. Бибкод : 2015GeoFr...6...23G . дои : 10.1016/j.gsf.2014.03.001 .
- ^ Брайан, SE; Феррари, Л. (2013). «Крупные магматические провинции и кислые крупные магматические провинции: прогресс в нашем понимании за последние 25 лет» . Бюллетень ГСА . 125 (7–8): 1055. Бибкод : 2013GSAB..125.1053B . дои : 10.1130/B30820.1 .
- ^ Толло, РП; Корриво, Л.; Маклелланд, Дж.; Варфоломей, MJ (2004). «Протерозойская тектоническая эволюция Гренвиллского орогена в Северной Америке: Введение» . В Толло, РП; Корриво, Л.; Маклелланд, Дж.; Варфоломей, MJ (ред.). Протерозойская тектоническая эволюция Гренвиллского орогена в Северной Америке . Мемуары Геологического общества Америки. Том. 197. стр. 1–18. ISBN 978-0-8137-1197-3 .
- ^ Брейзер, доктор медицины (1998). «Миллиард лет стабильности окружающей среды и появление эукариот: новые данные из северной Австралии». Геология . 26 (6): 555–558. Бибкод : 1998Geo....26..555B . doi : 10.1130/0091-7613(1998)026<0555:ABYOES>2.3.CO;2 . ПМИД 11541449 .
- ^ Jump up to: а б с Фиорелла, Р.; Шелдон, Н. (2017). «Равный климат конца мезопротерозоя при отсутствии высокого содержания CO2» . Геология . 45 (3): 231–234. Бибкод : 2017Geo....45..231F . дои : 10.1130/G38682.1 .
- ^ Jump up to: а б с д Вейзер, Дж. (2005). «Драйвер небесного климата: взгляд на четыре миллиарда лет углеродного цикла» . Геонаука Канады . 32 (1). ISSN 1911-4850 .
- ^ Jump up to: а б Ках, ЛК; Верховая езда, Р. (2007). «Уровни углекислого газа в мезопротерозое, определенные по кальцинированным цианобактериям». Геология . 35 (9): 799–802. Бибкод : 2007Geo....35..799K . дои : 10.1130/G23680A.1 . S2CID 129389459 .
- ^ Хартли, А.; Куржански, Б.; Пагсли, Дж.; Армстронг, Дж. (2020). «Ледяной сплав по озерам в раннем неопротерозое: камни в формации Диабайг, группа Торридон, северо-запад Шотландии». Шотландский геологический журнал . 56 (1): 47–53. Бибкод : 2020ScJG...56...47H . дои : 10.1144/sjg2019-017 . hdl : 2164/15493 . S2CID 214436923 .
- ^ Хрен, Монтана; Шелдон, Северная Дакота (2020). «Земные микробиалиты ограничивают мезопротерозойскую атмосферу» . Депозитарный протокол . 6 (1): 4–20. Бибкод : 2020DepRe...6....4H . дои : 10.1002/деп2.79 . hdl : 2027.42/154283 .
- ^ Шавив, Нир Дж. (2003). «Спиральная структура Млечного Пути, космические лучи и эпохи ледникового периода на Земле». Новая астрономия . 8 (1): 39–77. arXiv : astro-ph/0209252 . Бибкод : 2003NewA....8...39S . дои : 10.1016/S1384-1076(02)00193-8 . S2CID 879406 .
- ^ Jump up to: а б Планавский, Ной Дж.; Рейнхард, Кристофер Т.; Ван, Сянли; Томсон, Даниэль; Макголдрик, Питер; Рейнберд, Роберт Х.; Джонсон, Томас; Фишер, Вудворд В.; Лайонс, Тимоти В. (31 октября 2014 г.). «Низкий уровень кислорода в атмосфере в середине протерозоя и задержка подъема животных» (PDF) . Наука . 346 (6209): 635–638. Бибкод : 2014Sci...346..635P . дои : 10.1126/science.1258410 . ISSN 0036-8075 . ПМИД 25359975 . S2CID 37395258 .
- ^ Эйлс, Н. (2008). «Гляциоэпохи и цикл суперконтинента после ~ 3,0 млрд лет назад: тектонические граничные условия для оледенения». Палеогеография, Палеоклиматология, Палеоэкология . 258 (1–2): 89–129. Бибкод : 2008PPP...258...89E . дои : 10.1016/j.palaeo.2007.09.021 .
- ^ Кастинг, Дж. Ф.; Оно, С. (2006). «Палеоклиматы: первые два миллиарда лет» . Философские труды Лондонского королевского общества B: Биологические науки . 361 (1470): 917–929. дои : 10.1098/rstb.2006.1839 . ISSN 0962-8436 . ПМЦ 1868609 . ПМИД 16754607 .
- ^ Кэтлинг, округ Колумбия; Кастинг, Дж. Ф. (2017). Эволюция атмосферы на обитаемых и безжизненных мирах . Издательство Кембриджского университета. п. 291. ИСБН 978-1-316-82452-8 .
- ^ Jump up to: а б с д Беральди-Кампези, Х. (2013). «Ранняя жизнь на суше и первые наземные экосистемы» . Экологические процессы . 2 (1): 1–17. Бибкод : 2013ЭкоПр...2....1Б . дои : 10.1186/2192-1709-2-1 .
- ^ Анбар, AD; Нолл, АХ (2002). «Химия и эволюция протерозойского океана: биоинорганический мост?». Наука . 297 (5584): 1137–1142. Бибкод : 2002Sci...297.1137A . CiteSeerX 10.1.1.615.3041 . дои : 10.1126/science.1069651 . ISSN 0036-8075 . ПМИД 12183619 . S2CID 5578019 .
- ^ Фенхель, К .; Следует, М; Фальковски, П.Г. (2005). «Коэволюция циклов азота, углерода и кислорода в протерозойском океане» . Американский научный журнал . 305 (6–8): 526–545. Бибкод : 2005AmJS..305..526F . дои : 10.2475/ajs.305.6-8.526 . ISSN 0002-9599 .
- ^ Кипп, Массачусетс; Стюкен, Э.Э. (2017). «Переработка биомассы и ранний цикл фосфора Земли» . Достижения науки . 3 (11): eaao4795. Бибкод : 2017SciA....3O4795K . дои : 10.1126/sciadv.aao4795 . ПМЦ 5706743 . ПМИД 29202032 .
- ^ Jump up to: а б Парнелл, Дж.; Спринсс, С.; Эндрюс, С.; Тайалан, В.; Боуден, С. (2015). «Высокая доступность молибдена для эволюции в мезопротерозойской озерной среде» . Природные коммуникации . 6 (6996): 6996. Бибкод : 2015NatCo...6.6996P . дои : 10.1038/ncomms7996 . hdl : 2164/5221 . ПМИД 25988499 .
- ^ Jump up to: а б с д и ж г Кэнфилд, Делавэр (1998). «Новая модель химии протерозойского океана». Природа . 396 (6710): 450–453. Бибкод : 1998Natur.396..450C . дои : 10.1038/24839 . ISSN 0028-0836 . S2CID 4414140 .
- ^ Jump up to: а б с д и ж Лайонс, Тимоти В.; Рейнхард, Кристофер Т.; Планавский, Ной Дж. (2014). «Повышение содержания кислорода в раннем океане и атмосфере Земли». Природа . 506 (7488): 307–315. Бибкод : 2014Natur.506..307L . дои : 10.1038/nature13068 . ПМИД 24553238 . S2CID 4443958 .
- ^ Jump up to: а б с д Большой, Р.; Халпин, Дж.А.; Данюшевский, Л.В. (2014). «Содержание микроэлементов в осадочном пирите как новый показатель глубокой эволюции океана и атмосферы». Письма о Земле и планетологии . 389 : 209–220. Бибкод : 2014E&PSL.389..209L . дои : 10.1016/j.epsl.2013.12.020 .
- ^ Jump up to: а б с Слэк, Дж. Ф.; Кэннон, ВФ (2009). «Внеземная гибель полосчатых железных образований 1,85 миллиарда лет назад». Геология . 37 (11): 1011–1014. Бибкод : 2009Geo....37.1011S . дои : 10.1130/G30259A.1 .
- ^ Jump up to: а б Де Барр, HJW; немецкий, ЧР; Элдерфилд, Х; ван Ганс, П. (1988). «Распределение редкоземельных элементов в бескислородных водах желоба Кариако». Акта геохимии и космохимии . 52 (5): 1203–1219. Бибкод : 1988GeCoA..52.1203D . дои : 10.1016/0016-7037(88)90275-X .
- ^ Галлардо, Вирджиния; Эспиноза, К. (2008). Гувер, Ричард Б; Левин, Гилберт V; Розанов Алексей Юрьевич; Дэвис, Пол С. (ред.). «Эволюция цвета океана» (PDF) . Труды Международного общества оптической инженерии . Инструменты, методы и задачи астробиологии XI. 7097 : 1–7. Бибкод : 2008SPIE.7097E..0GG . дои : 10.1117/12.794742 . S2CID 53984439 . Архивировано из оригинала (PDF) 9 августа 2017 г. Проверено 3 декабря 2019 г.
- ^ Jump up to: а б Цю, Джейн (2014). «Колебания кислорода остановили жизнь на Земле» . Природа . дои : 10.1038/nature.2014.15529 . S2CID 131404410 . Проверено 24 февраля 2017 г.
- ^ Jump up to: а б Дойл, Кэтрин А.; Поултон, Саймон В.; Ньютон, Роберт Дж.; Подковыров Виктор Н.; Беккер, Андрей (октябрь 2018 г.). «Аноксия на мелководье в мезопротерозойском океане: данные Башкирского мегантиклинория, Южный Урал» . Докембрийские исследования . 317 : 196–210. Бибкод : 2018PreR..317..196D . doi : 10.1016/j.precamres.2018.09.001 . S2CID 134378820 . Проверено 17 декабря 2022 г.
- ^ Jump up to: а б с Джонстон, DT (2009). «Аноксигенный фотосинтез модулирует протерозойский кислород и поддерживает средний возраст Земли» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (40): 16925–16929. Бибкод : 2009PNAS..10616925J . дои : 10.1073/pnas.0909248106 . ПМЦ 2753640 . ПМИД 19805080 .
- ^ Партин, Калифорния; Беккер, А.; Планавски, Нью-Джерси; Скотт, Коннектикут; Гилл, Британская Колумбия; Ли, К.; Подковыров В.; Маслов А.; Конхаузер, КО (1 мая 2013 г.). «Крупномасштабные колебания уровня кислорода в атмосфере и океане докембрия на основе записей U в сланцах». Письма о Земле и планетологии . 369–370: 284–293. Бибкод : 2013E&PSL.369..284P . дои : 10.1016/j.epsl.2013.03.031 .
- ^ Беккер, А.; Голландия, HD (1 февраля 2012 г.). «Переизбыток кислорода и восстановление в раннем палеопротерозое». Письма о Земле и планетологии . 317–318: 295–304. Бибкод : 2012E&PSL.317..295B . дои : 10.1016/j.epsl.2011.12.012 .
- ^ Шредер, С.; Беккер, А.; Бьюкс, Нью-Джерси; Штраус, Х.; Ван Никерк, HS (1 апреля 2008 г.). «Рост концентрации сульфатов в морской воде, связанный с палеопротерозойским положительным выбросом изотопов углерода: данные по сульфатным эвапоритам в мелководной морской формации Лакхнау ~ 2,2–2,1 млрд лет, Южная Африка» . Терра Нова . 20 (2): 108–117. Бибкод : 2008TeNov..20..108S . дои : 10.1111/j.1365-3121.2008.00795.x . ISSN 1365-3121 .
- ^ Хардисти, Далтон С.; Лу, Зунли; Беккер, Андрей; Даймонд, Чарльз В.; Гилл, Бенджамин К.; Цзян, Ганьцин; Ка, Линда С.; Нолл, Эндрю Х.; Ллойд, Шон Дж.; Осберн, Магдалена Р.; Планавский, Ной Дж.; Ван, Чуньцзян; Чжоу, Сяоли; Лайонс, Тимоти В. (1 апреля 2017 г.). «Перспективы окислительно-восстановительного потенциала протерозойской поверхности океана по содержанию йода в древних и современных карбонатах» . Письма о Земле и планетологии . 463 : 159–170. Бибкод : 2017E&PSL.463..159H . дои : 10.1016/j.epsl.2017.01.032 . hdl : 1912/8872 . S2CID 13136821 .
- ^ Чжан, Шуйчан; Ван, Хуацзянь; Ван, Сяомэй; Йе, Юньтао (25 октября 2021 г.). «Мезопротерозойское событие оксигенации» . Наука Китай Науки о Земле . 64 (12): 2043–2068. Бибкод : 2021ScChD..64.2043Z . дои : 10.1007/s11430-020-9825-x . S2CID 239891560 . Проверено 9 мая 2023 г.
- ^ Йе, Юнтао; Ван, Хуацзянь; Ван, Сяомэй; Ли, Цзе; У, Чаодун; Чжан, Шуйчан (1 мая 2023 г.). «Региональные и глобальные показатели изменения окислительно-восстановительных условий океана на протяжении ~ 1,57 млрд лет назад: причинно-следственная связь с выветриванием, вызванным вулканизмом» . Геосистемы и геосреда . 2 (2): 100173. Бибкод : 2023GsGe....200173Y . дои : 10.1016/j.geogeo.2022.100173 . ISSN 2772-8838 .
- ^ Jump up to: а б Мукерджи, Индрани; Лардж, Росс Р. (август 2016 г.). «Химия микроэлементов пирита в формации Велкерри, группа Ропер, бассейн Макартур: свидетельства насыщения атмосферы кислородом во время Бурного миллиарда» . Докембрийские исследования . 281 : 13–26. Бибкод : 2016PreR..281...13M . doi : 10.1016/j.precamres.2016.05.003 . Проверено 8 ноября 2022 г.
- ^ Он, Ютинг; Чжу, Сиян; Цю, Ифань; Панг, Ланьинь; Чжао, Тайпин (декабрь 2022 г.). «Экстремальные изменения климата повлияли на эволюцию ранней жизни примерно в 1,4 млрд лет назад: последствия сланцев формации Сямалин на севере Северо-Китайского кратона» . Докембрийские исследования . 383 : 106901. Бибкод : 2022PreR..38306901H . doi : 10.1016/j.precamres.2022.106901 . S2CID 253787177 . Проверено 17 декабря 2022 г.
- ^ Лю, Аньци; Тан, Дунцзе; Ши, Сяоин; Чжоу, Сицян; Чжоу, Лимин; Шан, Мохан; Ли, Ян; Фанг, Хао (1 июня 2020 г.). «Мезопротерозойская насыщенная кислородом глубоководная морская вода, зафиксированная раннедиагенетическими карбонатными конкрециями из пачки IV формации Сямалин, Северный Китай» . Докембрийские исследования . 341 : 105667. Бибкод : 2020PreR..34105667L . doi : 10.1016/j.precamres.2020.105667 . ISSN 0301-9268 . S2CID 212948463 . Проверено 11 сентября 2023 г.
- ^ Ках, ЛК; Лайонс, ТВ; Фрэнк, Т.Д. (2004). «Низкая морская сульфатность и длительная оксигенация протерозойской биосферы» . Природа . 438 (7010): 834–838. Бибкод : 2004Natur.431..834K . дои : 10.1038/nature02974 . ПМИД 15483609 . S2CID 4404486 .
- ^ Оч, ЛМ; Шилдс-Чжоу, Джорджия (2012). «Неопротерозойское событие оксигенации: экологические возмущения и биогеохимический цикл». Обзоры наук о Земле . 110 (1–4): 26–57. Бибкод : 2012ESRv..110...26O . doi : 10.1016/j.earscirev.2011.09.004 .
- ^ Лайонс, Тимоти В.; Рейнхард, Кристофер Т. (2009). «Ранний продуктивный океан, непригодный для аэробики» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (43): 18045–18046. Бибкод : 2009PNAS..10618045L . дои : 10.1073/pnas.0910345106 . ISSN 0027-8424 . ПМЦ 2775325 . ПМИД 19846788 .
- ^ Бенигк, Дж.; Водник, С.; Глюксман, Э. (2015). Биоразнообразие и история Земли . Спрингер. стр. 58–59. ISBN 978-3-662-46394-9 .
- ^ Чжэн, Ван; Жиллодо, Джеффри Дж.; Ка, Линда С.; Анбар, Ариэль Д. (16 октября 2018 г.). «Сигнатуры изотопов ртути фиксируют эвксинию фотической зоны в мезопротерозойском океане» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (42): 10594–10599. Бибкод : 2018PNAS..11510594Z . дои : 10.1073/pnas.1721733115 . ISSN 0027-8424 . ПМК 6196510 . ПМИД 30275325 .
- ^ Планавский, Ной Дж.; Макголдрик, Питер; Скотт, Клинтон Т.; Ли, Чао; Рейнхард, Кристофер Т.; Келли, Эми Э.; Чу, Сюэлэй; Беккер, Андрей; С любовью, Гордон Д.; Лайонс, Тимоти В. (сентябрь 2011 г.). «Распространенные богатые железом условия в среднепротерозойском океане» . Природа . 477 (7365): 448–451. Бибкод : 2011Natur.477..448P . дои : 10.1038/nature10327 . ISSN 1476-4687 . ПМИД 21900895 . S2CID 205225926 .
- ^ Жиллодо, Джеффри Дж.; Романьелло, Стивен Дж.; Ло, Генмин; Кауфман, Алан Дж.; Чжан, Фейфей; Клаэбе, Роберт М.; Ка, Линда С.; Азми, Карем; Бартли, Джули К.; Чжэн, Ван; Нолл, Эндрю Х.; Анбар, Ариэль Д. (01 сентября 2019 г.). «Изотопные доказательства наличия ограниченной эвксинии в океанах среднего протерозоя» . Письма о Земле и планетологии . 521 : 150–157. Бибкод : 2019E&PSL.521..150G . дои : 10.1016/j.epsl.2019.06.012 . ISSN 0012-821X . S2CID 198401867 .
- ^ Ву, Яовэй; Тянь, Хуэй; Инь, Жуньшэн; Чен, Ди; Грасби, Стивен Э.; Шен, Цзюнь; Ли, Тэнфэй; Джи, Суй; Пэн, Пинъань (15 августа 2023 г.). «Высокофракционированные изотопы Hg свидетельствуют о динамической эвксинии на мелководье мезопротерозойского океана» . Письма о Земле и планетологии . 616 : 118211. Бибкод : 2023E&PSL.61618211W . дои : 10.1016/j.epsl.2023.118211 . S2CID 259004695 . Проверено 3 июля 2023 г.
- ^ Спинкс, Сэмюэл К.; Шмид, Сюзанна; Пажес, Анаис (декабрь 2016 г.). «Отсроченная эвксиния во внутриконтинентальных морях палеопротерозоя: жизненно важные пристанища для эволюции эукариот?» . Докембрийские исследования . 287 : 108–114. Бибкод : 2016PreR..287..108S . doi : 10.1016/j.precamres.2016.11.002 . Проверено 19 мая 2024 г. - через Elsevier Science Direct.
- ^ Jump up to: а б Кэнфилд, Делавэр ; Чжан, С.; Ван, Х.; Ван, X.; Чжао, В.; Су, Дж.; Бьеррум, CJ; Хаксен, скорая помощь; Хаммарлунд, ЕС (2018). «Мезопротерозойское железное образование» . Труды Национальной академии наук . 115 (17): 3895–3904. Бибкод : 2018PNAS..115E3895C . дои : 10.1073/pnas.1720529115 . ПМЦ 5924912 . ПМИД 29632173 .
- ^ Брейзер, доктор медицины; Линдси, Дж. Ф. (1998). «Миллиард лет стабильности окружающей среды и появление эукариот: новые данные из северной Австралии». Геология . 26 (6): 555–558. Бибкод : 1998Geo....26..555B . doi : 10.1130/0091-7613(1998)026<0555:ABYOES>2.3.CO;2 . ПМИД 11541449 .
- ^ Jump up to: а б Генели, Н.; Маккенна, AM; Окоучи, Н.; Борэм, CJ; Бегин, Дж.; Яво, Э.Дж.; Брокс, Джей Джей (2018). «Порфирины возрастом 1,1 миллиарда лет создают морскую экосистему, в которой доминируют бактериальные первичные продуценты» . Труды Национальной академии наук . 115 (30): 6978–6986. Бибкод : 2018PNAS..115E6978G . дои : 10.1073/pnas.1803866115 . ПМК 6064987 . ПМИД 29987033 .
- ^ Jump up to: а б Яво, Э.Дж.; Лепот, К. (2018). «Палеопротерозойская летопись окаменелостей: последствия для эволюции биосферы в средний возраст Земли» . Обзоры наук о Земле . 176 : 68–86. Бибкод : 2018ESRv..176...68J . doi : 10.1016/j.earscirev.2017.10.001 . hdl : 20.500.12210/62416 .
- ^ Санчес-Баракальдо, П. (2015). «Происхождение морских планктонных цианобактерий» . Научные отчеты . 5 : 17418. Бибкод : 2015NatSR...517418S . дои : 10.1038/srep17418 . ПМК 4665016 . ПМИД 26621203 .
- ^ Ширмейстер, Беттина Э.; Санчес-Баракальдо, Патрисия; Уэйси, Дэвид (2016). «Эволюция цианобактерий в докембрии» . Международный журнал астробиологии . 15 (3): 187–204. Бибкод : 2016IJAsB..15..187S . дои : 10.1017/S1473550415000579 .
- ^ Ранняя жизнь на Земле, ограниченная ферментом | Новости UCL
- ^ Аллен, Джон Ф.; Тэйк, Бренда; Мартин, Уильям Ф. (2019). «Ингибирование нитрогеназы, ограниченное окисление протерозойской атмосферы Земли» . Тенденции в науке о растениях . 24 (11): 1022–1031. Бибкод : 2019TPS....24.1022A . doi : 10.1016/j.tplants.2019.07.007 . ПМИД 31447302 .
- ^ Фахри, Моджтаба; Тархан Лидия Георгиевна; Рейнхард, Кристофер Т.; Кроу, Шон А.; Лайонс, Тимоти В.; Планавски, Ной Дж. (май 2023 г.). «Окисление поверхности Земли и возникновение эукариотической жизни: новый взгляд на связь с экскурсией положительных изотопов углерода в Ломагунди» . Обзоры наук о Земле . 240 : 104398. Бибкод : 2023ESRv..24004398F . doi : 10.1016/j.earscirev.2023.104398 . S2CID 257761993 .
- ^ Рейнхард, Коннектикут; Планавски, Нью-Джерси; Роббинс, LJ; Партин, Калифорния; Гилл, Британская Колумбия; Лалонд, СВ; Беккер, А.; Конхаузер, КО; Лайонс, ТВ (2013). «Окислительно-восстановительный потенциал протерозойского океана и биогеохимический застой» . Труды Национальной академии наук . 110 (14): 5357–5362. Бибкод : 2013PNAS..110.5357R . дои : 10.1073/pnas.1208622110 . ISSN 0027-8424 . ПМК 3619314 . ПМИД 23515332 .
- ^ Анбар, AD (2002). «Химия и эволюция протерозойского океана: бионеорганический мост». Наука . 297 (5584): 1137–1142. Бибкод : 2002Sci...297.1137A . дои : 10.1126/science.1069651 . ПМИД 12183619 . S2CID 5578019 .
- ^ Чжан, Фэнлянь; Йе, Юньтао; Лю, Итун; Ван, Сяомэй; У, Хуайчунь; Чжан, Шуйчан» . эукариот в г. высокая не низкий температура ли эволюции . изд O2 Препятствовала " апреля а докембрии , 29 2023 уровень ?
- ^ Jump up to: а б Бенгтсон, С.; Сальстедт, Т.; Беливанова В.; Уайтхаус, М. (2017). «Трехмерная сохранность клеточных и субклеточных структур позволяет предположить, что красным водорослям коронной группы возрастом 1,6 миллиарда лет» . ПЛОС Биология . 15 (3): e2000735. doi : 10.1371/journal.pbio.2000735 . ПМК 5349422 . ПМИД 28291791 .
- ^ Jump up to: а б с Хеджес, Южная Каролина; Блэр, Дж. Э.; Вентури, ML; Шу, Дж.Л. (2004). «Молекулярная временная шкала эволюции эукариот и возникновения сложной многоклеточной жизни» . Эволюционная биология BMC . 4 (2): 2. дои : 10.1186/1471-2148-4-2 . ПМК 341452 . ПМИД 15005799 .
- ^ Бенгтсон, С.; Беливанова В.; Расмуссен, Б.; Уайтхаус, М. (2009). «Спорные «кембрийские» окаменелости виндианцев реальны, но старше их более чем на миллиард лет» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (19): 7729–7734. Бибкод : 2009PNAS..106.7729B . дои : 10.1073/pnas.0812460106 . ПМЦ 2683128 . ПМИД 19416859 .
- ^ Лейминг, Инь; Биан, Лизенг; Сюньлай, Юань (октябрь 2004 г.). «Открытие разветвленных трубчатых водорослей и микроскопических трубок с кольцеобразно-спиральными утолщениями из мезопротерозойской группы Руян провинции Шаньси, Северный Китай» . Наука Китай Науки о Земле . 47 (10): 880–885. дои : 10.1360/02yd0356 . S2CID 129336775 . Проверено 13 октября 2022 г.
- ^ Панг, Ке; Тан, Цин; Юань, Сюнь-Лай; Ван, Бин; Сяо, Шухай (сентябрь 2015 г.). «Биомеханический анализ ранней эукариотической ископаемой Валерии и нового появления микрокаменелостей с органическими стенками из палео-мезопротерозойской группы Руян» . Палеомир . 24 (3): 251–262. дои : 10.1016/j.palwor.2015.04.002 .
- ^ Лейминг, Инь; Сюньлай, Юань; Фанвэй, Мэн; Цзе, Ху (7 ноября 2005 г.). «Протисты верхнемезопротерозойской группы Руян в провинции Шаньси, Китай» . Докембрийские исследования . 141 (1–2): 49–66. Бибкод : 2005PreR..141...49L . doi : 10.1016/j.precamres.2005.08.001 . Проверено 13 октября 2022 г.
- ^ Jump up to: а б Воробьева Наталья Георгиевна; Сергеев Владимир Н.; Петров, Петр Ю. (январь 2015 г.). «Комплекс котуйканской свиты: разнообразная микробиота с органическими стенками в мезопротерозойской анабарской последовательности, север Сибири» . Докембрийские исследования . 256 : 201–222. Бибкод : 2015PreR..256..201В . дои : 10.1016/j.precamres.2014.11.011 . Проверено 15 октября 2022 г.
- ^ Агич, Хеда; Мочидловска, Малгожата; Инь, Лейминг (август 2017 г.). «Разнообразие микрокаменелостей с органическими стенками из ранней мезопротерозойской группы Руян, Северо-Китайский кратон - окно в раннюю эволюцию эукариот» . Докембрийские исследования . 297 : 101–130. Бибкод : 2017PreR..297..101A . doi : 10.1016/j.precamres.2017.04.042 . Проверено 13 октября 2022 г.
- ^ Лорон, CC; Франсуа, К.; Рейнберд, Р.Х.; Тернер, ЕС; Боренштейн, С.; Яво, EJ (2019). «Ранние грибы протерозоя в Арктической Канаде» . Природа . 70 (7760): 232–235. Бибкод : 2019Natur.570..232L . дои : 10.1038/s41586-019-1217-0 . ПМИД 31118507 . S2CID 162180486 .
- ^ Купер, GM (2000). «Происхождение и эволюция клеток» . Клетка: молекулярный подход (2-е изд.). Синауэр Ассошиэйтс.
- ^ Никлас, KJ (2014). «Эволюционно-развитие истоков многоклеточности». Американский журнал ботаники . 101 (1): 6–25. дои : 10.3732/ajb.1300314 . ПМИД 24363320 .
- ^ Бернштейн, Х.; Бернштейн, К.; Мишо, RE (2012). «Репарация ДНК как основная адаптивная функция пола у бактерий и эукариот» . В Кимуре, С.; Симидзу, С. (ред.). Восстановление ДНК: новые исследования . Нова Биомедикал. стр. 1–49. ISBN 978-1-62100-756-2 . Архивировано из оригинала 29 октября 2013 года.
- ^ Гибсон, Тимоти М.; Ши, Патрик М.; Камминг, Вивьен М.; Фишер, Вудворд В.; Крокфорд, Питер В.; Ходжскисс, Малкольм С.В.; Верндле, Сара; Кризер, Роберт А.; Рейнберд, Роберт Х.; Скульски, Томас М.; Халверсон, Гален П. (08 декабря 2017 г.). «Точный возраст Bangiomorpha pubescens датирует возникновение эукариотического фотосинтеза» . Геология . 46 (2): 135–138. дои : 10.1130/g39829.1 . ISSN 0091-7613 .
- ^ Эгель, Р.; Пенни, Д. (2007). «О происхождении мейоза в эволюции эукариот: коэволюция мейоза и митоза с самого начала» . Динамика и стабильность генома . 3 (249–288): 249–288. дои : 10.1007/7050_2007_036 . ISBN 978-3-540-68983-6 .
- ^ Jump up to: а б Мартин, В.; Мюллер, М. (1998). «Водородная гипотеза первых эукариот». Природа . 392 (6671): 37–41. Бибкод : 1998Natur.392...37M . дои : 10.1038/32096 . ISSN 0028-0836 . ПМИД 9510246 . S2CID 338885 .
- ^ Тиммис, Дж. Н.; Эйлифф, Майкл А.; Хуанг, Китай; Мартин, В. (2004). «Эндосимбиотический перенос генов: геномы органелл создают эукариотические хромосомы». Обзоры природы Генетика . 5 (2): 123–135. дои : 10.1038/nrg1271 . ПМИД 14735123 . S2CID 2385111 .
- ^ Ментель, Марек; Мартин, Уильям (27 августа 2008 г.). «Энергетический обмен среди эукариотических анаэробов в свете химии океана протерозоя» . Философские труды Лондонского королевского общества B: Биологические науки . 363 (1504): 2717–2729. дои : 10.1098/rstb.2008.0031 . ISSN 0962-8436 . ПМК 2606767 . ПМИД 18468979 .
- ^ Мочидловска, М.; Лендинг, Э.; Занг, В.; Паласиос, Т. (2011). «Протерозойский фитопланктон и время возникновения хлорофитных водорослей» . Палеонтология . 54 (4): 721–733. Бибкод : 2011Palgy..54..721M . дои : 10.1111/j.1475-4983.2011.01054.x .
- ^ Стэнли, С.М. (2008). «Хищничество побеждает конкуренцию на морском дне». Палеобиология . 31 (1): 12. Бибкод : 2008Pbio...34....1S . дои : 10.1666/07026.1 . S2CID 83713101 .
- ^ Санчес-Баракальдо, Патрисия; Риджвелл, Энди; Рэйвен, Джон А. (17 марта 2014 г.). «Неопротерозойский переход в морском азотном цикле» . Современная биология . 24 (6): 652–657. Бибкод : 2014CBio...24..652S . дои : 10.1016/j.cub.2014.01.041 . ПМИД 24583016 . S2CID 16756351 .
- ^ Хоманн, М.; и др. (2018). «Микробная жизнь и биогеохимический круговорот на суше 3220 миллионов лет назад» (PDF) . Природа Геонауки . 11 (9): 665–671. Бибкод : 2018NatGe..11..665H . дои : 10.1038/s41561-018-0190-9 . S2CID 134935568 .
- ^ Баумгартнер, Р.Дж.; ван Кранендонк, MJ; и др. (2019). «Нанопористый пирит и органическое вещество в строматолитах возрастом 3,5 миллиарда лет свидетельствуют о первозданной жизни» (PDF) . Геология . 47 (11): 1039–1043. Бибкод : 2019Geo....47.1039B . дои : 10.1130/G46365.1 . S2CID 204258554 .
- ^ Ватанабэ, Юмико; Мартини, Жак Э.Ж.; Омото, Хироши (30 ноября 2000 г.). «Геохимические доказательства существования наземных экосистем 2,6 миллиарда лет назад». Природа . 408 (6812): 574–578. Бибкод : 2000Natur.408..574W . дои : 10.1038/35046052 . ISSN 0028-0836 . ПМИД 11117742 . S2CID 4427507 .
- ^ Jump up to: а б Городиский, Р.Дж.; Кнаут, LP (1994). «Жизнь на суше в докембрии». Наука . 263 (5146): 494–498. Бибкод : 1994Sci...263..494H . дои : 10.1126/science.263.5146.494 . ISSN 0036-8075 . ПМИД 17754880 . S2CID 37687767 .
- ^ Реталлак, Грегори Дж.; Миндсенти, Андреа (1 апреля 1994 г.). «Хорошо сохранившиеся палеопочвы позднего докембрия из северо-западной Шотландии» . Журнал осадочных исследований . 64 (2а): 264–281. doi : 10.1306/D4267D7A-2B26-11D7-8648000102C1865D . ISSN 1527-1404 .
- ^ Городиский, Р.Дж.; Кнаут, LP (1994). «Жизнь на суше в докембрии». Наука . 263 (5146): 494–498. Бибкод : 1994Sci...263..494H . дои : 10.1126/science.263.5146.494 . ISSN 0036-8075 . ПМИД 17754880 . S2CID 37687767 .
- ^ Праве, Энтони Роберт (2002). «Жизнь на суше в протерозое: свидетельства торридонских пород северо-западной Шотландии» . Геология . 30 (9): 811. Бибкод : 2002Geo....30..811P . doi : 10.1130/0091-7613(2002)030<0811:LOLITP>2.0.CO;2 . Архивировано из оригинала 7 марта 2016 года . Проверено 5 марта 2016 г.
- ^ Jump up to: а б Хекман, Д.С.; Гейзер, Д.М.; Эйделл, БР; Стауффер, РЛ; Кардос, Нидерланды; Хеджес, С.Б. (2001). «Молекулярные доказательства ранней колонизации суши грибами и растениями». Наука . 293 (5532): 494–498. Бибкод : 1994Sci...263..494H . дои : 10.1126/science.263.5146.494 . ПМИД 17754880 . S2CID 37687767 .
- ^ Стротер, ПК; Баттисон, Л.; Брейзер, Мэриленд ; Веллман, Швейцария (2011). «Самые ранние неморские эукариоты Земли» . Природа . 473 (7348): 505–509. Бибкод : 2011Natur.473..505S . дои : 10.1038/nature09943 . ПМИД 21490597 . S2CID 4418860 .
- ^ Кнаут, LP (2009). «Позднедокембрийское позеленение Земли». Природа . 460 (7256): 728–732. Бибкод : 2009Natur.460..728K . дои : 10.1038/nature08213 . ПМИД 19587681 . S2CID 4398942 .